Introduction
Acute myocardial infarction is a prevalent public
health problem which results in severe morbidity and mortality
globally (1,2). At present, the standard treatment
for acute myocardial infarction is early reperfusion therapy,
including thrombolysis, percutaneous coronary intervention and
coronary bypass grafting (3).
However, the process of reperfusion may cause additional
cardiomyocyte dysfunction and death, and this process is generally
referred to as myocardial reperfusion injury (MI/R) (4). MI/R injury is a major limitation of
certain clinical therapies for coronary artery disease.
The underlying mechanisms of MI/R injury are
complex, and at present multiple factors, including excessive
generation of reactive oxygen species (ROS), calcium overload and
vascular endothelial dysfunction are viewed as being relevant
(5). Among these contributors,
oxidative stress induced by the burst of ROS that occurs during
reperfusion is the critical trigger of MI/R injury (6). An appropriate amount of ROS usually
has a cardioprotective effect, while high levels of ROS are
deleterious and lead to cardiomyocyte death (7,8).
Excessive ROS generated during the MI/R process not only directly
and non-specifically oxidize biological macromolecules including
DNA, lipids, and proteins, but also damage cells by activating
redox-regulated signaling cascades that ultimately lead to cell
death (9). The generation of ROS
and the clearance of ROS are important factors that affect ROS
outbreaks (10). Compared with
the ability of ROS clearance, inhibition of ROS generation may be
more important in terms of avoiding ROS outbreaks (10). Previous studies have identified
major sources of ROS production in heart disorders: The
mitochondrial electronic transport chain (ETC) of mitochondria,
NADPH oxidases (NOX) and xanthine oxidase (XO) in the cytoplasm
(11). Therefore, inhibition of
mitochondrial and cytoplasmic ROS formation during the MI/R
process, which avoids the outbreak of ROS from their site of
origin, may be a key therapeutic approach to effectively protect
the myocardium during the MI/R process.
Pigment epithelium-derived factor (PEDF), a 50-kDa
secreted glycoprotein, belongs to the superfamily of serine
protease inhibitors (12). PEDF
is expressed in multiple tissues but is expressed prominently in
heart tissue, where it exerts diverse physiological activities
(13-15). Previous studies by our group have
demonstrated that PEDF protects cardiomyocytes against
hypoxia-induced apoptosis and necroptosis via anti-oxidative
effects, and PEDF improves cardiac function in rats with acute
myocardial infarction via inhibition of vascular permeability and
cardiomyocyte apoptosis (14,16). Multiple studies have focused on
the effects of PEDF in hypoxic cardiomyocytes. However, the
function of PEDF across the whole process of MI/R has rarely been
investigated.
PEDF is mainly deposited in the cell membrane, where
it interacts with its receptors (17). The PEDF receptor (PEDF-R) and
laminin receptor are two crucial receptors among PEDF binding sites
(17,18). PEDF-R, an enzyme protein of
roughly 55 KDa in size and 504 amino acids in length, exerts potent
phospholipase A2 enzymatic activity and lipase activity when it is
combined with PEDF (19,20). Previous studies by our group have
demonstrated that the effect of PEDF, that may protect
cardiomyocytes under hypoxic conditions, is mediated by PEDF-R
(21,22). As a result, it is possible to
conclude that PEDF-R mediates the protective effect of PEDF in MI/R
Injury.
In the present study, the adult Sprague-Dawley rat
MI/R model and the H9c2 cardiomyocyte hypoxia/reoxygeneration (H/R)
model were established to mimic MI/R injury. The hypothesis that
the protective effect of PEDF in MI/R injury and the antioxidative
effects of PEDF are based on the inhibition of mitochondrial and
cytosolic ROS generation via PEDF-R was investigated. The present
study may provide a novel therapeutic target for MI/R injury.
Materials and methods
Reagents
Cleaved caspase-3 antibodies (cat. no. 9664) was
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Monoclonal actin (α-sarcomeric; α-sa) antibodies (cat. no. A2172),
Dobutamine, Evans Blue dye and 2,3,5-triphenyl-tetrazolium (TTC)
were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
The in situ cell death detection kit (cat. no. 11684795910)
was purchased from Roche Diagnostics (Indianapolis, IN, USA).
β-actin antibodies (cat. no. 66009-1-Ig), PEDF-R rabbit polyclonal
antibodies (cat. no. 55190-1-AP) and RAC family small GTPase 1
(rac1) antibodies (cat. no. 24072-1-AP) were purchased from
ProteinTech Group, Inc. (Chicago, IL, USA). The MitoSOX™ Red
mitochondrial superoxide indicator was purchased from Invitrogen;
Thermo Fisher Scientific, Inc. (Waltham, MA, USA). Hoechst 33342
and the Annexin V-allophycocyanin (APC)/propidium iodide (PI)
Apoptosis Detection kit were purchased from Nanjing KeyGen Biotech
Co., Ltd. (Nanjing, China). Malondialdehyde (MDA; cat. no. S0131),
Dihydroethidium (DHE; cat. no. S0063), Dabco 4 and
6-diamidino-2-phenylindole (DAPI; cat. no. C1005) were purchased
from Beyotime Institute of Biotechnology (Haimen, China). The
TIANamp Genomic DNA kit was purchased from Tiangen Biotech Co.,
Ltd. (Beijing, China). The NADPH oxidase activity assay kit was
purchased from Shanghai Genmed Pharmaceutical Technology Co., Ltd.
(Shanghai, China). The xanthine oxidase activity assay kit was
purchased from Cayman Chemical Company (Ann Arbor, MI, USA).
Recombinant lentivirus constructs and
viral production
Recombinant lentivirus (LV; Shanghai GeneChem Co.,
Ltd, Shanghai, China) was prepared as described previously
(16). PEDF overexpression
plasmid (Shanghai GeneChem Co., Ltd.) was successfully constructed
and then packaged in 293T cells (American Type Culture Collection,
Manassas, VA, USA). PEDF was cloned and ligated into the pGC287
plasmid using AgeI and BamHI sites. The sequences of
the PEDF primers used were as follows: Forward,
5′-CGACCGGTGCCACCATGCAGACCCTGG-3′ and reverse,
5′-GGAATTCGGATCCTTAAGTGCTGCTGG-3′. The concentrated titer of virus
suspension was 2×1012 TU/l. Transient transfection of
H9c2 cells with 20 μM small interfering (si)RNA targeting
the PEDF-R genes (5′-GCGGCATTTCAGACAACTTGC-3′) was performed using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols. A
free-combination sequence of siPEDF-R was used as negative control
(5′-ACGGTTATGCTCGAACTCGCA-3′), all the siRNA were obtained from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Preparation of PEDF protein
Recombinant rat PEDF (GenBank accession no.
NM_177927) was synthesized by Cusabio Biotech, Co., Ltd. (Wuhan,
China). In brief, 293T cells were transfected with the 20 μg
recombinant vector pGEX 6P-1 (GE Healthcare, Chicago, IL, USA),
containing glutathione S-transferase (GST)-tagged PEDF using
Lipofectamine® 3000 according to the manufacturer's
protocols. GST-tagged PEDF proteins were purified by high-pressure
liquid chromatography purification (>90% purity) and
amino-terminal sequence determination (23,24). The resultant proteins were soluble
in aqueous solutions.
Animals
Adult male Sprague-Dawley rats (8-10 weeks old,
weighing 200-250 g, n=85) were purchased from the Experimental
Animal Centre of Xuzhou Medical University (Xuzhou, China) and
housed in a controlled environment (humidity, 50-60%). A total of 3
rats were housed per cage and were maintained at room temperature
under a 12 h light/dark cycle; rats were provided free access to
food and water. The experiments described in this manuscript
conformed to the Guide for the Care and Use of Laboratory Animals
published by the National Institutes of Health (Publication, 8th
Edition, 2011, Bethesda, MD, USA) (25). All animal care and experimental
protocols were approved by the Animal Care and Use Committee of
Xuzhou Medical University (license no. SYXK 2002-0038, Jiangsu,
China) and also followed the international guidelines (European
Council Directive 2010/63/EU) on the ethical use of animals
(26).
Rat MI/R model and intramyocardial gene
delivery
The rat MI/R model was produced as described
previously (27). Intramyocardial
gene delivery was performed one week prior to the MI/R experiment
in the rats. PEDF-LV (2×107 TU) in 20 μl enhanced
infection solution (pH 7.4, Shanghai GeneChem Co., Ltd.) was
delivered with a 20-μl syringe and 25-gauge needle into the
myocardium along the left-anterior descending coronary artery
(LAD). Sham-operated animals underwent an identical surgical
procedure without artery ligation. For the rat MI/R model, after 30
min of ischemia treatment, reperfusion was allowed for 24 h by
releasing the ligatures. The animal models were randomly divided
into five groups as follows: i) Sham group, surgical procedure
without artery ligation (n=18); ii) Sham+P group, surgical
procedure without artery ligation, PEDF-LV was transferred prior to
surgery (n=18); iii) MI group, 0.5 h ischemia (n=6); iv) MI/R
group, 0.5 h ischemia and 24 h reperfusion (n=18); v) MI/R+P group,
0.5 h ischemia and 24 h reperfusion, PEDF-LV was transferred prior
to surgery (n=18).
