Introduction
Resveratrol
(3,5,4′-trihydroxy-trans-stilbene) is a plant polyphenol
that is present in grapes, red wine, blueberries, mulberries, and
cranberries (1). In vitro
studies have shown that resveratrol can inhibit proliferation and
induce apoptosis in different types of cancer cells (1). In vivo, resveratrol has been
reported to be an effective agent against breast, esophageal, lung,
and colon cancers in animal models (2–6).
Furthermore, resveratrol has been reported to sensitize
neuroblastoma, glioblastoma, breast carcinoma, prostate carcinoma,
leukemia, and pancreatic carcinoma cells to the actions of multiple
traditional chemotherapeutic agents in vitro(7). At a molecular level, resveratrol has
been reported to act as an antioxidant, inhibit transcription
factor and kinase activation required for cell growth, and inhibit
cell cycle progression (1,8–10).
High-risk acute lymphoblastic leukemia (ALL) with
chromosomal translocation t(4;11) has a poor prognosis for
patients. A significant problem that limits survival of patients
with t(4;11) ALL is the relapse of chemotherapy-resistant leukemia
(11,12). The t(4;11) ALL is a pre-B cell
leukemia found in 60–85% of infants with ALL. Several cell lines
have been established from patients with t(4;11) ALL and these
lines have been used in vitro to evaluate novel therapeutic
agents (13–15). The nonobese diabetic x severe
combined immunodeficient (NOD/SCID) mouse model has been useful for
evaluating different chemotherapeutic agents against leukemia
(16–19). The SCID background has an absence
of T and B lymphocyte populations and the NOD background provides
reduced natural killer lymphocyte activity and absence of
circulating complement that increase engraftment efficiency of
human cells. The NOD/SCID mouse model for leukemia mimics the human
disease by homing to, and engrafting in, the bone marrow, spleen,
and liver. The engraftment in organ sites correlates well with the
presence of leukemia cells in the peripheral blood (20).
We have reported that resveratrol, as well as
several other plant-derived polyphenols, such as carnosol,
curcumin, and quercetin, are effective in vitro in inducing
apoptotic cell death in t(4;11) and other ALL-derived cell lines
(21–23). While resveratrol has been shown to
act by stimulating CD95− signaling in some cancer cells,
the induction of apoptosis in the t(4;11) ALL lines by resveratrol
occurred exclusively by disruption of mitochondrial homeostasis
(21). These in vitro
studies supported the concept that resveratrol has potential as a
preventative agent against high-risk leukemia in vivo. In
the current study, we determined whether dietary resveratrol
prevented the engraftment and growth of the t(4;11) ALL cells in
NOD/SCID mice. In addition, we tested the hypothesis that it could
increase the efficacy of the chemotherapeutic agent vincristine; a
standard chemotherapeutic agent used to treat this type of leukemia
(24).
Materials and methods
Cells and reagents
SEM is a cell line that was established from a
patient diagnosed with high-risk pre-B ALL containing the
chromosomal translocation t(4;11)(q21;q23) (13). The cells were grown at 37°C, 5%
CO2 in RPMI-1640 (Invitrogen, Carlsbad, CA) that was
supplemented with 10% fetal bovine serum (Sigma, St. Louis, MO), 50
IU/ml penicillin, 50 mg/l streptomycin, 0.25 mg/l amphotericin B, 1
mmol/l sodium pyruvate, and 2 mmol/l L-glutamine (Invitrogen). For
injection into mice, SEM cells were collected and washed 2 times in
Dulbecco’s PBS without Ca2+ or Mg+ (Sigma).
The cells were resuspended in PBS at a final concentration of
50×106 cells/ml.
Vincristine sulfate (Sigma) was dissolved in PBS and
stored at −20°C. To monitor leukemia burden in the blood,
peripheral blood leukocytes were stained with phycoerythrincyanin 7
(PE-Cy7)-conjugated anti-human CD19 and allophycocyanin-Cy7
(APC-Cy7) conjugated anti-mouse CD45 (Becton-Dickinson, San Jose,
CA). Trans-resveratrol, tetra-deuterated trans-resveratrol
(resveratrol-d4), resveratrol-3-O-D-glucuronide,
resveratrol-4′-O-D-glucuronide, and resveratrol-3-O-sulfate, and
1-cyclohexyluriedo-3-dodecanoic acid (CUDA) were purchased from
Cayman Chemical Co. (Ann Arbor, MI). Sulfatase from Aerobacter
aerogenes, β-glucuronidase (Type IX-A) from Escherichia
coli, formic acid, glycerol, potassium 4-nitrophenyl sulfate,
and 4-nitrophenyl β-D-glucuronide were obtained from Sigma.
Ammonium hydroxide and LC/MS grades of methanol, acetonitrile, and
water were purchased from Fisher Scientific (Fair Lawn, NJ). Normal
mouse serum was obtained from United States Biological (Swampscott,
MA).
