Introduction
Inflammatory bowel disease (IBD), including
ulcerative colitis (UC) and Crohn's disease (CD), are chronic
relapsing intestinal pathologies triggered by undefined etiological
factors (1,2). The pathogenesis of IBD is complex,
and no consensus theory is yet available to fully elucidate the
development of IBD. The development and progression of IBD may
involve a number of factors, such as the environment, genetic and
intestinal infections, immunity dysregulation and mucosal barrier
defects (3-5). Currently, the therapies for IBD
focus on controlling active inflammatory reactions and regulating
intestinal immune disorders with the use of immunosuppressive
agents for example, which are considered the most effective drugs
for the treatment of IBD; however, these have significant
side-effects (6).
Previous studies have demonstrated that mesenchymal
stem cells (MSCs) exert therapeutic effects against several immune
disorders, including IBD due to their potent immunomodulatory
effects and tissue regenerative potential (7-9).
The paracrine theory of stem cell action has provided new insight
into the mechanisms of MSCs in the treatment of several diseases
(10,11). Increasing evidence has indicated
that MSCs regulate the repair of damaged tissues and immune balance
through their paracrine soluble factors (12-15). Extracellular vesicles (EVs), tiny
membrane vesicles that can be secreted by the majority of cells,
have a lipid bilayer membrane structure of approximately 30-200 nm
in diameter (12). Recent studies
have found that EVs released by cells, acting as mediators of MSC
paracrine actions, can be used to communicate between cells
(16-18). MSC-derived EVs (MSC-EVs) contain a
wide variety of bioactive substances, such as mRNAs, microRNAs
(miRNAs or miRs) and proteins, which are not easily degraded
(18-20). Furthermore, several studies have
indicated that MSC-EVs exert therapeutic effects similar to those
of MSCs (11,12,21,22). It has recently been found that the
paracrine function of MSCs is mediated by EVs at least to a certain
extent. EVs are mainly released from the endosomal compartment and
contain certain bioactive substances, such as mRNAs, miRNAs and
proteins from their cells of origin (23). Researchers have further confirmed
the endosomal origin of MSC-EVs by using the method of lipid raft
composition detection (24). MSCs
can promote the healing of tissue ischemic diseases by secreting
proangiogenic secretory proteins (10). It has recently been further
demonstrated that MSC-derived EVs act as paracrine effectors of
angiogenesis; however, which components of the EVs proteome are
responsible for the identification of this role remains unknown. A
previous study performed proteomics analysis and demonstrated that
MSC-derived EVs included angiogenic paracrine effectors, which have
the potential to treat tissue ischemic diseases (25). Compared with stem cell therapy,
EVs do not cause acute immune rejection, do not constitute a risk
of tumor formation, and can be easily stored and transported
(13). Furthermore, it has been
demonstrated that the administration of EVs derived from umbilical
cord-MSCs can alleviate colitis in mice, and EVs are administered
via tail vein injection to mice (26). Thus, MSC-EVs are likely to become
a novel and more efficient cell-free therapy approach for IBD.
In the present study, it was hypothesized that the
in situ injection of human placental (hP-)MSC-derived EVs
would significantly improve the clinical symptoms and exert
beneficial effects by promoting mucosal healing via the inhibition
of inflammation and oxidative stress in mice with colitis. It was
also hypothesized that the method of administration of the EVs
would ensure that they localized directly and rapidly in the
damaged intestine for a longer period of time in order to enhance
their therapeutic effects. To examine this hypothesis, the
therapeutic effects of EVs were assessed in a murine model of
colitis induced by trinitrobenzene sulfonic acid (TNBS).
Inflammation, oxidative stress and mucosal healing were also
evaluated. The findings suggest that EVs derived from hP-MSCs may
represent a novel therapeutic approach for IBD by in situ
injection.
Materials and methods
Cells and cell culture
The hP-MSCs used in the present study were provided
by Cell Products of National Engineering Research Center/Tianjin
Amcellgene Engineering Co., Ltd. The hP-MSCs were cultured in
Dulbecco's modified Eagle's medium (DMEM)/F12 medium (Gibco; Thermo
Fisher Scientific, Inc.) with 10% bovine EV-free fetal bovine serum
(FBS; HyClone; Cytiva), 1% L-glutamine (Gibco; Thermo Fisher
Scientific, Inc.), 1% non-essential amino acid (NEAA) and 1% 100
U/ml penicillin/streptomycin (Gibco; Thermo Fisher Scientific,
Inc.). Bovine EV-free FBS was obtained by ultracentrifugation at
1,00,000 × g for 70 min at 4°C, and filtered using a 100 nm filter
(27).
Isolation and characterization of
EVs
EVs were purified from supernatants of hP-MSCs by
differential centrifugation (12,13,15). In brief, hP-MSCs were cultured in
medium containing 10% EV-free FBS for 48 h, and the supernatant was
then collected. The supernatant was then subjected to sequential
centrifugation steps at 500 × g for 10 min to remove any cell
contaminations, at 2,000 × g for 20 min to remove apoptotic bodies,
and at 5,000 × g for 30 min to remove cell debris at 4°C. The
resulting supernatant was then filtered using 0.2 µm filters
(Merck KGaA). The EVs were then harvested by centrifugation at
130,000 × g for 2 h in a SW32 Ti rotor (L-100XP Ultracentrifuge,
Beckman Coulter) at 4°C. Finally, the EV pellets were resuspended
in PBS and ultracentrifuged again at 130,000 × g for 2 h to discard
the contaminating proteins, and the purified EVs were harvested.
The morphology of the EVs was examined by transmission electron
microscopy (TEM; Talos F200C, Thermo Fisher Scientific, Inc.). The
particle size of the EV pellets was measured by dynamic light
scattering using a BI-200SM laser scattering instrument (ZetaPALS,
Brookhaven Instruments) at 20°C. The protein concentrations of the
EVs were measured using a BCA Protein assay kit (Promega
Corporation) according to the manufacturer's instructions. The
presence of known EV markers, including CD63 (1:1,000 dilution,
ab216130, Abcam), CD9 (1:1,000 dilution, ab92726, Abcam), ALIX
(1:1,000 dilution, ab186429, Abcam) and GM130 (1:1,000 dilution,
ab52649, Abcam) were examined by western blot analysis.
