Introduction
Prostate cancer is a significant health problem,
accounting for approximately 900,000 new cases and more than
258,000 cancer-related deaths worldwide in 2011 (1). Currently, prostate-specific antigen
(PSA) test is clinically used for early detection of prostate
cancer and for surveillance of disease progression, even though it
may not decrease the mortality of the disease (2). Most prostate cancers are slowly
growing, but aggressive prostate cancer cases do occur, especially
in cancers with a high Gleason score, and can metastasize to other
sites of the body, such as the bone and lymph nodes. Thus,
treatment of prostate cancer depends on the severity of the
disease. For aggressive prostate cancer, the treatments are
surgery, radiation therapy, hormonal therapy, chemotherapy or their
combination in order to increase patients’ survival and improve
their quality of life. However, there is currently no cure therapy
available once prostate cancer is metastasized, and androgen
deprivation therapy is one of the standard therapies (3). Since the 1940s, estrogens have been
used for androgen deprivation treatment of prostate cancer as
pioneered by Huggins et al(4). Estrogens inhibit testosterone
biosynthesis through the negative feedback of the
hypothalamic-pituitary-gonadal axis (5) and directly modulate androgen actions
through estrogen receptors (ERs) in prostate cancer cells (6,7).
However, the long-term use of estrogens in treatment of prostate
cancer is limited due to their cardiovascular side-effects, such as
thrombosis and cardiovascular events (8,9).
The mechanisms responsible for estrogen-induced
cardiovascular side-effects are not fully understood. Several
previous studies have documented that estrogens were able to
directly or indirectly induce dysfunction/injury of the
endothelium, resulting in thrombosis and atherosclerosis (10–12).
Functionally, estrogens display their cellular and biological
actions through binding to ERs (13). To date, two distinct ER isoforms
(i.e., ERα and ERβ) have been identified, and several variants for
each isoform have been discovered in humans or other mammals
(14,15). Studies over the last decade have
shown that the effects of estrogens are dependent on the receptor
isoform as well as on the ratio of ER isoforms or the variants
expressed in the target cells (16–18).
In the cardiovascular system, estrogens can significantly impact
cardiovascular functions (19,20)
and ERβ may play a major role in the regulation of vascular
function and blood pressure although the mechanism remains to be
elucidated (21). In prostate
cancer cells such as LAPC-4 and LNCaP cells, ERβ was highly
expressed, while ERα was relatively low or undetectable (7,22).
Thus, ERβ could mediate the direct actions of estrogens in these
prostate cancer cells (6,7,23).
Our previous data showed that estrogens acting on ERs produced a
receptor-ligand and receptor-isoform specific modulation of
androgen actions on gene expression and cell growth in prostate
cancer cells (6,7,24).
In this study, we further compared the receptor-ligand and
receptor-isoform specificity of estrogen receptor ligands in the
modulation of dihydrotestosterone (DHT) actions in prostate cancer
cells and endothelial cells, aiming to develop novel therapeutic
agents for prostate cancer therapy with minimized cardiovascular
side-effects.
Materials and methods
Reagents
Dihydrotestosterone (DHT), 17β-estradiol (βE2),
17α-estradiol (αE2), diethylstilbestrol (DES), genistein and
tamoxifen were purchased from Sigma Co. (St. Louis, MO, USA) and
dissolved in absolute ethanol at 10−2 M stock solutions.
4,4′,4″-(4-Propyl-[1H]-pyrazole-1,3,5-triyl) trisphenol (PPT) and
diarylpropionitrile (DPN) were obtained from Tocris Bioscences
(Minneapolis, MN, USA). ICI182780 (ICI), a pure estrogen
antagonist, was kindly provided by Dr A.E. Wakeling of Zeneca
Pharmaceuticals (Macclesfield, UK). Reagents for real-time PCR were
purchased from Invitrogen (Carlsbad, CA, USA). Antibodies against
ERα (HC-20: sc-543) and ERβ (N-19: SC-6820) were obtained from
Santa Cruz Biotechnology (Santa Cruz, CA, USA) and antibodies
against cyclin A (cat no. C4710) and β-actin (cat no. A5316) were
obtained from Sigma Co.