Diets
Resveratrol (>98% pure) was purchased in bulk
from Cayman Chemical Co. Rodent Diet 7013 (Harlan Teklad, Madison,
WI), a commercial NIH-31 modified diet, was used as the base diet
because it is similar in composition to the diet used by Jackson
Laboratory for maintenance of the NOD. CB17-Prkdcscid/J mouse
strain. The diets used in the current study were prepared by Harlan
Teklad. Diet 1 was the base diet (control) and diet 2 was the base
diet containing 0.2% w/w resveratrol. A dietary concentration of
0.2% w/w resveratrol is equivalent to approximately 300 mg/kg body
weight/day assuming a 20-g mouse and consumption of 3 g of
food/day. This dietary concentration was chosen to both minimize
the risk for resveratrol-induced renal toxicity (25), and ensure that reasonable blood
concentrations of resveratrol would reach the leukemia cells at
engraftment sites. For the addition of the supplement, Rodent Diet
7013 was ground, the resveratrol was added, and the diet was
repelleted. The control diet was also ground and repelleted so that
the consistency of the food for each group was similar. The diets
were γ-irradiated to sterilize and packaged in 2 kg vacuum sealed
bags to reduce exposure to air. Diets were stored at −20°C until
use and fresh food was given to the mice weekly.
Mice
Experimental procedures using mice were approved by
the University of California, Davis Institutional Animal Care and
Use Committee. Female NOD.CB17-Prkdcscid/J mice (5 weeks of age)
were purchased from the Jackson Laboratory (Bar Harbor, ME, common
name NOD/SCID). Upon arrival, the mice were randomly sorted into 2
groups (n=16 per group) and given the control diet or diet
containing 0.2% w/w resveratrol. Mice were given sterilized food
and water ad libitum. Mice were housed under 12 h light-dark
cycle, pathogen-free, temperature controlled conditions in
ventilator racks at a University of California, Davis vivarium.
Mice were weighed weekly. Weighing of mice and injections of
leukemia cells or chemotherapeutic agent were performed in a
biosafety cabinet to maintain pathogen-free conditions. Additions
of food and water, and cage changes were performed in a laminar
flow change-out cabinet. Mice were euthanized by carbon dioxide
asphyxiation.
Leukemia cell engraftment
After receiving the diets for 3 weeks, each mouse
was injected with 5×106 SEM cells (100 μl volume)
through the tail vein. Approximately 2 weeks after injecting the
leukemia cells, blood was collected from the tail artery of each
mouse once per week using heparinized Microvette tubes (Sarstedt,
Newton, NC). Blood from each mouse (approximately 50 μl) was
transferred to a separate 1.5 ml microfuge tube and the red blood
cells were lysed using PharmLyse (Becton-Dickinson) according to
the manufacturer’s suggestions. The resulting peripheral blood
leukocytes (PBLs) were stained with PE-Cy7 conjugated anti-human
CD19 and APC-Cy7 conjugated anti-mouse CD45. The cells were
incubated with antibodies at room temperature for 20 min, washed in
PBS containing 0.1% BSA and 7 mmol/l sodium azide (Sigma), and then
fixed in 1% paraformaldehyde (Sigma) before analysis on a
FACSCanto™ fluorescence-activated cell sorter (FACS) using
FACSDiva™ software (Becton-Dickinson). Analysis of leukocytes was
performed using appropriate scatter gates to exclude cellular
debris and aggregated cells. The negative control was PBLs isolated
from NOD/SCID mice that had not been injected with leukemia cells.
The positive control was made by spiking an aliquot of PBLs
(isolated from mice without leukemia) with SEM cells. Both the
negative and positive control cells were stained with PE-Cy7
conjugated anti-human CD19 and APC-Cy7 conjugated anti-mouse CD45
and used to set the gates for human CD19+ cells. Thirty
thousand events were collected for each sample. Positive
engraftment was defined as 1% or greater human CD19+
cells present in the murine PBL population (18,19).
Treatment with vincristine
All mice were injected intraperitoneally (i.p.) with
vincristine at a concentration of 0.5 mg/kg body weight 3 times per
week beginning approximately 4.5 weeks after injection of leukemia
cells (26). The total volume for
each injection of vincristine was approximately 100 μl and
was adjusted weekly according to the body weight of each mouse. All
animals were fed the control or resveratrol diets during the
chemotherapeutic treatment, and the percentage of human leukemia
cells was monitored by flow cytometry as described above.
Liquid chromatography (LC) - tandem mass
spectrometry (MS) analysis
Serum samples from NOD/SCID mice (n=5) were prepared
at the time of euthanization and stored at −70°C until use. Serum
from each mouse was separated into three 25 μl aliquots for
enzymatic digestion using a mock control (no enzyme),
β-glucuronidase, or sulfatase. The serum samples were processed and
analyzed as previously described (27).