Western blot analysis
Mouse colon tissues (described below) used for
western blot analysis were lysed on ice in radioimmunoprecipitation
assay (RIPA) buffer (Beijing Solarbio Science & Technology Co.,
Ltd.), and its quantification was measured using the BCA™ Protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The equal amount proteins (30
µg) were separated by 12% sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE; Invitrogen; Thermo
Fisher Scientific, Inc.) and transferred to polyvinylidene
difluoride membranes (PVDF; EMD Millipore). The membranes were
blocked with 5% non-fat milk in tri-sec-buffered saline/Tween-20
(TBST) buffer (20 mM Tris-HCl, pH 7.6, 136 mM NaCl and 0.1%
Tween-20). After blocking with 5% non-fat milk for 2 h, the
membranes were incubated with primary antibodies overnight at 4°C,
and then for 2 h at room temperature with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibodies
(1:5,000 dilution, ab97051, Abcam). The Pierce enhanced
chemiluminescence western blotting substrate (EMD Millipore) was
used to detect the signal. The primary antibodies used for western
blot analysis were as follows: Rabbit anti-CD9 (1:1,000 dilution,
ab92726, Abcam), CD63 (1:1,000 dilution, ab216130, Abcam), GM130
(1:1,000 dilution, ab52649, Abcam), and rabbit anti-interleukin
(IL)-1β (1:1,000 dilution, WL02257, Wanleibio), IL-6 (1:1,000
dilution, ab229381, Abcam), tumor necrosis factor (TNF)-α (1:1,000
dilution, WL01581, Wanleibio), IL-10 (1:1,000 dilution, ab271261,
Abcam), interferon (IFN)-γ (1:1,000 dilution, WL02440, Wanleibio),
EpCAM (1:1,000 dilution, WL01375, Wanleibio). The above-mentioned
antibodies were used following the manufacturer's instructions.
Clarity Western ECL substrate (Bio-Rad Laboratories, Inc.) and
ChemiDoc™ MP Imaging System (Bio-Rad Laboratories, Inc.) were
applied to detect the blots and for visualization. In addition,
ImageJ version 1.46 (Rawak Software, Inc.) was used to quantify the
blots.
Animal model
For all the experiments, 8-week-old male BALB/c mice
were utilized that were purchased from the Laboratory Animal Center
of the Academy of Military Medical Science. The mice were housed in
a standard animal laboratory where the temperature was maintained
at 25°C with a humidity level of 30-60%. The animals were provided
with free access to food and water. The protocols involved animals
and the experimental procedures of the present study were approved
by the Nankai University Animal Care and Use Committee guidelines
(approval no. 20170022) and conducted according to the
international regulations of the usage and welfare of laboratory
animals. All experimental procedures were conducted in accordance
with the Tianjin Committee for the Use and Care of Laboratory
Animals. The male BALB/c mice were randomly divided into 3 groups
(n=8/group) as follows: The sham-operated group (Sham group), the
IBD group (PBS group) and the EV-treated IBD group (EV group). As
mentioned above, colitis was induced in the mice with TNBS
(Sigma-Aldrich; Merck KGaA) (28). The TNBS-induced model of colitis
can be established by a single enema without prior sensitization
and with a long duration of inflammation, and it is a dynamic
process of transformation from acute inflammation to chronic
inflammation. The advantages of the TNBS-induced model of colitis
include the reproducibility and the technical simplicity. In brief,
TNBS (100 mg/kg) dissolved in 50% ethanol was slowly administered
into the colon via a catheter equipped approximately 3.5 cm into
the anus. The mouse was suspended for 1 min to allow the drug to
fully absorb. On the first day after modeling, the mice were
anesthetized by 4% chloral hydrate (350 mg/kg) and a laparotomy was
performed. A total of 200 µg EVs suspended in PBS were
injected into the mice in the EV group in situ by injection
at a 60 µl total volume. The position of in situ
administration were the injury colon mesangial margin. The same
volume of PBS was administered to the mice in the PBS group as the
control. The sham-operated mice were subjected to the same surgical
procedure without colitis or EV injection. The body weight, stool
consistency and mental state of the mice were measured daily. The
mice were monitored every 12 h within a period of 1 week to examine
their health and behavior. The mice were then euthanatized by
cervical dislocation when the following humane endpoints were
reached: The mice lost weight rapidly, and the loss of body weight
were 15-20% of their original weight; exhibited a complete loss of
appetite for up to 24 h; were unable to eat and drink by
them-selves; exhibited severe persistent gastrointestinal symptoms
(persistent diarrhea, intestinal obstruction, intussusception and
peritonitis). Respiratory arrest, no heartbeat and no blink reflex
were used to confirm mouse death. No mice died before meeting these
endpoints. The animal experiment strictly adhered to the principles
of using the least number of animals to complete the experiment and
minimizing the pain of the experimental animals. All mice were
euthanized by cervical dislocation at 3 days after EVs treatment
(apart from those included in the survival analysis), and the colon
tissues were then collected for subsequent analysis. For in
vivo EV trafficking assay, 200 µg EVs were incubated
with 1 µmol/l Dil (Beijing Ouhe Technology Co., Ltd.). At 30
min after incubation, the labeled EVs were collected and injected
into the mice. The presence of labeled EVs in colon tissues was
detected in a live animal imaging system.