Quantification of myocardial infarct
size
Myocardial infarct size was measured by Evans
Blue/TTC staining as previously described (28). Briefly, at the end of reperfusion
or ischemia, the LAD was again occluded and 1 ml 3% Evans Blue dye
was retrogradely injected into the ascending aorta to demarcate the
ischemic area at risk (AAR). The heart was rapidly excised and
frozen at −20°C prior to being cut into 1 mm thick sections
perpendicular to the axis of the LAD. At this point, sections were
immediately incubated in 1% TTC in phosphate buffer (pH 7.4) at
37°C for 15 min to discriminate infarcted tissue from viable
myocardium. All sections were scanned from both sides using a color
CCD camera (FV-10; Fujifilm Holdings Corporation, Tokyo, Japan),
and in each slide, infarct area was compared with the AAR using
digital planimetry software (Image-Pro Plus 6.0; Media Cybernetics,
Inc., Rockville, MD, USA). Following correction with the weight of
the sections, the myocardial infarct size was measured and
expressed as a percentage of infarct size over total AAR.
Determination of cardiac function and
dobutamine stress
Echocardiography was conducted under anesthesia with
sodium pentobarbital (30 mg/kg, intraperitoneal; Sigma-Aldrich;
Merck KGaA), as described previously (22). Two-dimensional-guided M-mode
echocardiography was performed at rest and during dobutamine stress
24 h after reperfusion in order to determine left ventricular (LV)
chamber volume at systole and diastole and contractile parameters,
including left ventricular end-diastolic dimension (LVEDD), left
ventricular end-systolic dimension (LVESD), left ventricular
end-diastolic volume (LVEDV) and left ventricular end-systolic
volume (LVESV). Left ventricular ejection fraction (EF) and
fractional shortening (LVFS) were calculated as follows: EF(%) =
(EDV-ESV)/EDV×100. FS(%) = (LVEDD-LVESD)/LVEDD ×100. All
measurements represent the mean of at least 3 consecutive cardiac
cycles. Dobutamine (1 μg/g body weight; Sigma-Aldrich; Merck
KGaA) was given intraperitoneally. Cardiac reserve was investigated
10 min after dobutamine injection.
Cell culture
The embryonic rat heart-derived H9c2 cell line was
obtained from the American Type Culture Collection, and the cells
were cultured in DMEM medium supplemented with 10% foetal bovine
serum (both from Gibco; Thermo Fisher Scientific, Inc.) and 100
mg/ml penicillin/streptomycin at 37°C in a humidified atmosphere
containing 5% CO2. The medium was replaced every 2-3
days, and cells were subcultured or subjected to experimental
procedures at 80-90% confluence.
Establishment of H/R model
To establish the H/R model, the culture medium was
changed to serum-free low glucose DMEM (Gibco; Thermo Fisher
Scientific, Inc.) and placed into a tri-gas incubator (Heal Force
Bio-meditech Holdings, Ltd., Shanghai, China) that was purged with
94% N2, 5% CO2, and 1% O2 for 4, 8
and 12 h respectively. Following hypoxia, reoxygenation was
initiated by incubating in serum-free high glucose DMEM (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C with 5% CO2 for
a further 0 or 2 h. H9C2 cells (~80-90% confluence) were treated
with or without 10 nM PEDF 1 h prior to H/R. The following
experimental groups were included: N group (control, normoxia), N+P
group (PEDF, normoxia), H/R group (control, 8 h hypoxia and 2 h
reoxygenation), H/R+P group (PEDF, H/R), H/R+PEDF+siPEDF-R group
(PEDF+siPEDF-R, H/R), H/R+PEDF+vector group (PEDF+vector, H/R).
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) staining for apoptosis in vivo and
in vitro
Myocardial samples from the rat left ventricle were
embedded in optimum cutting temperature compound tissue medium
(Sakura Finetek Europe B. V., Flemingweg, The Netherlands),
snap-frozen on dry ice and stored at −80°C. Cardiomyocyte apoptosis
in vivo was determined by double-labeling TUNEL
immunofluorescence staining, which was performed with an In
Situ Cell Death Detection kit according to the manufacturer's
protocol. Cardiomyocytes were identified using monoclonal actin
(α-sa) antibodies. Specimens were blocked with 5% bovine serum
albumin (Vicmed Life Sciences, Xuzhou, China) for 30 min at room
temperature prior to incubation with primary antibody. Specimens
were incubated with anti-α-sa (1:300) overnight at 4°C. Specimens
were then washed three times in PBS and incubated with the Goat
anti-Mouse immunoglobulin G (IgG; H+L) Cross-Adsorbed secondary
antibody (Alexa Fluor®488/green; 1:200; A11001; Thermo
Fisher Scientific, Inc.) under light-protected conditions for 1 h
at room temperature. DAPI staining conducted at room temperature
for 15 min was used to count the total number of nuclei.
Cardiomyocyte nuclei with a relatively large diameter are located
within myofibers (16). In
addition, 2×104 H9c2 cells with Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.) were
seeded into each of a 48-well plate. Following the H/R treatments,
H9c2 cells were treated with the In Situ Cell Death
Detection kit according to the manufacturer's protocol. H9c2 cells
were counterstained with DAPI for 30 min at room temperature. The
cardiomyocytes and H9c2 cells were observed under a fluorescence
microscope (Olympus Corporation, Tokyo, Japan). The percentage of
apoptotic cells was calculated as the ratio of the number of
TUNEL-positive cells to the total number of cells, which were
counted in three different random fields of view.
Western blot analysis
For western blot analysis, the cells were
solubilized in lysis buffer (100 mmol/l Tris-HCl, 4% SDS, 20%
glycerine, 200 mmol/l dithiothreitol and protease inhibitors; pH
6.8). Total cellular protein was denatured by boiling for 10 min
with an equal volume of 2X Tris-glycine SDS buffer. Protein
concentration from the supernatant was determined using a BCA
protein assay kit (Pierce; Thermo Fisher Scientific, Inc.). A total
of 50 ng protein per lane was separated by 12% SDS-PAGE and
transferred to nitrocellulose membranes (EMD Millipore, Billerica,
MA, USA). Following blocking with 5% non-fat milk/PBS-T for 3 h at
room temperature, the membranes were incubated with primary
antibodies overnight at 4°C. Cleaved caspase-3, PEDF-R rabbit
polyclonal and rac1 antibodies were used at a dilution of 1:1,000;
β-actin antibody was used at a dilution of 1:5,000. Then,
fluorescence-labeled secondary antibodies was added for 1 h at
37°C: Anti-rabbit IgG H+L DyLight™ 800 4X PEG (1:30,000; cat. no.
5151) and anti-mouse IgG (H+L DyLight™ 680, 1:15,000; cat. no.
5470) (both from Cell Signaling Technology, Inc.) and membranes
were scanned by the Odyssey Infrared Imaging System (LI-COR
Biosciences, Lincoln, NE, USA). ImageJ software (v1.50; NIH,
Bethesda, MD, USA) was used for quantification.
Determination of apoptosis by flow
cytometry
The Annexin V-APC/PI Apoptosis Detection kit was
used for the following assay. H9c2 cardiomyocytes (1×106
per group) were collected, washed twice with PBS, resuspended with
500 μl 1X binding buffer, treated with 5 μl Annexin
V-APC and 5 μl PI, and placed in the dark at room
temperature for 5-15 min. Then, the cells were analyzed using a
flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Detection of intracellular ROS
generation
DHE was used to stain for intracellular ROS, as
previously described (29). DHE,
an oxidant-sensitive probe, is widely used for the detection of
ROS. Harvested cells (2×104 per well) were incubated
with 10 μM DHE for 30 min at 37°C, according to the
manufacturer's protocol, then washed with DMEM without FBS three
times. Fluorescence was observed under a fluorescence microscope.
Fluorescence was calculated by viewing in four randomly selected
fields for each group; Image-Pro Plus software (v6.0; Media
Cybernetics, Inc.) was used for quantification.