Statistical analysis
Statistical analyses were performed with GraphPad
software (GraphPad Software, Inc., San Diego, CA) and data are
displayed as arithmetic means ± SEM. Event free survival (EFS) was
used for comparisons between the treatment groups, and was defined
by overt clinical illness requiring euthanization, that included
greater than 20% weight loss, lethargy, severe weakness, or
inability to reach food or water for 24 h. The EFS was calculated
beginning with the day of injection of the leukemia cells.
Kaplan-Meier survival curves were used to determine differences in
EFS by log-rank test. Differences in the body weights and
percentage of CD19+ cells between the dietary groups
were analyzed by 2-way ANOVA with Bonferroni posttests. The
confidence interval for significant differences was set at 95%.
Results
Dietary resveratrol does not inhibit
engraftment of leukemia
Mice were fed specialized diets for 3 weeks prior to
injecting the leukemia cells to determine if dietary resveratrol
delayed engraftment. Engraftment of SEM leukemia cells was 100% in
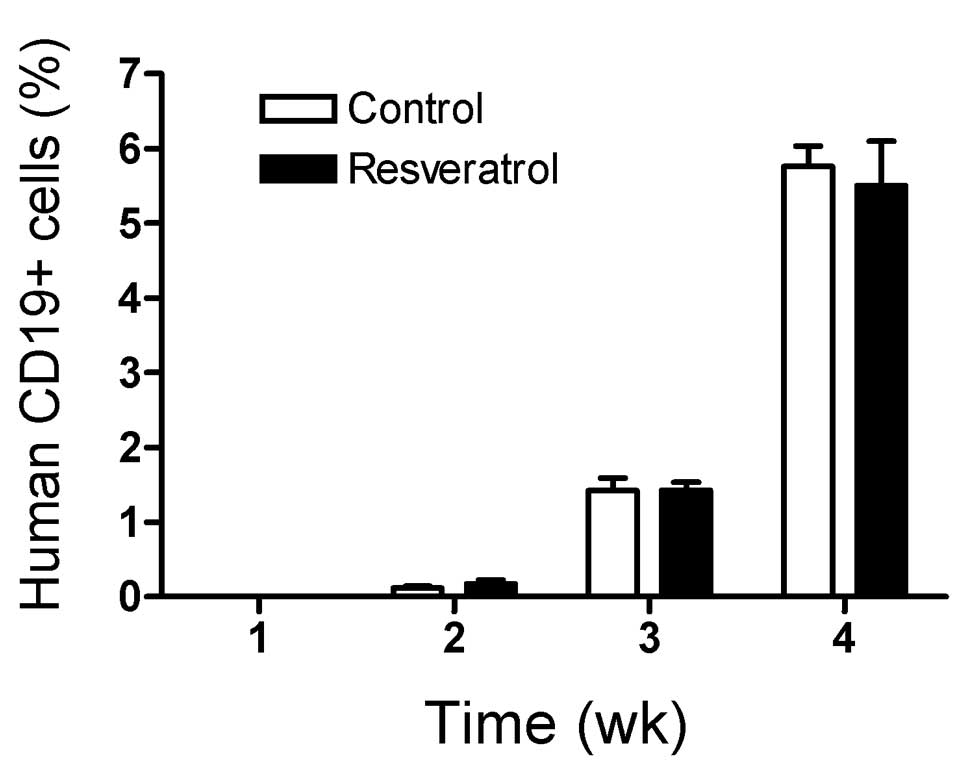
both control and resveratrol fed mice. The rate of engraftment as
measured by the percentage of SEM leukemia cells in the PBL
population in control and resveratrol fed mice was similar over a
4-week period after injection of the leukemia cells and prior to
treatment with vincristine (Fig.
1, P=0.78).
Dietary resveratrol does not sensitize
the leukemia cells to vincristine
Mice were treated with vincristine at week 4 when
the mean percentage of human leukemia cells in the mouse PBL
population reached approximately 5.8±0.3% for the control group and
5.5±0.6% in the resveratrol fed group (Fig. 2). Mice in both feeding groups
initially responded to the vincristine treatment, resulting in a
temporary decrease in leukemia burden. One mouse in the resveratrol
fed group had a leukemia burden of approximately 10% in the
peripheral blood and showed a delayed response to the vincristine
treatment, resulting in the large standard error at week 5
(Fig. 2A). Removal of this mouse
from the data analysis showed that vincristine treatment produced a
similar reduction in the leukemia burden between the two dietary
groups (Fig. 2B). Overall, there
was no difference in the percentage of human leukemia cells in the
mouse PBL population between the mice fed the control or
resveratrol diets at weeks 5 and 6 after vincristine treatment
(P=0.39, Fig. 2A, all data). After
2 weeks of treatment with vincristine, the leukemia burden began to
increase and the mice became clinically ill.
