Introduction
Abnormal proliferation of vascular smooth muscle
cells (VSMCs) is believed to serve an important role in the
formation of atherosclerotic plaques and restenosis following
percutaneous coronary intervention (1,2).
Vascular remodeling has recently garnered increasing attention due
to its critical role in the progression of vascular occlusion
diseases (3). During vascular
lesion formation, phenotypic switching of VSMCs from the
physiological contractile phenotype to the pathophysiological
synthetic phenotype occurs, and VSMCs migrate to the intima
(4). Under normal conditions,
VSMCs constitute the major structural component of the vasculature,
and are crucial for maintaining vessel tone, blood pressure and
blood flow. In the media layer of mature blood vessels, VSMCs
exhibit a differentiated and contractile phenotype, characterized
by the expression of contractile proteins, including α-smooth
muscle actin (α-SMA), calponin and smooth muscle heavy chain
(5). In various vascular
pathologies, VSMCs undergo a rapid and reversible alteration from a
quiescent contractile phenotype to a proliferative and secretory
phenotype, which is characterized by increased proliferation,
migration and extracellular matrix (ECM) production (6), and osteopontin (OPN) is the main
protein marker of synthetic phenotypes. The synthetic phenotype of
VSMCs acts as a critical factor in various cardiovascular diseases,
including atherosclerosis, restenosis after angioplasty or bypass,
and hypertension (7,8).
It is well known that numerous cytokines and growth
factors are released to stimulate VSMC proliferation during
vascular injury repair (9,10).
Angiotensin II (Ang II), which has previously been reported to
serve an important role in normal vascular physiology and
cardiovascular disease, is a biologically active peptide of the
renin-angiotensin system, which modulates vascular tone and
structure. Ang II interacts with the type 1 receptor on SMCs and
promotes a switch from a contractile to a synthetic phenotype of
VSMCs, leading to changes in the contractile machinery and VSMC
proliferation (11,12). In addition, Ang II may promote the
production of various growth-stimulating factors, including
fibroblast growth factor, transforming growth factor-β,
insulin-like growth factor and platelet-derived growth factor,
leading to proliferation of VSMCs, as well as migration of VSMCs to
the intima, thus contributing to the formation of
atherosclerosis.
Liuwei Dihuang (LWDH) is a traditional Chinese
herbal formulation that has been used for >1,000 years in China.
According to traditional Chinese medicine, LWDH formula (LWDHF) has
the ability to tonify kidney 'Yin', and can be used clinically to
treat hypertension, diabetes and numerous diseases associated with
'Yin' deficiency in the kidney of perimenopausal and postmenopausal
women. LWDHF is comprised of six Chinese herbs: Shu Di Huang (Radix
Rehmanniae Preparata), Shan Zhu Yu (Fructus Corni), Shan Yao
(Rhizoma Dioscoreae), Ze Xie (Rhizoma Alismatis), Mu Dan Pi (Cortex
Moutan) and Fu Ling (Poria). LWDHF is orally administered as a
decoction, or in pill form, for the treatment of aging-associated
functional decline and geriatric diseases, particularly for
diabetes, cognition and memory decline in China and Japan (13,14). Recently, numerous studies have
revealed the beneficial effects of LWDHF on the cardiovascular
system in postmenopausal women. Our previous study demonstrated
that LWDHF exerted preventive and therapeutic effects on
atherosclerosis in ovariectomized rats (15,16), and that LWDHF-medicated serum
protected human umbilical vein endothelial cells (HUVECs) against
hydrogen peroxide (H2O2)-induced apoptosis
and upregulated the expression of estrogen receptors (ERs)
following vascular endothelial cell injury in vitro
(16). A significant increase in
the incidence and mortality of cardiovascular diseases has been
noted in postmenopausal women, thus indicating that estrogen may
have an important protective effect on the vasculature (17). Furthermore, numerous studies
demonstrated that estrogen could lower the risk of cardiovascular
disease in women (18) and
inhibit VSMC proliferation following injury (19-21). Therefore, the present study aimed
to investigate the effects of LWDHF on Ang II-induced VSMC
proliferation and migration, and to explore the role of ERs in the
effects of LWDHF. To the best of our knowledge, the present study
is the first to demonstrate the suppressive effects of LWDHF on Ang
II-induced proliferation and migration of VSMCs. In addition, the
molecular mechanism by which LWDHF inhibits proliferation and
migration may be associated with modulating the phenotypic
modulation of VSMCs, which was partially mediated by the
ERα-activated estrogen signaling pathway. Understanding the
cellular and molecular pathways of LWDHF may result in the
identification of novel therapeutic strategies for the treatment of
atherosclerosis and restenosis in perimenopausal or postmenopausal
women.
Materials and methods
Reagents and antibodies
Human Ang II, tamoxifen and cell proliferation
reagent 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) were purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). Antibodies used to detect the protein
expression levels of α-SMA (ab124964), OPN (ab8448), ERβ (ab92306),
β-actin and β-tubulin were obtained from Abcam (Cambridge, MA,
USA). Anti-ERα antibody was purchased from Cell Signaling
Technology, Inc.(Danvers, MA, USA). ERα small interfering (si)RNA
and control siRNA were purchased from Santa Cruz Biotechnology,
Inc. (Dallas. TX, USA).
Preparation of LWDH
The method of LWDH preparation was reported by Yang
et al (22). Briefly,
Rehmannia glutinosa Libosch. (Scrophulariaceae family),
Cornus officinalis Sieb. (Cornaceae family), Dioscorea o
pposita Thunb. (Dioscoreaceae family), Alisma orientale
(G. Samuelsson) Juz (Alismataceae family), Poria cocos
(Schw.) Wolf (Polyporaceae family) and Paeonia suffruticosa
Andrews (Paeoniaceae family) were mixed at a ratio of 8:4:4:3:3:3.
The mixture was twice decocted in distilled water for 30 min. The
water extracts were concentrated to 2 g/ml for further use.
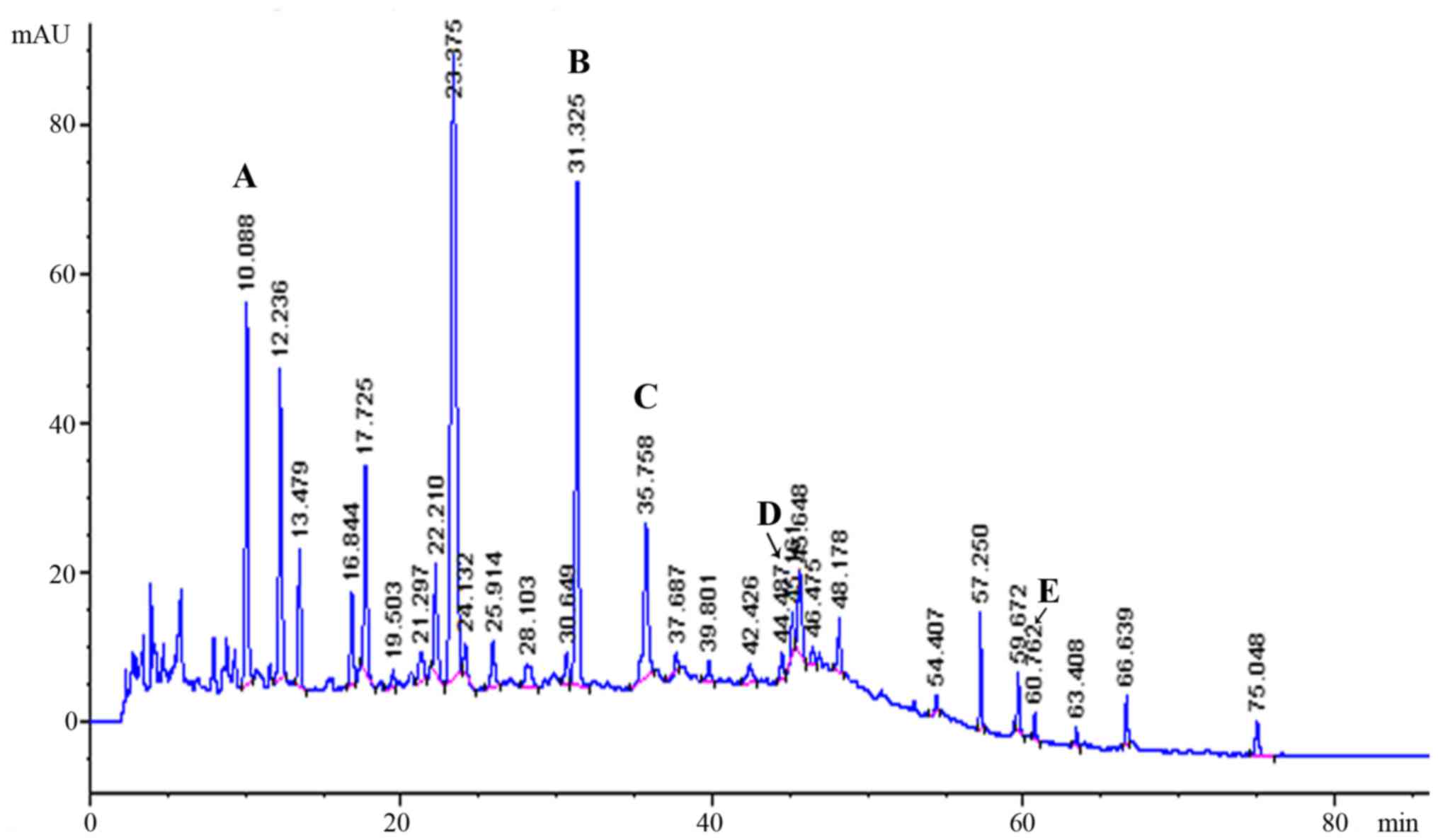
High-performance liquid chromatography was used to analyze the
constituents of LWDH (22). Five
major constituents, including gallic acid, paeonoside,
verbascoside, loganin and paeoniflorin, were identified in LWDHF
(Fig. 1). Their contents were
2.74, 0.05, 0.06, 0.74 and 0.43 mg/g, respectively. The standard
samples of gallic acid, paeonoside, verbascoside, loganin and
paeoniflorin were purchased from Beijing Beina Chuanglian
Biotechnology Research Institute (Beijing, China).