Detection of mitochondrial (mt)ROS
production
To assess the levels of mtROS, MitoSOX™, a
fluorochrome specific to anion superoxide produced in the inner
mitochondrial compartment (Invitrogen; Thermo Fisher Scientific,
Inc.) was used. H9c2 cells (1×104 per well) were seeded
into each well of a 48-well plate, underwent their respective
treatments as aforementioned, and subsequently loaded with 200
μl MitoSOX™ (5 mM stock in ethanol dissolved in Hank's
Balanced Salt Solution to a working solution of 5 μM) for 10
min at 37°C. After washing three times with PBS, nuclei were
counter-stained with Hoechst 33342 for 15 min at 37°C. Following
three washes with PBS, the sample was observed by a fluorescence
microscope (Olympus IX73; Olympus Corporation). Fluorescence was
calculated by viewing in five randomly selected fields for each
group. Image-Pro Plus software (v6.0; Media Cybernetics, Inc.) was
used for quantification.
Quantitative polymerase chain reaction
(qPCR)
Total DNA was isolated from cells using the TIANamp
Genomic DNA kit, according to the manufacturer's protocol. As
mitochondrial DNA (mtDNA) is a primary target of ROS and a
reflection of ETC, mtDNA copy number was detected by qPCR using
SYBR Green PCR Master Mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.). PCR was conducted with a final volume of 20
μl containing 10 μl 2× SYBR-Green PCR Master Mix, 0.1
μM of each primer and 100 μg genomic DNA. The mixture
was subjected to qPCR amplification for 95°C for 10 min, 45 cycles
(95°C for 10 sec, 60°C for 10 sec, 72°C for 20 sec), 1 cycle (95°C
for 1 min, 65°C for 1 min and 97°C with continuous) and then cooled
to 40°C for 30 sec using a Roche Light Cycler 480 (Roche
Diagnostics). Gene expression was normalized to that of 18s RNA.
Gene expression was quantified by using the 2−∆∆Cq
method (30). The following
primers, synthesized by GenScript (Piscataway, NJ, USA) were used:
D-loop forward, 5′-TGGTTCATCGTCCATACGTT-3′ and reverse,
5′-TGACGGCTATGTTGAGGAAG-3′; 18sRNA forward,
5′-CATTCGAACGTCTGCCCTATC-3′ and reverse,
5′-CCTGCTGCCTTCCTTGGA-3′.
Measurement of MDA levels, XO activity
and NOX activity
MDA levels, XO activity and NOX activity were
measured using the respective detection kits, according to the
manufacturers' protocol.
Statistical analysis
The results are expressed as the mean ± standard
error of the mean. All statistical analyses were conducted with
SPSS 19.0 for Windows (IBM Corp., Armonk, NY, USA). The results
were analyzed using unpaired Student's t-test or repeated-measures
one-way analysis of variance followed by Duncan's new multiple
range method or Fisher's least significant difference test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
PEDF reduces myocardial infarct size and
improves cardiac function under MI/R condition
First, the protective effects of PEDF were verified
in a MI/R rat model. As presented in Fig. 1A and B, an irregular small
myocardial infarct area was observed in 3 of 6 hearts in the MI
group, and infarcted myocar-dium was not observed in the rest of
the MI group. However, the MI/R group had a significantly increased
myocardial infarct size compared with the MI group. In addition,
PEDF treatment resulted in a reduced myocardial infarct size
compared with the MI/R group. Next, TUNEL staining was used to
analyze the anti-apoptotic effect of PEDF in the I/R myocardium.
There was a marked increase in cardiomyocyte apoptosis in MI/R
hearts compared with the sham group, and treatment with PEDF
reduced this effect (Fig. 1C and
D). There was no significant difference between the Sham group
and Sham+P group.
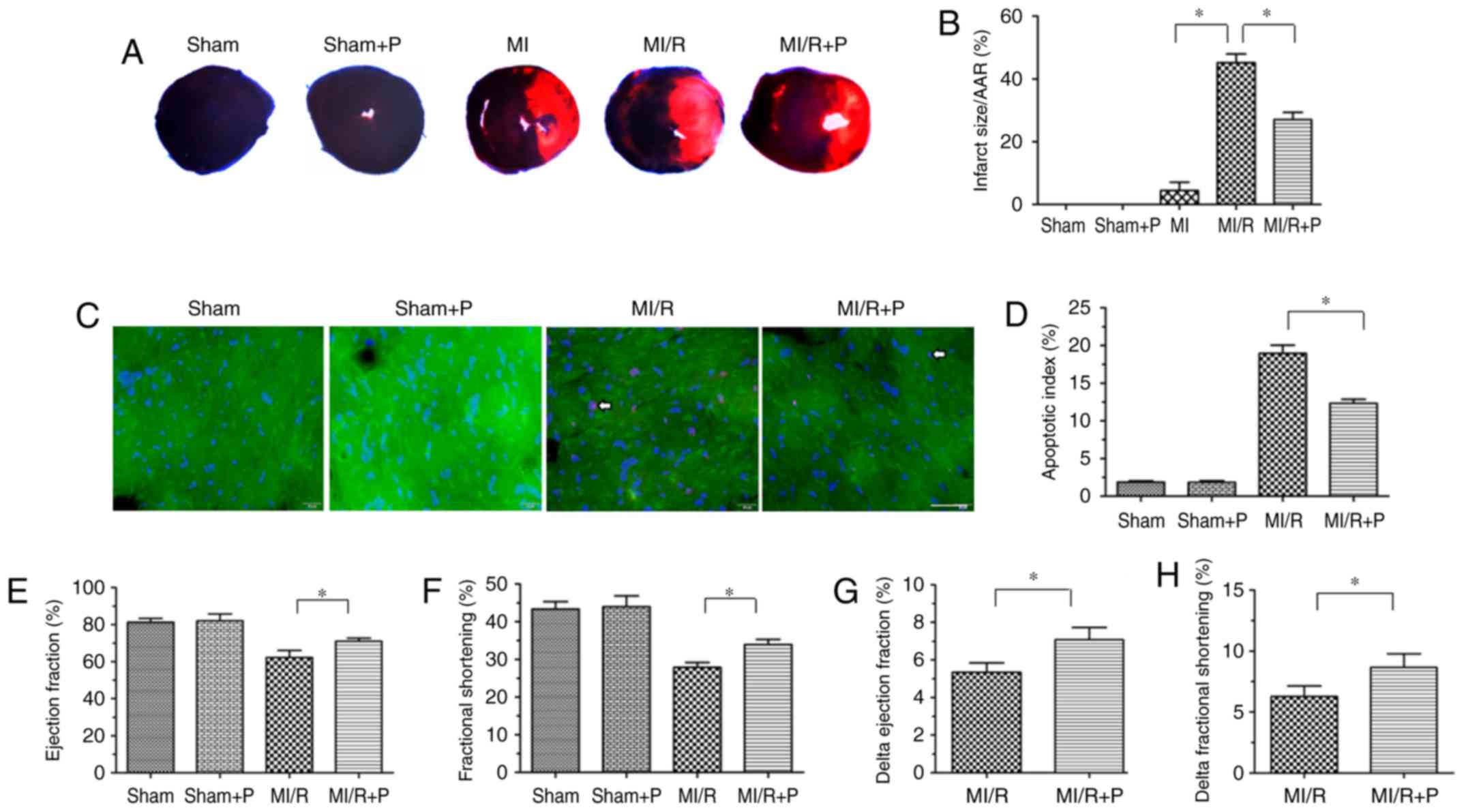 | Figure 1PEDF reduces myocardial infarct size
and improves cardiac function under MI/R conditions. (A)
Representative figures of the Evans Blue/TTC-stained myocardial
tissues in each indicated experimental condition, with (B)
quantification of the infarct size (n=6). (C) Representative images
of TUNEL staining of cardiomyocyte apoptosis (white arrow), with
(D) quantification. Cardiomyocyte apoptosis was measured by TUNEL
staining; cardiomyocytes were identified using α-sarcomeric actin
antibodies. TUNEL staining for cardiomyocyte apoptosis (red), DAPI
for nuclear staining (blue) and α-sarcomeric actin for
cardiomyocytes (green) in the border zone of the infarcted left
ventricle from all experimental groups (scale bar=20 μm;
n=6). (E) Left ventricular ejection fraction prior to dobutamine
injection determination by echocardiography (n=6). (F) Left
ventricular fractional shortening prior to dobutamine injection
determination by echocardiography (n=6). (G) Δ left ventricular
ejection fraction following dobutamine injection by
echocardiography (n=6). (H) Δ left ventricular fractional
shortening following dobutamine injection by echocardiography
(n=6). Data are expressed as the mean ± standard error of the mean.
*P<0.05, with comparisons indicated by lines. PEDF,
pigment epithelium-derived factor; MI/R, myocardial
ischemia/reperfusion; TTC, 2,3,5-triphenyltetrazolium; TUNEL,
terminal deoxynucleotidyl transferase dUTP nick end labeling. |
Cardiac function and cardiac functional reserve were
measured using transthoracic M-mode echocardiography prior to and
following dobutamine (1 μg/g) injection. The values of
ejection fraction and fractional shortening, which are signs of
myocardial function, were significantly increased in PEDF-treated
rats compared with untreated MI/R hearts (Fig. 1E and F). The Δ ejection fraction
and Δ fractional shortening to dobutamine infusions were increased
in hearts transfected with PEDF compared with MI/R rats in
vivo, which revealed that PEDF may increase cardiac functional
reserve (Fig. 1G and H).