Dietary resveratrol does not increase
survival of leukemic mice
Mice were fed the control or resveratrol
supplemented diets 3 weeks before injection of leukemia cells and
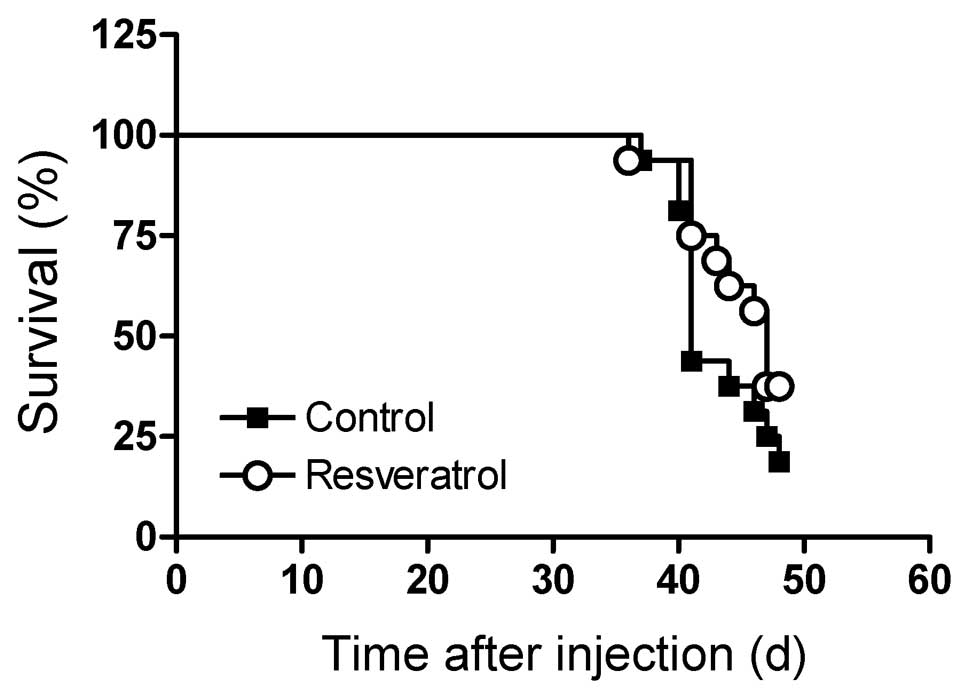
throughout the experimental period. Survival was similar between
the control and resveratrol supplemented mice (Fig. 3, P=0.74). The mice succumbed to the
disease rapidly about 6 weeks after injection of the leukemia
cells, and 2 weeks after treatment with vincristine. Mean body
weights were similar between the control and resveratrol
supplemented mice before and after injection of the leukemia cells
at 8 weeks of age (Fig. 4,
P=0.09). Body weight loss was evident by 4–5 weeks after the
injection of leukemia cells, and the mice continued to lose weight
during the vincristine treatment.
Dietary resveratrol was extensively
metabolized to glucuronides and sulfates
Due to the lack of anti-leukemia activity of
resveratrol, the metabolism of resveratrol was evaluated by LC-MS
in a subpopulation of surviving mice that were showing signs of
clinical illness, but were still mobile and able to reach food
(n=5). The mice were euthanized and the serum was either mock
digested or digested with β-glucuronidase or sulfatase. The
resulting increases in resveratrol aglycone after enzymatic
digestion were used to determine the concentrations of
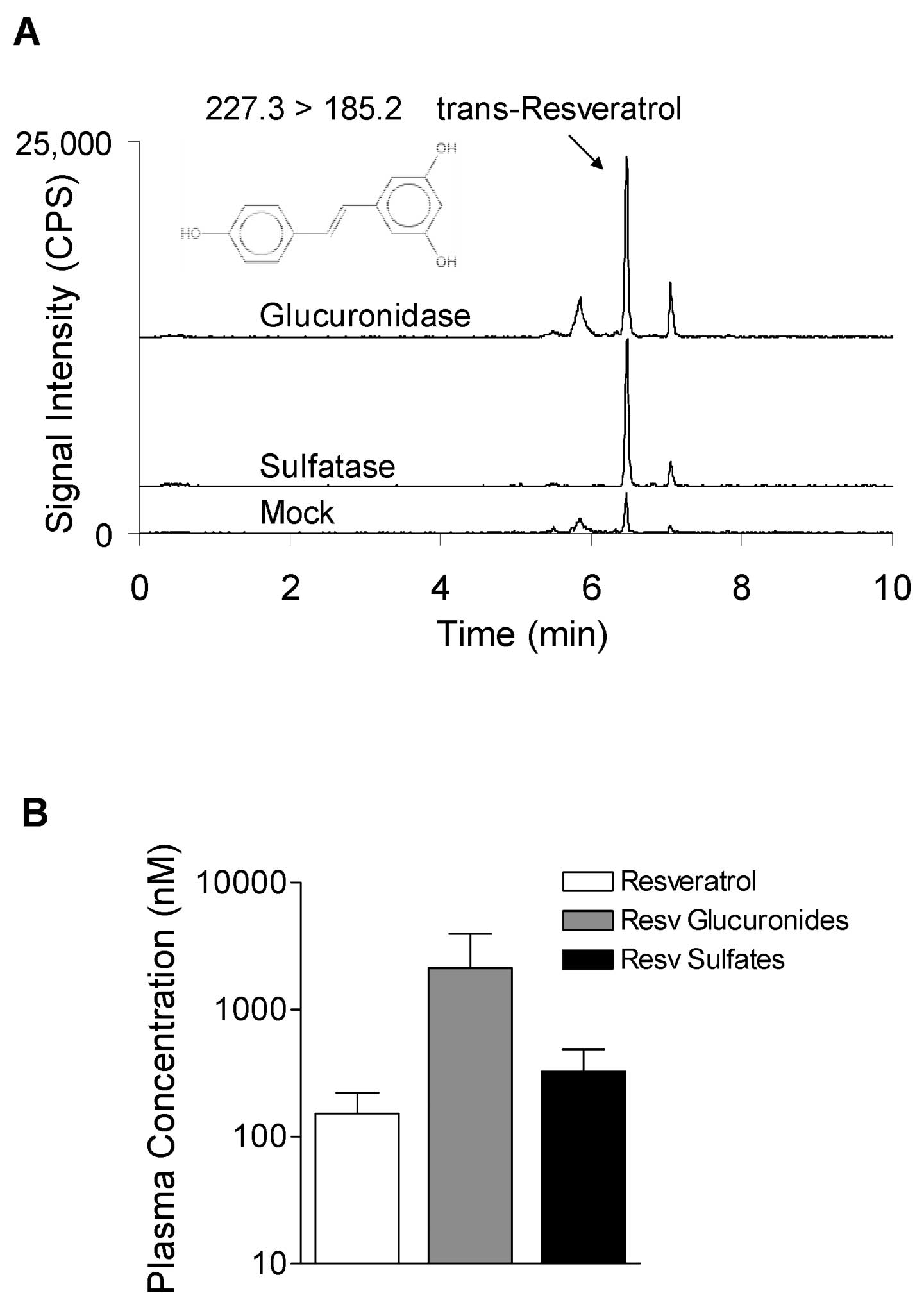
glucuronidated and sulfated metabolites. Incubation of serum with
either β-glucuronidase or sulfatase increased the peak intensities
of extractable resveratrol compared to the mock digested serum
(Fig. 5A). The majority of the
total resveratrol in the serum was present as metabolites. The mean
percentage of resveratrol aglycone was 16±6%, whereas 57±9% and
27±6% of resveratrol was present as monoglucuronides and
monosulfates, respectively (Fig.
5B). The 4-nitrophenyl glucuronide and sulfate conjugates
(digestion controls) showed greater than 99% deconjugation
efficiency. Deuterated resveratrol (surrogate) recoveries were
45±6%, 50±2%, and 62±3% for the mock, β-glucuronidase and sulfatase
digestions, respectively.
Discussion
In human feeding studies, resveratrol has been used
at levels up to 5 g/day, with some adverse results, including
diarrhea, nausea, and abdominal pain, being reported at doses
ranging from 2.5 to 5 g/day (28,29).
