Introduction
Acute myocardial infarction (AMI) is a key cause of
high mortality in various types of cardiovascular disease (1,2).
Although timely reperfusion therapy to salvage cardiomyocytes
effectively improves patient outcomes, many studies have
demonstrated that reperfusion also results in additional myocardial
damage and dysfunction of tissue, resulting in myocardial
ischemia/reperfusion injury (IRI) (3,4).
The pathophysiological processes underlying development of IRI are
complicated and involve numerous pathogenic factors, including
inflammatory responses and excess accumulation of reactive oxygen
species (ROS) (5). Novel
treatments are required to improve clinical prognosis following AMI
and IRI. Traditional Chinese medicine (TCM) has shown protective
effects against cardiovascular disease, type 2 diabetes mellitus,
and malaria (6). Therefore, the
protective role of TCM for IRI needs further research.
As a TCM, Danshen (Salvia miltiorrhiza) has
been applied to prevent tissue damage and organ dysfunction induced
by cardiovascular disease, including MI and arrhythmia (7). Tanshinone IIA (TSN) is extracted
from the rhizome of Danshen and serves an important role as an
anti-inflammatory, antioxidant, and antithrombotic agent in
vascular smooth muscle cells, improving AMI (8-10).
Additionally, TSN confers protection against IRI in vivo,
mediated by the PI3K/Akt/mTOR signaling pathway. The infarct size
and apoptosis rate of cardiac cells is significantly reduced in an
IRI model following pretreatment with TSN (11). To the best of our knowledge,
previous studies have only focused on biomarkers of apoptosis and
oxidative stress; the underlying molecular mechanisms of TSN and
involvement of ferroptosis are unknown.
Apoptosis has been reported as a primary form of
cell death caused by IRI (12).
In several studies, it has been reported that ferroptosis also
participates in the development of IRI (1,13).
As a novel form of regulated cell death, ferroptosis is an iron-
and lipotoxicity-dependent type of programmed cell death, which is
primarily caused by excessive generation of intracellular ROS
(14,15). Ferroptosis and apoptosis are
activated by the upregulation of voltage-dependent anion channel 1
(VDAC1) in IRI (16,17). As a key component of the outer
membrane of mitochondria, VDAC1 participates in the transportation
of metabolites, nucleotides and ions (16). Moreover, previous studies have
shown that expression of VDAC1 is upregulated and indicates more
severe myocardial damage after anoxia/reoxygenation (A/R) treatment
(18,20).
Therefore, the present study aimed to explore
whether TSN could induce myocardial protection by inhibiting
ferroptosis and apoptosis and whether the protective effects of TSN
are regulated by VDAC1.
Materials and methods
Materials and chemicals
TSN (purity: 98%) was obtained from the National
Institutes for Food and Drug Control (Beijing, China). Adenovirus
pAd/VDAC1 and pAd/negative control (NC) were purchased from Suzhou
GenePharma Co., Ltd. Ferrostatin-1 (Fer-1, ferroptosis inhibitor),
erastin (ferroptosis inducer), Z-Val-Ala-DL-Asp-fluoromethylketone
(Z-VAD, apoptosis inhibitor) and N-acetylcysteine (NAC; ROS
inhibitor) were obtained from MedChemExpress. The primary
antibodies against glutathione peroxidase 4 (GPX4), Bcl-2 and Bax
were obtained from Chengdu Zhengneng Biotechnology Co., Ltd. and
VDAC1, prostaglandin endoperoxide synthase 2 (PTGS2) and β-actin
were obtained from Proteintech Group, Inc. Goat anti-mouse and goat
anti-rabbit secondary antibodies were obtained from Beyotime
Institute of Biotechnology.
H9c2 cardiomyocyte culture and
construction of A/R injury model
Rat H9c2 cardiomyocytes, purchased from Cell
Bank/Stem Cell Bank (Beijing, China), were cultured as a monolayer
in high-glucose Dulbecco's modified Eagle's medium (H-DMEM;
HyClone, Cytiva) with 10% fetal bovine serum (FBS; Gibco, Thermo
Fisher Scientific, Inc.) at 37°C under normal conditions (95%
humidity, 21% O2 and 5% CO2), as described by
Pooja et al (19).
As described in previous studies (20,21), the cell A/R liquid method was
applied to establish an in vitro A/R model. H9c2 cells were
incubated in anoxia medium (CaCl2 1.0, HEPES 20.0, KCl
10.0, MgSO4 1.2, NaCl 98.5,
NaH2PO4 0.9, NaHCO3 6.0 and sodium
lactate 40.0 mM, pH 6.8) and 95% N2 and 5%
CO2 at 37°C for 3 h. Reoxygenation medium
(CaCl2 1.0, glucose 5.5, HEPES 20.0, KCl 5.0,
MgSO4 1.2 mM, NaCl 129.5, NaH2PO4
0.9 and NaHCO3 20.0 mM, pH 7.4) was used to simulate IRI
and cells were incubated under 95% O2 and 5%
CO2 at 37°C for 2 h.
Adenovirus preparation and
transduction
Adenoviral vector CMV-ADV6 (Suzhou GenePharma Co.,
Ltd.) was digested using EcoRI and BamHI. The open
reading frame sequence of rat VDAC1 gene [NCBI (National Center for
Biotechnology Information) reference sequence NM_031353.1]
(22) was amplified using PCR
using Power SYBR Green PCR MasterMix (Thermo Fisher Scientific,
Inc.) and used to establish CMV-ADV6-VDAC1 by linearizing and
inserting into the CMV-ADV6 (Suzhou GenePharma Co., Ltd.). PCR
process involved initial denaturation at 95°C for 10 min, followed
by thermocycling 95°C for 15 sec, and 60°C for 1 min, repeated 40
cycles. To acquire positive clones, the recombinant plasmid was
tranduced into DH5α-competent cells (Tiangen) at 37°C for 16 h and
the positive clone was confirmed by liquid sequencing. Plasmid Midi
Preparation kit (Beijing CW Biotech Co., Ltd.) was used to prepare
the recombinant plasmids following the manufacturer's instructions.
Next, CMV-ADV6-VDAC1 was transduced into 293 cells (ATCC, cat. no.
CRL-1573) using RNAi-mate (Suzhou GenePharma Co., Ltd.) and a 2nd
generation system under 95% O2 and 5% CO2 at
37°C for 6 h (quantity of adenovirus plasmid used for transfection
is 10 μg and the ratio used for adenovirus, packaging and
envelope plasmids is 1:3:4). Finally, the supernatant was collected
after centrifugation at 7,000 × g for 5 min at 4°C following virus
amplification and the generated adenoviral vectors were amplified
until the viral titer reached 1×109 plaque-forming units
(PFU)/ml.
pAd/VDAC1 or pAd/NC were transduced into H9c2
cardiomyocytes cultured in fresh H-DMEM supplemented with 10% FBS
(multiplicity of infection, 100) under 95% O2 and 5%
CO2 at 37°C for 48 h. H9c2 cells not transduced with
adenovirus were used as Control. Then, subsequent experiments were
conducted.
H9c2 cardiomyocyte treatment
H9c2 cells were randomly divided into the following
groups: i) Control, H9c2 cardiomyocytes cultured under normal
conditions; ii) A/R, H9c2 cardiomyocytes exposed to A/R; iii) TSN,
H9c2 cardiomyocytes pretreated with TSN (1, 2, 4, 8, 16 and 32
μM) at 37°C for 48 h; iv) TSN + A/R, H9c2 cardiomyocytes
pretreated with TSN (1, 2, 4, 8, 16 and 32 μM) at 37°C for
48 h before A/R injury; v) 8 μM TSN + A/R, H9c2
cardiomyocytes pretreated with 8 μM TSN at 37°C for 48 h
before A/R injury; vi) Fer-1 + A/R, H9c2 cardiomyocytes pretreated
with 10 μM Fer-1 at 37°C for 2 h before A/R; vii) erastin,
H9c2 cardiomyocytes treated with 10 μM Erastin at 37°C for
24 h; viii) erastin + TSN, H9c2 cardiomyocytes pretreated with 8
μM TSN for 48 h before erastin treatment; ix) Z-VAD + A/R,
H9c2 cells pretreated with 40 μM Z-VAD at 37°C for 24 h
before A/R; x) NAC + A/R, H9c2 cardiomyocytes pretreated with 6 mM
NAC at 37°C for 6 h before A/R injury; xi) pAd/VDAC1 + TSN + A/R,
H9c2 cardiomyocytes transfected with pAd/VDAC1 and pretreated with
8 μM TSN for 48 h before A/R and xii) pAd/NC + TSN + A/R,
H9c2 cardiomyocytes transfected with pAd/NC and pretreated with 8
μM TSN for 48 h before A/R.
Assessment of cell viability
Cell viability was determined by colorimetric assay
using Cell Counting Kit-8 assay (CCK-8; GlpBio Technology).
Briefly, H9c2 cardiomyocytes were seeded in 96-well plates at a
density of 1×104 cells/well, and incubated with 10
μl CCK-8 reagent/100 μl H-DMEM (HyClone, Cytiva)
added with 10% FBS (Gibco, Thermo Fisher Scientific, Inc.) for 1 h
at 37°C. Finally, the absorbance was measured at 450 nm using a
microplate reader (Thermo Fisher Scientific, Inc.).
Assessment of cytotoxicity
Following different treatments, the cell culture
medium was harvested, and the lactate dehydrogenase (LDH) levels
were determined by LDH assay kits (Beyotime Institute of
Biotechnology, cat. no. #C0016) following the manufacturer's
instructions. Furthermore, cytotoxicity was calculated by dividing
absorbance of the experimental wells by 100%.
Measurement of malondialdehyde (MDA),
total iron and glutathione (GSH)/glutathione disulfide (GSSG)
levels
Following treatment, H9c2 cardiomyocytes from each
group were digested with 0.25% trypsin, washed twice with
phosphate-buffered saline (PBS) and lysed by ultrasound (20 kHz for
3 min) to obtain cell homogenates. The supernatant was obtained
after centrifugation at 12,000 × g for 15 min at 4°C. The
supernatant was used to determine the intracellular levels of MDA,
total iron, GSH and glutathione disulfide (GSSG) and the GSH/GSSG
ratio using MDA assay kit (Beyotime Institute of Biotechnology,
cat. no. #S0131S), total iron assay kit [Applygen (https://www.applygen.com/), cat. no. #E1042] and GSH
and GSSG assay kits (Beyotime Institute of Biotechnology, cat. no.
#S0053), according to the manufacturer's instructions.
Caspase-3 activity assay
Following treatment, the activity of caspase-3 was
determined using caspase-3 activity assay kit (Beyotime Institute
of Biotechnology, cat. no. #C1115) according to the manufacturer's
instruction. In brief, 40 reaction buffer, 50 cell homogenate
extracted from each group and 10 μl caspase-3 substrates
were added to a 96-well plate and incubated for 2 h at 37°C. Next,
absorbance was measured using a microplate reader at 405 nm.
Finally, the Bradford method was used to assess the protein
concentration in each group (23).
Determination of intracellular ROS
The levels of intracellular ROS were determined by
an Olympus IX 73 microscope (Olympus Corporation) using DCFH-DA
(Beyotime Institute of Biotechnology, cat. no. S0033S). DCFH-DA
enters the cell and can be lysed into DCFH by esterases that cannot
cross the cell membrane and be oxidized by intracellular ROS into
fluorescent DCF (24). Briefly,
DCFH-DA (10 μM) dye was added to cells and incubated for 20
min in the dark at 37°C. Next, levels of intracellular ROS were
detected using the fluorescence microscope (magnification,
×200).