Together, these results indicated that PEDF has a protective effect
against I/R damage in rat hearts.
PEDF suppresses H/R-induced apoptosis in
H9c2 cardiomyocytes
To investigate whether PEDF suppresses H9c2 cell
apoptosis under H/R conditions, the level of the cleaved caspase-3
following H/R was dected in H9c2 cells at different time points
using western blotting. Onset of hypoxia for up to 8 h resulted in
an increase in cleaved caspase-3 protein expression, and cleaved
caspase-3 expression was also significantly increased in the
hypoxia (8 h)/reoxygenation (2 h) group in comparison with the
hypoxia (8 h) group (Fig. 2A and
B). Treatment with PEDF could significantly reduce the level of
the cleaved caspase-3 compared with the H/R group (Fig. 2C and D). To further confirm the
observation that PEDF treatment inhibited H9c2 cell apoptosis under
H/R condition, flow cytometric detection of early apoptosis was
performed. The results demonstrated similar trends (Fig. 2E and F). These observations
indicated that PEDF prevented H/R-induced H9c2 cell apoptosis.
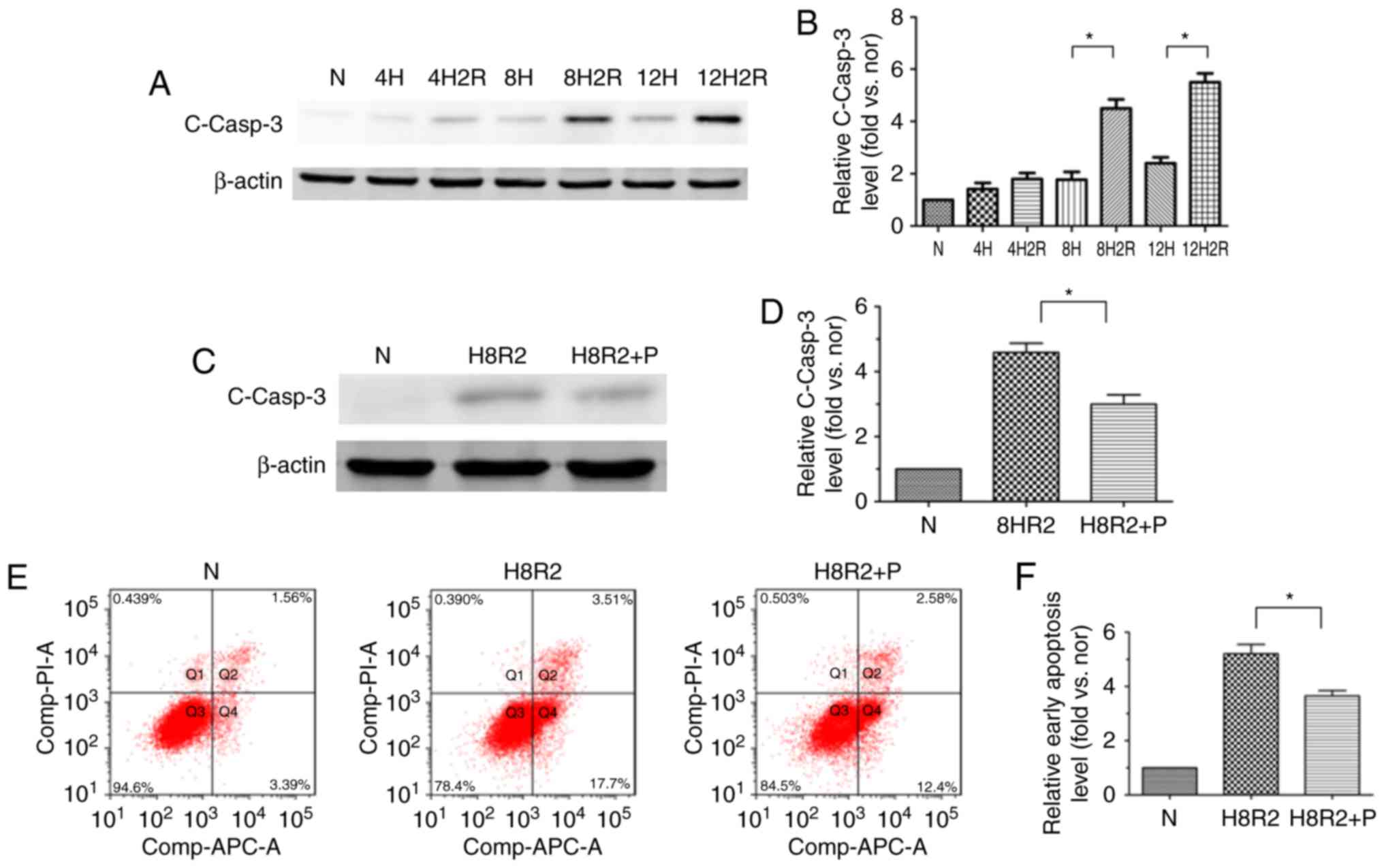 | Figure 2PEDF protects H9c2 cells against
H/R-induced apoptosis. H9c2 cells were exposed to H/R conditions
for various durations (0/0, 4/0, 4/2, 8/0, 8/2, 12/0 and 12/2 h).
In addition, H9c2 cells treated with or without PEDF (10 nM) were
exposed to normoxic or hypoxic/reoxygenation condition for 8/2 h.
(A) Western blotting detected the level of the cleaved casp3
protein, with (B) quantification (n=3). (C) Samples were collected
and analyzed for the expression of the cleaved casp3 protein by
western blotting analysis, with (D) quantification (n=4). (E) Flow
cytometric detection of early apoptosis
(APC+/PI−), with (F) quantification (n=3).
Data are expressed as the mean ± standard error of the mean.
*P<0.05, with comparisons indicated by lines. PEDF,
pigment epithelium-derived factor; H/R, hypoxia/reoxygenation;
casp3, caspase-3; APC, allophycocyanin; PI, propidium iodide; N,
negative control. |
PEDF protects H9c2 cells against
H/R-induced apoptosis via PEDF-R
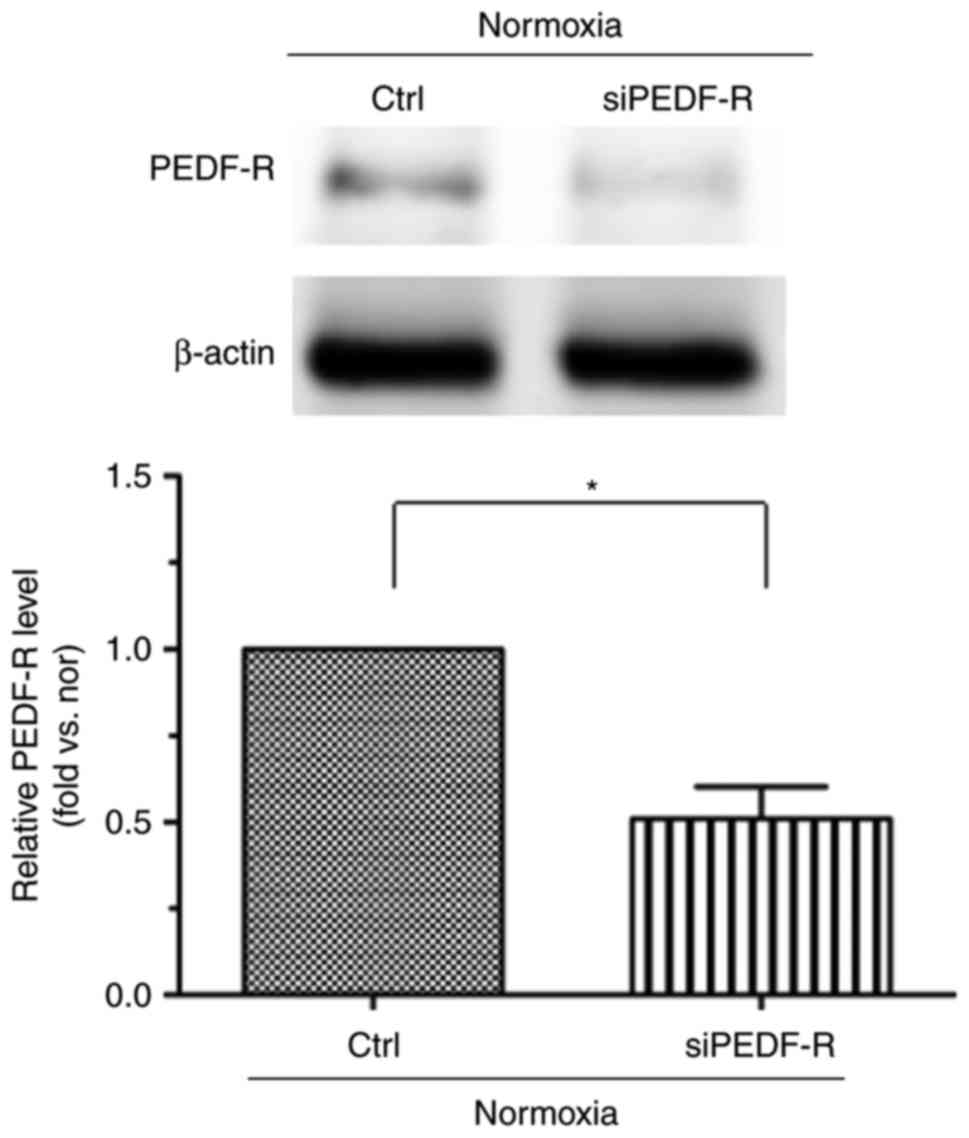
Next, to investigate whether PEDF-R is involved in
the PEDF-mediated repression of cell injury, H9c2 cells were
treated with PEDF under normoxic and H/R conditions, and RNA
interference assays were used to silence PEDF-R. PEDF-R siRNA
significantly reduced PEDF-R expression levels under normoxic
conditions in H9c2 cells (Fig.
