Introduction
Age-related macular degeneration (AMD) is an
age-associated macular disease, and the most common disease of
blindness in people >60 years old (1). Its main feature is the retinopathy
of the retina and choroid, which causes decreased visual function
and reduced central vision in particular (2). Oxidative stress serves a vital role
in the pathogenesis of AMD (3).
Retinal pigment epithelial (RPE) cells, as the most metabolically
active type of cellin eye tissue, can engulf the outer disc of the
retina photoreceptor cells and produce a large number of lipid
peroxides and H2O2 (4). Furthermore, the photo-oxidation
effect occurs when RPE is illuminated over long durations (5). Therefore, RPE has a higher
susceptibility to oxidative stress. In addition, due to aging, the
resistance of the antioxidant system of RRE declines (6,7).
Oxidative stress and decreased antioxidant capacity may lead to
functional disorders and structural abnormalities of the RPE, which
have been identified as important pathological alterations
associated with AMD (3,6,7).
Genipin (GP), is a glycosidic ligand derived from
iridoid glycosides and is widely distributed in plants, including
Mast and Eucommia ulmoides. GP is the main metabolite of
geniposide in humans or animals, and is also the main active form
with pharmacokinetic function (8). Studies have demonstrated that GP has
certain properties, including anti-infection, anti-inflammation,
antioxidation and antitumor (9-12).
In addition, GP has been widely regarded as a specific inhibitor of
uncoupling protein 2 (UCP2) (13). UCP2 is a functional protein in the
mitochondrial inner membrane, which regulates the proton pump of
mitochondria (14). Specifically,
UCP2 is involved in modulating the opening of the ion channels on
the mitochondrial membrane, inhibiting the production of reactive
oxygen species (ROS), there by suppressing the apoptosis of cells
and damage to mitochondria (15).
However, the role of GP on RPE cell injury induced by oxidative
stress is unknown.
Nuclear factor-erythroid 2-related factor-2 (Nrf2),
as a transcription factor, serves a vital function in opposing cell
damage due to endogenous and exogenous stresses (16). It is well known that Nrf2 is a
main regulator of the antioxidant reaction, which can control the
antioxidant response element-regulated the expression of phase II
and antioxidant enzymes, including heme oxygenase-1 (HO-1) and
NAD(P)H: Quinine oxidoreductase 1 (NQO1) (17,18). It has been reported that
antioxidants protect oxidative stress-induced RPE cells by
activating Nrf2 signaling (19).
The present study, aimed to establish an RPE cell
oxidative stress injury model; H2O2 acts as
an inducer of oxidative damage to RPE cells (20). In addition, the effects of GP on
H2O2-induced RPE cells were determined and
whether the underlying molecular mechanism is associated with Nrf2
signaling was investigated in the present study.
Materials and methods
Cell culture and transfection
Human RPE cell lines (ARPE-19 cells) were obtained
from the American Type Culture Collection (Manassas, VA, USA),
which were maintained in Dulbecco’s modified Eagle’s
medium/Nutrient F12 Ham (DMEM/F12; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), which contained 10% fetal bovine serum (FBS;
HyClone; GE Healthcare Life Sciences, Logan, UT, USA), 100
µg/ml streptomycin and 100 U/ml penicillin (Beijing Solarbio
Science & Technology, Co., Ltd., Beijing, China) in a
humidified incubator with 5% CO2 at 37°C (80G-2,
Shanghai Huafu Instrument Co. Ltd.).
ARPE-19 cells were transfected with pSilencer 2.1
vector (1 µg, Thermo Fisher Scientific, Inc., Waltham, MA,
USA) containing small interfering (si)RNA negative control (NC;
5′-CAC ACT GGA TGG CCT AGG AGG ATA T-3′) or siRNA Nrf2
(siNrf2;5′-CAC ACT GGA TCA GAC AGG AGG ATA T-3′) vectors using
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.).
After transfection, cells were incubated for 24 h and then used for
subsequent experimentation; untreated cells served as the
control.
Methylthiazolyldiphenyl-tetrazolium
bromide (MTT) assay
An MTT kit (Beyotime Institute of Biotechnology,
Shanghai, China) was employed to measure cell viability. ARPE-19
cells were seeded into 96-well plates (2×103 cell/well)
for 24 h. Cells were exposed to various concentrations of
H2O2 (0, 10, 50, 100, 200, 400 and 800
µM) and GP (0, 5, 10, 30, 50 and 100 µM) for 24 h at
37°C, respectively. Other cells were transfected with NC and siNrf2
vectors as aforementioned. Then, cells were incubated with MTT
solution for 4 h at 37°C. Subsequently, the supernatant was
removed; dimethyl sulfoxide was then added to the cells and the
optical density at 490 nm was evaluated using a microplate reader
(SpectraMax iD3, Molecular Devices, LLC, Sunnyvale, CA, USA); 200
µM H2O2 was finally used to treat
cells in the subsequent experiments.
Flow cytometry
For the analysis of ROS, a ROS assay kit (Beyotime
Institute of Biotechnology) was employed according to the
manufacturer’s protocols. In brief, cells were seeded into 6-well
plates (5×104 cell/well) for 24 h. Then, cells were
treated with 200 µM H2O2, 30 µM
GP, NC vector or siNrf2 vector for 24 h as aforementioned.