In a 50–70 kg adult, the dose of 5 g/day corresponds to 71–100
mg/kg body weight/day in humans. Due to the adverse responses in
humans, it has been suggested that levels of dietary resveratrol
for humans should not exceed 1 g/day. Toxicity studies have been
performed in rats with oral administration of 100, 300, and 3000
mg/kg body weight/day and resveratrol was reported to have renal
toxicity at 3000 mg/kg body weight (25). In the present study with mice, we
chose a dietary concentration of 0.2% w/w resveratrol that is
approximately 300 mg/kg body weight/day assuming a 20-mouse and
consumption of 3 g of food/day diet. This dietary concentration was
used to reduce renal toxicity in the mice, while ensuring there was
a high potential of the resveratrol to reach the organs and tissues
that harbored the leukemia cells. Despite the high amount (0.2% of
the diet) of resveratrol given in the current study, there was no
indication that its inclusion in the diet inhibited the engraftment
or growth of high-risk t(4;11) leukemia in NOD/SCID mice.
Studies have been performed in both humans and
experimental animals to determine the tissue distribution,
excretion rates, and general bioavailability of resveratrol after
its oral administration. In humans, after 25 mg of
14C-labelled resveratrol was given as a single oral
dose, the concentration of resveratrol peaked at approximately 500
ng/ml in the plasma (2 μmol/l) at 1 h, with a second peak of
approximately 300 ng/ml (1.3 μmol/l) at 6 h (30). In these experiments, there was at
least 70% absorption of resveratrol and most of the oral dose was
recovered in the urine. Although the plasma levels were low with
the single oral dose of 25 mg, absorption was rapid and efficient
and the plasma half-life of resveratrol and metabolites was
calculated at 9.2 h. The major forms of resveratrol observed in
this study were sulfate and glucuronide conjugates. More recently,
humans given daily doses of 0.5, 1, 2.5, or 5 g for 29 days showed
maximal plasma concentrations of the resveratrol aglycone at 0.19,
0.62, 1.45, and 4.24 μmol/l, respectively (28). However, the time of maximal plasma
concentration of resveratrol aglycone in the human study was
approximately 1 h and the majority of the resveratrol had been
converted to the resveratrol-4′-O-glucuronide,
resveratrol-3-O-glucuronide, and resveratrol-3-O-sulfate.