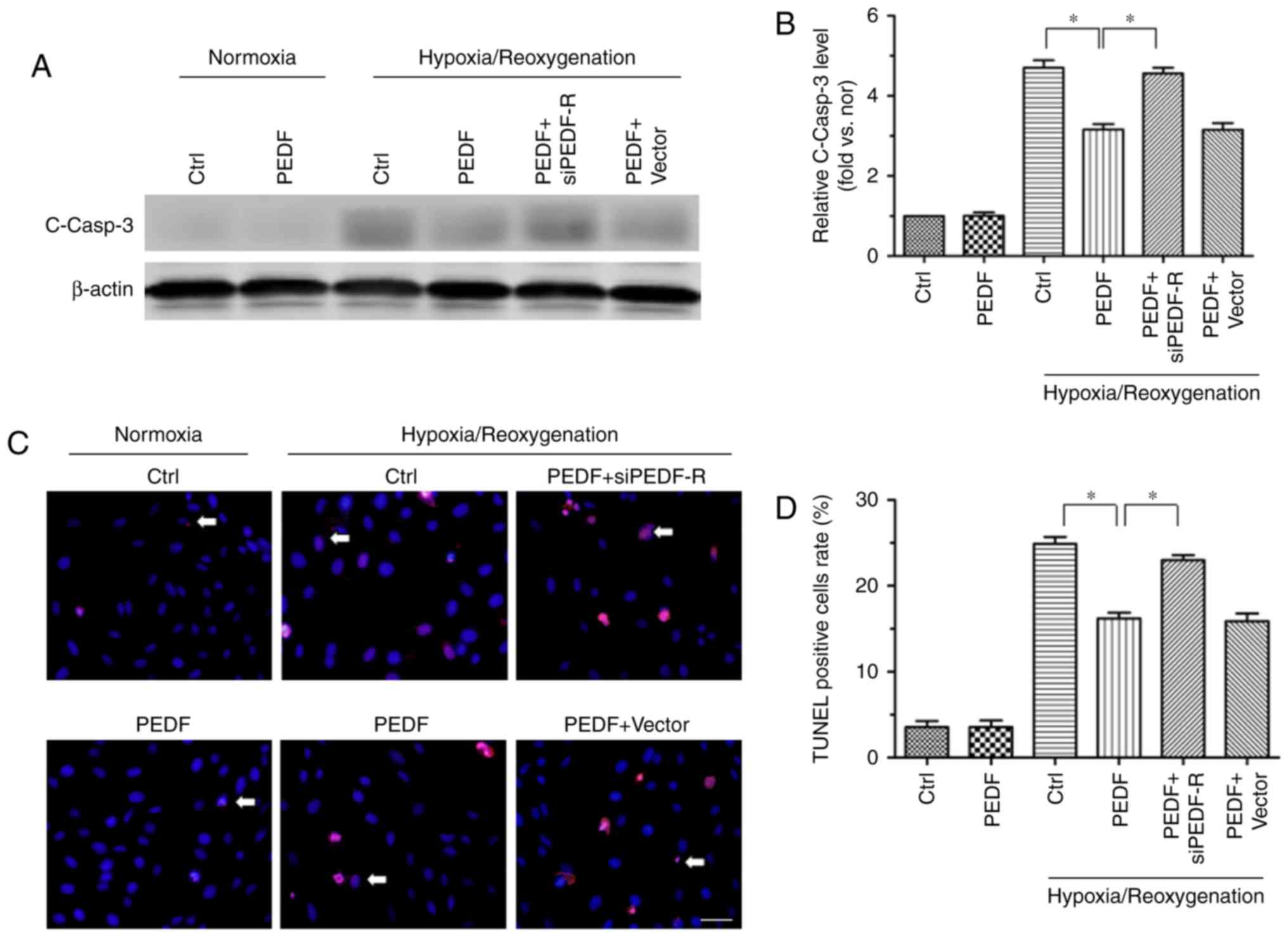
3). As presented in Fig. 4A and
B, PEDF-R siRNA prevented the PEDF-induced reduction of cleaved
caspase-3 protein expression under H/R conditions in H9c2 cells.
This was not observed in the PEDF+vector group. TUNEL staining was
also used to identify apoptotic H9c2 cells, and the results
revealed the critical involvement of PEDF-R in the effect of PEDF
in H/R injury (Fig. 4C and D).
These results suggested that the anti-apoptotic effect of PEDF is
dependent on PEDF-R.
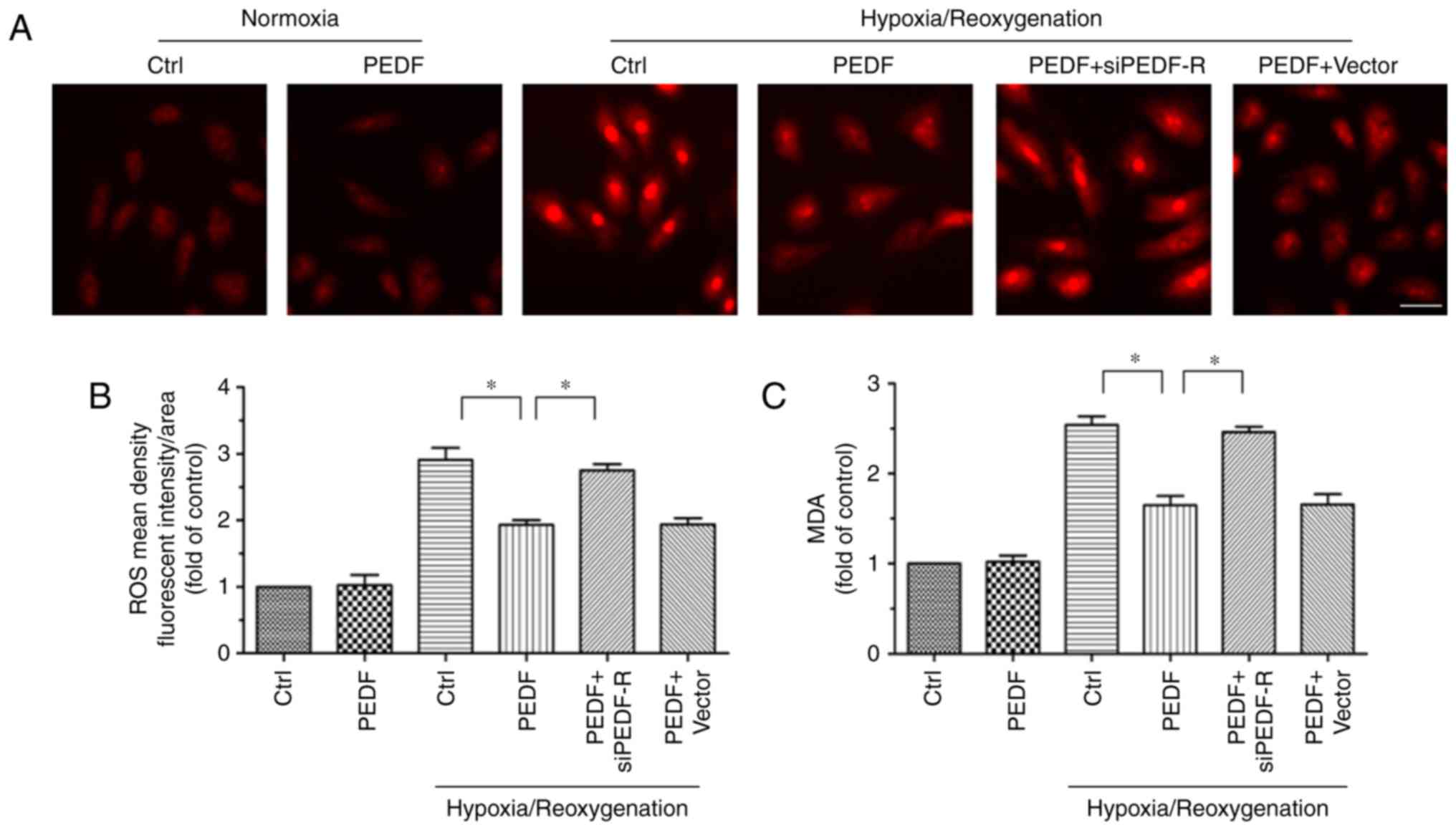
PEDF reduces H/R-induced the burst of ROS
via PEDF-R
The ROS burst is a key factor of myocardial
ischemia/reperfusion injury. Therefore, whether PEDF inhibited the
H/R-induced burst of ROS via PEDF-R was investigated. The changes
in ROS levels in H9c2 cells are presented in Fig. 5A and B. Compared with the normal
group, ROS levels were significantly increased in the H/R groups.