Following treatment, cells were incubated with
2′,7′-dichlorofluorescein diacetate (Sigma-Aldrich; Merck KGaA) at
37°C for 30 min and then washed with PBS for three times. The
levels of ROS were determined using a MoFlo flow cytometer (Beckman
Coulter, Inc., Brea, CA, USA) and the data was analyzed using
SUMMIT Software V4.3 (Dako; Agilent Technologies, Santa Clara,
USA).
For the cell apoptosis assay, an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis
detection kit (Dalian Meilunbio Biotechnology Co., Ltd., Dalian,
China) was employed according to the manufacturer’s protocols.
Briefly, cells were seeded into 6-well plates (5×104
cell/well) for 24 h. Then, cells were treated with 200 µM
H2O2, 30 µM GP, NC vector and siNrf2
vector for 24 h as aforementioned. Following treatment, cells were
digested with 0.25% EDTA-trypsin (Beijing Solarbio Science &
Technology, Co., Ltd.) at room temperature for 2 min and
resuspended in DMEM/F12. The cells were centrifuged at 1,000 × g
for 5 min at 4°C, and then incubated with Annexin V-FITC and PI in
darkness for 25 min at room temperature. Cell apoptosis was
analyzed with a MoFlo flow cytometer (Beckman Coulter, Inc.) and
the data was analyzed using SUMMIT Software V4.3. The number of
apoptotic cells was calculated by adding data in the second and the
fourth quadrants of the flow cytometry data.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
Total RNA was extracted from cells using Trizol
reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer’s protocols. RNA (1 µg) was used to synthesize
cDNA by using the Quantscript RT Kit (Promega Corporation, Madison,
WI, USA) under the following conditions: 25°C for 10 min, 42°C for
50 min, 85°C for 15 min and 4°C for 10 min. cDNA was amplified
using a SYBR Premix Taq™ II kit (Takara Bio, Inc., Otsu, Japan) on
an ABI 7500 thermocycler (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The reaction conditions were as follows: 95°C
for 3 min, followed by 30 cycles of at 95°C for 15 sec and at 62°C
for 30 sec, and extension for 60 sec at 72°C. The sequences of
primers were presented in Table
I. GAPDH was used as an internal control. The quantification of
gene expression was performed using the 2−ΔΔCq method
(21).
 | Table ISequences of the primers employed for
reverse transcription-quantitative polymerase chain reaction. |
Table I
Sequences of the primers employed for
reverse transcription-quantitative polymerase chain reaction.
| Primer | Sequence
(5′-3′) |
|---|
| HO-1-forward |
CGTTCCTGCTCAACATCCAG |
| HO-1-reverse |
TGAGTGTAAGGACCCATCGG |
| NQO1-forward |
AGAAAGGATGGGAGGTGGTG |
| NQO1-reverse |
ATATCACAAGGTCTGCGGCT |
| Bax-forward |
AACATGGAGCTGCAGAGGAT |
| Bax-reverse |
CCAATGTCCAGCCCATGATG |
| Bcl-2-forward |
TTCTTTGAGTTCGGTGGGGT |
| Bcl-2-reverse |
CTTCAGAGACAGCCAGGAGA |
| GAPDH-forward |
CCATCTTCCAGGAGCGAGAT |
| GAPDH-reverse |
TGCTGATGATCTTGAGGCTG |
Western blot analysis
Cells were lysed with radioimmunoprecipitation assay
buffer (Beijing Solarbio Science & Technology, Co., Ltd.) to
obtain protein extracts. A Bradford’s protein assay kit (Beyotime
Institute of Biotechnology) was employed to detect the
concentrations of protein extracts. Each protein (25 µg per
lane) was separated by 10% SDS-PAGE and transferred to a
polyvinylidene difluoride membrane (Merck KGaA). The membrane was
blocked using tris-buffered saline with TBS solution (0.05%
Tween-20) containing 5% non-fat milk for 1 h at 4°C. The membrane
was incubated with anti-HO-1 (AF3169, 1:600; R&D Systems, Inc.,
Minneapolis, MN, USA), anti-Nrf2 (MAB3925, 1:800; R&D Systems,
Inc.), anti-NQO1 (AF7567, 1:1,200; R&D Systems, Inc.),
anti-cleaved-caspase-3 (AF835, 1:1,000; R&D Systems, Inc.),
anti-B-cell lymphoma-2 (Bcl-2; AF810, 1:800; R&D Systems,
Inc.), anti-Bcl-2 associated X (Bax; AF820, 1:1,000; R&D
Systems, Inc.) and anti-GAPDH (2275-PC-100, 1:600; R&D Systems,
Inc.) at 4°C overnight. Then, the membrane was washed with TBST for
three times. Following washing, the membrane was incubated with
corresponding secondary antibodies [rabbit anti-goat
IgG-horseradish peroxidase (HRP), sc-2768, 1:6,000; mouse
anti-rabbit, IgG-HRP, sc-2357, 1:7,000; donkey anti-goat IgG-HRP,
sc-2020, 1:8,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
at 37°C for 60 min. The membrane was washed with TBST for three
times. Subsequently, the proteins were detected by using an
enhanced chemiluminescence detection reagent (GE Healthcare,
Chicago, IL, USA)and exposed under an E-Gel Imager (Invitrogen;
Thermo, Fisher, Scientific, Inc.). The blot density was analyzed by
Quantity One software version 4.6.2 (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Statistical analysis
The data were presented as the mean ± standard
deviation using SPSS software (version 20; IBM, Corp., Armonk, NY,
USA). The differences among groups were assessed by one-way
analysis of variance followed by a Tukey’s post hoc test. All the
experiment was independently conducted at least for three times.