Cell lines and culture
Human aortic endothelial cells (HAECs) were
purchased from Lonza Walkersville Inc. (Walkersville, MD, USA) and
grown in EGM-2 medium (Lonza) supplemented with 2% fetal bovine
serum (FBS), hydrocortisone, human epidermal growth factor, bovine
brain extract and gentamicin/amphotericin-B as described previously
(25). The HAECs with less than
four passages in the laboratory were used for the experiments. All
experiments were carried out in the EBM™-phenol red free medium
(Lonza) containing 2% stripped FBS (Gemini Bio-Products, Calabasas,
CA, USA).
Prostate cancer LAPC-4 cells, an androgen-dependent
cell line (a gift from Dr C. Sawyer of Memorial Sloan-Kettering
Cancer Center, New York, NY, USA) were cultured in Iscove’s
modified Eagle’s medium (IMEM) supplemented with 15% FBS, 2 mM
L-glutamine, 1 nM R1881, 50 U/ml of penicillin, and 50 μg/ml
of streptomycin as described previously (7,26).
R1881 was withdrawn 48 h before cell passage to conduct the
experiments (6). LNCaP cells were
cultured in RPMI-1640 medium (Sigma) with supplements as previously
described (24). All cells were
cultured at 37°C in a 5% CO2, 95% air-humidified
atmosphere incubator.
Cell viability assay
To determine cell viability after treatment with
different steroids, HAECs, LAPC-4 and LNCaP cells were plated in
96-well plates at a density of approximately 25% in EBM-phenol
red-free medium (Lonza) containing 2% stripped FBS, or in phenol
red-free IMEM supplemented with 5% stripped FBS, or in phenol
red-free RPMI-1640 medium supplemented with 5% stripped FBS,
respectively. Twenty-four hours after plating, cells were treated
with various hormones alone or in combination as indicated in each
experiment. The concentrations of hormones and treatment durations
were selected based on previous studies in these cells (6,24,25).
The number of viable cells was determined using Cell Titer
96® Aqueous One Solution Cell Proliferation Assay kit
from Promega (Madison, WI, USA) according to the manufacturer’s
instruction.
Reverse transcriptase-polymerase chain
reaction (RT-PCR) and quantitative RT-PCR
To determine gene expression, RT-PCR and qRT-PCR
were performed. Briefly, total cellular RNA was isolated using
TriPure reagents (Roche Diagnostic Inc., Indianapolis, IN, USA),
and the concentration of RNA was quantified using the ultraviolet
absorbance at 260 nm. cDNA was synthesized following the protocol
from Invitrogen with 1 μg of total cellular RNA, and PCR was
carried out according to the protocol from Promega in a PCR mixture
containing 1.5 mM MgCl2, 0.5 μM of each primer,
200 μM dNTPs, 2.5 units of GoTaq® Flexi DNA
polymerase (Promega) and 2.5 μl of cDNA. The primers used
are listed in Table I. The PCR
conditions were 94°C for 2 min, and then 35 cycles of 94°C for 30
sec, 63°C for 30 sec for ERα or 60°C for 30 sec for ERβ, 72°C for
30 sec, and a final extension of 72°C for 5 min. The PCR products
were then fractionated in a 2% agarose gel and visualized by
ethidium bromide staining. pSG5-ERα and pSG5-ERβ expression
plasmids were used as positive controls, and yeast tRNA was used as
a negative control.
 | Table I.Primers for RT-PCR and qRT-PCR. |
Table I.
Primers for RT-PCR and qRT-PCR.
| Gene | Primers |
|---|
| ERα | F:
5′-ATGAGAGCTGCCAACCTTTG-3′ |
| R:
5′-AGAAATGTGTACACTCCAGAAT-3′ |
| ERβ | F:
5′-GATGAGGGGAAATGCGTAGA-3′ |
| R:
5′-CTTGTTACTCGCATGCCTGA-3′ |
| PSA | F:
5′-TTGTCTTCCTCACCCTGTCC-3′ |
| R:
5′-CAGGGTTGGGAATGCTTCT-3′ |
| GAPDH | F:
5′-GAAGGTGAAGGTCGGAGTC-3′ |
| R:
5′-GAAGATGGTGATGGGATTTC-3′ |
qRT-PCR was performed using the comparative Ct
method according to the instructions from the manufacturer on the
ABI Prism 7900 Sequence Detection System (Applied Biosystems,
Foster City, CA, USA) in our institutional core facility as
described previously (6).
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an
internal control. The difference between samples was calculated
following the instructions of the manufacturer (Applied
Biosystems).