Nonetheless, PEDF markedly decreased the level of ROS induced by
H/R, while this effect was reversed by PEDF-R siRNA. Intracellular
MDA levels were measured to further confirm the antioxidative
function of PEDF. In the H/R group, MDA content was increased
compared with the normal group. PEDF dampened the MDA increase, but
PEDF-R siRNA attenuated this effect (Fig. 5C). These results suggested that
PEDF has antioxidant activities in response to H/R injury in H9c2
cells, and these activities are dependent on PEDF-R.
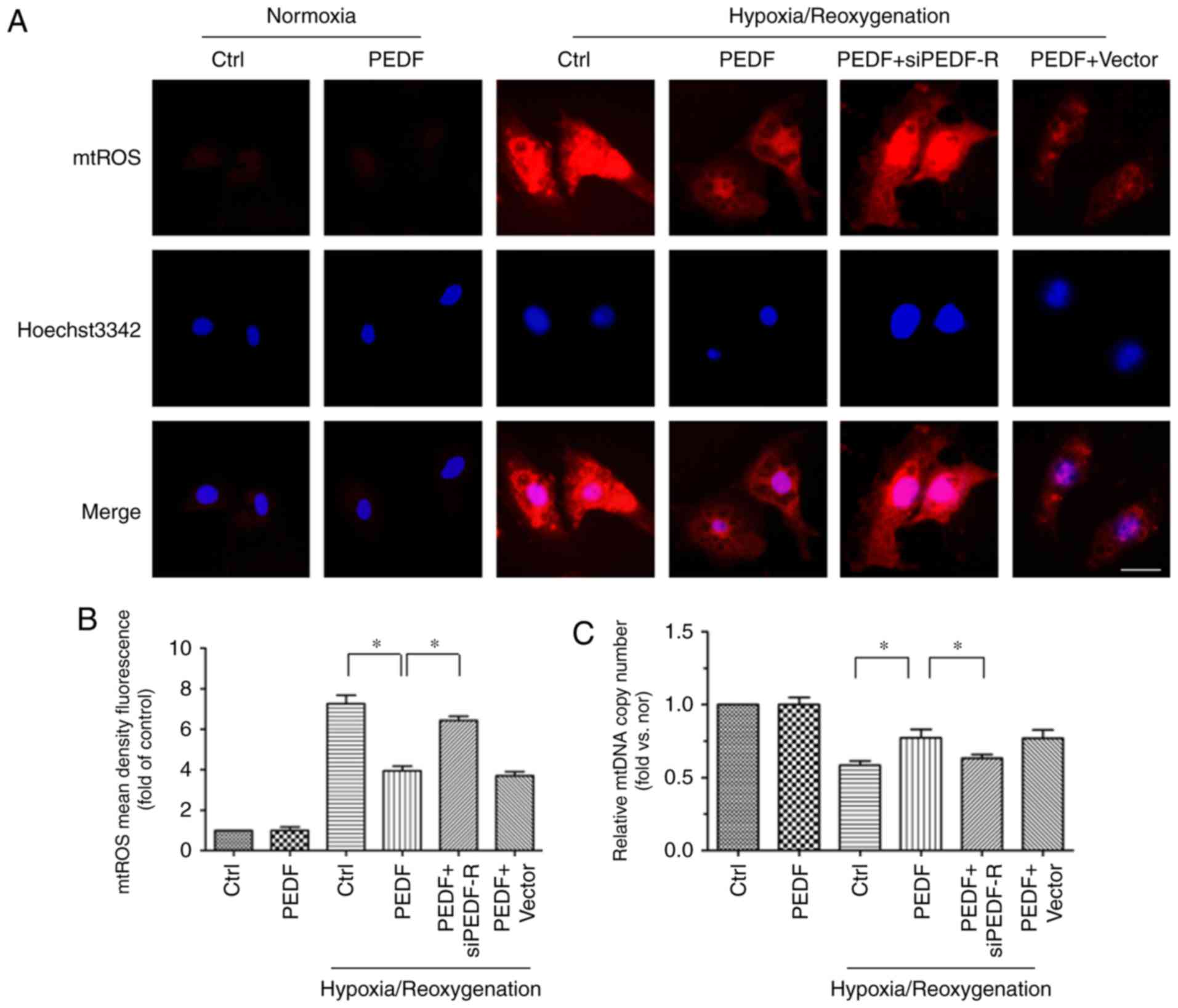
PEDF decreases H/R-induced mtROS
generation via PEDF-R
To study the detailed mechanisms underlying the
antioxidative effect of PEDF, which protects cardiomyocytes against
H/R-induced apoptosis, our group developed a novel hypothesis: That
PEDF attenuates H/R-induced oxidative stress via PEDF-R through
inhibition of ROS generation. The main source of ROS is the
mitochondria, which produce ROS primarily through single electron
transport to molecular oxygen in the ETC (11). In addition, mtDNA is the primary
target of the ROS, and a reflection of ETC function (31). Thus, mtROS level and the mtDNA
copy number were measured to investigate mitochondrial-derived ROS
synthesis. Under H/R conditions, mtROS levels were increased while
the mtDNA copy number was decreased. This effect was reversed by
PEDF treatment, but PEDF-R siRNA inhibited this effect (Fig. 6). The results demonstrated that
PEDF attenuates the H/R-induced burst of ROS by reducing mtROS
generation via PEDF-R.
PEDF decreases H/R-induced cytoplasmic
ROS generation via PEDF-R
NOX and XO are the main sources of ROS formation in
the cytoplasm (11), thus,
examination of XO and NOX activity permits the evaluation of
cytoplasmic ROS production. XO and NOX activity was increased in
H/R and H9c2 cells and, while PEDF reduced thid activity, PEDF-R
siRNA reversed this effect (Fig. 7A
and B). In addition, the expression of rac1, which has an
important effect on NOX activation (32), was detected. Rac1 protein
expression was altered in a similar manner to NOX activity
(Fig. 7C and D). These
observations suggested that PEDF attenuates the H/R-induced ROS
burst by reducing cytoplasmic ROS generation via PEDF-R.