Assessment of intracellular ferrous iron
levels
The intracellular Fe2+ level was measured
using FerroOrange (Dojindo Laboratories, Inc.) based on the
manufacturer's instructions. Briefly, treated H9c2 cardiomyocytes
were treated with 1 μM FerroOrange and 0.5 μg/ml
Hoechst 33342 (Beyotime Institute of Biotechnology) at 37°C for 30
min in the dark. Excess FerroOrange and Hoechst 33342 were removed
by washing twice with PBS. Finally, the fluorescence microscope
(magnification, ×200) was used to assess the levels of ferrous
iron.
Mitochondrial membrane potential (MMP)
assay
MMP was determined using the JC-1 MMP Detection kit
(BestBio) following the manufacturer's instructions. Briefly,
following treatments, H9c2 cells were collected and incubated with
JC-1 and quenching agent for 30 min at 37°C in the dark, washed
twice with PBS and the levels of MMP were measured using a Cytomics
FC500 flow cytometer [Beckman Coulter, Inc.; 530/580 and 485/530
nm] (analyte detector is Annexin V and PI; the analyte reporter is
FITC and PE). The red (JC-1 aggregates) to green (JC-1 monomers)
fluorescence intensity ratio of the cells reflects the levels of
MMP. NovoExpress (v.6.2; Agilent Technologies, Inc.) was used to
analyze the flow cytometry data.
Mitochondrial permeability transition
pore (mPTP) opening assay
mPTPs were assessed by BBcellProbe M61 mPTP
Detection kit (BestBio). In brief, following treatment, H9c2
cardiomyocytes were collected and cultured with BBcellProbe M61 and
a quenching agent for 15 min at 37°C in the dark, washed twice with
PBS and the levels of mPTP were determined using a Cytomics FC500
flow cytometer [excitation (Ex), 488; emission (Em), 558 nm] (The
analyte detector is Annexin V; the analyte reporter is FITC). The
green fluorescence intensity of BBcellProbe M61 probe [(Ex), 495
nm; (Em), 515 nm] labelled cells represented the levels of mPTP
opening. NovoExpress (v.6.2; Agilent Technologies, Inc.) was used
to analyze the flow cytometry data.
Apoptosis assay
Cell apoptosis was assayed using the Annexin V-FITC
Apoptosis Detection kit (BestBio). In brief, following treatment,
H9c2 cardiomyocytes were collected and resuspended in 1X Annexin V
binding buffer at a cell density of 1×106 cells/ml. The
cell suspension was incubated with 5 Annexin V-FITC and 10
μl PI at 4°C for 20 min in the dark and detected using a
Cytomics FC500 flow cytometer (Ex, 488 nm; Em, 578 nm). Q1 quadrant
represented necrotic cells and Q4 quadrant represented live cells.
The total apoptosis rate was calculated as the sum of Q2 (late
apoptotic cells) and Q3 (early apoptotic cells). NovoExpress
(v.6.2; Agilent Technologies, Inc.) was used to analyze data.
Lipid ROS assay
The levels of intracellular lipid ROS were measured
using a Cytomics FC500 flow cytometer (analyte detector is Annexin
V and PI; the analyte reporter is FITC and PE) with
C11-BODIPY581/591 (GlpBio Technology). Briefly, treated H9c2
cardiomyocytes were gathered and incubated with 10 μM
C11-BODIPY581/591 in the dark for 30 min at 37°C and washed twice
with PBS. The cells were resuspended in PBS containing 10% FBS. The
shift of fluorescence emission peak from ~590 to ~510 nm reflects
the increase in lipid ROS levels. NovoExpress (v.6.2; Agilent
Technologies, Inc.) was used to analyze the flow cytometry
data.
Mitochondrial ultrastructural
assessment
Following the different treatments, H9c2 cells were
rapidly gathered and incubated in 2% glutaraldehyde for 2 h. Then,
the cells were observed by transmission electron microscopy after
washing, dehydration, embedding in Epon 812, sectioning (60 nm),
and stained with 2% uranyl acetate and 2.6% lead citrate for 8 min
at 37°C. Additionally, the Flameng score method was applied to
determine the mitochondrial ultrastructural injury (25).
Western blot analysis
Following treatment, total cell lysates obtained
using radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology) with 1% phenylmethylsulfonyl fluoride,
incubated at 4°C for 15 min and the protein concentration was
quantified using bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). An equal amount of total protein (40
μg/lane) in each sample was separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, transferred to
polyvinylidene fluoride membranes and blocked with 5% non-fat dry
milk in Tris-buffered saline with 0.1% Tween-20 buffer at room
temperature for 2 h. Subsequently, membranes were incubated with
primary antibodies against PTGS2 (ProteinTech Group, Inc.; cat. no.
#12375-1-AP; 1:1,000), VDAC1 (ProteinTech Group, Inc.; cat. no.
#55259-1-AP; 1:1,000), GPX4 (ZENBIO; cat. no. #381958; 1:1,000),
Bcl-2 (ZENBIO; cat. no. #250412; 1:1,000), Bax (ZENBIO; cat. no.
#380709; 1:1,000) and β-actin (ProteinTech Group, Inc.; cat. no.
#20536-1-AP; 1:1,000) at 4°C overnight in a shaker. Then, the PVDF
membranes were washed three times for 10 min and incubated with
secondary HRP-conjugated antibodies (Beyotime Institute of
Biotechnology, cat. no. #A0208; 1:1,000) for 2 h at 25°C. Finally,
an ultra-high-sensitivity ECL kit (Beyotime Institute of
Biotechnology) was used to visualize protein bands. The signal
intensities of bands were quantified using ImageJ 1.8.0 (National
Institutes of Health).
Molecular docking simulation
The molecular docking simulation was performed using
Discovery Studio 4.5 software (26). The three-dimensional structure of
VDAC1 for molecular docking was obtained from AlphaFold database
(UniProt ID: P21796) (27) and
the structure of TSN was obtained from PubChem (PubChem CID:
164676) (28). TSN was docked
into VDAC1 using LibDock mode in Discovery Studio 4.5 software with
parameters selected by a default procedure. Thus, molecular docking
scores was calculated using LibDock custom scoring function to
evaluate the binding affinity between TSN and VDAC1.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 8.0 (Dotmatics). All experiments were repeated 3 times and
the data are expressed as the mean ± standard deviation. Three or
more groups were compared by one-way analysis of variance followed
by Dunnett's or Tukey's post hoc multiple comparisons test. P≤0.05
was considered to indicate a statistically significant
difference.
Results
TSN protects H9c2 cardiomyocytes against
A/R injury
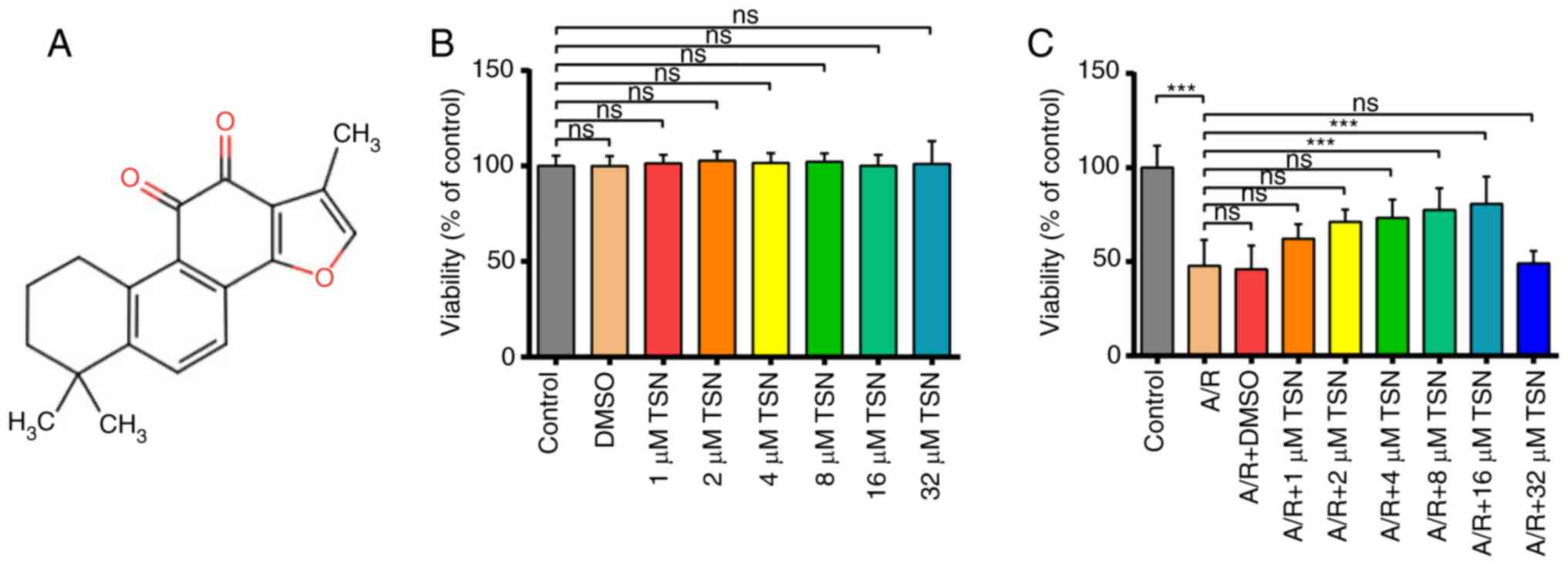
The chemical structure of TSN is presented in
Fig. 1A. CCK-8 assay was used to
measure the viability of H9c2 cardiomyocytes following pretreatment
with different concentrations of TSN (0, 1, 2, 4, 8, 16 and 32
μM) and A/R injury. After culturing H9c2 cardiomyocytes with
TSN at different concentrations, there was no difference in the
viability of H9c2 cells compared with Control group (Fig. 1B). Additionally, A/R injury
significantly decreased the cell viability, which was increased
significantly after treatment with 8 and 16 μM TSN before
A/R. Based on the principle of drug dosage selection (29), 16 μM was considered the
limiting level for safe drug concentrations. By contrast, 8
μM TSN optimally enhanced viability of A/R-induced H9c2
cardiomyocytes (Fig. 1C).
TSN alleviates ferroptosis of A/R-induced
H9c2 cardiomyocytes via downregulation of VDAC1
TSN at a concentration of 8 μM was selected
for subsequent experiments. Pretreatment with TSN and Fer-1 before
A/R injury significantly increased the cell viability that was
decreased by A/R injury and decreased levels of LDH activity that
were increased significantly by A/R injury, suggesting pretreatment
with 8 μM TSN and 10 μM Fer-1 could effectively
protect H9c2 cardiomyocytes against A/R injury (Fig. 2A and B). It has previously been
reported that iron overload and abnormal lipid metabolism are key
features of ferroptosis (30,31). The total iron content and products
of lipid peroxidation such as MDA were significantly increased in
A/R group and reduced by TSN and Fer-1 pretreatment (Fig. 2C and D). In addition, as an
important part of the non-enzymatic antioxidant system, GSH is used
by glutathione peroxidase 4 (GPX4) to remove excess lipid
metabolites (32). A/R injury
significantly increased the levels of GSSG and decreased the levels
of GSH and the GSH/GSSG ratio in H9c2 cardiomyocytes, while
pretreatment with TSN and Fer-1 reversed these changes (Fig. 2E). Accumulation of excess ROS not
only damages lipids and triggers abnormal lipid metabolism leading
to ferroptosis but is also the critical outcome of ferroptosis
(5). Pretreatment with TSN and
Fer-1 significantly attenuated the increase in intracellular ROS
induced by A/R (Fig. 2F).