Protein extraction and western blot
analysis
Protein extraction and western blot analysis were
performed as described previously (25,27)
with minor modifications. Briefly, LAPC-4 cells treated with
various agents as indicated in each experiment were harvested for
total cellular protein extraction using the passive lysis buffer
from Promega. The protein concentrations were determined using the
Bio-Rad Protein Assay kit following the manufacturer’s instruction
(Bio-Rad, Hercules, CA, USA). Equal amounts (20 μg) of total
cellular proteins were fractionated on a 10% SDS-PAGE and
transferred to a nitrocellulose membrane (Amersham Pharmacia
Biotech, Piscataway, NJ, USA). The membrane was blocked with TBS-T
buffer [500 mM NaCl, 20 mM Tris-HCl (pH 7.4) and 0.1% Tween-20]
containing 5% non-fat dry milk overnight at 4°C and then incubated
with specific antibodies against ERα (1:200) or ERβ (1:400) or
cyclin A (1:1,000) in TBS-T buffer containing 5% non-fat dry milk
for 2 h at room temperature. Following the secondary antibody
incubation (1:2,000), the positive signal was visualized using the
SuperSignal West Pico Chemiluminescent kit (Pierce Biotechnology
Inc., Rockford, IL, USA) and exposed to Kodak X-Max film. β-actin
was used as an internal control. The specific signals of ERα, ERβ,
cyclin A and β-actin were quantified using Image J (NIH, Bethesda,
MD, USA). The data are presented as fold changes of the control
after normalizing with β-actin levels.
Construction of small interference RNA
(siRNA) and gene transfection
To knockdown ERβ expression in cells, we used ERβ
siRNA and gene transfection. We first searched GenBank for human
ERβ gene sequences (GenBank accession no. NM_001437) and designed a