Cell culture
Primary VSMCs were isolated from thoracic aortas of
7-week-old male Sprague-Dawley rats by explant technique, and were
then cultured in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% fetal bovine serum (FBS), 100 kU/l
benzylpenicillin and 100 mg/l streptomycin at 37°C in a humidified
chamber containing 5% CO2 (23). The identification of VSMCs was
performed by α-SMA immunostaining; >90% of cells were
α-SMA-positive and exhibited a spindle-shaped appearance. VSMCs
were passaged by trypsinization, and cells at passages 3-7 were
used for subsequent experiments to ensure genetic stability of the
culture. All animal experimental protocols were approved by the
Nanjing University of Chinese Medicine Committee on Laboratory
Animal Care and all animals received humane care according to the
National Institutes of Health guidelines. The animals were housed
under diurnal lighting conditions (12:12) and had access to food
and water ad libitum. Female Sprague-Dawley rats (weighting
200±30g) were provided by Zhejiang Experimental Animal Center
(Nanjing, China).
Cell proliferation assay
Cell proliferation was analyzed using the MTT assay.
VSMCs (1×104/well) were seeded in a 96-well microplate
and were cultured with 200 µl DMEM supplemented with 10%
FBS. Once the cells had reached 60% confluence they were
serum-starved for 16 h at 37°C in a humidified chamber containing
5% CO2. VSMCs were then treated with 100 nM Ang II or
100 nM Ang II + LWDHF (3, 6 and 12 µg/ml) for 24 h; cells
were incubated with MTT (5 mg/ml) for the last 4 h at 37°C in a
humidified chamber containing 5% CO2 and then dissolved
into 150 µl DMSO. Untreated cells served as the control
group. MTT was dissolved in PBS at a concentration of 5 mg/ml.
Subsequently, optical density was measured at 490 nm using a
Universal Microplate Spectrophotometer (BioTek Instruments, Inc.,
Winooski, VT, USA). All experiments were performed in
triplicate.
Western blot analysis
Cells were lysed in radioimmunoprecipitation assay
(RIPA) buffer with a protease and phosphatase inhibitor cocktail
(protease inhibitor : phosphatase inhibitor = 100:1/1). Solubilized
protein samples were centrifuged at 12,000 x g for 15 min at 4°C,
and protein concentration within the supernatant was determined
using a Bradford protein assay. Equal amounts of protein (30-50
µg) were separated by 10% SDS-PAGE and were transferred to
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA). The membranes were then blocked with 5% non-fat milk powder
in Tris-buffered saline containing 0.1% Tween-20 (TBST) for 2 h at
room temperature, and were incubated with various primary
antibodies (all antibodies were diluted in 1X TBST) overnight at
4°C. After washing with TBST, the membranes were incubated for 90
min at room temperature with a rabbit anti-goat secondary antibody
(sc-2004; 1:5,000; Santa Cruz Biotechnology, Inc.). After washing
three times with TBST, the membranes were detected using a
chemiluminescence kit (EMD Millipore). The blots were finally
semi-quantified by densitometric analysis (Image Lab Software,
version 3.0 Beta 3). Specific protein expression levels were
normalized to β-tubulin or β-actin for total protein analyses.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from VSMCs was isolated using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). RNA concentration was quantified by measuring the absorbance
at 260 nm using a spectrophotometer, and RNA purity was assessed
according to the 260 and 280 nm ratio. RT was conducted according
to the manufacturer's protocol. RNA (5 µg) was then reverse
transcribed into cDNA using a First Strand cDNA Synthesis kit
(Thermo Fisher Scientific, Inc.). The obtained cDNA was used to
determine the mRNA expression levels of α-SMA, OPN, ERα and ERβ by
qPCR using a Thermo Fisher Scientific ABI7500 instrument (Thermo
Fisher Scientific, Inc.) with SuperScript II First-Strand Synthesis
SuperMix (Thermo Fisher Scientific, Inc.), and relative target gene
expression was normalized to GAPDH or β-tubulin. The primer
sequences were designed by Biogot Technology Co., Ltd. (Nanjing,
China) and are presented in Table
I. RT-qPCR was performed using AceQ qPCR SYBR Green Master Mix.
Amplification was initiated at 95°C for 10 min, followed by 40
cycles at 95°C for 10 sec, and was finally terminated at 60°C for
40 sec and analysed using the ΔΔCt method on an ABI 7500 Thermal
Cycler (both from Invitrogen; Thermo Fisher Scientific, Inc.). The
quantification cycle (Cq) value of the gene of interest was
obtained and was initially normalized to the housekeeping gene
GAPDH or β-tubulin (ΔCq), after which the value was further
normalized to the control (ΔΔCq). The fold-change of each target
gene was calculated using the 2−ΔΔCq method (24).
 | Table IPrimer sequences used for polymerase
chain reaction. |
Table I
Primer sequences used for polymerase
chain reaction.
| Gene | Sense primer | Antisense
primer |
|---|
| β-tubulin |
5-CGCAAGCTAGCTGTCAACAT-3 |
5-CTGCTCATCCACCTCCTTCA-3 |
| α-SMA |
5-CATCATGCGTCTGGACTTGG-3 |
5-CCAGGGAAGAAGAGGAAGCA-3 |
| OPN |
5-AGCCATGAGTCAAGTCAGCT-3 |
5-ACTCGCCTGACTGTCGATAG-3 |
| GAPDH |
5-CCTCTATGCCAACACAGTGC-3 |
5-CCTGCTTGCTGATCCACATC-3 |
| ERα |
5-TCGCTACTGTGCTGTGTGTA-3 |
5-GCCTTTCATCATGCCCACTT-3 |
| ERβ |
5-TGAGCACCTTGAGTCCAGAG-3 |
5-AGTCCCACCATTAGCACCTC-3 |
Transmission electron microscopy
The cells were fixed in 2% glutaraldehyde in neutral
phosphate buffer, post-fixed in osmium tetraoxide, and embedded in
Epon. Sections were cut at 80 nm and examined under a Philips
Tecnai 10 electron microscope.
Flow cytometry
Primary VSMCs were seeded in 6-well plates at a
density of 2×105 cells and were incubated at 37°C for 24
h. After VSMCs were treated with Ang II and/or LWDHF at 37°C for 24
h, they were trypsinized, collected and washed twice with cold PBS
at 4°C for 5 min. Precipitated cells were fixed in 1 ml ice-cold
70% ethanol overnight at 4°C. The fixed cells were then washed in
PBS, treated with RNase A (10 µg/ml), and DNA was stained
with propidium iodide (50 µg/ml) for 30 min at 37°C in the
dark. Cells (1×104) were finally analyzed by flow
cytometry (FACSCalibur; BD Biosciences, San Jose, CA, USA).