P<0.05 was considered to indicate a statistically significant
difference.
Results
GP alleviates decreased viability of
ARPE-19 cells induced by H2O2
To investigate the effects of GP and
H2O2 on the viability of ARPE-19 cells, an
MTT assay was conducted to determine cell viability. The results
revealed that cell viability was significantly suppressed in
response to treatment with 200, 400 and 800 µM
H2O2 compared with the control (Fig. 1A), while 30, 50 and 100 µM
GP significantly promoted cell viability compared with the control;
the increased cell viability at 30 µM GP was similar to that
of 50 and 100 µM (Fig.
1B). Therefore, 100 and 200 µM
H2O2 and 5, 10, 30 µM GP was selected
to further detect the effects of GP on
H2O2-treated cells. Upon treatment with
H2O2 (10, 50 and 100 µM) or GP (5 and
10 µM), no notable alterations in cell viability were
observed. Treatment with 200 µM H2O2
and GP (0, 5, 10 and 30 µM) exhibited significantly reduced
cell viability compared with the untreated control (Fig. 1C). No significant difference was
observed between the GP + 200 µM H2O2
and GP + 100 µM H2O2 groups. Thus, 30
µM GP was used to treat cells for subsequent experiments.
The present study reported that cell viability was significantly
increased in the 30 µM GP + 200 µM
H2O2 group compared with the 200 µM
H2O2 group (Fig. 1C). The results indicated that GP
increased the viability of H2O2-treated
cells.
 | Figure 1GP alleviates decreased viability of
ARPE-19 cells induced by H2O2. (A-C) ARPE-19
cells were exposed to various concentrations of
H2O2 (0, 10, 50, 100, 200, 400+ and 800
µM) and GP (0, 5, 10, 30, 50 and 100 µM). Cell
viability was assessed by an MTT assay. *P<0.05,
**P<0.01, vs. 0 µM group.
#P<0.05, vs. 200 µM H2O2
group. n=5. GP, genipin. |
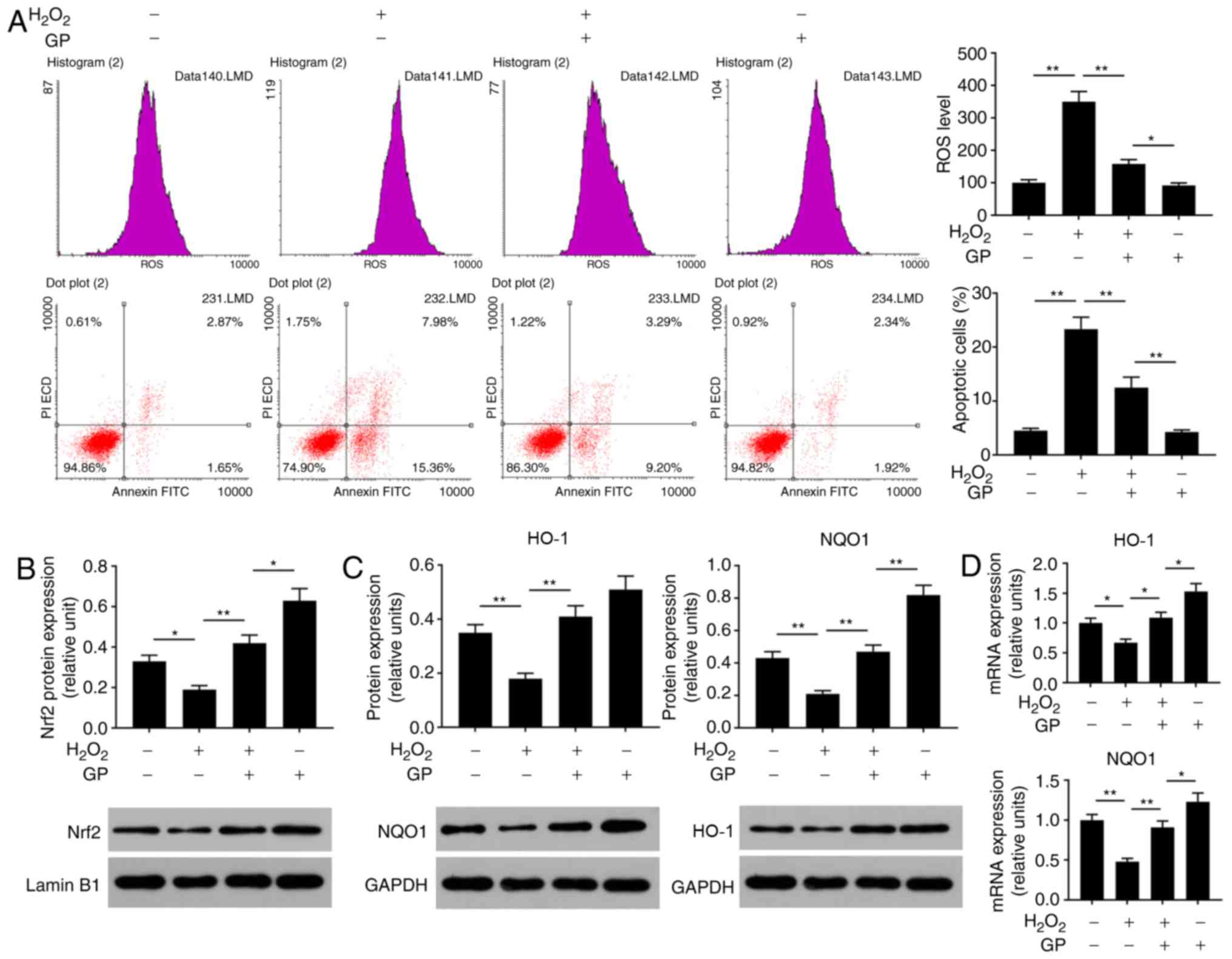
GP suppresses the effects of
H2O2 on the levels of ROS and apoptosis of
ARPE-19 cells by activating Nrf2 signaling
Oxidative stress is an important cause of injury to
RPE cells (22). To analyze the
effects of GP on H2O2-induced ARPE-19 cell
injury, the ROS levels, cell apoptosis and Nrf2 signaling were
detected. The data of flow cytometry revealed that ROS levels and
the number of apoptotic cells were significantly enhanced in cells
treated with H2O2 compared with in untreated
and H2O2-induced cells treated with GP
(Fig. 2A). In addition,
H2O2 significantly reduced the protein