custom stealth RNAi oligonucleotide at 25 base pairs in length
(Invitrogen). The sequence for ERβ was
5′-GUCAAGGCCAUGAUCCUGCUCAAUU-3′ and the control siRNA was
5′-CCAUGGCGCCAAUUCCAAACA GUUU-3′. For RNAi transfection, LAPC-4
cells were seeded in a 96-well plate or a 6-well plate in phenol
red-free IMEM medium containing 5% stripped FBS without
antibiotics. Twenty-four hours later, the cells were transfected
with various concentrations of siRNA using Lipofectamine 2000 (0.25
μl/well in 96-well plate) according to the instruction from
the manufacturer (Invitrogen) in OPTI-MEM medium. Sixteen hours
after transfection, transfection reagents were replaced with normal
medium and cells were treated with various hormones for 72 h as
indicated in each experiment. At the end of the experiments, the
number of viable cells was determined using the cell viability
assay described above. The efficiency of lipofectamine was tested
before siRNA transfection, and the knockdown of ERβ expression was
verified using western blot analysis.
Statistical analysis
The data are presented as the mean ± SE of the mean
(SEM). One-way analysis of variance (ANOVA) followed by a post hoc
Student-Newman-Keuls test was used to determine the difference
among multiple groups. A p<0.05 was considered as statistically
significant.
Results
DHT induction of endothelial and prostate
cancer cell proliferation and PSA expression
In this study, we first assessed the effect of DHT
on regulation of both endothelial HAEC and prostate cancer cell
proliferation. We found that consistent with our previous studies
(6,7,24,25),
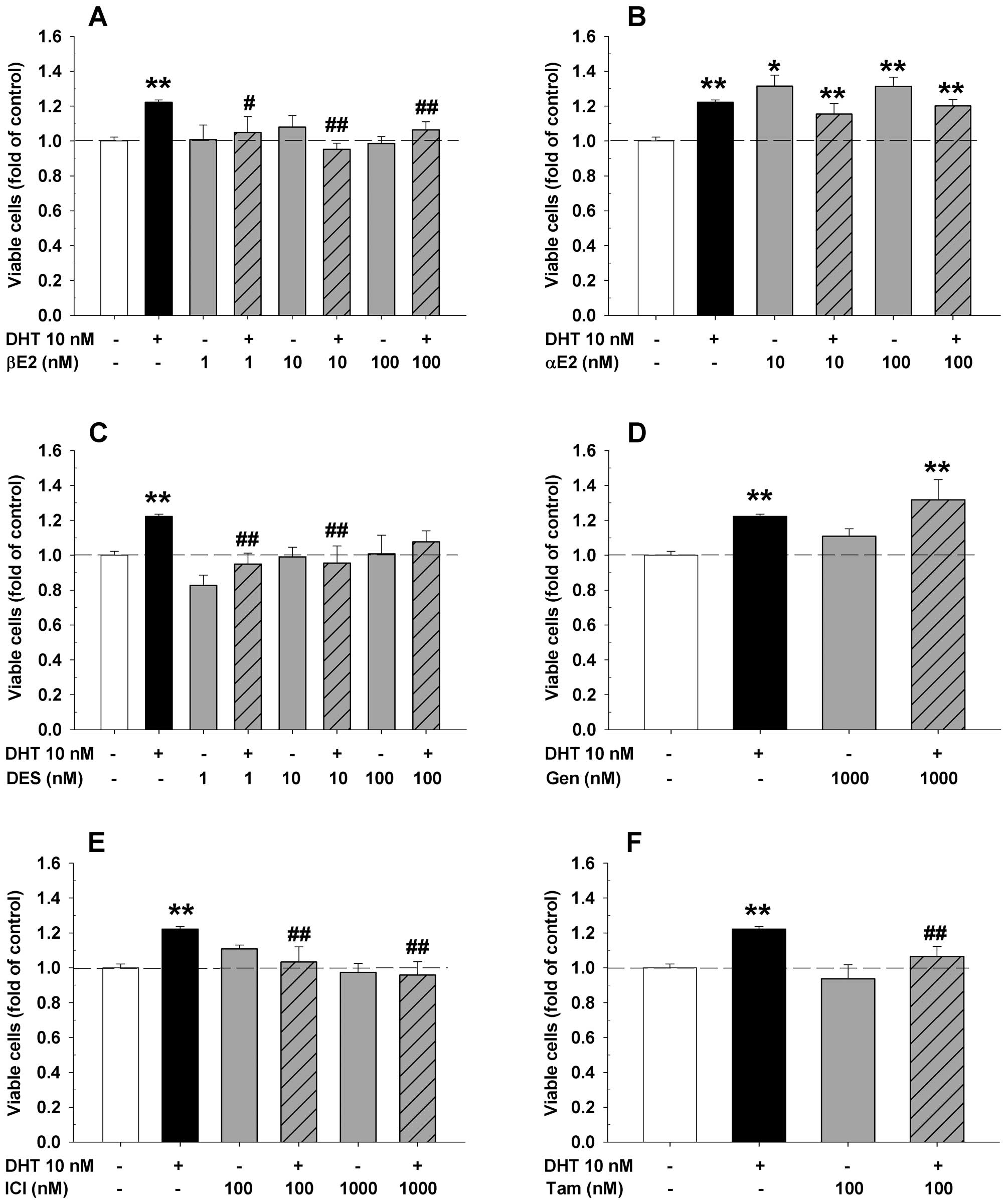
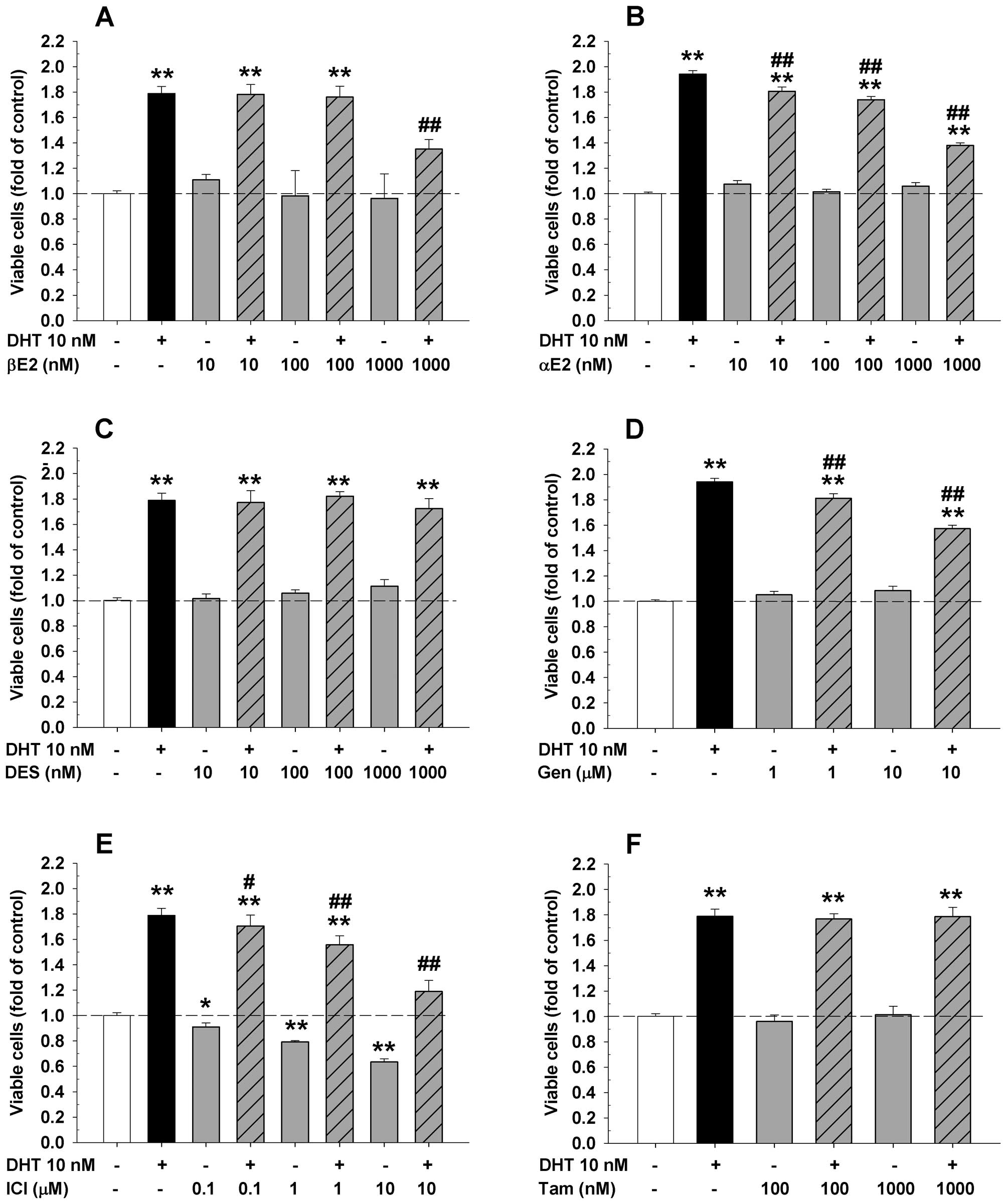