Wound-healing assay
Primary VSMCs were seeded in 6-well plates at a
density of 2×105 cells for 24 h to ensure a single cell
distribution. Confluent cells were scratched along the edge of the
attached cells to create a double-sided wound using a 10 µl
plastic pipette tip and were rinsed twice with PBS to remove cell
debris. Cells were then grown in serum-free medium supplemented
with Ang II and/or LWDHF for 24 h. Cell gaps were observed under a
Zeiss microscope (CarlZeiss, Hallbergnoos, Germany) and images were
captured of the wound sites. VSMC migration was assessed on the
basis of the injured area covered by cells counted from the wound
borders.
Transwell migration assay
The effects of LWDHF on VSMC migration were examined
by Transwell chamber assay. Primary VSMCs cells (1×105
cells/well) at passages 3–7 were seeded in a 36 mm culture dish for
24 h. Cells were then trypsinized, resuspended in 0.5% FBS medium
in the upper chamber and incubated with LWDHF (12 µg/ml),
DMEM medium containing Ang II (100 nM) was added to the lower
chamber. The cells were incubated at 37°C in an atmosphere
containing 5% CO2 for 24 h and were allowed to migrate
through the micropores to the bottom side of the Transwell
apparatus. The remaining cells in the upper chamber were removed
with a cotton swab, the cell membrane surface was wiped, and the
lower side of the filter harboring the migrated VSMCs was fixed
with 4% paraformaldehyde for 30 min. The migrating cells were then
stained with 0.5% Coomassie Brilliant Blue for 10 min. Stained
cells in five random visual fields from each of the Transwell
filters were selected and images were captured under a Zeiss
microscope (×200 magnification; CarlZeiss).
Analysis of F-actin cytoskeleton by
fluorescence microscopy
Primary VSMCs were cultured on glass coverslips
treated with indicated agents at 37°C for 24 h. Cells were then
fixed with pre-cooled 4% paraformaldehyde and rinsed three times
with PBS at room temperature. Cells were permeabilized in 0.1%
Triton X-100 and incubated with 1% bovine serum albumin
(10735108001; Biosharp, Hefei, China)/PBS to block nonspecific
binding, after which they were incubated with
phalloidin-fluorescein isothiocyanate for 60 min at room
temperature in the dark. After three washes with PBS at 4°C for 5
min, images were captured and were analyzed using the ZEN 2011
imaging software on a Zeiss inverted microscope (Carl Zeiss AG,
Oberkochen, Germany) under 400-fold magnification.
Transfection of siRNA
Human VSMCs were seeded in a 6-well culture plate at
a density of 2×105 cells in 2 ml antibiotic-free normal
growth medium supplemented with 10% FBS. The cells were incubated
at 37°C in a CO2 incubator until the cells reached 60%
confluence. Cells were then transfected with a transfection mixture
composed of ERα siRNA (sc-29305) or control siRNA (sc-37007) and
siRNA transfection reagent (sc-29528) (all Santa Cruz
Biotechnology, Inc.) according to the manufacturer's protocols.
After 6 h, cells were washed and cultured for 18 h in complete
medium, and were treated with Ang II and/or LWDHF for 24 h. The
cells were subsequently lysed and collected for western blot
analyses.
Statistical analysis
SPSS 10.0 (SPSS, Inc., Chicago, IL, USA) was used
for all statistical analyses. All data are presented as the mean ±
standard deviation of at least three independent experiments.
Differences in the results between two groups were evaluated using
either two-tailed Student's t-test or one-way ANOVA followed by
Dunnett's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
LWDHF inhibits Ang II-induced VSMC
proliferation and induces cell cycle arrest
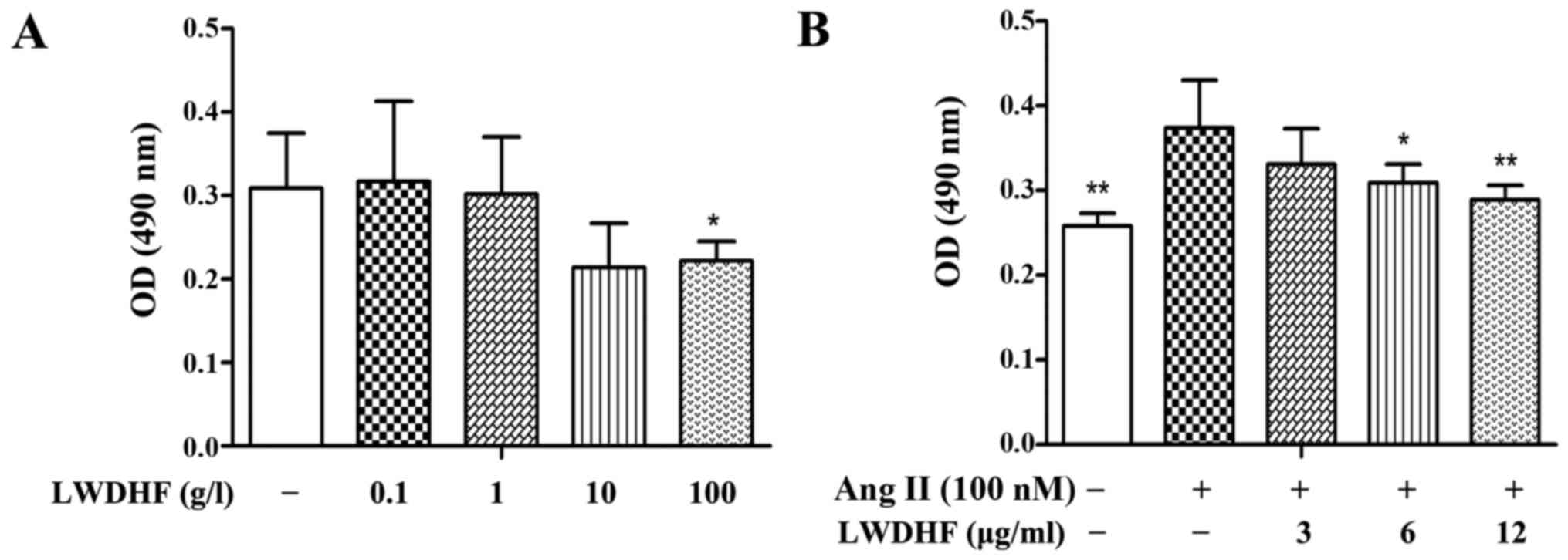
The present study evaluated the effects of LWDHF on
normal primary VSMC proliferation using the MTT assay.
Growth-arrested cells were treated with LWDHF (0.1–100 g/l) in the
presence of 10% FBS; the results indicated that no significant
difference in cell viability was observed following treatment with
≤10 g/l LWDHF (Fig. 2A),
suggesting that ≤10 g/l LWDHF did not exert significant
cytotoxicity to normal cells. However, treatment with Ang II (100
nM) induced a 1.45-fold increase in VSMC proliferation, whereas
treatment with LWDHF attenuated the effects of Ang II on VSMC
proliferation in a dose-dependent manner (Fig. 2B); treatment with the highest
concentration of LWDHF (12 µg/ml) significantly inhibited
Ang II-induced VSMC proliferation. In addition, flow cytometric
analysis was performed to observe the effects of LWDHF on cycle
progression of VSMCs. As shown in Table II, the percentage of cells at
G0/G1, S and G2/M phases in the
Ang II-stimulated group was 58.85±5.01, 21.82±2.19 and 19.34±2.86%,
respectively. LWDHF, at concentrations of 3, 6 and 12 µg/ml,
effectively increased the proportion of cells in the
G0/G1 phase and simultaneously decreased the
S and G2/M phase cell populations. These results
indicated that LWDHF may exert suppressive effects on Ang
II-induced VSMC proliferation; these effects may be associated with
the induction of cell cycle arrest at G0/G1
phase.
| Table IIEffects of LWDHF on Ang II-induced
cell cycle progression. |
Table II
Effects of LWDHF on Ang II-induced
cell cycle progression.