Histological analysis
At day 3 following TNBS enema, the mice were
euthanized, and colonic segments were harvested. In order to
determine the injury to the colon mucosa, the paraffin-embedded
sections were stained with hematoxylin and eosin (H&E)
(Beyotimebio) according to standardized procedures. Colonic
segments were washed in PBS, fixed in 4% paraformaldehyde (pH 7.4),
embedded in paraffin, and stained with H&E. The colon sections
were stained using hematoxylin for 5-10 min, rinsed with distilled
water for 1 min, separated color with 0.5% alcohol hydrochloric
acid at 37°C. The colon sections were then stained with eosin for
2-5 min, dehydrated using graded ethanol, vitrification by
dimethylbenzene, and mounted with neutral balsam at 37°C.
Immunohistochemical staining was carried out to determine the
inflammation, and proliferation and apoptosis of the damaged
intestinal tissue from the mice in the different groups. The
paraffin-embedded sections were dewaxed by xylene and hydrated in a
graded series of ethanol. The sections were treated with the
appropriate amount of endogenous peroxidase blocker and incubate
for 10 min at room temperature (37°C) to inactivate endogenous
peroxidase activity. The sections were then incubated with primary
antibodies overnight at 4°C for myeloperoxidase (MPO) (1:150
dilution, WL02355, Wanleibio), Ki67 (1:100 dilution, WL01384a,
Wanleibio) and EpCAM (1:1,000 dilution, WL01375, Wanleibio). The
sections were then incubated with horseradish peroxidase-conjugated
goat anti-rabbit secondary antibody (1:5,000 dilution, ab97051,
Abcam) for 20 min at 37°C, and DAB was used as the substrate.
Immunohistochemical positive staining was brown. The sections
obtained were examined with an optical micro-scope (Olympus BX51).
For immunofluorescence staining, the colon samples were embedded
into optimal cutting temperature (OCT) compound (Sakura Finetek).
Samples were cut into 6-µm-thick sections. The sections were
then incubated with primary antibodies overnight at 4°C for EpCAM
(1:200 dilution, ab71916, Abcam). The sections were incubated with
fluorescently labeled secondary antibodies (Alexa Fluor 488 goat
antimouse, 1:1,000 dilution, A-21151, Thermo Fisher Scientific,
Inc.) for 2 h at 37°C, and the sections were then counterstained
with DAPI (Abcam) for 5-10 min at 37°C to identify cell nuclei
(Olympus BX51). Images were analyzed using ImageJ version 1.46
(Rawak Software, Inc.).
Bioluminescence imaging
To examine the severity of injury to the intestine,
reactive oxygen species (ROS) were detected in the mice for 1 week
via an intraperitoneal injection of luminol (10 mg/kg;
Sigma-Aldrich; Merck KGaA). The luminol stock solutions were
prepared in normal saline prior to injection. The mice were imaged
with the IVIS Lumina II system (Xenogen Corp.) 5 min after the
injection of the substrate. Bioluminescence signals were quantified
in units of maximum photons per second per centimeter squared per
steradian (photons/sec/cm2/sr) as previously described
(29-31). All mice were euthanized by
cervical dislocation at 7 days after the experiment.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
The extracted colon tissue were suspended in TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and total RNA
was extracted from the colon samples according to the
manufacturer's instructions. The BioScript All-in-One cDNA
Synthesis SuperMix (Bimake) was used to produce cDNA by reverse
transcription. Gene expression was analyzed by ABI 7500 Fast
Real-Time PCR System (Bio-Rad Laboratories, Inc.) using a FastStart
Universal SYBR-Green Master (Roche Diagnostics) according to the
manufacturer's instructions. The relative gene expression fold
changes were calculated using the 2−ΔΔCq method
(32). The primers used for
RT-qPCR in the present study are presented in Table SI.
MPO activity assay
MPO is a marker of activation of neutrophils, and
changes in its activity represent the function and activity status
of neutrophils. MPO is a clinical examination index for
inflammatory bowel disease. In the present study, the measurement
of MPO activity in the colon in the homogenates was evaluated using
an MPO kit (Nanjing Jiancheng Technology Co., Ltd.) according to
the instructions of the manufacturer.
Statistical analyses
The experimental data are expressed as the means ±
standard deviation (SD). Multiple groups were compared by one-way
analysis of variance (ANOVA), followed by a Tukey's post hoc test.
Data were analyzed using GraphPad software (GraphPad Prism
Software, Inc.). Differences were considered statistically
significant at P-values <0.05.
Results
Characterization of EVs
EVs were isolated from the supernatant of hP-MSC
culture by sequential ultracentrifugation. The morphology of the
EVs was observed by TEM. As shown in Fig. 1A, the EVs exhibited a
'saucer'-like or hemispherical shape with a depression on one side.
The results of dynamic light scattering analysis revealed that the
diameter of the EVs was approximately 110 nm (Fig. 1B). The results of western blot
analysis indicated that the specific marker of EVs, including CD9,
CD63 and Alix were normally expressed and that they were negative
for GM130 protein expression (Fig.
1C-F). These results confirmed that the EVs are successfully
isolated from the hP-MSCs.
EVs ameliorate TNBS-induced colitis in
mice
Following the establishment of the model, the
survival, weight loss, colon length, disease activity index (DAI)
were observed, and a histological evaluation of the mice with
TNBS-induced colitis was performed. The results revealed that the
administration of hP-MSC-derived EVs significantly improved
clinical parameters, such as the survival rate of the mice with
colitis, changes in body weight and DAI compared to the mice
treated with PBS (Fig. 2A-C). On
the 3rd day following administration, the mice were sacrificed and
the colon were collected. As shown in Fig. 2D and E, the length of the colon in
the EV group was longer than that in the PBS group. The colon
segments of the mice in each group were stained with H&E to
observe the pathological changes. For the histological examination,
the results revealed that compared with the PBS group, inflammatory
cell infiltration significantly decreased, the number of intestinal
glands markedly increased, and crypt abscesses were reduced in the
EV-treated group (Fig. 2G).
Grading was performed in a blinded manner by a pathologist and the
scoring result is presented in Fig.