expression levels of Nrf2, HO-1 and NQO1, and the mRNA expression
levels of HO-1 and NQO1 compared with the control (Fig. 2B-D). On the contrary, treatment
with GP significantly increased the expression of Nrf2, HO-1 and
NQO1 in H2O2-induced cells compared with
H2O2 treatment alone (Fig. 2B-D). The results suggested that GP
suppressed H2O2-induced RPE cell
injuries.
Nrf2 silencing enhances
H2O2-induced damage to ARPE-19 cells
In the present study, to determine the effects of
Nrf2 on ARPE cells, cell viability and the expression levels of
Nrf2, HO-1 and NQO1 were respectively analyzed by an MTT assay,
RT-qPCR and western blotting. When cells were transfected with
siRNA Nrf2 vector, no notable alterations in cell viability were
observed (Fig. 3A). As the
demonstrated by western blotting, the expression levels of NQO1,
HO-1 and Nrf2 proteins were significantly suppressed in the siNrf2
group compared with in the NC group (Fig. 3B and C). In addition, the mRNA
expression profiles of NQO1 and HO-1 were similar to the protein
expression profile in each group (Fig. 3D). In order to further investigate
the effects of siNrf2 on ARPE cells induced by
H2O2, the ROS levels, cell apoptosis and Nrf2
signaling were analyzed. The present study reported that siNrf2
significantly and markedly enhanced the ROS levels and apoptosis in
ARPE-19 cells induced by H2O2, respectively,
compared with the H2O2 + NC group (Fig. 4A). The expression levels of
apoptosis-associated factors were evaluated by RT-qPCR and western
blotting. SiNrf2 significantly downregulated the expression levels
of Nrf2, NQO1 and HO-1 in cells treated with
H2O2, compared with the
H2O2 + NC group (Fig. 4B, C and E). The results also
revealed that, compared with the H2O2 and
H2O2 + NC groups, siNfr2-transfected cells
treated with H2O2 exhibited increased Bax and
cleaved-caspase-3 expression levels; the protein expression levels
of Bcl-2 were significantly decreased in the
H2O2 + siNrf2 group compared with the
H2O2 and H2O2 + NC
groups (Fig. 4D and F). These
results suggested that Nrf2 knockdown enhanced
H2O2-induced RPE cell injury.
 | Figure 4Nrf2 silencing enhances
H2O2-induced damage to ARPE-19 cells. (A)
ARPE-19 cells were subjected to treatment with 200 µM
H2O2, NC vector and siNrf2 vector. Flow
cytometry was performed to analyze the levels of ROS and the number
of apoptotic cells. *P<0.05 vs.