| Group | Dose | Phases
|
|---|
|
G0/G1 | S |
G2/M |
|---|
| Control | | 70.81±0.39a | 16.24±0.53a | 12.96±0.44a |
| Ang II | 100 nM | 58.85±5.01 | 21.82±2.19 | 19.34±2.86 |
| LWDHF | 3 µg/ml | 67.01±0.27b | 19.20±1.03 | 13.79±0.77a |
| 6 µg/ml | 67.90±0.47b | 18.27±0.59b | 13.84±0.37a |
| 12
µg/ml | 68.73±0.29a | 17.68±0.42b | 13.59±0.61a |
LWDHF suppresses Ang II-stimulated VSMC
migration
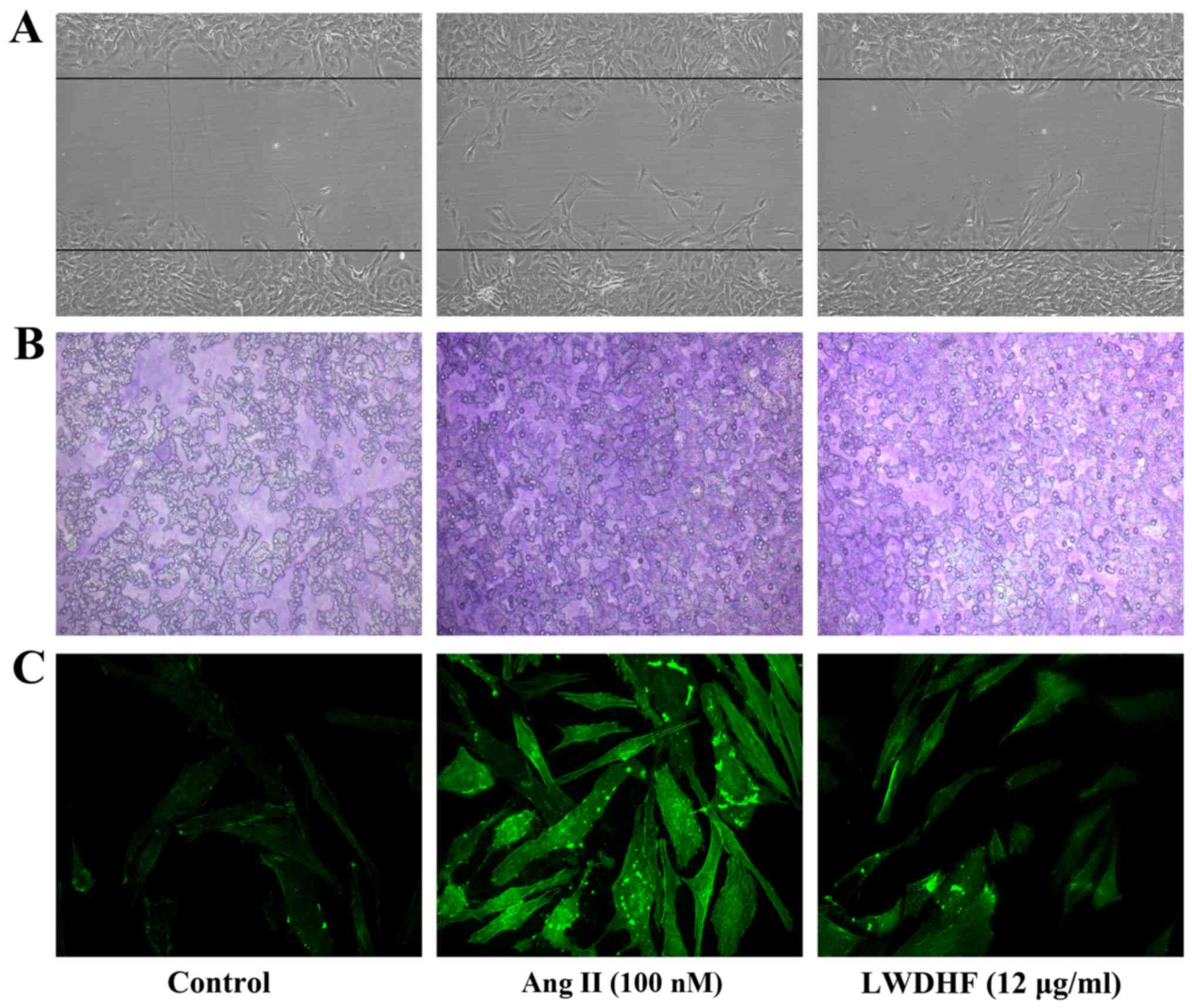
The effects of LWDHF on Ang II-stimulated VSMC
migration were observed using a wound-healing assay. Equal numbers
of confluent VSMCs were scratched to create a double-sided wound
using a 10 µl pipette tip. The cells were then treated with
Ang II (100 nM) or Ang II (100 nM) + LWDHF (12 µg/ml). As
presented in Fig. 3A, Ang II
promoted VSMCs to migrate into the wound, whereas cell migration
was reduced in the LWDHF-treated group compared with in the Ang
II-stimulated group, thus suggesting that LWDHF may reduce VSMC
migration. In addition, a Transwell chamber assay was conducted to
verify the effects of LWDHF on Ang II-induced VSMC migration. As
shown in Fig. 3B, LWDHF treatment
markedly decreased migration of VSMCs to the bottom chamber in
response to Ang II compared with those treated with Ang II alone.
Taken together, these results indicated that LWDHF may reduce Ang
II-induced VSMC migration.
Effects of LWDHF on Ang II-induced
F-actin organization in VSMCs
The present study demonstrated that LWDHF inhibited
the migration of Ang II-stimulated VSMCs. Therefore, in order to
determine whether LWDHF decreased cell migration by influencing
reorganization of the cytoskeleton, F-actin was visualized by
phalloidin staining, 24 h after Ang II (100 nM)-stimulated VSMC
spreading. Treatment with Ang II resulted in a substantial increase
in the number of stress fibers, as well as the rearrangement of
these structures into ordered parallel arrays, in cultured VSMCs.
Conversely, treatment with LWDHF (12 µg/ml) abolished
reorganization of the actin cytoskeleton induced by Ang II
(Fig. 3C).
LWDHF inhibits Ang II-induced VSMC
phenotypic switching
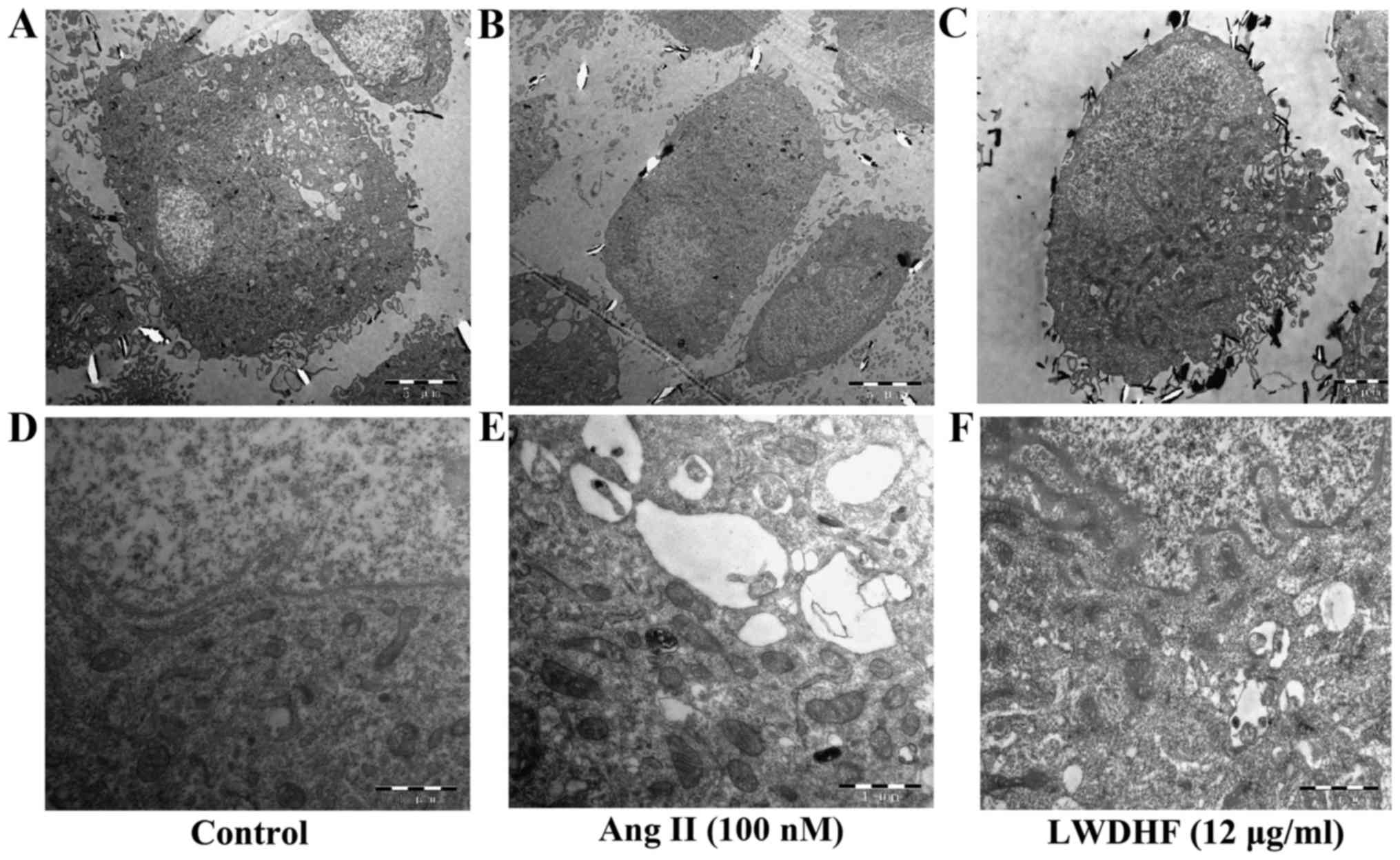
VSMCs transform from a differentiated contractile
phenotype into a synthetic phenotype, which is associated with low
expression of contractile proteins and high expression of rough
endoplasmic reticulum (RER) and ECM proteins (25). The ultrastructure of VSMCs was
observed under transmission electron microscopy (Fig. 4). The cytoplasm in normal VSMCs
was filled with myofilaments, while homogeneous distribution of
chromatin in nucleus. In cells treated with Ang II for 24 h, the
the muscle filament in the cytoplasm obviously decreased compared
to that in the control group, but the Golgi bodyobviously
increased, and the endoplasmic reticulum was obviously dilated.