2F. On the whole, the administration of hP-MSC-derived EVs
markedly relieved the symptoms of TNBS-induced IBD in mice. To
examine the biodistribution of EVs in the injured colon tissues of
mice with colitis, EVs were labeled with Dil (Fig. S1A) to monitor the retention of
transplanted EVs by the live animal imaging system. The data
revealed the strong red fluorescence signals from the colon tissues
at days 0 and 1 following the injection of the Dil-EVs. In
comparison with the obtained signals on days 0 and 1, the
fluorescence signals on day 2 were weak. There was almost no
fluorescence observed on day 3 (Fig.
S1B).
 | Figure 2hP-MSC-derived EVs attenuate colitis
in mice. (A) Survival rate of mice in the sham, PBS and EV groups
(n=8). (B) Percentage of body weight loss following treatment. (C)
The disease activity index (DAI) score was used to estimate the
severity of symptoms of mice with TNBS induced colitis, seven days
in a row (n=8). (D) Macroscopic images of colonic tissues at day 3
after treatment. (E) Colon length in the sham, PBS and EV groups.
(F) Histopathological score in the sham, PBS and EV groups. (G)
H&E staining of representative histological sections of mouse
colons in the sham, PBS and EV groups. Scale bar, 100 µm. In
the images, the numbered arrows indicate the following: 1,
Inflammatory cells; glands; and 3, crypt abscesses. Data are
expressed as the means ± SD; (n=8). *P<0.05,
**P<0.01, ***P<0.001 vs. sham group;
#P<0.05, ##P<0.01 vs. PBS group. EVs,
extracellular vesicles; hP-MSCs, human placenta-derived mesenchymal
stem cells. |
EVs reduce intestinal inflammation in
mice
To further investigate the mechanisms responsible
for the alleviation of the symptoms of colonic inflammation in mice
by EVs, the expression of pro-inflammatory and anti-inflammatory
cytokines in mouse colon tissues at the gene and protein level were
examined by RT-qPCR and western blot analysis, respectively. The
results demonstrated that the gene expression levels of
pro-inflammatory cytokines (IL-1β, TNF-α, IL-6 and IFN-γ) in the
EV-treated group were significantly lower than those in the PBS
group (Fig. 3A). To further
corroborate these results, the protein expression levels of
pro-inflammatory cytokines were detected by western blot analysis
and the amount of target protein was calculated by gray scanning
(Figs. 3B and S2). Subsequently, the expression of
anti-inflammatory cytokines in the colon tissues of the mice in
each group were examined. The results revealed that the expression
of anti-inflammatory cytokines (IL-10 and TGF-β) in the EV-treated
group were significantly higher than those in the PBS group
(Fig. 3C). The results of western
blot analysis revealed that the levels of anti-inflammatory factors
(IL-10 and TGF-β) increased significantly in the EV-treated group
compared with the PBS group (Fig.
3D). These results indicated that the hP-MSC-derived EVs
attenuated colitis by regulating the balance between
pro-inflammatory and anti-inflammatory cytokines in mice with
TNBS-induced colitis.
Effect of EV administration on
antioxidant defenses in the colitis mice
To examine the therapeutic effects of hP-MSC-derived
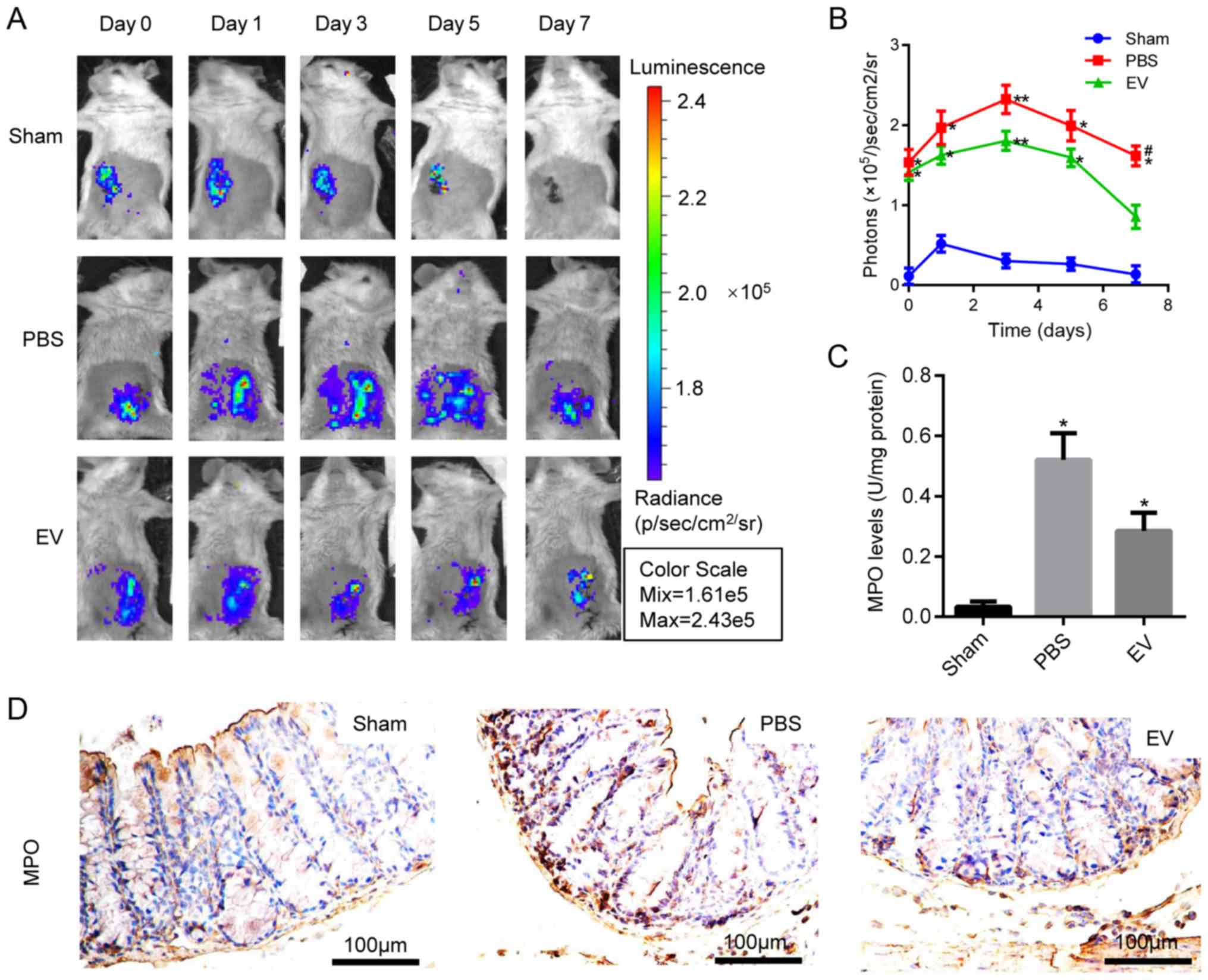
EVs on oxidative stress in TNBS-induced colitis, the IVIS Lumina
Imaging System was used to measure the levels of ROS in colon
tissues. EV administration significantly decreased the ROS content,
and the light signal on the 7th day was extremely low, indicating a
lower ROS content and a reduced inflammatory response (Fig. 4A and B). Moreover, MPO activity
was measured in the colon homogenate to deter-mine the number of
activated polymorphonuclear leukocytes. The results revealed that
MPO activity was significantly downregulated in the EV-treated
group compared with the PBS group (Fig. 4C). The above-mentioned results
were further confirmed by immunohistochemical staining for MPO
(Figs. 4D and S3). These results suggested that EVs
inhibited the oxidative stress reaction in the mice with
colitis.