Expression of PTGS2 and GPX4, positive/negative molecular markers
of ferroptosis, was measured in cell lysate (33). Following A/R treatment, PTGS2
expression was significantly increased and GPX4 expression was
decreased; these effects were reversed by pretreatment with TSN and
Fer-1 (Fig. 2G and H).
Furthermore, VDAC1 expression in H9c2 cardiomyocytes in A/R group
increased significantly compared with that in control group and was
decreased in H9c2 cardiomyocytes pretreated with TSN and Fer-1
(Fig. 2G and H). Together, the
results showed that VDAC1 was involved in ferroptosis during A/R
injury and may mediate the protective effect of TSN against
ferroptosis in A/R injury.
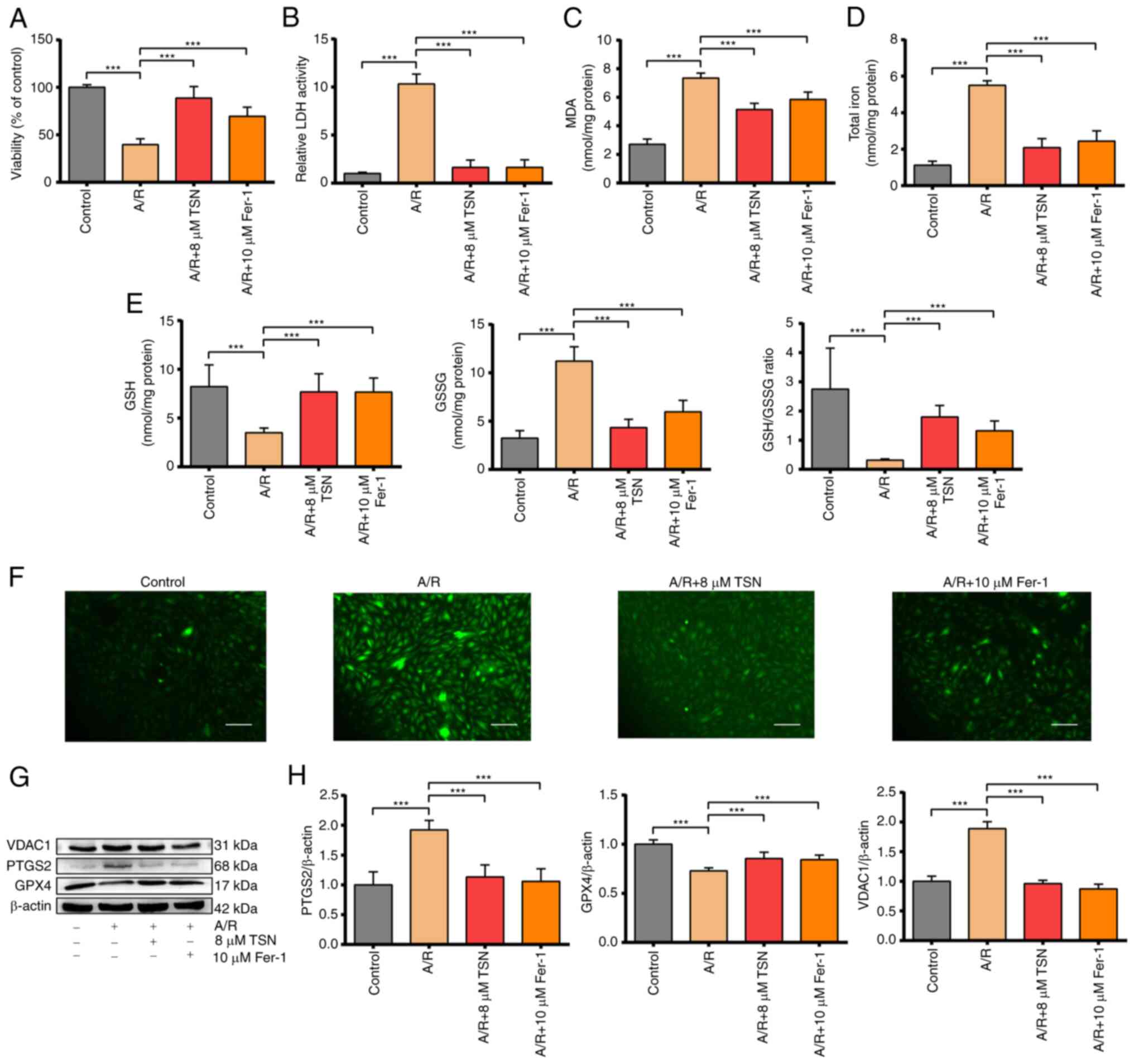 | Figure 2TSN alleviates ferroptosis of
A/R-induced H9c2 cardiomyocytes via downregulation of VDAC1. (A)
Cell Counting Kit-8 detection of viability in A/R-induced cells
following TSN or Fer-1 pretreatment. (B) LDH, (C) MDA, (D) total
iron, (E) GSH, GSSG, GSH/GSSG and (F) ROS were determined by
quantitative kits in A/R-induced cells following TSN or Fer-1
treatment (magnification, ×200; scale bar, 50 μm). (G)
Expression of (H) ferroptosis-related proteins and VDAC1 were
detected by western blot analysis in A/R-induced cells following
TSN or Fer-1 pretreatment. Data are expressed as the mean ± SD
(n=3). ***P<0.05. TSN, Tanshinone IIA; A/R,
Anoxia/reoxygenation; VDAC1, Voltage-dependent anion channel 1;
Fer-1, ferrostatin-1; LDH, lactate dehydrogenase; MDA,
malondialdehyde; GSH, Glutathione; GSSG, Glutathione disulfide;
ROS, reactive oxygen species; PTGS2, Prostaglandin endoperoxide
synthase 2; GPX, Glutathione peroxidase 4. |
TSN attenuates erastin-induced
ferroptosis and inhibits apoptosis of A/R-induced H9c2 cells
To confirm that TSN alleviates A/R injury by
inhibiting ferroptosis and apoptosis, the present study explored
the protective role of TSN on erastin-induced H9c2 cell injury.
Viability of erastin-induced H9c2 cardiomyocytes was significantly
decreased and levels of lipid ROS, ferrous iron, MDA and total iron
were significantly increased in erastin group (Fig. 3A-F). Pretreatment with TSN
effectively reversed the harmful effects of erastin (Fig. 3A-F). Based on the main biological
signs (lipid peroxidation and iron overload) of ferroptosis, TSN
served a biological role in attenuating ferroptosis and TSN
alleviated A/R injury by inhibiting ferroptosis (Figs. 2 and 3A-F). Excess ROS generation has been
reported to play a key role in the pathological process of IRI and
triggers multiple forms of regulated cell death, including
apoptosis and ferroptosis (34,35). Therefore, the protective effect of
NAC on A/R injury was investigated. A/R treatment significantly
reduced the cell viability, while NAC or TSN pretreatment
effectively abolished the damaging effects of A/R (Fig. 3G).
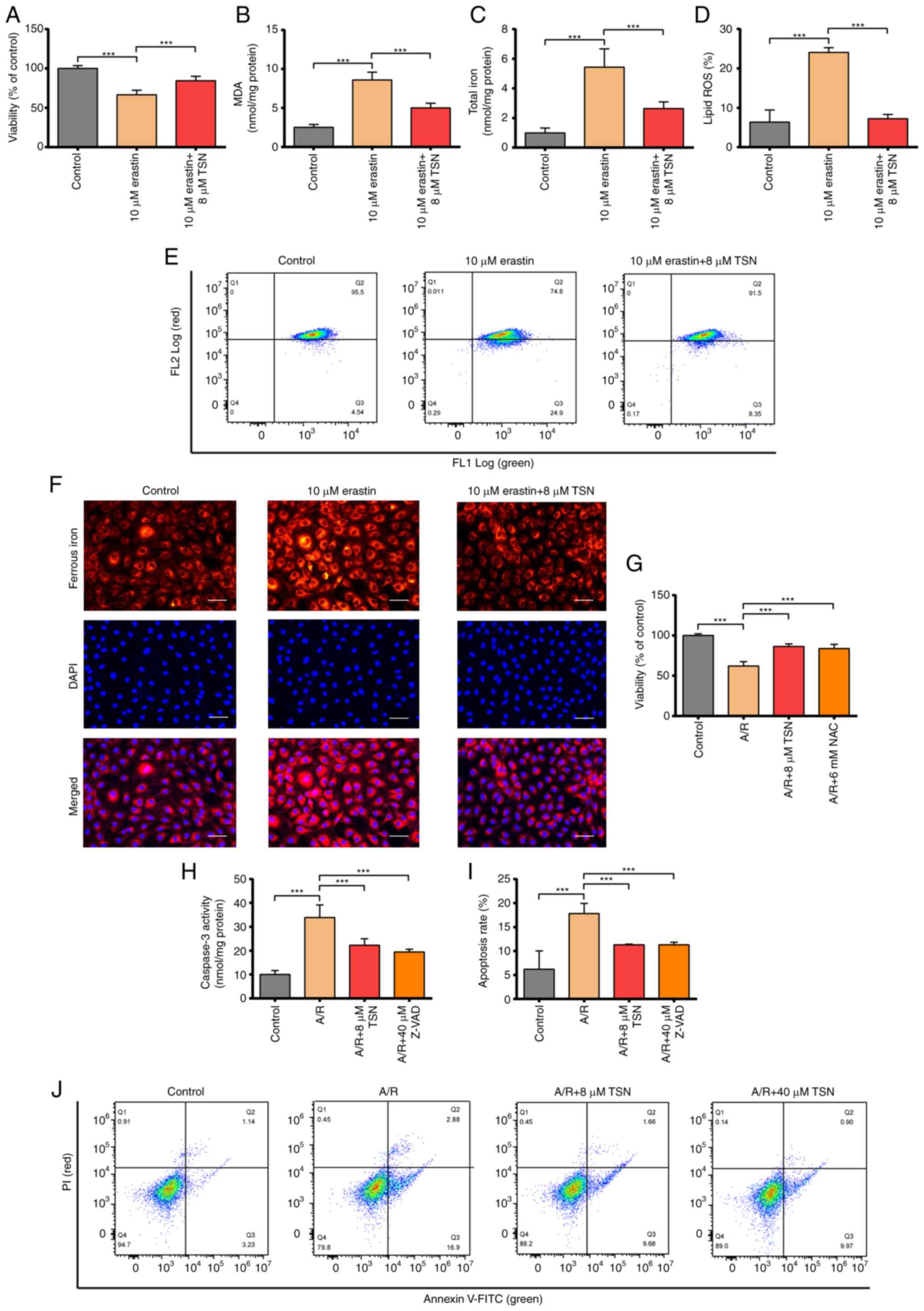 | Figure 3TSN attenuates ferroptosis of
erastin-induced H9c2 cells and inhibits apoptosis of A/R-induced
H9c2 cardiomyocytes. (A) CCK-8 detection of viability in H9c2 cells
following erastin or TSN treatment. (B) MDA, (C) total iron, (D)
lipid ROS were determined by quantitative kits in H9c2 cells after
erastin or TSN treatment. Quantitative analysis for (E) the levels
of intracellular lipid ROS. (F) Ferrous iron was measured by
quantitative kits in H9c2 cells after erastin or TSN treatment
(magnification, ×200; scale bar, 50 μm). (G) CCK-8 detected
viability in A/R-induced cells after TSN or NAC pretreatment. (H)
Caspase-3 activity was measured using a Caspase-3 quantitative kit
in A/R-induced cells following TSN or Z-VAD treatment. (I)
Apoptotic rate was (J) measured by Annexin V-FITC/PI detected by
flow cytometry. Data are expressed as the mean ± SD (n=3).