The tissue distribution of orally administered
resveratrol has been reported in animal models. In mice, a single
oral dose of 14C-labelled resveratrol (5 mg/kg body
weight) showed distribution in the duodenum, colon, liver, kidney,
lung, spleen, heart, brain, and testis by 3 h, with the highest
content in the duodenum (31).
Longer term experiments utilizing rats showed the presence of
resveratrol metabolites in the plasma after an 8-week feeding of
300 mg resveratrol/kg body weight per day (32). The plasma, liver, kidney, urine,
and feces contained mainly the sulfated and glucuronidated forms of
resveratrol in these rats. By the end of the 8-week feeding study,
the concentrations of the different resveratrol conjugates in the
plasma ranged from 0.37 to 7.46 mg/l (1.6–32.7 μmol/l). In
the present study, only nmol/l levels of resveratrol aglycone were
detected, but approximately 2 μmol/l glucuronide metabolites
were present in the serum of leukemic NOD/SCID mice. Taken
together, the SEM leukemia cells in the target organs (spleen,
liver, and bone marrow) would have been mostly exposed to
resveratrol metabolites rather than the resveratrol aglycone.
Further investigations will be required to evaluate the apoptotic
activities of resveratrol metabolites against leukemia cells.
However, our data suggest that the leukemia cells were not exposed
to high enough levels of resveratrol aglycone and/or metabolites to
induce leukemia cell death.
The majority of in vitro studies on
resveratrol focused on the study of the putative anti-cancer
activity of this nutrient have been conducted using
nonphysiological μmol/l concentrations of the resveratrol
aglycone. We submit that these types of in vitro experiments
have limited value to elucidate the chemopreventive or therapeutic
potential of resveratrol against leukemia in vivo. However,
we note that resveratrol has been reported to have some efficacy in
decreasing tumor burden and metastatic potential of a number of
different cancers in rodent models (33). Resveratrol has been reported to
inhibit metastasis of lung cancer and melanoma after
intraperitoneal injection, and colon carcinoma metastasis after
oral administration (34–36). Resveratrol has been reported to
sensitize a number of cancers to chemotherapeutic agents in
vitro and the mechanisms of this sensitization include
down-regulation of multi-drug resistant protein expression,
modulating the expression of cell survival proteins such as Bcl-2,
down-regulating the transcription factor NF-κB, and cell cycle
arrest (7). Reversal of
doxorubicin resistance in acute myeloid leukemia in vitro
was reported to be mediated by down-regulation of the multi-drug
resistant protein MRP-1, but only at a nonphysiological dose of 50
μmol/l (37). Limited data
exist on the chemosensitizing effects of resveratrol in
vivo. It was recently reported that resveratrol administered
daily by gavage (at a dose of 40 mg/kg body weight) for 35 days
significantly reduced the size of ectopic pancreatic tumors in nude
mice, and that it potentiated the effects of gemcitabine in this
model of pancreatic cancer (38).
These experiments suggest that resveratrol may have potential as a
chemopreventive agent depending upon the type of cancer and/or
route of administration, and may be beneficial in the prevention of
metastasis.
High-risk t(4;11) ALL is an aggressive leukemia that
has a poor prognosis due to development of chemotherapy resistant
cells. The t(4;11) ALL line SEM was established from a relapsed
patient that had undergone chemotherapy (13). Engraftment of these cells in
NOD/SCID mice was achieved rapidly within 2–3 weeks after injection
of the leukemia cells. In vitro, the SEM cells are
responsive to vincristine-induced apoptosis, but upon engraftment
into NOD/SCID mice and treatment with vincristine, SEM cells revert
to a vincristine-resistant phenotype due in part by an increased
expression of the multi-drug resistant protein P-glycoprotein
(26). Dietary resveratrol did not
sensitize these leukemia cells to vincristine treatment, nor did
this dietary nutrient inhibit the rapid engraftment and growth of
SEM cells in the NOD/SCID mice. In summary, while dietary
resveratrol may have potential as a chemopreventive agent against
some cancers, the results presented in the current study suggest
that this agent has minimal value with respect to the treatment or
prevention of high-risk t(4;11) ALL in vivo.
Acknowledgements
This work was supported by an award
from National Institutes of Health, National Cancer Institute,
award no. 1R21CA122117-01. The content is solely the responsibility
of the authors and does not necessarily represent the official
views of the National Institutes of Health. No conflicts of
interest are present for any of the authors. USDA is an equal
opportunity provider and employer.