EVs downregulate the expression of
apoptotic proteins
The expression of caspase-3, caspase-8 and caspase-9
at the gene level was determined, as markers of colonic cell
apoptosis. In comparison to the PBS group, the levels of
apoptosis-related genes were markedly decreased in the mice with
colitis administered EVs (Fig.
5A). The results of the analysis based on the
immumohistochemical staining of caspase-3 protein expression were
in accordance with those of RT-qPCR (Fig. 5B). The mean of integrated option
density (IOD) of caspase-3 was measured with imaging analysis and
then statistically analyzed (Fig.
S4). All these data indicated that the administration of EVs
played an important role in protecting epithelial cells from injury
in mice with colitis.
EVs promote mucosal healing in mice with
colitis
The present study then investigated whether EVs play
a role in intestinal epithelial cells (IECs) in mice with colitis.
The assessment of Cldn1, ZO-1 and Occludin mRNA expression by
RT-qPCR revealed that their expression markedly increased in the EV
group compared with the PBS group (Fig. 6A). In contrast to the
above-mentioned results, the mRNA expression levels of MMP-2 and
MMP-9 were decreased in EV group (Fig. 6B). Subsequently, the expression of
EpCAM was examined in the different groups. The results of western
blot analysis revealed that the expression of EpCAM in EVs group
were significantly higher than that in the PBS group (Fig. 6C). In addition, EpCAM
immunofluorescence staining and immunohistochemical staining
revealed that the colonic tissues from the mice in the EV group
maintained a better epithelial integrity, compared with those from
mice in the PBS group (Fig. S5A and
B). The immunohistochemical-mediated examination of Ki67
protein expression revealed that the EVs promoted the proliferation
of IECs (Figs. 6D and S6).
Discussion
In the present study, EVs derived from hP-MSCs were
utilized for the treatment of colitis by in situ injection.
The results revealed that the hP-MSC-derived EVs attenuated
TNBS-induced colitis, as assessed by body weight loss and the DAI
histological scores. Furthermore, it was found that the
administration of hP-MSC-derived EVs markedly reduced intestinal
inflammation and oxidative stress.
A number of studies have demonstrated that
hP-MSC-based therapy could be developed as a potential treatment
for several diseases (10,33,34).
The application of hP-MSCs in various disease models has several
advantages (34). First, the
placenta has wide resources and the use of this is not associated
with any ethical concerns. Second, hP-MSCs are easily available and
have a low immunogenicity and viral infection rate, which have more
widespread application prospect than bone marrow-MSCs. Moreover,
MSCs derived from umbilical cord blood have unavoidable
immunogenicity, and the available amniotic fluid is limited.
Therefore, hP-MSCs are a better choice, compared with other
sources. In addition, a previous study demonstrated that hP-MSCs
retained a higher therapeutic efficacy than bone marrow-derived
MSCs in a model of hindlimb ischemia disease (35). MSC-derived EVs reflect the
characteristics of their source cells; thus, there are differences
between different stem cell-derived EVs. There are marked
differences in growth factors and cytokines secreted by MSCs from
different sources. Studies have reported that EVs derived from MSCs
mainly exert biological effects via the following mechanisms
(16,20,36,37): Surface protein molecules and lipid
ligands of EVs can directly bind to related receptors on the
surface of target cells and further activate the signaling pathways
in target cells; EVs can directly enter target cells through fusion
or endocytosis with the target cell, so that the bioactive
substances such as proteins, lipid and nucleic acid carried by them
can be brought into the target cell, to further regulate the
function of the target cell and exert the therapeutic effect. EVs
derived from MSCs carry a variety of biologically active substances
(18). The exact molecular
mechanisms of action EVs remain unknown, and in future studies, the
authors aim to investigate and clarify the types of components in
EVs which play therapeutic roles.
Cytokine responses play an important role in the
pathophysiology of IBD, including Crohn's disease and ulcerative
colitis (38). Furthermore, the
cytokine responses of IBD continue during the entire process of
inflammation, and are also a major pathophysiological factor in the
eventual resolution of inflammation (39). In particular, intestinal
inflammation and the destruction of the intestinal mucosa are
related to the imbalance in pro-inflammatory and anti-inflammatory
cytokines (40). Therefore, the
development of strategies with which to regulate this imbalance
would be an effective treatment for IBD. In the present study, it
was confirmed that the administration of EVs downregulated the
expression of pro-inflammatory cytokines (TNF-α, IL-1β, IFN-γ and
IL-6) in colon tissues of mice with colitis, compared to those
treated with PBS. On the contrary, the administration of EVs
upregulated the expression of anti-inflammatory cytokines, such as
IL-10 and TGF-β. These results indicated that EV administration
regulated the balance of pro-inflammatory and anti-inflammatory
cytokines in mice with TNBS-induced colitis.