***P<0.05. TSN, tanshinone IIA; A/R,
Anoxia/reoxygenation; CCK, Cell Counting Kit; MDA, malondialdehyde;
ROS, reactive oxygen species; NAC, N-acetylcysteine; Z-VAD,
Z-Val-Ala-DL-Asp-fluoromethylketone. |
To validate that the protective effects of TSN on
A/R-induced H9c2 cell injury were partly due to the inhibition of
apoptosis, the present study elucidated the protective effect of
TSN on A/R-induced apoptosis. TSN and Z-VAD pretreatment
significantly inhibited apoptosis of H9c2 cardiomyocytes and
significantly decreased the increased caspase3 activity in H9c2
cardiomyocytes caused by A/R (Fig.
3H-J).
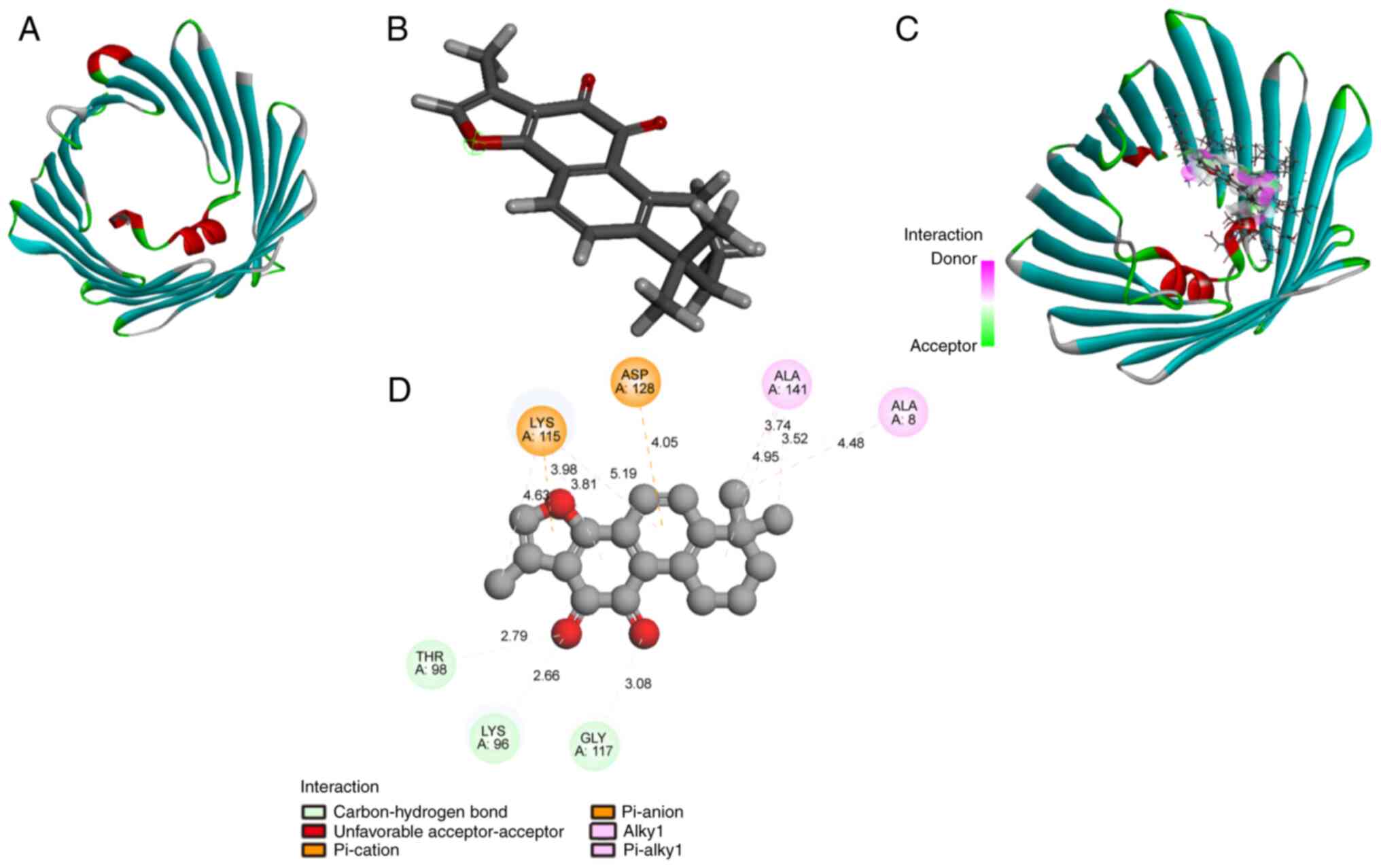
TSN binds to VDAC1
Molecular docking is widely used to assay potential
bioactivity of compounds (36,37). Computational modeling was
performed to investigate the direct interaction between TSN and
VDAC1. The structures of TSN and VDAC1 are shown in Fig. 4A and B, respectively. Molecular
docking of TSN-VDAC1 was performed using LibDock module of
Discovery Studio 4.5 software. TSN strongly bound to residues
Thr-98, Lys-96, and Gly-117 by hydrogen bonds (Fig. 4C and D) with a high combination
score (93.0347). Therefore, VDAC1 was a possible potential target
of TSN.
TSN inhibits ferroptosis of A/R-induced
H9c2 cardiomyocytes by downregulating VDAC1
VDAC1 is an important ferroptosis regulator in IRI
(16). Therefore, the present
study aimed to verify the role of TSN in the modulation of the
VDAC1 levels. After confirming successful transduction of pAd/VDAC1
and pAd/NC into H9c2 cells (Fig.
S1). A/R significantly decreased the cell viability and
increased the level of LDH activity, while pretreatment with TSN
inhibited the injury induced by A/R (Fig. 5A and B). However, the protective
effect of TSN was significantly eliminated by pAd/VDAC1, whereas
pAd/NC could not abolish the protective effect of TSN. Thus, TSN
might regulate VDAC1 to protect H9c2 cardiomyocytes against A/R
injury. It was hypothesized that pretreatment with TSN could
inhibit ferroptosis associated with IRI by downregulating VDAC1.
Levels of MDA, total iron, intracellular ROS, GSH and GSSG were
determined in H9c2 cardiomyocytes. Compared with the control group,
the levels of MDA, total iron, intracellular ROS and GSSG were
significantly increased in H9c2 cells after A/R injury and GSH and
the GSH/GSSG ratio were significantly decreased. However,
pretreatment with TSN could reverse the effects of A/R and
pAd/VDAC1 prevented this reversion (Fig. 5C-F).
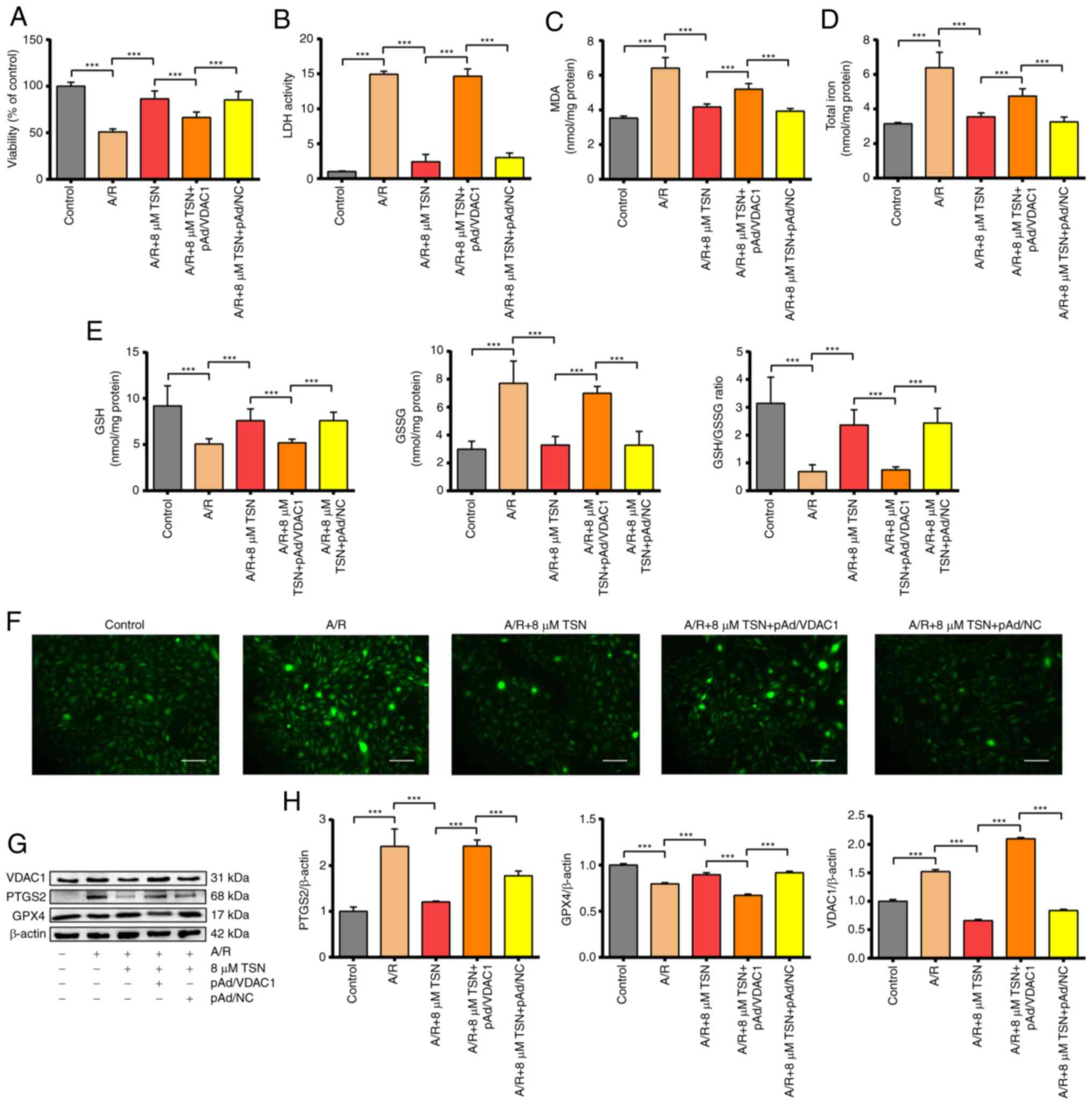 | Figure 5TSN inhibits ferroptosis of
A/R-induced H9c2 cardiomyocytes by downregulating VDAC1. (A) Cell
Counting Kit-8 detection of viability in A/R-induced cells after
TSN, pAd/VDAC1 and pAd/NC pretreatment. (B) LDH, (C) MDA, (D) total
iron, (E) GSH, GSSG, GSH/GSSG and (F) ROS were determined by
quantitative kits in A/R-induced cells following TSN, pAd/VDAC1 and
pAd/NC treatment (magnification, x200; scale bar, 50 μm).