H2O2 + NC group. (B) Proteins expression
levels of Nrf2, were determined using western blotting. (C) The
protein expression levels of NQO1 and HO-1 were detected by reverse
transcription-quantitative polymerase chain reaction. (D) The
protein expression levels of cleaved-caspase 3, Bax and Bcl-2 were
detected by western blotting. (E) The mRNA expression levels of
HO-1 and NQO1 were detected by reverse transcription-quantitative
polymerase chain reaction. (F) The mRNA expression levels of Bax
and Bcl-2 were detected by reverse transcription-quantitative
polymerase chain reaction. *P<0.05,
**P<0.01. n=4. Bcl-2, B-cell lymphoma 2; Bax,
Bcl-2-associated X; FITC, fluorescein isothiocyanate; HO-1, heme
oxygenase-1; NQO1, NAD(P)H: Quinine oxidoreductase; Nrf2, nuclear
factor-erythroid 2-related factor-2; PI, propidium iodide; siRNA,
small interfering RNA; NC, siRNA negative control; siNrf2, siRNA
Nrf2. |
Nrf2 silencing attenuates the protective
effects of GP on H2O2-induced ARPE-19
cells
In the present study, the role of Nrf2 in
GP-induced-effects on H2O2-treated cells was
investigated. Flow cytometry revealed that, compared with the
H2O2 + GP + NC group, the ROS levels and
number of apoptotic cells were significantly increased in the
H2O2 + GP + siNrf2 group; GP significantly
decreased ROS levels and apoptosis in
H2O2-induced cells compared with
H2O2 treatment alone (Fig. 5A). When cells were exposed to GP
and siNrf2 vector, the protein expression levels of Nrf2, NQO1,
HO-1, and Bcl-2 were significantly downregulated, while that of Bax
and cleaved-caspase-3 were significantly upregulated, compared with
the GP group. Additionally, compared with the
H2O2 + GP + NC group, the expression of Nrf2,
NQO1, HO-1, and Bcl-2 were inhibited in the
H2O2 + GP + siNrf2 group; the protein
expression levels of Bax and cleaved-caspase-3 were notably and
significantly increased, respectively (Fig. 5B-D). The mRNA expression profile
of Nrf2, NQO1, HO-1, Bcl-2 and Bax was similar to their respective
trend in protein levels (Fig. 5E and
F). The results of the present study suggested that the
activation of Nrf2 signaling was associated with the protective
effects of GP on H2O2-treated RPE cells.
 | Figure 5Nrf2 silencing attenuates the effects
of GP on H2O2-induced ARPE-19 cells. (A)
ARPE-19 cells were subjected to treatment with 200 µM
H2O2, 30 µM GP, NC vector and siNrf2
vector. Flow cytometry was applied to analyze the levels of ROS and
the number of apoptotic cells. *P<0.05,
**P<0.01. (B-D) Proteins expression levels of Nrf2,
HO-1, NQO1, Bax, Bcl-2 and cleaved-caspase 3 were determined by
western blotting. (E and F) mRNA expression levels of HO-1, NQO1,
Bax and Bcl-2 were detected by reverse transcription-quantitative
polymerase chain reaction. *P<0.05,
**P<0.01. n=4. Bcl-2, B-cell lymphoma 2; Bax,
Bcl-2-associated X; FITC, fluorescein isothiocyanate; HO-1, heme
oxygenase-1; NQO1, NAD(P)H: Quinine oxidoreductase; Nrf2, nuclear
factor-erythroid 2-related factor-2; PI, propidium iodide; siNrf2,
small interfering RNA Nrf2. |
Discussion
Oxidative stress is one of the most important
mechanisms underlying the pathogenesis of AMD (3). Oxidative damage of RPE is a key
process in the pathogenesis of AMD (4). H2O2 can induce
the production of ROS in cells, leading to oxidative damage
(23). Exogenous
H2O2 treatment is a simple and feasible cell
model for studying RPE oxidative damage, which can effectively
simulate the process of oxidative damage of RPE in AMD (24,25). Therefore, in the present study,
H2O2 was selected as an inducer of oxidative
damage to ARPE-19 cells. Additionally, 200 µM
H2O2 was identified as the optimal
concentration to generate a cellular oxidative damage model.
GP has been demonstrated to prevent or treat
cardiovascular diseases, diabetes, nervous system diseases,
pathogenic infections and inflammation (26-30). Lin et al (28) reported that GP protected renal
tissue from oxidative stress-associated injury by suppressing
oxidative stress and apoptosis. Shin and Lee (29) proposed that GP improved
age-associated insulin resistance by reducing oxidative stress in
LO2 cells; however, the role of GP in AMD remains unclear. In the
present study, it was suggested that GP exerted a protective effect
on H2O2-induced oxidative damage of ARPE-19
cells. The results of the present study revealed that GP (30, 50
and 100 µM) significantly increased the viability of ARPE-19
cells, indicating that the treatment of GP may induce some survival
signals in ARPE-19 cells; the increased cell viability in response
to 30 µM GP was similar to that of 50 and 100 µM.
Therefore, low, moderate and high concentrations of GP (5, 10 and
30 µM) were selected to analyze cell viability inhibited by
H2O2. The data demonstrated that 30 µM
GP significantly enhanced cell viability. Hence, 30 µM GP
was determined to be the optimal concentration for analysis in the
present study. The observations of the present study suggested that
GP exerted a protective effect on
H2O2-induced oxidative damage of ARPE-19
cells by enhancing cell viability.