References
|
1.
|
BB AggarwalA BhardwajRS AggarwalNP SeeramS
ShishodiaRole of resveratrol in prevention and therapy of cancer:
preclinical and clinical studiesAnticancer
Res2427832840200415517885
|
|
2.
|
S BanerjeeC Bueso-RamosBB
AggarwalSuppression of 7,12-dimethylbenz(a)anthracene-induced
mammary carcinogenesis in rats by resveratrol: role of nuclear
factor-κB, cyclo-oxygenase 2, and matrix metalloproteinase 9Cancer
Res6249454954200212208745
|
|
3.
|
ZG LiT HongY ShimadaI KomotoA
KawabeSuppression of N-nitrosomethylbenzylamine (NMBA)-induced
esophageal tumorigenesis in F344 rats by
resveratrolCarcinogenesis2315311536200210.1093/carcin/23.9.153112189197
|
|
4.
|
EO LeeHJ LeeHS HwangKS AhnC ChaePotent
inhibition of Lewis lung cancer growth by heyneanol A from the
roots of Vitis amurensis through apoptotic and anti-angiogenic
activitiesCarcinogenesis2720592069200610.1093/carcin/bgl05516675471
|
|
5.
|
HS LiuCE PanW YangXM LiuAntitumor and
immunomodulatory activity of resveratrol on experimentally
implanted tumor of H22 in Balb/c miceWorld J
Gastroenterol914741476200312854144
|
|
6.
|
L TessitoreA DavitI SarottoG
CaderniResveratrol depresses the growth of colorectal aberrant
crypt foci by affecting bax and p21(CIP)
expressionCarcinogenesis2116191622200010.1093/carcin/21.8.161910910967
|
|
7.
|
SC GuptaR KannappanS ReuterJH KimBB
AggarwalChemosensitization of tumors by resveratrolAnn NY Acad
Sci1215150160201110.1111/j.1749-6632.2010.05852.x21261654
|
|
8.
|
D ColinE LimagneS JeanningrosA JacquelG
LizardEndocytosis of resveratrol via lipid rafts and activation of
downstream signaling pathways in cancer cellsCancer Prev
Res410951106201110.1158/1940-6207.CAPR-10-027421467134
|
|
9.
|
G LiS HeL ChangH LuH ZhangGADD45α and
annexin A1 are involved in the apoptosis of HL-60 induced by
resveratrolPhytomedicine187047092011
|
|
10.
|
M KartalG SaydamF SahinY BaranResveratrol
triggers apoptosis through ceramide metabolizing genes in human
K562 chronic myeloid leukemia cellsNutr
Cancer63637644201110.1080/01635581.2011.53848521500096
|
|
11.
|
S FaderlHM KantarjianM TalpazZ
EstrovClinical significance of cytogenetic abnormalities in adult
acute lymphoblastic leukemiaBlood913995401919989596644
|
|
12.
|
MF GreavesJ WiemelsOrigins of chromosome
translocations in childhood leukaemiaNat
Rev3111200310.1038/nrc1164
|
|
13.
|
J GreilM GramatzkiR BurgerR MarschalekM
PeltnerThe acute lymphoblastic leukaemia cell line SEM with t(4;11)
chromosomal rearrangement is biphenotypic and responsive to
interleukin-7Br J
Haematol86275283199410.1111/j.1365-2141.1994.tb04726.x8199015
|
|
14.
|
RC StongSJ KorsmeyerJL ParkinDC ArthurJH
KerseyHuman acute leukemia cell line with the t(4;11) chromosomal
rearrangement exhibits B lineage and monocytic
characteristicsBlood65213119853917311
|
|
15.
|
B LangeM ValtieriD SantoliD CaraccioloF
MavilioGrowth factor requirements of childhood acute leukemia:
establishment of GM-CSF-dependent cell
linesBlood7019219919873496132
|
|
16.
|
G BaerschT MollersA HotteB
Dockhorn-DworniczakC RubeGood engraftment of B-cell precursor ALL
in NOD-SCID miceKlin
Padiatr209178185199710.1055/s-2008-10439479293448
|
|
17.
|
A BorgmannC BaldyA von StackelbergB
BeyermannI FichtnerChildhood ALL blasts retain phenotypic and
genotypic characteristics upon long-term serial passage in NOD/SCID
micePediatr Hematol
Oncol17635650200010.1080/0888001005021134911127395
|
|
18.
|
NLM LiemRA PapaCG MilrossMA SchmidM
TajbakhshCharacterization of childhood acute lymphoblastic leukemia
xenograft models for the preclinical evaluation of new
therapiesBlood10339053914200410.1182/blood-2003-08-291114764536
|
|
19.
|
RB LockN LiemML FarnsworthCG MilrossC
XueThe nonobese diabetic/severe combined immunodeficient (NOD/SCID)
mouse model of childhood acute lymphoblastic leukemia reveals
intrinsic differences in biologic characteristics at diagnosis and
relapseBlood9941004108200210.1182/blood.V99.11.4100
|
|
20.