A growing body of evidence has indicated that
oxidative stress plays a crucial role in the pathogenesis and
progression of IBD, and is considered to be involved in the
infiltration and activation of neutrophils in damaged intestinal
tissue (41,42). The increase in ROS generation and
the decrease in anti-oxidant activities could contribute to the
major pathogenesis of IBD (43).
MPO, as a biochemical indicator of neutrophil infiltration and
activation in the intestinal mucosa, is widely applied to evaluate
the severity of intestinal inflammation in IBD (44). MPO secreted by the activated
neutrophils, a potent oxidant, plays an important role in oxidative
stress by catalyzing hydrogen dioxide (H2O2)
to hypochlorite (HOCL) (41). The
present study demonstrated that the administration of
hP-MSC-derived EVs markedly decreased the accumulation of
neutrophils and MPO activity in injured sites in mice with colitis.
The dynamic equilibrium of ROS production and metabolism is crucial
for the maintenance of the normal function of cells and tissues, if
this balance is disrupted, it can lead to oxidative stress and a
series of tissue damage (43).
The results of the present study further confirmed that the
hP-MSC-derived EVs decreased the levels of ROS in mice with
TNBS-induced colitis and protected the cells against oxidative
damage.
Furthermore, the formation of intestinal damage in
IBD is closely related to the production of ROS, and the overload
of ROS alter tight junctions and lead to pathological changes in
epithelial permeability. Previous studies have found that oxidative
stress may cause apoptosis. Abnormal apoptosis may lead to the
disruption of intestinal mucosal integrity, and induce the invasion
of pathogenic agents and bacteria, resulting in the continuous
activation of intestinal T-cells, and pro-inflammatory cytokines
can further promote IEC apoptosis in this process (43,45). In order to investigate the
disorder of colonic epithelium apoptosis in mice with TNBS-induced
colitis, the present study detected the expression of certain genes
and protein related to apoptosis. Following the administration of
hP-MSC-derived EVs, it was found that the EVs decreased the gene
expression levels of caspase-3, caspase-8 and caspase-9, and
decreased the protein expression of caspase-3.
Mucosal healing has been regarded as an important
therapeutic goal in IBD, and the evaluation of mucosal healing is
based on the integrity of the gut epithelium. The integrity of the
epithelial monolayer is formed by IECs with intercellular junctions
between adjacent cells (46),
which creates the continuous physical barrier (47). The gastric mucosal barrier is the
first barrier to come in contact with the intestinal tract and
external environment, and plays an important role in reducing the
invasion of pathogens and the absorption of toxins. Although the
intestinal tract is a protective barrier, pathogens or other
substances in the intestinal tract can stimulate IECs and
polymorphonuclear neutrophils to secrete inflammatory mediators and
promote oxidative stress. When damaged, it can lead to an increase
in intestinal epithelial permeability, which plays an important
role in the pathogenesis of IBD. In order to maintain the integrity
of the gut epithelium and the intestinal barrier, IECs need to
proliferate constantly, which is crucial for mucosal healing
(48,49). Tight junction proteins, such as
Cldn1, Zo1 and Occludin, function as indicators for the evaluation
of colonic epithelial integrity in mice with colitis (50). In the present study, it was found
that hP-MSC-derived EVs maintained proper tight junctions in the
colons of the treated mice, and promoted the proliferation of the
colon epithelial cells. Taken together, these results indicated
that hP-MSC-derived EVs played an important role in promoting
mucosal healing.
In conclusion, the present study developed a method
for treatment of experimental colitis in mice with the use of
hP-MSC-derived EVs. EVs markedly relieved the clinical symptoms by
inhibiting inflammation and oxidative stress to promote mucosal
healing in mice with TNBS-induced colitis. Moreover, molecular
imaging used in the present study elucidated the effects of EV
administration on antioxidant defenses in mice with colitis by the
real-time tracking of ROS in living mice. In general, the findings
of the present study provide new insight into the treatment of IBD.
EVs derived from hP-MSCs may be used as a novel therapeutic
strategy for IBD.
Supplementary Data
Acknowledgments
Not applicable.
Funding
The present study was partially supported by granst
from the National Key R&D Program of China (no.
2017YFA0103200), National Natural Science Foundation of China (no.
81671734), Key Projects of Tianjin Science and Technology Support
Program (no. 18YFZCSY00010), and National Natural Science
Foundation of China (no. 81470808).
Availability of data and materials
All data generated and/or analyzed during this study
are available from the corresponding author upon reasonable
request.
Authors' contributions
ZL, ZG and XC conceived and designed the
experiments. LD performed the majority of the experiments. HH, XZ,
MZ, SC, CW, ZH and ZCH collected and analyzed the data. LD and ZL
wrote the main manuscript and prepared the figures. ZL provided
funding. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The protocols involved animals and the experimental
procedures of the present study were approved by the Nankai
University Animal Care and Use Committee guidelines (Approval no.