Oxidative damage is caused by the imbalance of the
intracellular redox state, which generates a large amount of active
oxygen and produces free radicals (31). Providing the concentration of ROS
exceeds the clearance capacity of the body, tissue or cell damage
may occur (32,33). It has been reported that
antioxidant genes and drugs inhibit
H2O2-induced oxidative damage by decreasing
ROS activity (26,34-37). Studies have demonstrated that
oxidative stress injury is one of the important factors of
apoptosis (38-40). Radi et al (39) have reported that taxifolin
alleviated H2O2-induced oxidative stress
injury of ARPE-19 cells by inhibiting apoptosis. Therefore, ROS
levels and the apoptosis of ARPE-19 cells treated with
H2O2 and GP were analyzed by flow cytometry
in the present study. Similar to previous reports (34,36,37,41), the results of the present study
revealed that GP significantly reduced the levels of ROS and
apoptosis in ARPE-19 cells induced by H2O2.
In addition, GP significantly suppressed the expression of Bax and
cleaved-caspase 3, and promoted Bcl-2 expression. These
observations indicated that GP opposed the effects of
H2O2 on ARPE-19 cell injury via
anti-apoptosis and antioxidation.
Nrf2 signaling serves a key role in regulating
antioxidant enzymes, and is also an important part of maintaining
oxidative and antioxidative homeostasis, and alleviating oxidative
stress damage (42). HO-1 and
NQO1 are the key downstream factors of Nrf2 signaling, and serve an
important role in protecting cells from oxidative damage (43,44). Hu et al (43) indicated that microRNA-455
activated the Nrf2 signaling pathway to protect osteoblasts against
H2O2-induced injury. Vurusaner et al
(44) proposed that laminarin
ameliorated H2O2-induced MRC-5 cell oxidative
injury by promoting the Nrf2 signaling pathway (45). Previous studies have also reported
the beneficial effects of the Nrf2 signaling pathway on RPE cells
(46,47). In the present study, it was
proposed that the potential antioxidative mechanism of GP may
comprise Nrf2 signaling. The results demonstrated that GP reversed
the inhibitory effects of H2O2 on the
expression of Nrf2, HO-1 and NQO1 in ARPE-19 cells. In addition,
the effects of Nrf2 on ARPE-19 cells were investigated in the
present study. The results revealed that Nrf2 knockdown exhibited
no toxicity to cells; the effects of Nrf2 knockdown may be mainly
produced on the molecular level rather than at the cellular level.
However, Nrf2 silencing enhanced the effects of
H2O2 on inducing cell damage via increasing
ROS levels and apoptosis. Furthermore, Nrf2 silencing attenuated
the protective effects of GP on H2O2-induced
ARPE-19 cell injury by promoting apoptosis and oxidation.
Therefore, the activation of Nrf2 signaling may be closely
associated with the protective effects of GP. In addition, it has
been reported that GP suppressed the growth of breast cancer cells
by inhibiting UCP2 (14). Thus,
it is possible that other signals may also associate with the
protective effects of GP.
In summary, the present study proposed the novel
functions of GP, which protected ARPE-19 cells against oxidative
damage induced by H2O2 via promoting cell
viability and suppressing ROS levels and apoptosis. The molecular
mechanism was associated with the activation of Nrf2 signaling.
These results suggested that GP may be considered as a therapeutic
agent for the treatment and prevention of AMD.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors’ contributions
HZ wrote the manuscript. HZ, RW, MY and LZ
performed the experiments and data analysis. HZ and LZ designed the
study and contributed to manuscript revisions. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Nowak JZ: Age-related macular degeneration
(AMD): Pathogenesis and therapy. Pharmacol Rep. 58:353–363.
2006.PubMed/NCBI
|
|
2
|
Jager RD, Mieler WF and Miller JW:
Age-related macular degeneration. N Engl J Med. 358:2606–2617.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hernández-Zimbrón LF, Zamora-Alvarado R,
Ochoa-De la Paz L, Velez-Montoya R, Zenteno E, Gulias-Cañizo R,
Quiroz-Mercado H and Gonzalez-Salinas R: Age-related macular
degeneration: New paradigms for treatment and management of AMD.
Oxid Med Cell Longev. 2018:83746472018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu L and Wu XW: Nobiletin protects human
retinal pigment epithelial cells from hydrogen peroxide-induced
oxidative damage. J Biochem Mol Toxicol. 32:e220522018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y, Kim HJ and Sparrow JR: Quercetin
and cyanidin-3-glucoside protect against photooxidation and
photo-degradation of A2E in retinal pigment epithelial cells. Exp
Eye Res. 160:45–55. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dun Y, Vargas J, Brot N and Finnemann SC:
Independent roles of methionine sulfoxide reductase A in
mitochondrial ATP synthesis and as antioxidant in retinal pigment
epithelial cells. Free Radic Biol Med. 65:1340–1351. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang H, Wu M, Liu Y, Song L, Li S, Wang
X, Zhang YF, Fang J and Wu S: Serine racemase deficiency attenuates
choroidal neovascularization and reduces nitric oxide and VEGF
levels by retinal pigment epithelial cells. J Neurochem.