|
BA NijmeijerP MollevangerSL van
Zelderen-BholaHC Kluin-NelemansR WillemzeMonitoring of engraftment
and progression of acute lymphoblastic leukemia in individual
NOD/SCID miceExp
Hematol29322329200110.1016/S0301-472X(00)00669-X11274760
|
|
21.
|
J DörrieH GerauerY WachterSJ
ZuninoResveratrol induces extensive apoptosis by depolarizing
mitochondrial membranes and activating caspase-9 in acute
lymphoblastic leukemia cellsCancer Res6147314739200111406544
|
|
22.
|
J DörrieK SapalaSJ ZuninoCarnosol-induced
apoptosis and downregulation of Bcl-2 in B-lineage leukemia
cellsCancer Lett1703339200111448532
|
|
23.
|
C KellnerSJ ZuninoNitric oxide is
synthesized in acute leukemia cells after exposure to phenolic
antioxidants and initially protects against mitochondrial membrane
depolarizationCancer
Lett2154352200410.1016/j.canlet.2004.06.046
|
|
24.
|
JB NachmanHN SatherMG SenselME TriggJM
CherlowAugmented post-induction therapy for children with high-risk
acute lymphoblastic leukemia and a slow response to initial
therapyN Engl J
Med33816631671199810.1056/NEJM1998060433823049614257
|
|
25.
|
JA CrowellPJ KorytkoRL MorrisseyTD BoothBS
LevineResveratrol-associated renal toxicityToxicol
Sci82614619200410.1093/toxsci/kfh26315329443
|
|
26.
|
SJ ZuninoDH StormsJM DucoreNovel in vivo
model of inducible multi-drug resistance in acute lymphoblastic
leukemia with chromosomal translocation t(4;11)Cancer
Lett2964954201010.1016/j.canlet.2010.03.01720381955
|
|
27.
|
SJ ZuninoDH StormsJW NewmanTL PedersenCL
KeenJM DucoreResveratrol given intraperitoneally does not inhibit
the growth of high-risk t(4;11) acute lymphoblastic leukemia cells
in a NOD/SCID mouse modelInt J Oncol4012771284201222200740
|
|
28.
|
VA BrownKR PatelM ViskadurakiJA CrowellM
PerloffRepeat dose study of the cancer chemopreventive agent
resveratrol in healthy volunteers: safety, pharmacokinetics, and
effect on the insulin-like growth factor axisCancer
Res7090039011201010.1158/0008-5472.CAN-10-236420935227
|
|
29.
|
KR PatelE ScottVA BrownAJ GescherWP
StewardClinical trials of resveratrolAnn NY Acad
Sci1215161169201110.1111/j.1749-6632.2010.05853.x21261655
|
|
30.
|
T WalleF HsiehMH De LeggeJE OatisUK
WalleHigh absorption but very low bioavailability of oral
resveratrol in humansDrug Metab
Dispos3213771382200410.1124/dmd.104.00088515333514
|
|
31.
|
X VitracA DesmouliereB BrouillaudS KrisaG
DeffieuxDistribution of [14C]-trans-resveratrol, a cancer
chemopreventive polyphenol, in mouse tissues after oral
administrationLife Sci72221922332003
|
|
32.
|
E WenzelT SoldoH ErbersdoblerV
SomozaBioactivity and metabolism of trans-resveratrol orally
administered to Wistar ratsMol Nutr Food
Res49482494200510.1002/mnfr.20050000315779067
|
|
33.
|
JA BaurDA SinclairTherapeutic potential of
resveratrol: the in vivo evidenceNat Rev Drug
Discov5493506200610.1038/nrd206016732220
|
|
34.
|
Y KimuraH OkudaResveratrol isolated from
Polygonum cuspidatum root prevents tumor growth and metastasis to
lung and tumor-induced neovascularization in Lewis lung
carcinoma-bearing miceJ Nutr13118441849200111385077
|
|
35.
|
S BusquetsE AmetllerG FusterM OlivanV
RaabResveratrol, a natural diphenol, reduces metastatic growth in
an experimental cancer modelCancer
Lett245144148200710.1016/j.canlet.2005.12.03516466851
|
|
36.
|
YL WengHF LiaoAF LiJC ChangRY ChiouOral
administration of resveratrol in suppression of pulmonary
metastasis of BALB/c mice challenged with CT26 colorectal
adenocarcinoma cellsMol Nutr Food
Res54259267201010.1002/mnfr.20090004919862773
|
|
37.
|
SH KweonJH SongTS KimResveratrol-mediated
reversal of doxorubicin resistance in acute myeloid leukemia cells
via down-regulation of MRP1 expressionBiochem Biophys Res
Commun395104110201010.1016/j.bbrc.2010.03.14720350534
|
|
38.
|
KB HarikumarAB Kunnumakkara1G SethiP
DiagaradjaneP AnandResveratrol, a multitargeted agent, can enhance
antitumor activity of gemcitabine in vitro and in orthotopic mouse
model of human pancreatic cancerInt J
Cancer127257268201019908231
|