These ultrastructural alterations are characteristics of the
synthetic phenotype, thus suggesting that Ang II induced phenotypic
modulation of VSMCs. However, after treatment with LWDHF (12
µg/ml), the myocytes in the cytoplasm were still rich, but
the organelles of the Golgi bodies were decreased. These
ultrastructural observations indicated that LWDHF reversed Ang
II-induced VSMC phenotypic switching. Furthermore, the present
study measured alterations in the expression of molecular markers
associated with contractile and synthetic VSMC phenotypes. RT-qPCR
and western blot analysis were used to measure the relative mRNA
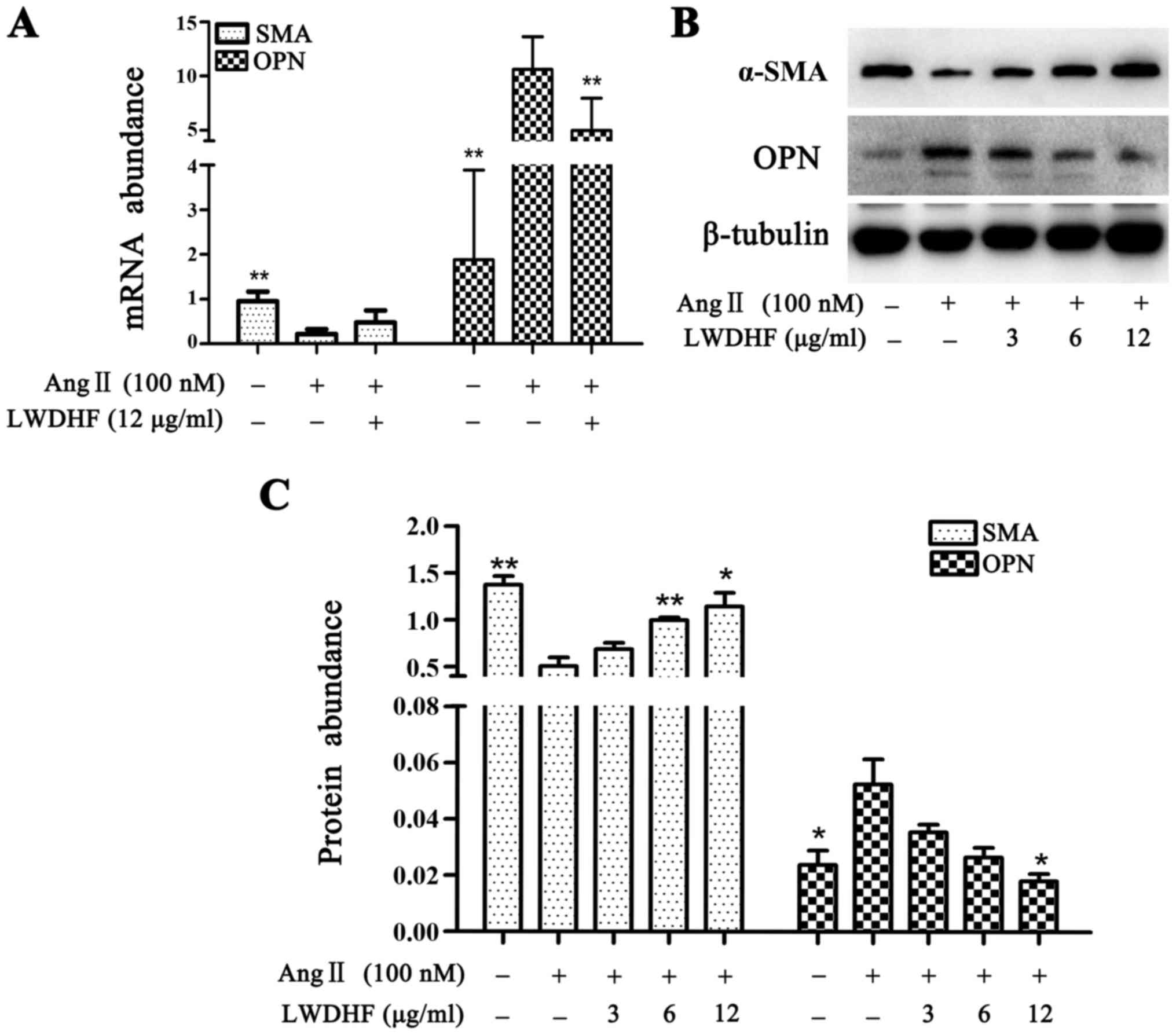
and protein abundance of α-SMA and OPN, respectively. The results
indicated that exposure of VSMCs to Ang II for 24 h resulted in a
75% decrease in the mRNA expression levels of α-SMA and a robust
upregulation of OPN (5-fold) (Fig.
5A). In addition, a decline in the protein expression levels of
α-SMA and an increase in OPN protein expression were determined by
western blotting (Fig. 5B and C).
Conversely, addition of LWDHF dose-dependently rescued the
downregulation of α-SMA and abolished the upregulation of OPN in
Ang II-treated VSMCs. These results confirmed that the
aforementioned ultrastructural alterations were due to
LWDHF-induced inhibition of VSMC phenotypic transition caused by
Ang II, and indicated that LWDHF suppressed VSMC proliferation and
migration via inhibiting VSMC phenotypic switching.