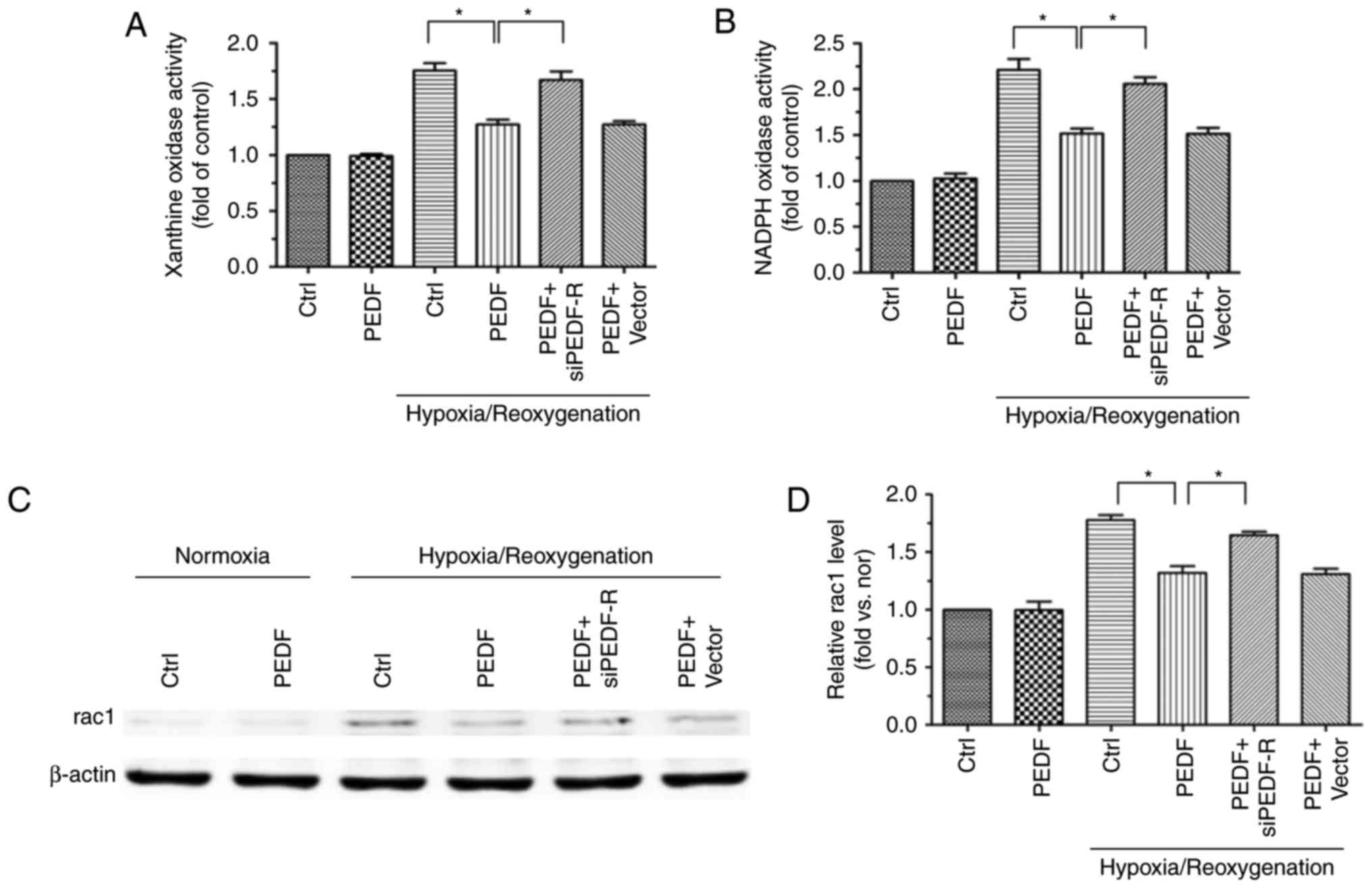 | Figure 7PEDF decreases H/R-induced
cytoplasmic ROS generation via PEDF-R. H9c2 cells were maintained
in normoxic or H/R conditions for 8/2 h with or without PEDF (10
nM). RNA interference assays were used to silence PEDF-R. (A) XO
activity was assessed in all experimental groups using the XO
activity assay kit (n=4). (B) NOX activity was assessed in all
experimental groups using the NOX activity assay kit (n=4). (C)
Western blot analysis of rac1 protein expression, with (D)
quantification (n=4). Data are expressed as the mean ± standard
error of the mean. *P<0.05, with comparisons
indicated by lines. PEDF, pigment epithelium-derived factor; H/R,
hypoxia/reoxygenation; ROS, reactive oxygen species; PEDF-R,
pigment epithelium-derived factor receptor; XO, xanthine oxidase;
NOX, NADPH oxidase; rac1, RAC family small GTPase 1; si, small
interfering. |
Discussion
In the present study, PEDF was demonstrated to
reduce myocardial infarct size and improve cardiac function in a
rat MI/R model. In addition, the mechanism underlying effect was
verified: PEDF reduced H/R-induced cell injury by attenuating ROS
generation, and in particular mtROS generation, via PEDF-R.
A large number of mitochondria are typically present
in cardiomyocytes (33). ROS,
initially produced by damaged mitochondria, induce the opening of
the mitochondrial membrane permeability transition pore, induce the
dissipation of mitochondrial membrane potential (ΔΨm) and block the
electron transport chain, which leads to the ROS burst (34,35). This process is known as
ROS-induced ROS release, and this is one of the factors that
induces cell injury (34,35). Mitochondria are not only the main
target of ROS damage, but also the primary site of ROS production
(6). Therefore, it is important
to reduce the generation of mitochondrial (mt)ROS in
cardiomyocytes. A previous study by our group demonstrated that
PEDF decreases mtROS by inhibiting mitochondrial fission in hypoxic
cardiomyocytes (36). However,
the explosive generation of mtROS appears during the reperfusion
process due to the transient increase of oxygen concentration and
the effect of inefficient electron transfer (6). In this case, whether PEDF is able to
continue to reduce mtROS generation is worthy of further
investigation. In the present study, PEDF was revealed to
significantly decreased mtROS levels, increase mitochondrial DNA
copy number, reduce xanthine oxidase and NADPH oxidase activity and
decrease rac1 protein expression, compared with the H/R group. The
results from the present study demonstrated that PEDF may attenuate
MI/R-induced mitochondrial and cytosolic ROS formation.
Under the condition of the present study, the
results of Evans Blue/TTC staining revealed that small myocardial
infarctions occurred in certain individuals from the MI (0.5 h)
group. However, the MI (0.5 h)/R (24 h) group had wider range of
myocardial infarction compared with the MI (0.5 h) group. In the
in vitro experiments, the apoptosis of H9c2 cardiomyocytes
was relatively mild 8 h after hypoxia, and was significantly
increased in the H (8 h)/R (2 h) group. Based on these results,
reperfusion (24 h) following ischemia (0.5 h) were selected in
vivo and reoxygenation (2 h) following hypoxia (8 h) were
selected in vitro as the optimal time points to study the
effect of PEDF in MI/R injury and the underlying mechanisms. The
protective effect of PEDF on cardiomyocytes during MI/R process may
be best reflected in the protective effect of PEDF on
cardiomyocytes during reperfusion.
Multiple previous studies have focused on the
function of PEDF in hypoxic cardiomyocytes, rather than the
functions and mechanisms of PEDF in the MI/R process. A previous
study has demonstrated that that PEDF reduces the levels of ROS in
hypoxia-reoxygenated human cardiomyocytes (37), consistent with the results of the
present study. Furthermore, in addition to detecting the decrease
in ROS levels, the present study demonstrated that PEDF has the
potential to inhibit the formation of ROS through further studies.
To the best of our knowledge, the present study has demonstrated
for the first time that PEDF significantly reduces myocardial
infarct size in rats undergoing the MI/R process, while protecting
cardiac function. In addition, the present study demonstrated that
PEDF significantly decreased H9c2 cardiomyocyte apoptosis caused by
H/R via PEDF-R in vitro, which is associated with the
suppression of ROS generation.
In the present study, a previously unknown
association between PEDF and ROS generation in the MI/R process was
demonstrated. However, further studies are required to fully
explore the detailed mechanisms underlying PEDF-induced regulation
of MI/R-induced ROS generation in vivo. In addition, the
results derived from H9c2 cells may not represent the same
mechanisms as in primary cardiomyocytes. This limitation of the
present study means that further research is required. In
conclusion, the present study confirmed that PEDF protects
cardiomyocytes against MI/R injury by reducing ROS production. The
results of the present study suggest that PEDF represents a
promising novel therapeutic approach for MI/R injury.
Acknowledgments
We would like to express our thanks to the Research
Facility Center for Morphology of Xuzhou Medical University
(Xuzhou, China), their research site and facilities provided an
effective means to conduct our research.
Notes
[1]
Funding
The present study was supported by the National
Nature Science Foundation of China (grant no. 81570242) and the
Natural Science Foundation of Jiangsu Province (grant no.
BK20150207).
[2] Availability
of data and materials
We declared that materials described in the
manuscript, including all relevant raw data, will be freely
available to any scientist wishing to use them for non-commercial
purposes, without breaching participant confidentiality.
[3] Authors'
contributions
QZ, ZL, HD and ZZ conceived and designed the
experiments. QZ, ZL, BH, YY, XL, HZ and YZ performed the
experiments. QZ, ZL, YL, HD and ZZ analyzed the data. QZ, ZL, FQ,
HM and YZ acquired the reagents, materials and tools for analysis.
QZ and ZL produced the manuscript.
[4] Ethics
approval and consent to participate
The experiments described in this manuscript conform
to the Guide for the Care and Use of Laboratory Animals published
by the National Institutes of Health (Publication, 8th Edition,
2011, Bethesda, MD, USA) (25).
All animal care and experimental protocols were approved by the
Animal Care and Use Committee of Xuzhou Medical University (license
no. SYXK 2002-0038, Jiangsu, China) and also followed the
international guidelines (European Council Directive 2010/63/EU) on
the ethical use of animals (26).