20170022) and conducted according to the international regulations
of the usage and welfare of laboratory animals. All experimental
procedures were conducted in accordance with the Tianjin Committee
for the Use and Care of Laboratory Animals. All research was
conducted in accordance with the provided protocol. The animal
experiment strictly adhered to the principles of using the least
number of animals to complete the experiment and minimizing the
pain of the experimental animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kim DH and Cheon JH: Pathogenesis of
inflammatory bowel disease and recent advances in biologic
therapies. Immune Netw. 17:25–40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen X, Cai C, Xu D, Liu Q, Zheng S, Liu
L, Li G, Zhang X, Li X, Ma Y, et al: Human mesenchymal stem
cell-treated regulatory CD23+CD43+ B cells alleviate intestinal
inflammation. Theranostics. 9:4633–4647. 2019. View Article : Google Scholar :
|
|
3
|
Manichanh C, Borruel N, Casellas F and
Guarner F: The gut microbiota in IBD. Nat Rev Gastroenterol
Hepatol. 9:599–608. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baumgart DC and Sandborn WJ: Crohn's
disease. Lancet. 380:1590–1605. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ordás I, Eckmann L, Talamini M, Baumgart
DC and Sandborn WJ: Ulcerative colitis. Lancet. 380:1606–1619.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chudy-Onwugaje KO, Christian KE, Farraye
FA and Cross RK: A state-of-the-art review of new and emerging
therapies for the treatment of IBD. Inflamm Bowel Dis. 25:820–830.
2019. View Article : Google Scholar :
|
|
7
|
Dave M, Mehta K, Luther J, Baruah A, Dietz
AB and Faubion WA Jr: Mesenchymal stem cell therapy for
inflammatory bowel disease: A systematic review and meta-analysis.
Inflamm Bowel Dis. 21:2696–2707. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
González MA, Gonzalez-Rey E, Rico L,
Büscher D and Delgado M: Adipose-derived mesenchymal stem cells
alleviate experimental colitis by inhibiting inflammatory and
autoimmune responses. Gastroenterology. 136:978–989. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ricart E: Current status of mesenchymal
stem cell therapy and bone marrow transplantation in IBD. Dig Dis.
30:387–391. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tao H, Han Z, Han ZC and Li Z:
Proangiogenic features of mesenchymal stem cells and their
therapeutic applications. Stem Cells Int. 2016:13147092016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cosenza S, Toupet K, Maumus M,
Luz-Crawford P, Blanc-Brude O, Jorgensen C and Noël D: Mesenchymal
stem cells-derived exosomes are more immunosuppressive than
microparticles in inflammatory arthritis. Theranostics.
8:1399–1410. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Y, Cui J, Wang H, Hezam K, Zhao X,
Huang H, Chen S, Han Z, Han ZC, Guo Z and Li Z: Enhanced
therapeutic effects of MSC-derived extracellular vesicles with an
injectable collagen matrix for experimental acute kidney injury
treatment. Stem Cell Res Ther. 11:1612020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang K, Zhao X, Chen X, Wei Y, Du W, Wang
Y, Liu L, Zhao W, Han Z, Kong D, et al: Enhanced therapeutic
effects of mesenchymal stem cell-derived exosomes with an
injectable hydrogel for hindlimb ischemia treatment. ACS Appl Mater
Interfaces. 10:30081–30091. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qi X, Zhang J, Yuan H, Xu Z, Li Q, Niu X,
Hu B, Wang Y and Li X: Exosomes secreted by human-induced
pluripotent stem cell-derived mesenchymal stem cells repair
critical-sized bone defects through enhanced angiogenesis and
osteogenesis in osteoporotic rats. Int J Biol Sci. 12:836–849.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tao H, Chen X, Cao H, Zheng L, Li Q, Zhang
K, Han Z, Han ZC, Guo Z, Li Z and Wang L: Mesenchymal stem
cell-derived extra-cellular vesicles for corneal wound repair. Stem
Cells Int. 2019:57385102019. View Article : Google Scholar
|
|
16
|
Tkach M and Théry C: Communication by
extracellular vesicles: Where we are and where we need to go. Cell.
164:1226–1232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bei Y, Das S, Rodosthenous RS, Holvoet P,
Vanhaverbeke M, Monteiro MC, Monteiro VVS, Radosinska J, Bartekova
M, Jansen F, et al: Extracellular vesicles in cardiovascular
theranostics. Theranostics. 7:4168–4182. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Du W, Zhang K, Zhang S, Wang R, Nie Y, Tao
H, Han Z, Liang L, Wang D, Liu J, et al: Enhanced proangiogenic
potential of mesenchymal stem cell-derived exosomes stimulated by a
nitric oxide releasing polymer. Biomaterials. 133:70–81. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu B, Zhang X and Li X: Exosomes derived
from mesenchymal stem cells. Int J Mol Sci. 15:4142–4157. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qiu G, Zheng G, Ge M, Wang J, Huang R, Shu
Q and Xu J: Mesenchymal stem cell-derived extracellular vesicles
affect disease outcomes via transfer of microRNAs. Stem Cell Res
Ther. 9:3202018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liao Z, Luo R, Li G, Song Y, Zhan S, Zhao
K, Hua W, Zhang Y, Wu X and Yang C: Exosomes from mesenchymal stem
cells modulate endoplasmic reticulum stress to protect against
nucleus pulposus cell death and ameliorate intervertebral disc
degeneration in vivo. Theranostics. 9:4084–4100. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Xu J, Liu S, Lim M, Zhao S, Cui
K, Zhang K, Wang L, Ji Q, Han Z, et al: Embryonic stem cell-derived
extracellular vesicles enhance the therapeutic effect of
mesenchymal stem cells. Theranostics. 9:6976–6990. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rani S, Ryan AE, Griffin MD and Ritter T:
Mesenchymal stem cell-derived extracellular vesicles: Toward
cell-free therapeutic applications. Mol Ther. 23:812–823. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tan SS, Yin Y, Lee T, Lai RC, Yeo RW,
Zhang B, Choo A and Lim SK: Therapeutic MSC exosomes are derived
from lipid raft microdomains in the plasma membrane. J Extracell
Vesicles. 2:2013.PubMed/NCBI
|
|
25
|
Anderson JD, Johansson HJ, Graham CS,
Vesterlund M, Pham MT, Bramlett CS, Montgomery EN, Mellema MS,
Bardini RL, Contreras Z, et al: Comprehensive proteomic analysis of
mesenchymal stem cell exosomes reveals modulation of angiogenesis
via nuclear factor-kappaB signaling. Stem Cells. 34:601–613. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mao F, Wu Y, Tang X, Kang J, Zhang B, Yan
Y, Qian H, Zhang X and Xu W: Exosomes derived from human umbilical
cord mesenchymal stem cells relieve inflammatory bowel disease in
mice. Biomed Res Int. 2017:53567602017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen G, Ran X, Li B, Li Y, He D, Huang B,
Fu S, Liu J and Wang W: Sodium butyrate inhibits inflammation and
maintains epithelium barrier integrity in a TNBS-induced
inflammatory bowel disease mice model. EBioMedicine. 30:317–325.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang S, Liu Y, Zhang X, Zhu D, Qi X, Cao
X, Fang Y, Che Y, Han ZC, He ZX, et al: Prostaglandin E2
hydrogel improves cutaneous wound healing via M2 macrophages
polarization. Theranostics. 8:5348–5361. 2018. View Article : Google Scholar :
|
|
30
|
Zhang K, Wang C, Wang R, Chen S and Li Z:
Dual bioluminescence imaging of tumor progression and angiogenesis.