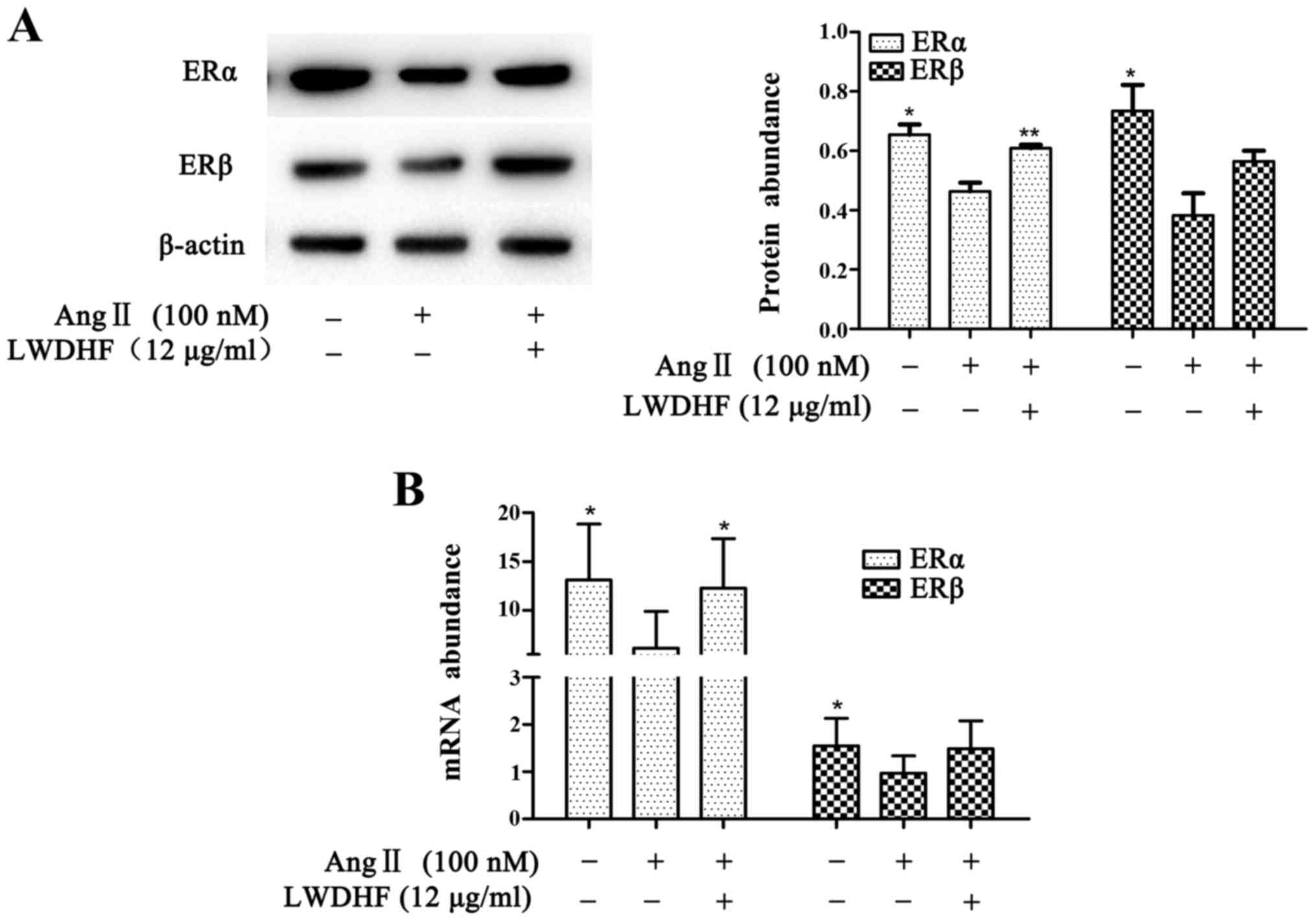
LWDHF regulates expression of ERs in
VSMCs
Our previous studies demonstrated that LWDHF exerts
estrogen-like effects on the prevention of cardiovascular diseases
in experimental rat models (15,16). The present study investigated
whether LWDHF could affect the expression of ERs in VSMCs. As shown
in Fig. 6, the mRNA expression
levels of ERα and ERβ were significantly downregulated in VSMCs
stimulated with Ang II (100 nM) compared with in untreated cells.
Conversely, VSMCs treated with LWDHF and exposed to Ang II for 24 h
exhibited a significant upregulation of ERα; in addition ERβ
exhibited higher expression in LWDHF-treated cells compared with in
cells treated with Ang II alone. The results of western blotting
were consistent with those of RT-qPCR.
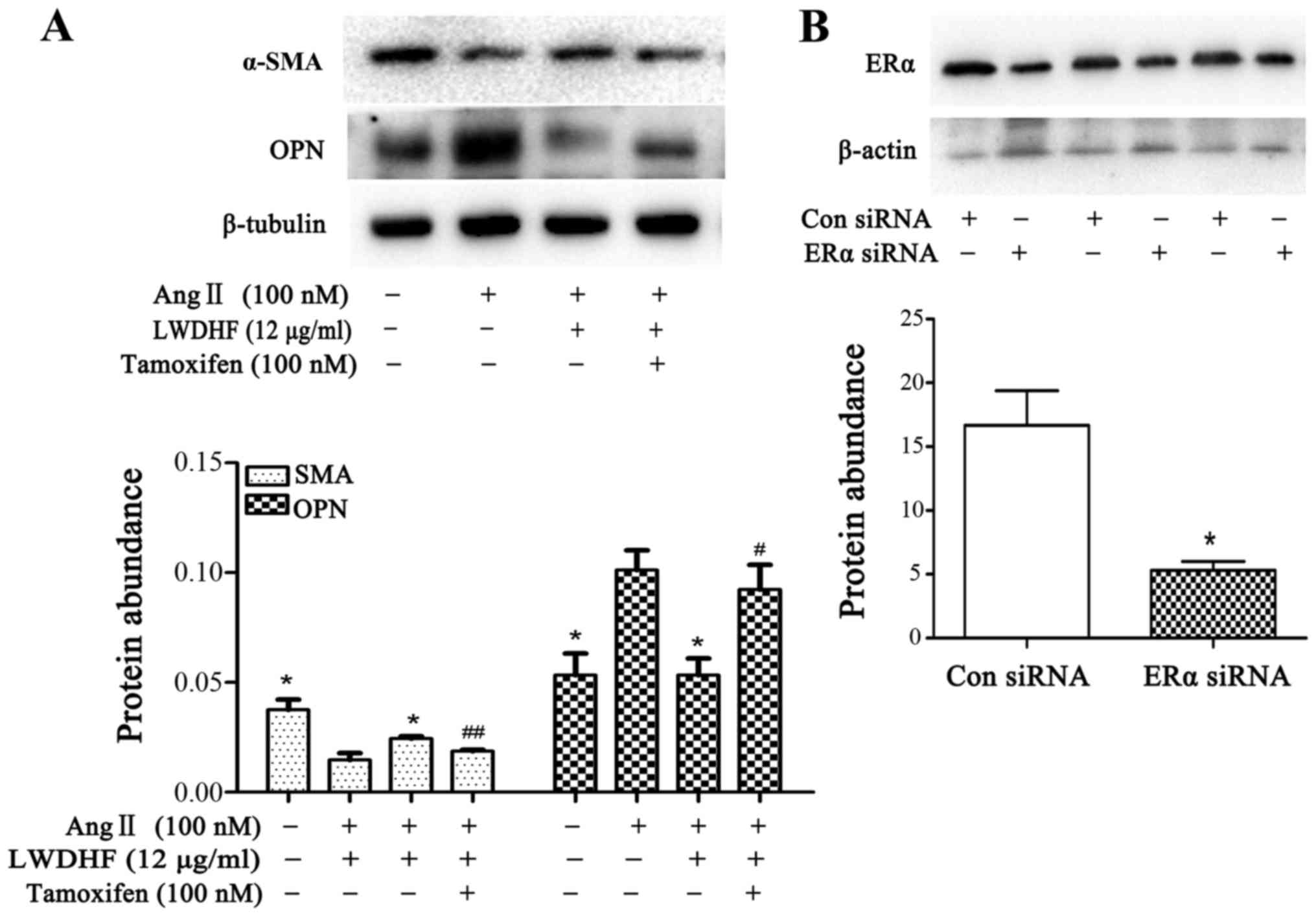
Role of ERs in Ang II-induced phenotypic
modulation of VSMCs
To examine whether ERs were involved in the
inhibitory effects of LWDHF on the phenotypic modulation of VSMCs,
tamoxifen, a nonselective estrogen antagonist, was used to
characterize the role of ER in LWDHF-mediated effects. As shown in
Fig. 7A, part suppression of ER
signaling by tamoxifen markedly attenuated the inhibitory effects
of LWDHF on VSMC phenotypic switching; the ability of LWDHF to
increase α-SMA protein expression and decrease OPN protein
expression was significantly abrogated by tamoxifen. Furthermore,
human aortic VSMCs (HAVSMCs) were transfected with ERα siRNA to
observe the role of ERα in LWDHF-mediated inhibition of VSMC
phenotypic switching. When 0.24 nM ERα siRNA was transfected into
HAVSMCs, the constitutive expression of ERα was significantly
reduced after 6 h (Fig. 7B),
indicating that HAVSMCs were successfully transfected with this
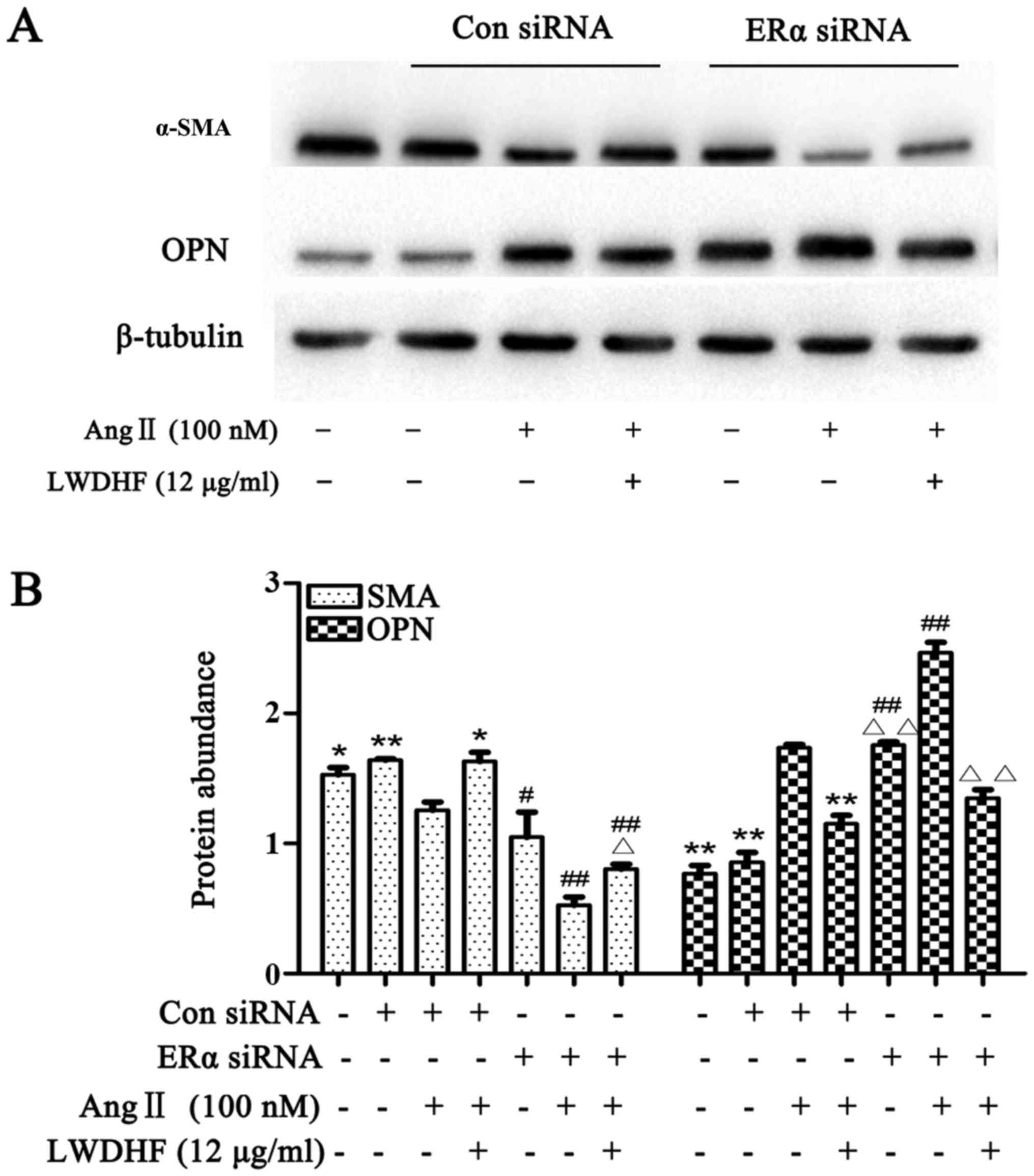
siRNA. HAVSMCs were transfected with ERα siRNA for 6 h and were
then stimulated with Ang II (100 nM) for 24 h; these cells
exhibited a significant reduction in α-SMA expression and a
significant increase in OPN expression compared with in cells
transfected with control siRNA and treated with Ang II. In
addition, HAVSMCs transfected with ERα siRNA and stimulated with
Ang II in the presence of LWDHF for 24 h exhibited weakened
upregulated SMA expression, while the expression of OPN was not
downregulated. The results showed that the expression of SMA and
OPN was related to ERα, and the change of SMA and OPN expression
were mediated by ERα (Fig. 8).
Taken together, these results indicated that ERα knockdown
abolished the inhibitory effects of LWDHF on Ang II-induced VSMC
phenotypic switching, and ERα may be involved in LWDHF-mediated
upregulation of α-SMA.
Discussion
Our previous study investigated the effects of LWDHF
on ovariectomized rats with atherosclerosis and demonstrated that
LWDHF improved lipid metabolism in the serum and reduced Ang II
levels through regulating ER expression, thereby producing
therapeutic effects (15). In
addition, our in vitro study demonstrated that
LWDHF-medicated serum protected HUVECs against
H2O2-induced apoptosis via ERα (16). Therefore, it may be hypothesized
that LWDHF exerts preventive and therapeutic effects against
atherosclerosis in menopausal women, and that ERs may mediate the
effects of LWDHF. The present study provided further evidence