DHT significantly increased viable cell numbers of HAECs (Fig. 1), LAPC-4 (Fig. 2), and LNCaP cells (Fig. 3). Compared to the corresponding
controls, cell proliferation was significantly increased to
approximately 22% (48 h), 94% (72 h) and 38% (144 h) in endothelial
HAECs (Fig. 1), and prostate
cancer LAPC-4 cells (Fig. 2)
treated with 10 nM DHT, and in prostate cancer LNCaP cells
(Fig. 3) treated with 0.1 nM DHT,
respectively. Moreover, treatment with DHT at 10 nM for 72 h
induced PSA mRNA expression by approximately 40-fold in LAPC-4
cells (Fig. 4).
Differential effects of ER ligands on
DHT-induced cell proliferation in endothelial HAEC and prostate
cancer LAPC-4 and LNCaP cells
To determine the effects of various ER ligands on
the regulation of DHT-induced cell proliferation in endothelial and
prostate cancer cells, endothelial HAECs or prostate cancer cells
were seeded in 96-well plates and treated with DHT plus or minus
various concentrations of αE2, βE2, DES, ICI, genistein, and
tamoxifen (Figs. 1–3) for 48 or 72 h, respectively. As shown
in Figs. 1 and 2, treatment with βE2, DES, genistein or
tamoxifen alone did not significantly affect the cell proliferation
of both endothelial HAECs (Fig. 1)
and prostate cancer LAPC-4 cells (Fig.
2). However, administration of αE2 significantly increased cell
proliferation in HAEC cells (Fig.
1B), but did not have any effect in LAPC-4 cells (Fig. 2B). Treatment with ICI alone did not
significantly affect HAEC cell proliferation (Fig. 1E), but decreased LAPC-4 cell
proliferation in a dose-dependent manner (Fig. 2E). In LNCaP prostate cancer cells,
treatment with either αE2 or genistein alone did not affect the
cell proliferation (Fig. 3B and
C), while βE2 significantly increased the viable cell number
(Fig. 3A).