(G) Expression of (H) ferroptosis-associated proteins and VDAC1
were detected by western blot analysis in A/R-induced cells after
TSN, pAd/VDAC1 and pAd/NC pretreatment. Data are expressed as the
mean ± SD (n=3). ***P<0.05. TSN, tanshinone IIA; A/R,
anoxia/reoxygenation; VDAC1, voltage-dependent anion channel 1; NC,
negative control; LDH, lactate dehydrogenase; MDA, malondialdehyde;
GSH, Glutathione; GSSG, Glutathione disulfide; ROS, reactive oxygen
species; PTGS2, Prostaglandin endoperoxide synthase 2; GPX4,
Glutathione peroxidase 4. |
In addition, ferroptosis-associated proteins (PTGS2
and GPX4) and VDAC1 were also determined in vitro using
western blot analysis; A/R injury remarkably increased the level of
PTGS2 and reduced the level of GPX4 compared with the control.
Furthermore, pretreatment with TSN obviously inhibited the effects
of A/R injury. In addition, pAd/VDAC1 significantly increased the
protein levels of VDAC1. Pretreatment with TSN and pAd/VDAC1
significantly increased levels of PTGS2 and decreased GPX4 compared
with pretreatment with TSN (Fig. 5G
and H). TSN effectively inhibited ferroptosis in IRI mediated
by VDAC1 and pAd/VDAC1 significantly prevented this inhibition.
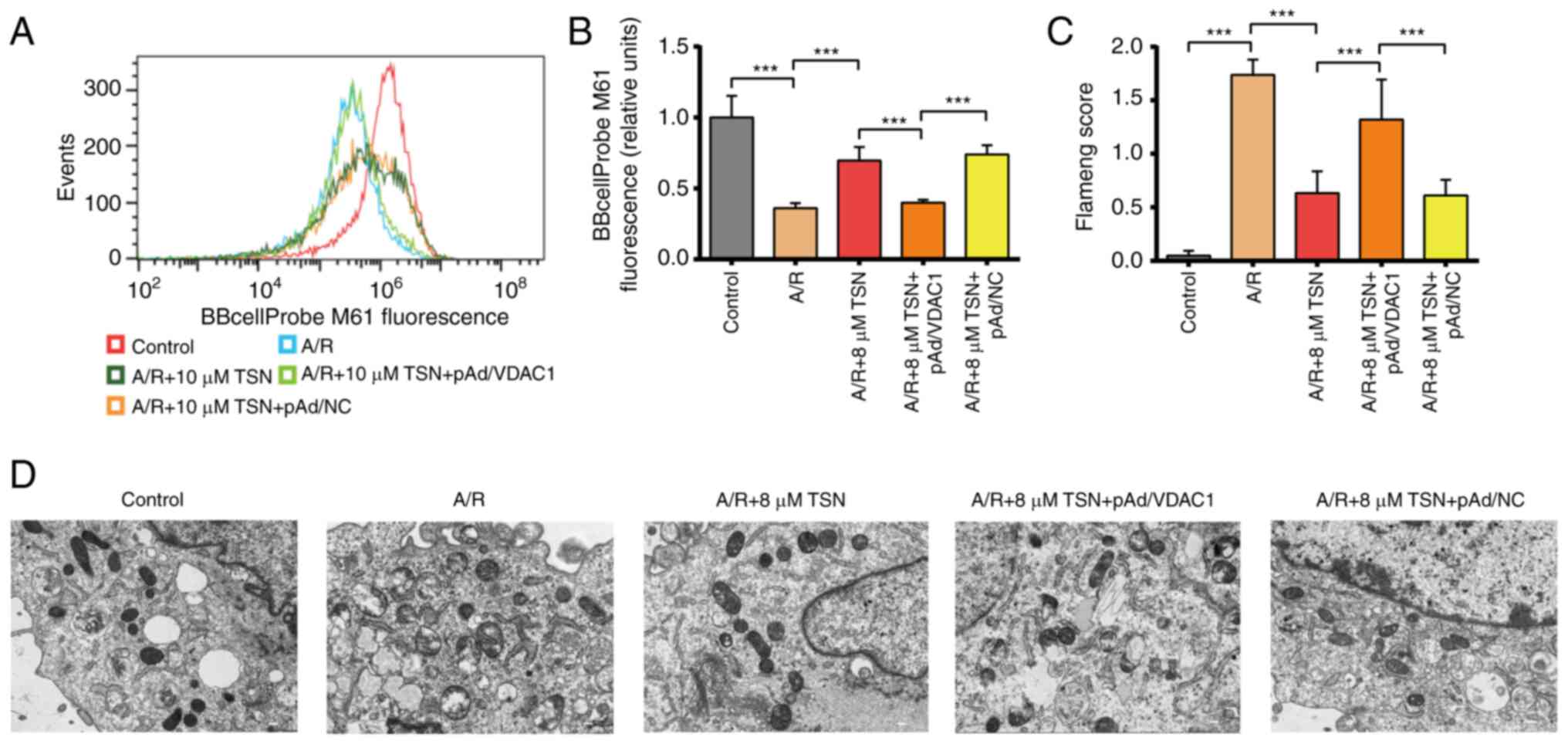
Previous studies have demonstrated that VDAC1 is key
for mPTP formation (16,17). Based on downregulated expression
of VDAC1, the present study aimed to assess mitochondrial
functional and morphological changes in TSN-treated H9c2
cardiomyocytes. TSN or pAd/NC pretreatment significantly decreased
mPTP opening in H9c2 cardiomyocytes following A/R injury;
pretreatment with pAd/VDAC1 reversed this effect (Fig. 6A and B). TEM indicated that the
mitochondrial structure in A/R-treated H9c2 cells was notably
distorted with fewer cristae, while the Flameng scores were
significantly increased; these A/R-induced effects were prevented
by TSN pretreatment, which was reversed by pAd/VDAC1 (Fig. 6C and D).
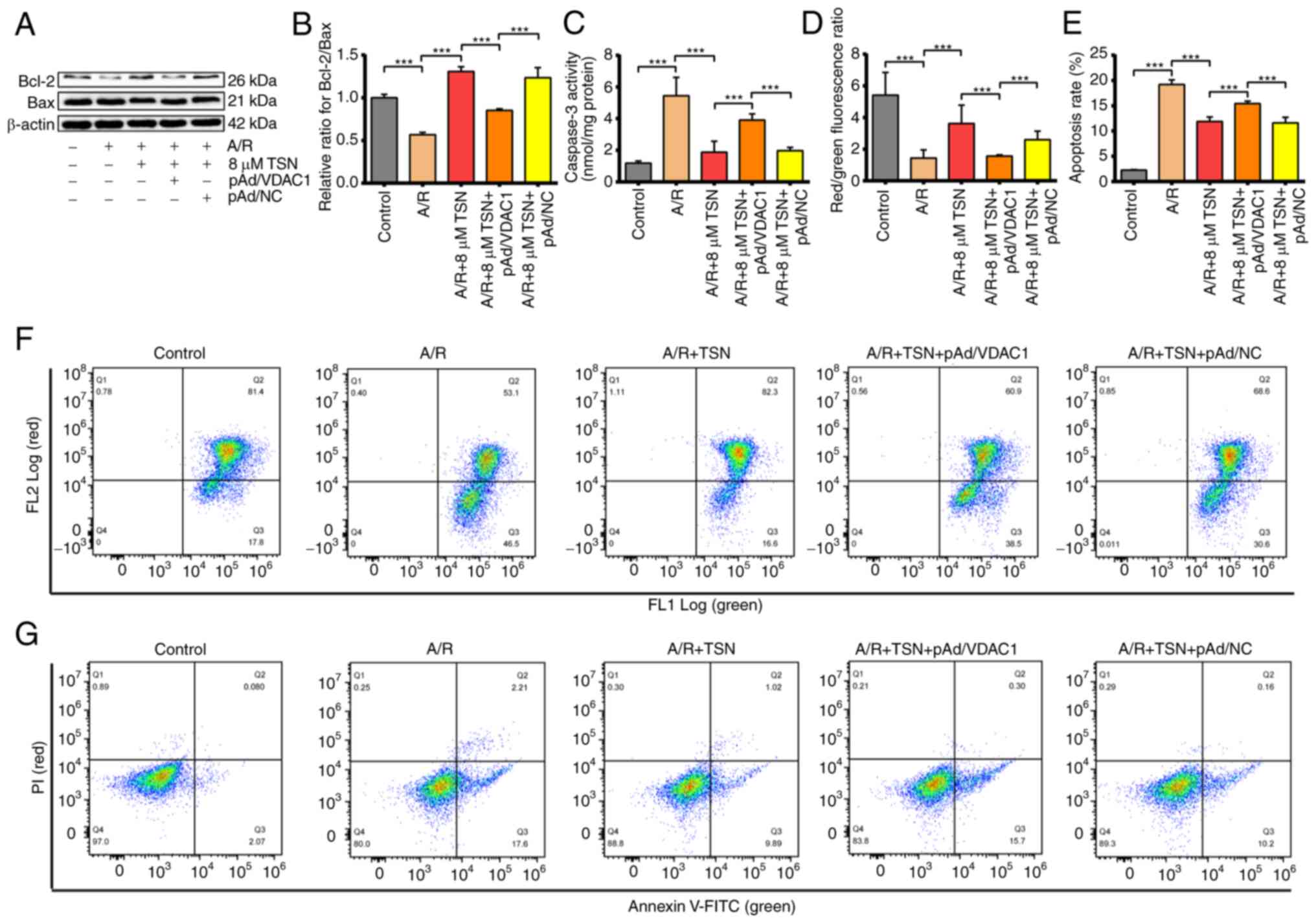
TSN inhibits apoptosis of A/R-induced
H9c2 cardiomyocytes by downregulating
VDAC1 Previous studies have reported that
apoptosis is a key form of cell death involved in IRI (2,38).
Upon investigating the association between the protective effects
of TSN on A/R injury and VDAC1, the present study demonstrated that
protein levels of Bcl-2 and Bax, molecular markers of apoptosis
(39), were notably decreased and
significantly increased, respectively. Thus, Bcl-2/Bax ratio was
significantly decreased in A/R-stimulated H9c2 cardiomyocytes
compared with the control. However, pretreatment with TSN
efficiently increased the Bcl-2/Bax ratio in H9c2 cardiomyocytes
following A/R injury (Fig. 7A and
B). Moreover, overexpression of VDAC1 significantly prevented
the effects of TSN (Fig. 7A and
B). Additionally, TSN suppressed the increased caspase-3
activity in H9c2 cardiomyocytes caused by A/R injury, which was
reversed by pAd/VDAC1 (Fig.
7C).
Furthermore, to confirm that TSN inhibited apoptosis
of A/R-induced H9c2 cardiomyocytes by adjusting VDAC1 (Fig. 8), MMP was determined by flow
cytometry. Changes in MMP are characteristic of early apoptosis,
reflect mitochondrial viability and can be detected by JC-1. In
live cells with high levels of MMP, JC-1 accumulates in the
mitochondrial matrix and produces red fluorescence, while in
apoptotic and dead cells with low levels of MMP, JC-1 cannot
aggregate in the mitochondrial matrix and only emits green
fluorescence. Thus, red/green fluorescence ratio is used to assay
loss of MMP (40). MMP levels
were significantly increased in the A/R group compared with those
in the control. Levels of MMP were significantly downregulated,
whereas pretreatment with TSN significantly increased levels of MMP
compared with A/R group. Furthermore, pAd/VDAC1 significantly
prevented the protective effect of TSN on the A/R-induced decrease
in MMP (Fig. 7D and F).