supporting the antiatherosclerotic effects of LWDHF, indicating
that LWDHF inhibited the Ang II-induced phenotypic modulation and
migration of VSMCs, and that the effects of LWDHF were associated
with ERs, particularly ERα.
The development of atherosclerosis- and
restenosis-associated advanced lesions is highly dependent on VSMC
proliferation. Therefore, inhibition of VSMC proliferation is
considered a potential strategy in the prevention of
atherosclerosis (7). LWDH is a
commonly used traditional Chinese formula; the present study
demonstrated that LWDH could inhibit Ang II-induced VSMC
proliferation in a dose-dependent manner, and the maximum
inhibition on proliferation was detected following treatment with
12 µg/ml LWDH (73.24%). It is well known that progression
from G1 to S phase is closely associated with cell
proliferation. The present study provided further evidence to
confirm that LWDHF may exert antiproliferative effects; Ang
II-induced cell cycle progression to S phase was markedly reduced
by 12 µg/ml LWDHF, as determined by flow cytometry.
Migration also has an important role in numerous vascular diseases.
In the present study, LWDHF exerted marked inhibitory effects on
VSMC migration, as determined using wound-healing and Transwell
chamber assays. Cell migration is associated with numerous
spatially and temporally coordinated cellular processes, and occurs
in the following four steps: i) Formation of actin-rich
protrusions, such as lamellipodia, ii) cell adhesion, iii)
translocation of the cell body and iv) rear detachment.
Cytoskeletal reorganization is required for leading-edge cellular
protrusion during locomotion, and VSMC migration requires
rearrangement in the VSMC cytoskeleton (26,27). Consistent with previous studies,
the present study demonstrated that treatment with LWDHF could
markedly suppress Ang II-stimulated rearrangement of F-actin.
Phenotypic modulation of VSMCs is the cytological
basis for vascular remodeling. Under normal conditions, VSMCs with
a contractile phenotype are highly specialized cells, which
function to regulate blood vessel diameter, blood pressure and
blood flow distribution. However, VSMCs exhibit a phenotypic
alteration, characterized by loss of contractility, and abnormal
proliferation and migration, in response to vascular injury or
disease. The present study hypothesized that LWDHF may inhibit
proliferation and migration of VSMCs via modulating the VSMC
phenotype. In general, α-SMA is considered a biomarker of VSMCs
with a contractile phenotype, whereas OPN is a biomarker of VSMCs
with a synthetic phenotype (28,29). VSMC phenotypic switching is
characterized by markedly increased expression of synthetic
phenotype markers, alongside increased VSMC proliferation and
migration. Consistent with the previous observations, treatment of
VSMCs with Ang II resulted in downregulation of α-SMA and
upregulation of OPN; these effects were suppressed by LWDHF in a
concentration-dependent manner. These results confirmed the
hypothesis that LWDHF may inhibit proliferation and migration of
VSMCs via modulating the VSMC phenotype. However, the addition of
tamoxifen to the culture medium partly abolished the inhibitory
effects of LWDHF on phenotypic switching, thus suggesting that ERs
may be involved in the effects of LWDHF on VSMC phenotype.