When ER ligands were administrated concomitantly
with DHT, ER ligands produced a differential regulation of
DHT-induced cell proliferation in a cell type-dependent manner. For
example, βE2 and ICI produced a dose-dependent inhibition of
DHT-induced cell proliferation in both HAECs (Fig. 1A and E) and LAPC-4 cells (Fig. 2A and E), while αE2 and genistein
significantly attenuated DHT-induced cell proliferation in LAPC-4
cells (Fig. 2B and D) without
significantly altering the DHT-induced cell proliferation in HAEC
cells (Fig. 1B and D). DES and
tamoxifen attenuated DHT-induced cell proliferation in HAEC cells
(Fig. 1C and F), but had no effect
on DHT-induced cell proliferation in LAPC-4 cells (Fig. 2C and F). Similar inhibition of
DHT-induced cell proliferation by αE2 and genistein was observed in
LNCaP cell (Fig. 3B and C).
However, the addition of βE2 did not affect DHT-induced cell
proliferation since treatment with βE2 alone greatly induced cell
growth in LNCaP cells (Fig.
3A).
Co-administration of ER ligands also produced a
ligand-specific modulation of DHT-induced PSA expression in LAPC-4
cells (Fig. 4), consistent with
our previous studies (7,24). Specifically, the DHT-induced PSA
mRNA expression in LAPC-4 cells was significantly inhibited by αE2
and βE2 (Fig. 4A and B) but not by
ICI or tamoxifen at the doses tested (Fig. 4C and D).
The role of ERβ in βE2 modulation of
DHT-induced cell proliferation in LAPC-4 cells
To investigate whether the effects of estrogens are
mediated through ERs, we first determined the mRNA and protein
levels of ERα and ERβ in HAEC and LAPC-4 cells using RT-PCR
(Fig. 5A) and western blot
analysis (Fig. 5B), respectively.
As shown in Fig. 5, ERβ was highly
expressed, whereas ERα expression was quite low or undetectable in
both HAEC and LAPC-4 cells. As a positive control, ERα was
expressed in MCF-7 cells (28)
(Fig. 5B).
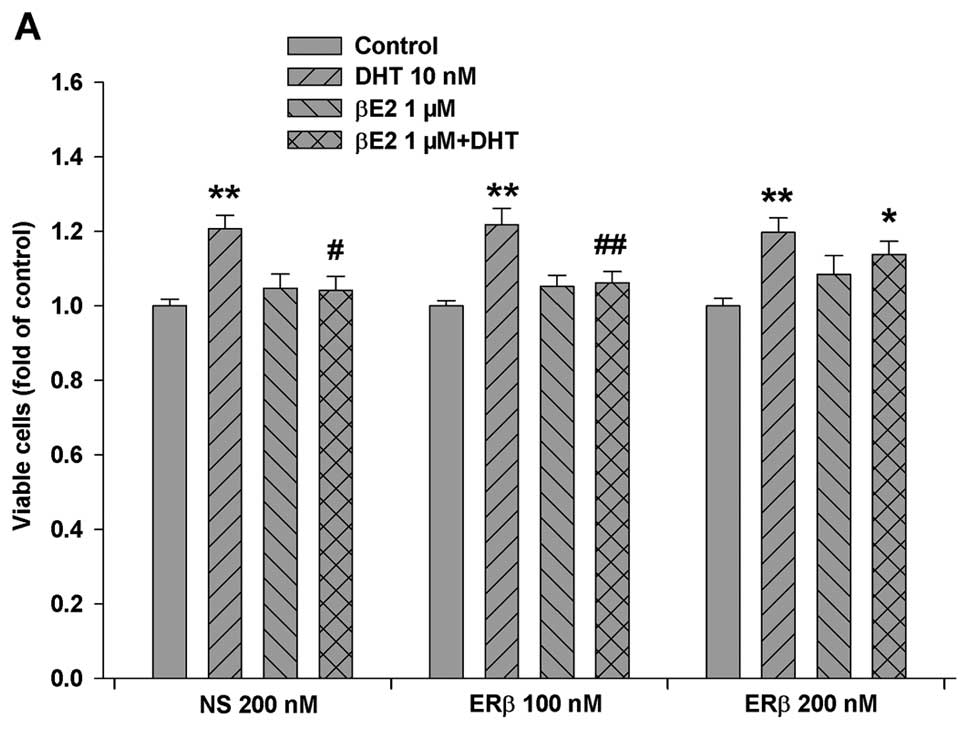
Based on this information, we knocked down ERβ
expression in LAPC-4 cells by transfection of a specific ERβ siRNA.