Consistently, apoptotic rate was significantly induced in A/R group
compared with that in control group and pretreatment with TSN
significantly reversed the effects on apoptotic rate compared with
the A/R group, whereas pAd/VDAC1 pretreatment abolished the
protective effects of TSN (Fig. 7E
and G).
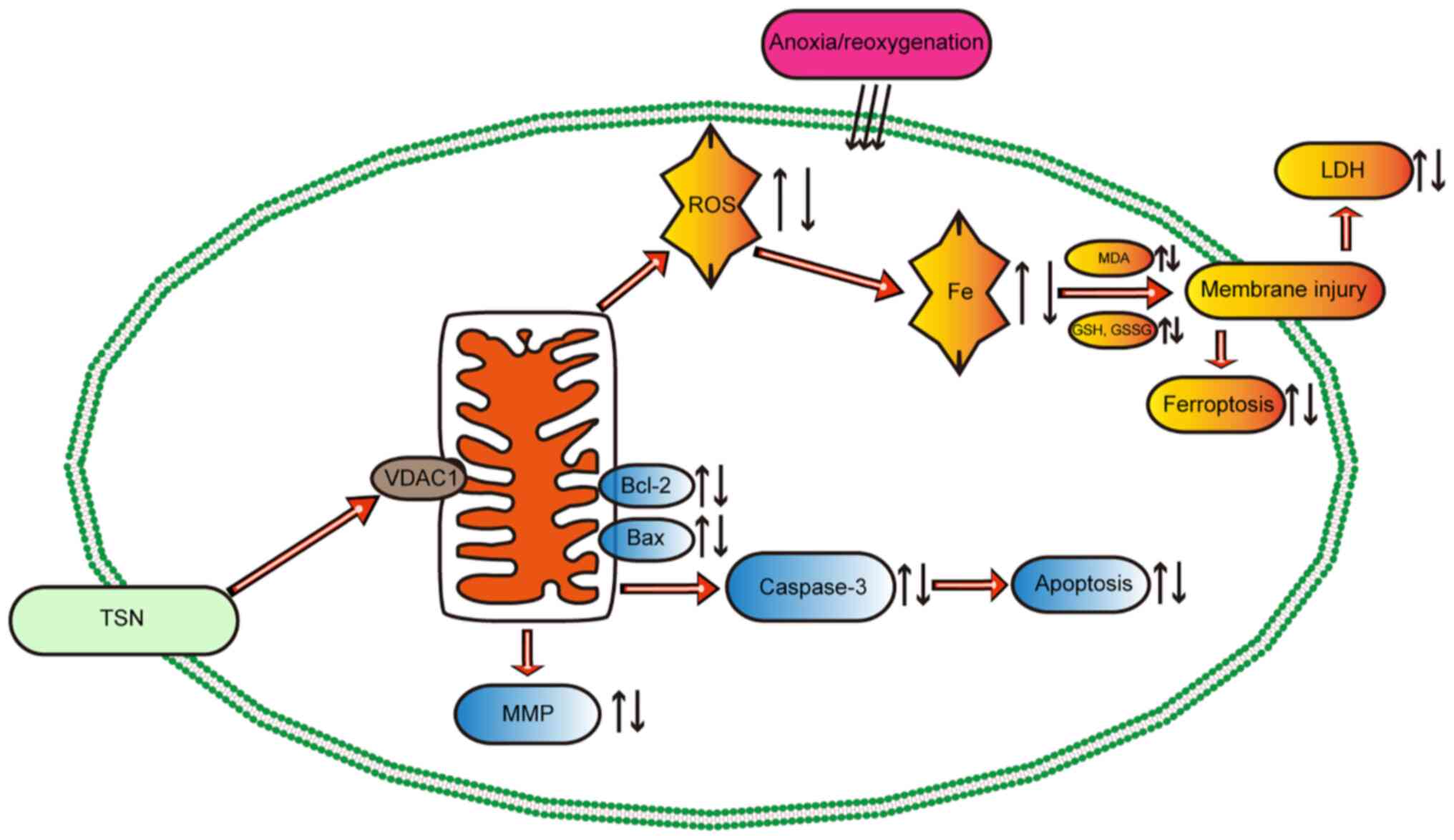 | Figure 8Potential mechanism of TSN in
myocardial ischemia/reperfusion injury. TSN pretreatment
upregulates the expression of VDAC1, thereby decreasing the
accumulation of ROS and iron and abnormal lipid metabolism,
maintaining mitochondrial function and protecting the myocardium
against anoxia/reoxygenation-induced ferroptosis and apoptosis.
TSN, tanshinone IIA; VDAC1, Voltage-dependent anion channel 1; ROS,
reactive oxygen species; MDA, malondialdehyde; GSH, Glutathione;
GSSG, Glutathione disulfide; LDH, lactate dehydrogenase; MMP,
Mitochondrial membrane potential. |
These results showed that TSN attenuated apoptosis
in A/R injury by downregulating VDAC1 (Fig. 8).
Discussion
Although reoxygenation or reperfusion is an
effective approach for AMI, it can lead to further serious tissue
damage that accounts for expansion of the infarcted area and
increased mortality in patients with AMI (1). Such injury is known as IRI (41). Previous studies have reported that
the pathogenesis of IRI involves excessive ROS generation and
inflammatory responses (42,43). The underlying molecular mechanisms
and pathways of IRI are complicated and associated with multiple
forms of regulated cell death, including pyroptosis, necroptosis,
ferroptosis and apoptosis (44,45). Elucidating the underlying
mechanisms is key in identifying therapeutic targets and drugs for
IRI (46). The present study
showed that in A/R-induced H9c2 cardiomyocytes, LDH activity
increased and cell viability decreased significantly, suggesting
that establishment of an in vitro model of IRI was
successful.
Since the 'nutritional preconditioning' hypothesis
was proposed, it has been the focus of many studies, which refers
to the pretreatment of cells with nutrients before injuries
(16,47). Pharmacological pretreatment is the
most common approach to improve the outcome of numerous types of
cardiovascular disease (48). For
example, Puerarin preconditioning protects cardiomyocytes from
sepsis-induced injury by inhibiting ferroptosis and apoptosis
(49). It is hypothesized that
TSN is also a candidate phytochemical. TSN, a bioactive natural
product primarily extracted from the root of Danshen, is considered
a nutriment and is used to produce soup or tea (9). However, it has also been used as a
medicine in the US and Europe (10). Danshen has been used to treat
stroke, skin disorders, spasms, and other ailments in Asian
countries for hundreds of years (50,51). Additionally, pretreatment with TSN
significantly alleviates apoptosis, oxidative stress and
inflammatory responses to reduce cardiomyocyte death following IRI
(52-54). Here, TSN pretreatment could
effectively increase cell viability and reduced apoptotic rate and
caspase-3 activity following A/R injury, which was similar to the
protective effects of the apoptosis inhibitor Z-VAD and the ROS
inhibitor NAC. However, TSN treatment at high concentration
severely inhibits heart development and leads to cardiac toxicity
(55). Here, TSN pretreatment
effectively protected the myocardium against A/R injury. Cell
viability following 32 μM TSN treatment was significantly
decreased compared with other TSN doses in the A/R injury model,
suggesting that high concentrations of TSN could not attenuate
A/R-induced cardiotoxicity.
Although a protective effect of TSN was identified,
the underlying molecular mechanisms of action are unknown. Here,
TSN and Fer-1 significantly downregulated A/R-induced VDAC1
upregulation and molecular docking simulation revealed the direct
interaction between TSN and VDAC1. Furthermore, the potential
association between VDAC1 and protection effect of TSN was
explored. The effects of TSN were significantly reversed after the
overexpression of VDAC1 using pAd/VDAC1. Therefore, it was
concluded that VDAC1 was the target of TSN.
VDAC1 participates in the synthesis of mPTP, a key
pore located in the outer membrane of the mitochondria and is
involved in signal transduction and transport of metabolites
(56,57). In numerous studies, it has been
suggested that upregulated VDAC1 expression serves an important
role in the pathological process of IRI, while overexpression of
VDAC1 aggravates myocardial damage through the activation of
mitochondrial apoptosis based on release of apoptotic factors
(16,58). Here, VDAC1 overexpression
prevented the protective effect of TSN against A/R injury. In
addition, overexpression of VDAC1 could decrease the levels of MMP
and apoptotic rate induced by TSN. Furthermore, overexpression of
VDAC1 could open mPTP and inhibit the protective effect of TSN
against mitochondrial damage during A/R injury. Therefore, TSN
attenuates mitochondrial dysfunction and mitochondrial-related
apoptosis induced by A/R injury via targeting VDAC1.
Apoptosis is a major form of cell death during IRI
that involves various genes and signaling pathways, including
Capase-3, Bax and Bcl-2 (59).
Bcl-2 is an antiapoptotic gene and Bax is a proapoptotic gene, both
of which belong to the Bcl-2 family, while apoptosis is mediated by
the interaction between Bax and Bcl-2 (60). Previous studies have demonstrated
that the apoptotic rate is associated with Bcl-2/Bax ratio; higher
Bcl-2/Bax ratio represents a lower apoptotic rate (61,62). As a member of the caspase protein
family, caspase-3 is involved in the activation and mediation of
apoptosis as a key protease (63). The present study showed that TSN
effectively increased the Bcl-2/Bax ratio and decreased Caspase-3
activity induced by A/R, while the protective effects of TSN
pretreatment were prevented by overexpression of VDAC1. Taken
together, TSN could prevent mitochondrial apoptosis following IRI
by inhibiting upregulation of VDAC1.
Mitochondria play a key role in regulated cell
death, including ferroptosis (iron-dependent) and apoptosis
(caspase-regulated). VDAC1, as a key part of mPTP, is involved in
ferroptosis during tissue damage and organ dysfunction induced by
various types of disease such as cancer, Alzheimer's disease, and
myocardial infraction (64,65). Ferroptosis has recently been
identified as a key contributor to cell death during IRI but the
underlying mechanisms of action of ferroptosis are yet to be
elucidated (66). As a primary
ferroptosis inhibitor, Fer-1 has been used in previous studies,
while Fer-1 inhibits ferroptosis, which is primarily dependent on
inhibiting lipid peroxidation (16,33). To the best of our knowledge,
however, few studies have reported that the protective effects of
Fer-1 are more effective and stable in vivo than in
vitro (13,67). Thus, a more stable and effective
ferroptosis inhibitor may further prevent tissue damage due to IRI.
Previous studies have demonstrated that GPX4 and PTGS2 are key
genes involved in the pathological process of ferroptosis and are
regarded ferroptosis-specific markers (14,68). Therefore, in the present study,
these two genes were selected as ferroptosis indicators. TSN
significantly alleviated cell damage following A/R injury, which
was similar to the protective effects of Fer-1. Additionally, VDAC1
overexpression effectively prevented the effects of TSN on the
damage of cardiomyocytes resulting from A/R injury.
The major pathological process of ferroptosis is
dependent on excess iron and lipoxygenase, which promote generation
of ROS and other superoxides that cause lipid peroxidation via the
Fenton reaction (69).