A previous study demonstrated that LWDHF may possess
estrogenic properties and regulate the expression of ERs (16). The present study demonstrated that
LWHDF could suppress VSMC proliferation in vitro. To further
investigate the association between LWHDF and ER, western blot
analysis and RT-qPCR were employed to examine the effects of LWDHF
on ERα and ERβ expression. The results indicated that LWDHF
upregulated the expression levels of ERs, particularly ERα. The
underlying mechanism of inhibitory actions with LWDHF on VSMCs has
been intensely investigated following the discovery of ERα. Our
earlier studies in HUVECs indicated that LWDHF exerted
anti-apoptotic effects, mainly via an ERα-mediated estrogen
signaling pathway (16).
Therefore, to determine whether ERα mediated the inhibitory effects
of LWDHF on phenotypic modulation of VSMCs, siRNA technology was
used to reduce the expression of ERα in HUVSMCs, which led to a
significant increase in the Ang II-induced synthetic phenotype. In
addition, ERα knockdown in HUVSMCs significantly attenuated the
effects of LWDHF on phenotypic modulation. LWDHF-elevated α-SMA
expression was markedly reduced in HUVSMCs treated with ERα siRNA,
whereas OPN expression was not markedly altered in ERα
siRNA-transfected cells treated with Ang II and LWDHF compared with
in control siRNA-transfected cells. These data suggested that LWDHF
may upregulate α-SMA expression predominantly via ERα-mediated
signaling pathways; however, another ER subtype may be responsible
for LWDHF-induced downregulation of OPN.
In conclusion, the present study suggested that
LWDHF possesses estrogenic properties and may mimic E2 to suppress
VSMC proliferation and migration in vitro. The effects
exerted by LWDHF were associated with phenotypic regulation. In
addition, analysis of the underlying mechanisms of action revealed
that ERα may act as a key mediator for LWDHF-induced inhibition of
phenotypic switching. However, whether ERβ is involved in the
inhibitory effects of LWDHF on phenotypic switching and migration
of VSMCs requires further research.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81774029).
References
|
1
|
Rivard A and Andrés V: Vascular smooth
muscle cell proliferation in the pathogenesis of atherosclerotic
cardiovascular diseases. Histol Histopathol. 15:557–571.
2000.PubMed/NCBI
|
|
2
|
Lacolley P, Regnault V, Nicoletti A, Li Z
and Michel JB: The vascular smooth muscle cell in arterial
pathology: a cell that can take on multiple roles. Cardiovasc Res.
95:194–204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Deatrick KB, Eliason JL, Lynch EM, Moore
AJ, Dewyer NA, Varma MR, Pearce CG, Upchurch GR Jr, Wakefield TW
and Henke PK: Vein wall remodeling after deep vein thrombosis
involves matrix metalloproteinases and late fibrosis in a mouse
model. J Vasc Surg. 42:140–148. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Owens GK, Kumar MS and Wamhoff BR:
Molecular regulation of vascular smooth muscle cell differentiation
in development and disease. Physiol Rev. 84:767–801. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rensen SS, Doevendans PA and van Eys GJ:
Regulation and characteristics of vascular smooth muscle cell
phenotypic diversity. Neth Heart J. 15:100–108. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schwartz SM: Perspectives series: cell
adhesion in vascular biology. Smooth muscle migration in
atherosclerosis and restenosis. J Clin Invest. 99:2814–2816. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Doran AC, Meller N and McNamara CA: Role
of smooth muscle cells in the initiation and early progression of
atherosclerosis. Arterioscler Thromb Vasc Biol. 28:812–819. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hao H, Gabbiani G and Bochaton-Piallat ML:
Arterial smooth muscle cell heterogeneity: implications for
atherosclerosis and restenosis development. Arterioscler Thromb
Vasc Biol. 23:1510–1520. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Babapulle MN and Eisenberg MJ: Coated
stents for the prevention of restenosis: Part I. Circulation.
106:2734–2740. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao Y, Liu YX, Xie SL, Deng BQ, Wang JF
and Nie RQ: Increased expression of granulocyte colony-stimulating
factor mediates mesenchymal stem cells recruitment after vascular
injury. Chin Med J (Engl). 124:4286–4292. 2011.
|
|
11
|
Watanabe T, Pakala R, Katagiri T and
Benedict CR: Synergistic effect of urotensin II with mildly
oxidized LDL on DNA synthesis in vascular smooth muscle cells.
Circulation. 104:16–18. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Intengan HD and Schiffrin EL: Vascular
remodeling in hypertension: roles of apoptosis, inflammation, and
fibrosis. Hypertension. 38:581–587. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu JP, Feng L, Zhang MH, Ma DY, Wang SY,
Gu J, Fu Q, Qu R and Ma SP: Neuroprotective effect of Liuwei
Dihuang decoction on cognition deficits of diabetic encephalopathy
in streptozotocin-induced diabetic rat. J Ethnopharmacol.
150:371–381. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kalu DN: The ovariectomized rat model of
postmenopausal bone loss. Bone Miner. 15:175–191. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu Y, L Y and Zhang QC: The effect of
LWDHF on menopausal atherosclerosis model in ovariectomized rats.
Chinese Traditional Patent Medicine. 34:553–556. 2012.In
Chinese.
|
|
16
|
Yin QY, Guo J and Meng QH: Effects of
Liuweidihuang formula mediated serum on H2O2
-injured human umbilical vascular endothelial cells. Chin Pharmacol
Bull. 29:1753–1757. 2013.In Chinese.
|
|
17
|
Mendelsohn ME and Karas RH: Molecular and
cellular basis of cardiovascular gender differences. Science.
308:1583–1587. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Farhat MY, Lavigne MC and Ramwell PW: The
vascular protective effects of estrogen. FASEB J. 10:615–624. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kawagoe J, Ohmichi M, Tsutsumi S, Ohta T,
Takahashi K and Kurachi H: Mechanism of the divergent effects of
estrogen on the cell proliferation of human umbilical endothelial
versus aortic smooth muscle cells. Endocrinology. 148:6092–6099.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sivritas D, Becher MU, Ebrahimian T, Arfa
O, Rapp S, Bohner A, Mueller CF, Umemura T, Wassmann S, Nickenig G,
et al: Antiproliferative effect of estrogen in vascular smooth
muscle cells is mediated by Kruppel-like factor-4 and manganese
superoxide dismutase. Basic Res Cardiol. 106:563–575. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu HM, Zhao XF, Guo LN, Tan Z and Wang
TH: Effects of caveolin-1 on the 17β-estradiol-mediated inhibition
of VSMC proliferation induced by vascular injury. Life Sci.
80:800–812. 2007. View Article : Google Scholar
|
|
22
|
Yang S, Zhou W, Zhang Y, Yan C and Zhao Y:
Effects of Liuwei Dihuang decoction on ion channels and synaptic
transmission in cultured hippocampal neuron of rat. J
Ethnopharmacol. 106:166–172. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rodríguez A, Fortuño A, Gómez-Ambrosi J,
Zalba G, Díez J and Frühbeck G: The inhibitory effect of leptin on
angiotensin II-induced vasoconstriction in vascular smooth muscle
cells is mediated via a nitric oxide-dependent mechanism.
Endocrinology. 148:324–331. 2007. View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)). Method Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Hedin U and Thyberg J: Plasma fibronectin
promotes modulation of arterial smooth-muscle cells from
contractile to synthetic phenotype. Differentiation. 33:239–246.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rousseau S, Houle F, Landry J and Huot J:
p38 MAP kinase activation by vascular endothelial growth factor
mediates actin reorganization and cell migration in human
endothelial cells. Oncogene. 15:2169–2177. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gunst SJ and Zhang W: Actin cytoskeletal
dynamics in smooth muscle: a new paradigm for the regulation of
smooth muscle contraction. Am J Physiol Cell Physiol.
295:C576–C587. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shanahan CM, Weissberg PL and Metcalfe JC:
Isolation of gene markers of differentiated and proliferating
vascular smooth muscle cells. Circ Res. 73:193–204. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hultgårdh-Nilsson A, Lövdahl C, Blomgren
K, Kallin B and Thyberg J: Expression of phenotype-and
proliferation-related genes in rat aortic smooth muscle cells in
primary culture. Cardiovascular research. 34:418–430. 1997.
View Article : Google Scholar
|