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Damiani G, Salvatori E, Silvestrini G,
Ivanova I, Bojovic L, Iodice L and Ricciardi W: Influence of
socioeconomic factors on hospital readmissions for heart failure
and acute myocardial infarction in patients 65 years and older:
Evidence from a systematic review. Clin Interv Aging. 10:237–245.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Crea F, Battipaglia I and Andreotti F: Sex
differences in mechanisms, presentation and management of ischaemic
heart disease. Atherosclerosis. 241:157–168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hearse DJ: Myocardial protection during
ischemia and reperfusion. Mol Cell Biochem. 186:177–184. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hearse DJ and Bolli R: Reperfusion induced
injury: Manifestations, mechanisms, and clinical relevance.
Cardiovasc Res. 26:101–108. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Turer AT and Hill JA: Pathogenesis of
myocardial ischemia-reperfusion injury and rationale for therapy.
Am J Cardiol. 106:360–368. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Murphy E and Steenbergen C: Mechanisms
underlying acute protection from cardiac ischemia-reperfusion
injury. Physiol Rev. 88:581–609. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Minamino T: Cardioprotection from
ischemia/reperfusion injury: Basic and translational research. Circ
J. 76:1074–1082. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Braunersreuther V and Jaquet V: Reactive
oxygen species in myocardial reperfusion injury: From
physiopathology to therapeutic approaches. Curr Pharm Biotechnol.
13:97–114. 2012. View Article : Google Scholar
|
|
9
|
Radak Z, Zhao Z, Goto S and Koltai E:
Age-associated neuro-degeneration and oxidative damage to lipids,
proteins and DNA. Mol Aspects Med. 32:305–315. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Campos JC, Bozi LH, Bechara LR, Lima VM
and Ferreira JC: Mitochondrial quality control in cardiac diseases.
Front Physiol. 7:4792016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Afanas'ev I: ROS and RNS signaling in
heart disorders: Could antioxidant treatment be successful? Oxid
Med Cell Longev. 2011:2937692011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Becerra SP, Sagasti A, Spinella P and
Notario V: Pigment epithelium-derived factor behaves like a
noninhibitory serpin. Neurotrophic activity does not require the
serpin reactive loop. J Biol Chem. 270:25992–25999. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rychli K, Kaun C, Hohensinner PJ, Dorfner
AJ, Pfaffenberger S, Niessner A, Bauer M, Dietl W, Podesser BK,
Maurer G, et al: The anti-angiogenic factor PEDF is present in the
human heart and is regulated by anoxia in cardiac myocytes and
fibroblasts. J Cell Mol Med. 14:198–205. 2010. View Article : Google Scholar :
|
|
14
|
Gao X, Zhang H, Zhuang W, Yuan G, Sun T,
Jiang X, Zhou Z, Yuan H, Zhang Z and Dong H: PEDF and PEDF-derived
peptide 44mer protect cardiomyocytes against hypoxia-induced
apoptosis and necroptosis via anti-oxidative effect. Sci Rep.
4:56372014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang H, Sun T, Jiang X, Yu H, Wang M, Wei
T, Cui H, Zhuang W, Liu Z, Zhang Z and Dong H: PEDF and
PEDF-derived peptide 44mer stimulate cardiac triglyceride
degradation via ATGL. J Transl Med. 13:682015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang H, Wang Z, Feng SJ, Xu L, Shi HX,
Chen LL, Yuan GD, Yan W, Zhuang W, Zhang YQ, et al: PEDF improves
cardiac function in rats with acute myocardial infarction via
inhibiting vascular permeability and cardiomyocyte apoptosis. Int J
Mol Sci. 16:5618–5634. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Notari L, Baladron V, Aroca-Aguilar JD,
Balko N, Heredia R, Meyer C, Notario PM, Saravanamuthu S, Nueda ML,
Sanchez-Sanchez F, et al: Identification of a lipase-linked cell
membrane receptor for pigment epithelium-derived factor. J Biol
Chem. 281:38022–38037. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bernard A, Gao-Li J, Franco CA, Bouceba T,
Huet A and Li Z: Laminin receptor involvement in the
anti-angiogenic activity of pigment epithelium-derived factor. J
Biol Chem. 284:10480–10490. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zimmermann R, Strauss JG, Haemmerle G,
Schoiswohl G, Birner-Gruenberger R, Riederer M, Lass A, Neuberger
G, Eisenhaber F, Hermetter A and Zechner R: Fat mobilization in
adipose tissue is promoted by adipose triglyceride lipase. Science.
306:1383–1386. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hirsch J, Johnson CL, Nelius T, Kennedy R,
Riese Wd and Filleur S: PEDF inhibits IL8 production in prostate
cancer cells through PEDF receptor/phospholipase A2 and regulation
of NFκB and PPARγ. Cytokine. 55:202–210. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang XY, Zhang YQ, Lu P, Zhang H, Li Y,
Dong H and Zhang Z: PEDF attenuates hypoxia-induced apoptosis and
necrosis in H9c2 cells by inhibiting p53 mitochondrial
translocation via PEDF-R. Biochem Biophys Res Commun. 465:394–401.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu P, Zhang YQ, Zhang H, Li YF, Wang XY,
Xu H, Liu ZW, Li L, Dong HY and Zhang ZM: Pigment
epithelium-derived factor (PEDF) improves ischemic cardiac
functional reserve through decreasing hypoxic cardiomyocyte
contractility through PEDF receptor (PEDF-R). J Am Heart Assoc.
5:e0031792016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Reim DF and Speicher DW: N-terminal
sequence analysis of proteins and peptides. Curr Protoc Protein
Sci. Chapter 11: Unit 11.10. 2001.
|
|
24
|
Chen L, Zhao X, Liang G, Sun J, Lin Z, Hu
R, Chen P, Zhang Z, Zhou L and Li Y: Recombinant SFRP5 protein
significantly alleviated intrahepatic inflammation of nonalcoholic
steatohepatitis. Nutr Metab (Lond). 14:562017. View Article : Google Scholar
|
|
25
|
National Research Council: Guide for the
Care and Use of Laboratory Animals. 8th edition. The National
Academies Press; Washington, DC: 2011
|
|
26
|
European Commission: Directive 2010/63/EU
of the European Parliament and of the council of 22 September 2010
on the protection of animals used for scientific purposes. Off J
Eur Union. L276:33–79. 2010.
|
|
27
|
Zhang Y, Li Y, Wang X, Zhao Q, Lu P, Zhang
H, Dong H and Zhang Z: A closed-chest rat model of myocardial
isch-aemia/reperfusion supports direct intramyocardial gene
delivery. Exp Ther Med. In press.
|
|
28
|
Black SC and Rodger IW: Methods for
studying experimental myocardial ischemic and reperfusion injury. J
Pharmacol Toxicol Methods. 35:179–190. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peshavariya HM, Dusting GJ and Selemidis
S: Analysis of dihydroethidium fluorescence for the detection of
intracellular and extracellular superoxide produced by NADPH
oxidase. Free Radic Res. 41:699–712. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Tsutsui H, Kinugawa S and Matsushima S:
Oxidative stress and mitochondrial DNA damage in heart failure.
Circ J. 72(Suppl A): A31–A37. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li JM, Zhu HQ, Shen E, Wan L, Arnold JMO
and Peng TQ: Deficiency of Rac1 blocks NADPH oxidase activation,
inhibits endoplasmic reticulum stress, and reduces myocardial
remodeling in a mouse model of type 1 diabetes. Diabetes.
59:2033–2042. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Garcia-Dorado D, Andres-Villarreal M,
Ruiz-Meana M, Inserte J and Barba I: Myocardial edema: A
translational view. J Mol Cell Cardiol. 52:931–939. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gustafsson AB and Gottlieb RA: Heart
mitochondria: Gates of life and death. Cardiovasc Res. 77:334–343.
2008. View Article : Google Scholar
|
|
35
|
Zorov DB, Filburn CR, Klotz LO, Zweier JL
and Sollott SJ: Reactive oxygen species (ROS)-induced ROS release:
A new phenomenon accompanying induction of the mitochondrial
permeability transition in cardiac myocytes. J Exp Med 1.
92:1001–1014. 2000. View Article : Google Scholar
|
|
36
|
Zhou ZX, Wang Z, Guan QH, Qiu F, Li Y, Liu
Z, Zhang H, Dong H and Zhang Z: PEDF inhibits the activation of
NLRP3 inflammasome in hypoxia cardiomyocytes through PEDF
receptor/phospholipase A2. Int J Mol Sci. 17:E20642016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kuo HF, Liu PL, Chong IW, Liu YP, Chen YH,
Ku PM, Li CY, Chen HH, Chiang HC, Wang CL, et al: Pigment
epithelium-derived factor mediates autophagy and apoptosis in
myocardial hypoxia/reoxygenation injury. PLoS One. 11:e01560592016.
View Article : Google Scholar : PubMed/NCBI
|