Additionally, depletion of GSH and excess generation of GSSG lead
to activation of ferroptosis (70). Increased total iron content, MDA
(products of lipid peroxidation), ROS and GSSG are associated with
ferroptosis (15). Following
pretreatment with TSN or Fer-1, upregulation of genes related to
ferroptosis and the levels of total iron, MDA, ROS and GSSG were
significantly decreased, while GSH levels and the GSH/GSSG ratio
were increased. However, the present study did not prove that TSN
could exert a protective effect against A/R injury by inhibiting
ferroptosis. Previous studies have reported that erastin treatment
could effectively cause ferroptosis in H9c2 cardiomyocytes
(71,72). The present study showed similar
results that erastin treatment could remarkably increase levels of
ferrous and total iron, lipid ROS and MDA and pretreatment with TSN
could significantly prevent the effect of erastin on H9c2 cells,
indicating that TSN could prevent ferroptosis. In summary, TSN
remarkably attenuated cardiomyocyte injury by inhibiting
ferroptosis during A/R. Overexpression of VDAC1 prevented the
protective effects of TSN. Together, these results demonstrated
that TSN effectively prevented ferroptosis following A/R injury via
downregulation of VDAC1.
Here, only ferroptosis-specific inhibitors were used
to investigate the molecular mechanisms of TSN's protection in
vitro. To understand the underlying mechanisms of action,
transgenic or knockout rat models or cells overexpressing VDAC1 and
ferroptosis-associated genes are necessary and the protective
effect of co-treatment with TSN and Fer-1 on A/R injury also
requires further study. Additionally, although a direct interaction
was predicted between TSN and VDAC1 using molecular docking, this
should be validated using ex vivo pull-down assay. Finally,
the ability of VDAC1 to transit between open and closed
configurations is involved in the regulation of ferroptosis and
apoptosis but was not confirmed in the present study (73).
The present study demonstrated that TSN protected
cardiomyocytes from A/R injury by inhibiting ferroptosis and
apoptosis. Additionally, protection of TSN was prevented by
overexpression of VDAC1. VDAC1 and ferroptosis-related proteins
play a vital role in the pathological process of IRI and clinical
application of TSN deserves further study.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HH, SQ-L and JC-L conceived and designed the study.
TH, HX-Z, HH, SQ-L and SY-L performed cell experiments and data
analysis and interpretation. YR-W, YM-Q and YY performed cell
experiments. All authors wrote the manuscript. All authors have
read and approved the final manuscript. HH and SQ-L revised the
manuscript. HH and SQ-L confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of Jiangxi, China (grant no. 20212BAB206021), Key
Projects of Jiangxi Natural Science Foundation (grant no.
20224ACB206002), National Natural Science Foundation of China
(grant nos. 81960059 and 82160073) and Jiangxi Provincial Natural
Science Foundation (grant no. 20232BAB206009).
References
|
1
|
Davidson SM, Ferdinandy P, Andreadou I,
Bøtker HE, Heusch G, Ibáñez B, Ovize M, Schulz R, Yellon DM,
Hausenloy DJ, et al: Multitarget strategies to reduce myocardial
ischemia/reperfusion injury: JACC review topic of the week. J Am
Coll Cardiol. 73:89–99. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hausenloy DJ, Bøtker HE, Ferdinandy P,
Heusch G, Ng GA, Redington A and Garcia-Dorado D: Cardiac
innervation in acute myocardial ischaemia/reperfusion injury and
cardioprotection. Cardiovasc Res. 115:1167–1177. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee JR, Park BW, Park JH, Lim S, Kwon SP,
Hwang JW, Kim H, Park HJ and Kim BS: Local delivery of a senolytic
drug in ischemia and reperfusion-injured heart attenuates cardiac
remodeling and restores impaired cardiac function. Acta Biomater.
135:520–533. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gumpper-Fedus K, Park KH, Ma H, Zhou X,
Bian Z, Krishnamurthy K, Sermersheim M, Zhou J, Tan T, Li L, et al:
MG53 preserves mitochondrial integrity of cardiomyocytes during
ischemia reperfusion-induced oxidative stress. Redox Biol.
54:1023572022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ren D, Quan N, Fedorova J, Zhang J, He Z
and Li J: Sestrin2 modulates cardiac inflammatory response through
maintaining redox homeostasis during ischemia and reperfusion.
Redox Biol. 34:1015562020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Duan W, Yang Y, Yan J, Yu S, Liu J, Zhou
J, Zhang J, Jin Z and Yi D: The effects of curcumin post-treatment
against myocardial ischemia and reperfusion by activation of the
JAK2/STAT3 signaling pathway. Basic Res Cardiol. 107:2632012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
LLi ZM, Xu SW and Liu PQ: Salvia
miltiorrhizaBurge (Danshen): A golden herbal medicine in
cardiovascular therapeutics. Acta Pharmacol Sin. 39:802–824. 2018.
View Article : Google Scholar
|
|
8
|
Zhang X, Ma Z, Liang Q, Tang X, Hu D, Liu
C, Tan H, Xiao C, Zhang B, Wang Y and Gao Y: Tanshinone IIA exerts
protective effects in a LCA-induced cholestatic liver model
associated with participation of pregnane X receptor. J
Ethnopharmacol. 164:357–367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo J, Song W, Yang G, Xu H and Chen K:
Compound Danshen (Salvia miltiorrhiza) dripping pill for coronary
heart disease: An overview of systematic reviews. Am J Chin Med.
43:25–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shang Q, Xu H and Huang L: Tanshinone IIA:
A promising natural cardioprotective agent. Evid Based Complement
Alternat Med. 2012:7164592012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Q, Shen L, Wang Z, Jiang HP and Liu LX:
Tanshinone IIA protects against myocardial ischemia reperfusion
injury by activating the PI3K/Akt/mTOR signaling pathway. Biomed
Pharmacother. 84:106–114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiao H, Zhang M, Wu H, Wu J, Hu X, Pei X,
Li D, Zhao L, Hua Q, Meng B, et al: CIRKIL exacerbates cardiac
ischemia/reperfusion injury by interacting with Ku70. Circ Res.
130:e3–e17. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao WK, Zhou Y, Xu TT and Wu Q:
Ferroptosis: Opportunities and challenges in myocardial
ischemia-reperfusion injury. Oxid Med Cell Longev.
2021:99296872021.PubMed/NCBI
|
|
14
|
Li J, Cao F, Yin HL, Huang ZJ, Lin ZT, Mao
N, Sun B and Wang G: Ferroptosis: Past, present and future. Cell
Death Dis. 11:882020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen X, Kang R, Kroemer G and Tang D:
Ferroptosis in infection, inflammation, and immunity. J Exp Med.
218:e202105182021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Feng Y, Madungwe NB, Imam Aliagan AD,
Tombo N and Bopassa JC: Liproxstatin-1 protects the mouse
myocardium against ischemia/reperfusion injury by decreasing VDAC1
levels and restoring GPX4 levels. Biochem Biophys Res Commun.
520:606–611. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He H, Wang L, Qiao Y, Zhou Q, Yang B, Yin
L, Yin D and He M: Vinegar/Tetramethylpyrazine Induces Nutritional
Preconditioning Protecting the Myocardium Mediated by VDAC1. Oxid
Med Cell Longev. 2021:66700882021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin D, Cui B, Ren J and Ma J: Regulation
of VDAC1 contributes to the cardioprotective effects of
penehyclidine hydrochloride during myocardial ischemia/reperfusion.
Exp Cell Res. 367:257–263. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pooja S, Pushpanathan M, Gunasekaran P and
Rajendhran J: Endocytosis-Mediated Invasion and Pathogenicity of
Streptococcus agalactiae in Rat Cardiomyocyte (H9C2). PLoS One.
10:e01397332015. View Article : Google Scholar
|
|
20
|
Wang L, Lai S, Zou H, Zhou X, Wan Q, Luo
Y, Wu Q, Wan L, Liu J and Huang H: Ischemic
preconditioning/ischemic postconditioning alleviates
anoxia/reoxygenation injury via the Notch1/Hes1/VDAC1 axis. J
Biochem Mol Toxicol. 36:e231992022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou XL, Wu X, Xu QR, Zhu RR, Xu H, Li YY,
Liu S, Huang H, Xu X, Wan L, et al: Notch1 provides myocardial
protection by improving mitochondrial quality control. J Cell
Physiol. 234:11835–11841. 2019. View Article : Google Scholar
|
|
22
|
Sayers EW, Beck J, Bolton EE, Bourexis D,
Brister JR, Canese K, Comeau DC, Funk K, Kim S, Klimke W, et al:
Database resources of the National Center for biotechnology
information. Nucleic Acids Res. 49:D10–D17. 2021. View Article : Google Scholar
|
|
23
|
Karimi F, Hamidian Y, Behrouzifar F,
Mostafazadeh R, Ghorbani-HasanSaraei A, Alizadeh M, Mortazavi SM,
Janbazi M and Naderi Asrami P: An applicable method for extraction
of whole seeds protein and its determination through Bradford's
method. Food Chem Toxicol. 164:1130532022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoshida Y, Shimakawa S, Itoh N and Niki E:
Action of DCFH and BODIPY as a probe for radical oxidation in
hydrophilic and lipophilic domain. Free Radic Res. 37:861–872.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Flameng W, Borgers M, Daenen W and
Stalpaert G: Ultrastructural and cytochemical correlates of
myocardial protection by cardiac hypothermia in man. J Thorac
Cardiovasc Sur. 79:413–424. 1980. View Article : Google Scholar
|
|
26
|
Liu X, Shi Y, Deng Y and Dai R: Using
molecular docking analysis to discovery Dregea sinensis Hemsl.
Potential mechanism of anticancer, antidepression, and
immunoregulation. Pharmacogn Mag. 13:358–362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
UniProt Consortium: UniProt: A hub for
protein information. Nucleic Acids Res. 43:D204–D212. 2015.
View Article : Google Scholar :
|
|
28
|
Hähnke VD, Kim S and Bolton EE: PubChem
chemical structure standardization. J Cheminform. 10:362018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng Y, Wang L, Zhang Z, He X, Fan Q,
Cheng X, Qiao Y, Huang H, Lai S, Wan Q, et al: Puerarin activates
adaptive autophagy and protects the myocardium against
doxorubicin-induced cardiotoxicity via the 14-3-3γ/PKCε pathway.
Biomed Pharmacother. 153:1134032022. View Article : Google Scholar
|
|
30
|
Van der Paal J, Neyts EC, Verlackt CCW and
Bogaerts A: Effect of lipid peroxidation on membrane permeability
of cancer and normal cells subjected to oxidative stress. Chem Sci.
7:489–498. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Camaschella C, Nai A and Silvestri L: Iron
metabolism and iron disorders revisited in the hepcidin era.
Haematologica. 105:260–272. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fang X, Wang H, Han D, Xie E, Yang X, Wei
J, Gu S, Gao F, Zhu N, Yin X, et al: Ferroptosis as a target for
protection against cardiomyopathy. Proc Natl Acad Sci USA.