143:375–388. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee CH, Kwak SC, Kim JY, Oh HM, Rho MC,
Yoon KH, Yoo WH, Lee MS and Oh J: Genipin inhibits RANKL-induced
osteoclast differentiation through proteasome-mediated degradation
of c-Fos protein and suppression of NF-κB activation. J Pharmacol
Sci. 124:344–353. 2014. View Article : Google Scholar
|
|
9
|
Kim ES, Jeong CS and Moon A: Genipin, a
constituent of Gardenia jasminoides Ellis, induces apoptosis and
inhibits invasion in MDA-MB-231 breast cancer cells. Oncol Rep.
27:567–572. 2012.
|
|
10
|
Lim W, Kim O, Jung J, Ko Y, Ha J, Oh H,
Lim H, Kwon H, Kim I, Kim J, et al: Dichloromethane fraction from
Gardenia jasminoides: DNA topoisomerase 1 inhibition and oral
cancer cell death induction. Pharm Biol. 48:1354–1360. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Machida K, Oyama K, Ishii M, Kakuda R,
Yaoita Y and Kikuchi M: Studies of the constituents of Gardenia
species. II. Terpenoids from Gardeniae Fructus. Chem Pharm Bull
(Tokyo). 48:746–748. 2000. View Article : Google Scholar
|
|
12
|
Zuo T, Zhu M, Xu W, Wang Z and Song H:
Iridoids with genipin stem nucleus inhibit
lipopolysaccharide-induced inflammation and oxidative stress by
blocking the NF-κB pathway in polycystic ovary syndrome. Cell
Physiol Biochem. 43:1855–1865. 2017. View Article : Google Scholar
|
|
13
|
Hoshovs’ka IUV, Shymans’ka TV and Sahach
VF: Effect of UCP2 activity inhibitor genipin on heart function of
aging rats. Fiziol Zh. 55:28–34. 2009.In Ukrainian.
|
|
14
|
Ayyasamy V, Owens KM, Desouki MM, Liang P,
Bakin A, Thangaraj K, Buchsbaum DJ, LoBuglio AF and Singh KK:
Cellular model of Warburg effect identifies tumor promoting
function of UCP2 in breast cancer and its suppression by genipin.
PLoS One. 6:e247922011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dando I, Fiorini C, Pozza ED, Padroni C,
Costanzo C, Palmieri M and Donadelli M: UCP2 inhibition triggers
ROS-dependent nuclear translocation of GAPDH and autophagic cell
death in pancreatic adenocarcinoma cells. Biochim Biophys Acta.
1833:672–679. 2013. View Article : Google Scholar
|
|
16
|
Kensler TW, Wakabayashi N and Biswal S:
Cell survival responses to environmental stresses via the
Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol. 47:89–116.
2007. View Article : Google Scholar
|
|
17
|
Nguyen T, Nioi P and Pickett CB: The
Nrf2-antioxidant response element signaling pathway and its
activation by oxidative stress. J Biol Chem. 284:13291–13295. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang M, An C, Gao Y, Leak RK, Chen J and
Zhang F: Emerging roles of Nrf2 and phase II antioxidant enzymes in
neuroprotection. Prog Neurobiol. 100:30–47. 2013. View Article : Google Scholar :
|
|
19
|
Sachdeva MM, Cano M and Handa JT: Nrf2
signaling is impaired in the aging RPE given an oxidative insult.
Exp Eye Res. 119:111–114. 2014. View Article : Google Scholar :
|
|
20
|
Kang KA, Wang ZH, Zhang R, Piao MJ, Kim
KC, Kang SS, Kim YW, Lee J, Park D and Hyun JW: Myricetin protects
cells against oxidative stress-induced apoptosis via regulation of
PI3K/Akt and MAPK signaling pathways. Int J Mol Sci. 11:4348–4360.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Patel AK and Hackam AS: Toll-like receptor
3 (TLR3) protects retinal pigmented epithelium (RPE) cells from
oxidative stress through a STAT3-dependent mechanism. Mol Immunol.
54:122–131. 2013. View Article : Google Scholar :
|
|
23
|
Sies H: Role of metabolic
H2O2 generation: Redox signaling and
oxidative stress. J Biol Chem. 289:8735–8741. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen XD, Su MY, Chen TT, Hong HY, Han AD
and Li WS: Oxidative stress affects retinal pigment epithelial cell
survival through epidermal growth factor receptor/AKT signaling
pathway. Int J Ophthalmol. 10:507–514. 2017.PubMed/NCBI
|
|
25
|
Du L, Chen J and Xing YQ: Eupatilin
prevents H2O2-induced oxidative stress and
apoptosis in human retinal pigment epithelial cells. Biomed
Pharmacother. 85:136–140. 2017. View Article : Google Scholar
|
|
26
|
Chen S, Sun P, Zhao X, Yi R, Qian J, Shi Y
and Wang R: Gardenia jasminoides has therapeutic effects on
L-NNA-induced hypertensio in vivo. Mol Med Rep. 15:4360–4373. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen XL, Tang WX, Tang XH, Qin W and Gong
M: Downregulation of uncoupling protein-2 by genipin exacerbates
diabetes-induced kidney proximal tubular cells apoptosis. Ren Fail.