As shown in Fig. 6, transfection
of a specific ERβ siRNA produced a dose-dependent decrease in ERβ
protein expression, and the knockdown of ERβ partially eliminated
the βE2 inhibition of DHT-induced cell proliferation in LAPC-4
cells (Fig. 6A).
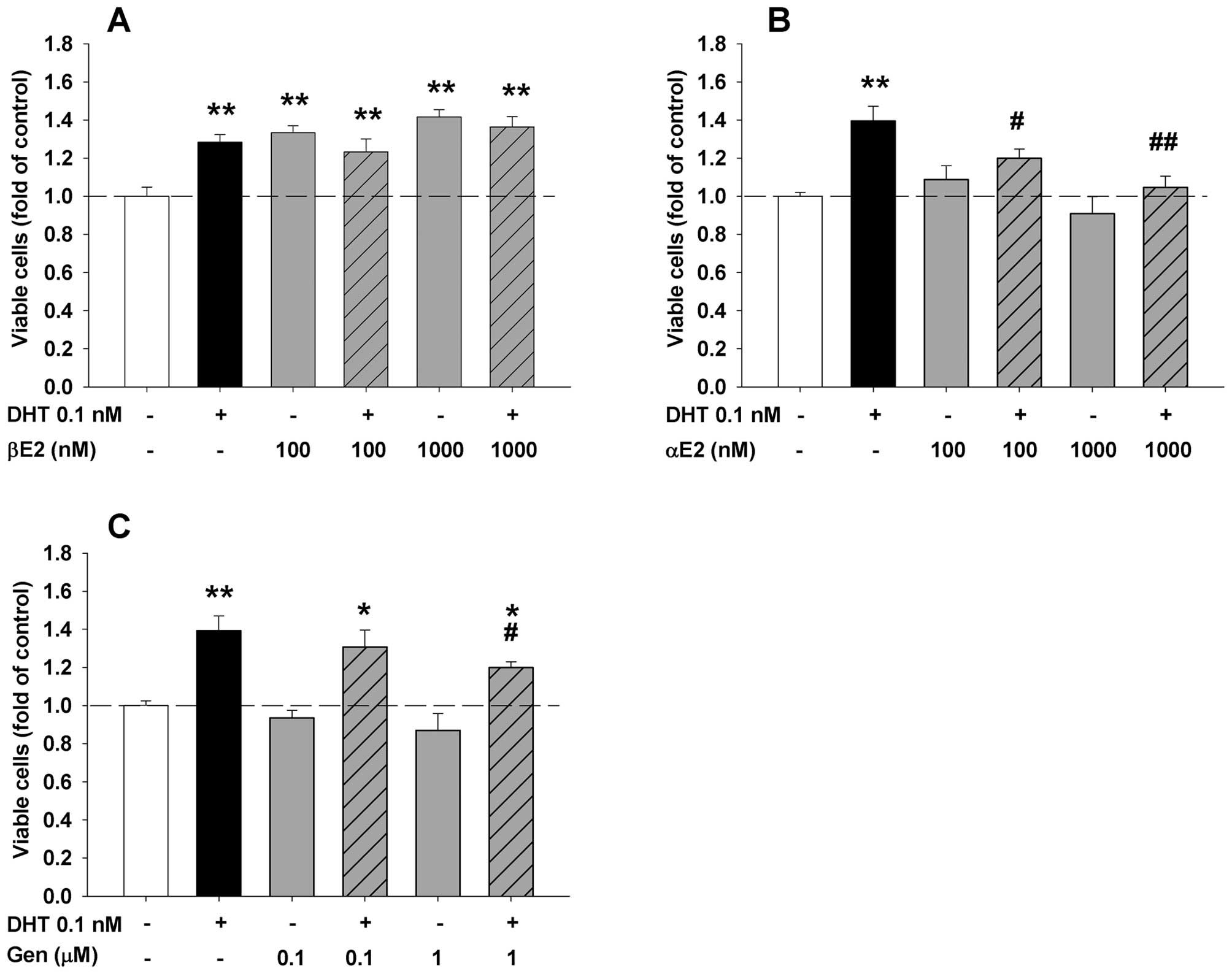
To explore whether a specific activation of ERβ is
sufficient to inhibit DHT-induced LAPC-4 cell proliferation, the
cells were treated with DHT plus or minus a specific ERα or ERβ
agonist. As expected, the addition of an ERα specific agonist, PPT,
failed to affect the DHT-induced cell proliferation at the doses
ranging from 0.1 to 1,000 nM (Fig.