J Vis Exp. e597632019.
|
|
31
|
Zhao N, Yue Z, Cui J, Yao Y, Song X, Cui
B, Qi X, Han Z, Han ZC, Guo Z, et al: IGF-1C domain-modified
hydrogel enhances therapeutic potential of mesenchymal stem cells
for hindlimb ischemia. Stem Cell Res Ther. 10:1292019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
33
|
Liang L, Li Z, Ma T, Han Z, Du W, Geng J,
Jia H, Zhao M, Wang J, Zhang B, et al: Transplantation of human
placenta-derived mesenchymal stem cells alleviates critical limb
ischemia in diabetic nude rats. Cell Transplant. 26:45–61. 2017.
View Article : Google Scholar :
|
|
34
|
Li Z and Han ZC: Introduction of perinatal
tissue-derived stem cells. Perinatal Stem Cells: Biology,
Manufacturing and Translational Medicine. Han ZC, Takahashi TA, Han
Z and Li Z: Springer; Singapore, Singapore: pp. 1–7. 2019
|
|
35
|
Jeon YJ, Kim J, Cho JH, Chung HM and Chae
JI: Comparative analysis of human mesenchymal stem cells derived
from bone marrow, placenta, and adipose tissue as sources of cell
therapy. J Cell Biochem. 117:1112–1125. 2016. View Article : Google Scholar
|
|
36
|
Keshtkar S, Azarpira N and Ghahremani MH:
Mesenchymal stem cell-derived extracellular vesicles: Novel
frontiers in regenerative medicine. Stem Cell Res Ther. 9:632018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hong P, Yang H, Wu Y, Li K and Tang Z: The
functions and clinical application potential of exosomes derived
from adipose mesenchymal stem cells: A comprehensive review. Stem
Cell Res Ther. 10:2422019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Neurath MF: Cytokines in inflammatory
bowel disease. Nat Rev Immunol. 14:329–342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Strober W and Fuss IJ: Proinflammatory
cytokines in the pathogenesis of inflammatory bowel diseases.
Gastroenterology. 140:1756–1767. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Guan Q and Zhang J: Recent advances: The
imbalance of cytokines in the pathogenesis of inflammatory bowel
disease. Mediators Inflamm. 2017:48102582017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Brito TV, Neto JP, Prudêncio RS, Batista
JA, Júnior JS, Silva RO, Franco AX, Aragão KS, Soares PM, Souza MH,
et al: Sulfated-polysaccharide fraction extracted from red algae
gracilaria birdiae ameliorates trinitrobenzenesulfonic acid-induced
colitis in rats. J Pharm Pharmacol. 66:1161–1170. 2014.PubMed/NCBI
|
|
42
|
Denson LA, Jurickova I, Karns R, Shaw KA,
Cutler DJ, Okou DT, Dodd A, Quinn K, Mondal K, Aronow BJ, et al:
Clinical and genomic correlates of neutrophil reactive oxygen
species production in pediatric patients with Crohn's disease.
Gastroenterology. 154:2097–2110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tian T, Wang Z and Zhang J:
Pathomechanisms of oxidative stress in inflammatory bowel disease
and potential antioxidant therapies. Oxid Med Cell Longev.
2017:45351942017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hansberry DR, Shah K, Agarwal P and
Agarwal N: Fecal myeloperoxidase as a biomarker for inflammatory
bowel disease. Cureus. 9:e10042017.PubMed/NCBI
|
|
45
|
Souza HS, Tortori CJ, Castelo-Branco MT,
Carvalho AT, Margallo VS, Delgado CF, Dines I and Elia CC:
Apoptosis in the intestinal mucosa of patients with inflammatory
bowel disease: Evidence of altered expression of FasL and perforin
cytotoxic pathways. Int J Colorectal Dis. 20:277–286. 2005.
View Article : Google Scholar
|
|
46
|
Martini E, Krug SM, Siegmund B, Neurath MF
and Becker C: Mend your fences: The epithelial barrier and its
relationship with mucosal immunity in inflammatory bowel disease.
Cell Mol Gastroenterol Hepatol. 4:33–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Peterson LW and Artis D: Intestinal
epithelial cells: Regulators of barrier function and immune
homeostasis. Nat Rev Immunol. 14:141–153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Boltin D, Perets TT, Vilkin A and Niv Y:
Mucin function in inflammatory bowel disease: An update. J Clin
Gastroenterol. 47:106–111. 2013. View Article : Google Scholar
|
|
49
|
Johansson ME, Sjövall H and Hansson GC:
The gastrointestinal mucus system in health and disease. Nat Rev
Gastroenterol Hepatol. 10:352–361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Marchiando AM, Shen L, Graham WV, Edelblum
KL, Duckworth CA, Guan Y, Montrose MH, Turner JR and Watson AJ: The
epithelial barrier is maintained by in vivo tight junction
expansion during pathologic intestinal epithelial shed-ding.
Gastroenterology. 140:1208–1218. e1–e2. 2011. View Article : Google Scholar
|