36:1298–1303. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin YH, Tsai SC, Lai CH, Lee CH, He ZS and
Tseng GC: Genipin-cross-linked fucose-chitosan/heparin
nanoparticles for the eradication of Helicobacter pylori.
Biomaterials. 34:4466–4479. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shin JK and Lee SM: Genipin protects the
liver from ischemia/reperfusion injury by modulating mitochondrial
quality control. Toxicol Appl Pharmacol. 328:25–33. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu D, Shi M, Bao J, Yu X, Li Y and Liu W:
Genipin ameliorates hypertension-induced renal damage via the
angiotensin II-TLR/MyD88/MAPK pathway. Fitoterapia. 112:244–253.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dias HKI, Milward M, Grant M, Chapple ILC
and Griffiths HR: Sulforaphane decreases neutrophil hyperactivity
by reducing intracellular oxidative stress. Free Rad Biol Med.
53:S492012. View Article : Google Scholar
|
|
32
|
Schieber M and Chandel NS: ROS function in
redox signaling and oxidative stress. Curr Biol. 24:R453–R462.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Slimen IB, Najar T, Ghram A, Dabbebi H,
Ben Mrad M and Abdrabbah M: Reactive oxygen species, heat stress
and oxidative-induced mitochondrial damage. A review. Int J
Hyperthermia. 30:513–523. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu H, Liu W, Zhou X, Long C, Kuang X, Hu
J, Tang Y, Liu L, He J, Huang Z, et al: Protective effect of lutein
on ARPE-19 cells upon H2O2-induced G2/M
arrest. Mol Med Rep. 16:2069–2074. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mitter SK, Song C, Qi X, Mao H, Rao H,
Akin D, Lewin A, Grant M, Dunn W Jr, Ding J, et al: Dysregulated
autophagy in the RPE is associated with increased susceptibility to
oxidative stress and AMD. Autophagy. 10:1989–2005. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu C, Dong Y, Liu H, Ren H and Cui Z:
Hesperetin protects against H2O2-triggered
oxidative damage via upregulation of the Keap1-Nrf2/HO-1 signal
pathway in ARPE-19 cells. Biomed Pharmacother. 88:124–133. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhu Y, Zhao KK, Tong Y, Zhou YL, Wang YX,
Zhao PQ and Wang ZY: Exogenous NAD(+) decreases oxidative stress
and protects H2O2-treated RPE cells against
necrotic death through the up-regulation of autophagy. Sci Rep.
6:263222016. View Article : Google Scholar
|
|
38
|
Areti A, Yerra VG, Naidu V and Kumar A:
Oxidative stress and nerve damage: Role in chemotherapy induced
peripheral neuropathy. Redox Biol. 2:289–295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Radi E, Formichi P, Battisti C and
Federico A: Apoptosis and oxidative stress in neurodegenerative
diseases. J Alzheimers Dis. 42(Suppl 3): S125–S152. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Su J, Shi HX, Wang LJ, Guo RX, Ren TK and
Wu YB: Chemical constituents of bark of Taxus chinensis var.
mairei. Zhong Yao Cai. 37:243–251. 2014.In Chinese. PubMed/NCBI
|
|
41
|
Xie X, Feng J, Kang Z, Zhang S, Zhang L,
Zhang Y, Li X and Tang Y: Taxifolin protects RPE cells against
oxidative stress-induced apoptosis. Mol Vis. 23:520–528.
2017.PubMed/NCBI
|
|
42
|
Huang Y, Li W, Su ZY and Kong AN: The
complexity of the Nrf2 pathway: Beyond the antioxidant response. J
Nutr Biochem. 26:1401–1413. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hu Y, Duan M, Liang S, Wang Y and Feng Y:
Senkyunolide I protects rat brain against focal cerebral
ischemia-reperfusion injury by up-regulating p-Erk1/2, Nrf2/HO-1
and inhibiting caspase 3. Brain Res. 1605:39–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vurusaner B, Gamba P, Gargiulo S, Testa G,
Staurenghi E, Leonarduzzi G, Poli G and Basaga H: Nrf2 antioxidant
defense is involved in survival signaling elicited by
27-hydroxycholesterol in human promonocytic cells. Free Radic Biol
Med. 91:93–104. 2016. View Article : Google Scholar
|
|
45
|
Liu X, Liu H, Zhai Y, Li Y, Zhu X and
Zhang W: Laminarin protects against hydrogen peroxide-induced
oxidative damage in MRC-5 cells possibly via regulating NRF2.
PeerJ. 5:e36422017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang K, Jiang Y, Wang W, Ma J and Chen M:
Escin activates AKT-Nrf2 signaling to protect retinal pigment
epithelium cells from oxidative stress. Biochem Biophys Res Commun.
468:541–547. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hu H, Hao L, Tang C, Zhu Y, Jiang Q and
Yao J: Activation of KGFR-Akt-mTOR-Nrf2 signaling protects human
retinal pigment epithelium cells from Ultra-violet. Biochem Biophys
Res Commun. 495:2171–2177. 2018. View Article : Google Scholar
|