116:2672–2680. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao T, Wu W, Sui L, Huang Q, Nan Y, Liu J
and Ai K: Reactive oxygen species-based nanomaterials for the
treatment of myocardial ischemia reperfusion injuries. Bioact
Mater. 7:47–72. 2022. View Article : Google Scholar
|
|
35
|
Zhang Z, Dalan R, Hu Z, Wang JW, Chew NW,
Poh KK, Tan RS, Soong TW, Dai Y, Ye L and Chen X: Reactive oxygen
species scavenging nanomedicine for the treatment of ischemic heart
disease. Adv Mater. 34:e22021692022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou T, Wang Z, Guo M, Zhang K, Geng L,
Mao A, Yang Y and Yu F: Puerarin induces mouse mesenteric
vasodilation and ameliorates hypertension involving endothelial
TRPV4 channels. Food Funct. 11:10137–10148. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li T, Guo R, Zong Q and Ling G:
Application of molecular docking in elaborating molecular
mechanisms and interactions of supramolecular cyclodextrin.
Carbohydr Polym. 276:1186442022. View Article : Google Scholar
|
|
38
|
Del Re DP, Amgalan D, Linkermann A, Liu Q
and Kitsis RN: Fundamental mechanisms of regulated cell death and
implications for heart disease. Physiol Rev. 99:1765–1817. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ye J, Huang Y, Que B, Chang C, Liu W, Hu
H, Liu L, Shi Y, Wang Y, Wang M, et al: Interleukin-12p35 knock out
aggravates doxorubicin-induced cardiac injury and dysfunction by
aggravating the inflammatory response, oxidative stress, apoptosis
and autophagy in mice. EBioMedicine. 35:29–39. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
He H, Zhou Y, Huang J, Wu Z, Liao Z, Liu
D, Yin D and He M: Capsaicin protects cardiomyocytes against
anoxia/reoxygenation injury via preventing mitochondrial
dysfunction mediated by SIRT1. Oxid Med Cell Longev.
2017:10357022017. View Article : Google Scholar
|
|
41
|
Lu M, Jia M, Wang Q, Guo Y, Li C, Ren B,
Qian F and Wu J: The electrogenic sodium bicarbonate cotransporter
and its roles in the myocardial ischemia-reperfusion induced
cardiac diseases. Life Sci. 270:1191532021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ren D, Quan N, Fedorova J, Zhang J, He Z
and Li J: Sestrin2 modulates cardiac inflammatory response through
maintaining redox homeostasis during ischemia and reperfusion.
Redox Biol. 34:1015562020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang H, Zheng B, Che K, Han X, Li L, Wang
H, Liu Y, Shi J and Sun S: Protective effects of safranal on
hypoxia/reoxygenation-induced injury in H9c2 cardiac myoblasts via
the PI3K/AKT/GSK3β signaling pathway. Exp Ther Med. 22:14002021.
View Article : Google Scholar
|
|
44
|
Heusch G: Myocardial ischaemia-reperfusion
injury and cardioprotection in perspective. Nat Rev Cardiol.
17:773–789. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Maslov LN, Popov SV, Naryzhnaya NV,
Mukhomedzyanov AV, Kurbatov BK, Derkachev IA, Boshchenko AA,
Khaliulin I, Prasad NR, Singh N, et al: The regulation of
necroptosis and perspectives for the development of new drugs
preventing ischemic/reperfusion of cardiac injury. Apoptosis.
27:697–719. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ma W, Wei S, Zhang B and Li W: Molecular
Mechanisms of cardiomyocyte death in drug-induced cardiotoxicity.
Front Cell Dev Biol. 8:4342020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Abdukeyum GG, Owen AJ and McLennan PL:
Dietary (n-3) long-chain polyunsaturated fatty acids inhibit
ischemia and reperfusion arrhythmias and infarction in rat heart
not enhanced by ischemic preconditioning. J Nutr. 138:1902–1909.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hausenloy DJ and Yellon DM: The
therapeutic potential of ischemic conditioning: An update. Nat Rev
Cardiol. 8:619–629. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhou B, Zhang J, Chen Y, Liu Y, Tang X,
Xia P, Yu P and Yu S: Puerarin protects against sepsis-induced
myocardial injury through AMPK-mediated ferroptosis signaling.
Aging (Albany NY). 14:3617–3632. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cheng TO: Cardiovascular effects of
Danshen. Int J Cardiol. 121:9–22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang Z, He H, Qiao Y, Huang J, Wu Z, Xu
P, Yin D and He M: Tanshinone IIA pretreatment protects H9c2 cells
against Anoxia/reoxygenation injury: Involvement of the
translocation of Bcl-2 to mitochondria mediated by 14-3-3η. Oxid
Med Cell Longev. 2018:35839212018. View Article : Google Scholar
|
|
52
|
Feng J, Li S and Chen H: Tanshinone IIA
inhibits myocardial remodeling induced by pressure overload via
suppressing oxidative stress and inflammation: Possible role of
silent information regulator 1. Eur J Pharmacol. 791:632–639. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tong Y, Xu W, Han H, Chen Y, Yang J, Qiao
H, Hong D, Wu Y and Zhou C: Tanshinone IIA increases recruitment of
bone marrow mesenchymal stem cells to infarct region via
up-regulating stromal cell-derived factor-1/CXC chemokine receptor
4 axis in a myocardial ischemia model. Phytomedicine. 18:443–450.
2011. View Article : Google Scholar
|
|
54
|
Chen L, Wei L, Yu Q, Shi H and Liu G:
Tanshinone IIA alleviates hypoxia/reoxygenation induced
cardiomyocyte injury via lncRNA AK003290/miR-124-5p signaling. BMC
Mol Cell Biol. 21:202020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang T, Wang C, Wu Q, Zheng K, Chen J, Lan
Y, Qin Y, Mei W and Wang B: Evaluation of Tanshinone IIA
developmental toxicity in Zebrafish embryos. Molecules. 22:6602017.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhou H, Zhang Y, Hu S, Shi C, Zhu P, Ma Q,
Jin Q, Cao F, Tian F and Chen Y: Melatonin protects cardiac
microvasculature against ischemia/reperfusion injury via
suppression of mitochondrial fission-VDAC1-HK2-mPTP-mitophagy axis.
J Pineal Res. 63:e124132017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jiang L, Wang H, Chen G, Feng Y, Zou J,
Liu M, Liu K, Wang N, Zhang H, Wang K and Xiao X: WDR26/MIP2
interacts with VDAC1 and regulates VDAC1 expression levels in H9c2
cells. Free Radic Biol Med. 117:58–65. 2018. View Article : Google Scholar
|
|
58
|
Liao Z, Liu D, Tang L, Yin D, Yin S, Lai
S, Yao J and He M: Long-term oral resveratrol intake provides
nutritional preconditioning against myocardial ischemia/reperfusion
injury: Involvement of VDAC1 downregulation. Mol Nutr Food Res.
59:454–464. 2015. View Article : Google Scholar
|
|
59
|
Zhang Y, Yang X, Ge X and Zhang F:
Puerarin attenuates neurological deficits via Bcl-2/Bax/cleaved
caspase-3 and Sirt3/SOD2 apoptotic pathways in subarachnoid
hemorrhage mice. Biomed Pharmacother. 109:726–733. 2019. View Article : Google Scholar
|
|
60
|
Nishikawa S, Tatsumi T, Shiraishi J,
Matsunaga S, Takeda M, Mano A, Kobara M, Keira N, Okigaki M,
Takahashi T and Matsubara H: Nicorandil regulates Bcl-2 family
proteins and protects cardiac myocytes against hypoxia-induced
apoptosis. J Mol Cell Cardiol. 40:510–519. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Misao J, Hayakawa Y, Ohno M, Kato S,
Fujiwara T and Fujiwara H: Expression of bcl-2 protein, an
inhibitor of apoptosis, and Bax, an accelerator of apoptosis, in
ventricular myocytes of human hearts with myocardial infarction.
Circulation. 94:1506–1512. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang L, Wang YN, Ju JM, Shabanova A, Li
Y, Fang RN, Sun JB, Guo YY, Jin TZ, Liu YY, et al: Mzb1 protects
against myocardial infarction injury in mice via modulating
mitochondrial function and alleviating inflammation. Acta
Pharmacolo Sin. 42:691–700. 2021. View Article : Google Scholar
|
|
63
|
Liou CM, Tsai SC, Kuo CH, Ting H and Lee
SD: Cardiac Fas-dependent and mitochondria-dependent apoptosis
after chronic cocaine abuse. Int J Mol Sci. 15:5988–6001. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Niu B, Lei X, Xu Q, Ju Y, Xu D, Mao L, Li
J, Zheng Y, Sun N, Zhang X, et al: Protecting mitochondria via
inhibiting VDAC1 oligomerization alleviates ferroptosis in
acetaminophen-induced acute liver injury. Cell Biol Toxicol.
38:505–530. 2022. View Article : Google Scholar
|
|
65
|
Nagakannan P, Islam MI, Karimi-Abdolrezaee
S and Eftekharpour E: Inhibition of VDAC1 protects against
glutamate-induced oxytosis and mitochondrial fragmentation in
hippocampal HT22 cells. Cell Mol Neurobiol. 39:73–85. 2019.
View Article : Google Scholar
|
|
66
|
Liu P, Feng Y, Li H, Chen X, Wang G, Xu S,
Li Y and Zhao L: Ferrostatin-1 alleviates
lipopolysaccharide-induced acute lung injury via inhibiting
ferroptosis. Cell Mol Biol Lett. 25:102020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Miotto G, Rossetto M, Di Paolo ML, Orian
L, Venerando R, Roveri A, Vučković AM, Bosello Travain V, Zaccarin
M, Zennaro L, et al: Insight into the mechanism of ferroptosis
inhibition by ferrostatin-1. Redox Biol. 28:1013282020. View Article : Google Scholar
|
|
68
|
Zhang Z, Guo M, Li Y, Shen M, Kong D, Shao
J, Ding H, Tan S, Chen A, Zhang F and Zheng S: RNA-binding protein
ZFP36/TTP protects against ferroptosis by regulating autophagy
signaling pathway in hepatic stellate cells. Autophagy.
16:1482–1505. 2020. View Article : Google Scholar :
|
|
69
|
He YJ, Liu XY, Xing L, Wan X, Chang X and
Jiang HL: Fenton reaction-independent ferroptosis therapy via
glutathione and iron redox couple sequentially triggered lipid
peroxide generator. Biomaterials. 241:1199112020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Fu C, Wu Y, Liu S, Luo C, Lu Y, Liu M,
Wang L, Zhang Y and Liu X: Rehmannioside A improves cognitive
impairment and alleviates ferroptosis via activating PI3K/AKT/Nrf2
and SLC7A11/GPX4 signaling pathway after ischemia. J
Ethnopharmacol. 289:1150212022. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li S, Lei Z, Yang X, Zhao M, Hou Y, Wang
D, Tang S, Li J and Yu J: Propofol protects myocardium from
ischemia/reperfusion injury by inhibiting ferroptosis through the
AKT/p53 signaling pathway. Front Pharmacol. 13:8414102022.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Xu S, Wu B, Zhong B, Lin L, Ding Y, Jin X,
Huang Z, Lin M, Wu H and Xu D: Naringenin alleviates myocardial
ischemia/reperfusion injury by regulating the nuclear
factor-erythroid factor 2-related factor 2 (Nrf2)/System
xc-/glutathione peroxidase 4 (GPX4) axis to inhibit ferroptosis.
Bioengineered. 12:10924–10934. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Vander Heiden MG, Chandel NS, Li XX,
Schumacker PT, Colombini M and Thompson CB: Outer mitochondrial
membrane permeability can regulate coupled respiration and cell
survival. Proc Natl Acad Sci USA. 97:4666–4671. 2000. View Article : Google Scholar : PubMed/NCBI
|