7B). Surprisingly, the concomitant administration of an ERβ
specific agonist, DPN, did not inhibit the DHT action while it
slightly but significantly potentiated DHT-induced cell
proliferation at a 1 μM dose (Fig. 7A). Of note, treatment with DPN
alone produced a dose-dependent biphasic effect in LAPC-4 cells. At
low doses from 0.1 to 1 nM, it slightly but significantly decreased
cell proliferation while at a high dose of 1 μM, it
significantly increased cell proliferation (Fig. 7A). In contrast, both PPT and DPN at
doses of 100 nM and 1 μM completely blocked DHT-induced cell
proliferation in LNCaP cells as shown in Fig. 7C and D. Treatment with PPT or DPN
alone did not significantly alter the cell proliferation in LNCaP
cells.
Parallel changes in estrogen modulation
of DHT-induced cyclin A expression and cell proliferation in LAPC-4
cells
Previous studies demonstrate that cyclin A
expression is induced by DHT, which is related to DHT-induced cell
proliferation in both HAECs (25)
and LAPC-4 cells (6). To decipher
the possible molecular events responsible for the differential
effects of ER ligands on the modulation of DHT-induced cell
proliferation in LAPC-4 cells, we assessed the cyclin A expression
after treating LAPC-4 cells with DHT and various ER ligands alone
or in combination. As shown in Fig.
8, treatment of LAPC-4 cells with 10 nM DHT for 72 h
significantly upregulated the expression of cyclin A. This DHT
effect was significantly attenuated by the co-administration of
αE2, βE2 or ICI, but not by tamoxifen at the doses used, resulting
in changes parallel to the modulation of DHT-induced LAPC-4 cell
proliferation (Fig. 2).
Discussion
It has been documented that a major side-effect of
androgen deprivation therapy of prostate cancer especially using
estrogens is the development of thrombosis and cardiovascular
events (8,9,29).
The development of new therapeutic strategies and/or agents with
minimal side-effects for the androgen deprivation therapy of
advanced prostate cancer has been a continuing effort of the
scientists around the world for the last 6 decades. With the
discovery of ERβ and the elucidation of various ER ligand
conformations, it is getting clearer that the effects of ER ligands
are dependent not only on the receptor ligands but also on the
receptor isoforms (7,16–18).
Based on the recent findings that ER ligands can directly modulate
androgen actions in prostate cancer cells in a receptor-ligand and
receptor-isoform specific manner (6,7,24)
and that androgens can directly stimulate endothelial cell
proliferation in a gender-specific manner (25,30),
we have compared the effects of various ER ligands on the
modulation of androgen actions between endothelial HAECs and
prostate cancer cells in the present study. Our data demonstrated
that different ER ligands had differential effects on the
regulation of DHT-inducted cell proliferation in both HAECs and
LAPC-4 cells, presumably mediated through ERβ and associated with
their modulation of DHT-induced cyclin A expression. These findings
provide the first evidence that the effects of ER ligands in
endothelial HAECs and prostate cancer cells could be dissociated
and support the feasibility for the development of a novel
therapeutic agent for anti-androgen therapy of prostate cancer with
minimal cardiovascular side-effects.
Interactions between androgens and estrogens via
their corresponding receptors play an important role in prostate
and endothelial physiology and pathophysiology (6,7,31,32).
The biological outcome of this hormonal interaction is not only
receptor-ligand and receptor-isoform specific (6,7,25)
but also cell specific as demonstrated in our current study.
Consistent with previous reports (6,24,25),
we have observed that treatment with DHT stimulated cell
proliferation in HAECs (Fig. 1)
and induced PSA gene expression (Fig.
4) and cell proliferation in LAPC-4 (Fig. 2) and LNCaP cells (Fig. 3). This DHT induced cell
proliferation in endothelial HAECs and prostate LAPC-4, LNCaP
cancer cells is differentially modulated by ER ligands in a
cell-dependent manner. In HAECs, βE2, DES, ICI and tamoxifen
blocked the DHT-induced cell proliferation, whereas αE2 and
genistein did not have such effect. Interestingly, treatment with
αE2 alone significantly increased the cell proliferation in HAECs,
a potential beneficial effect in the endothelium to repair
endothelial damage/injury. On the other hand, both αE2 and
genistein inhibited DHT-induced cell proliferation in LAPC-4 and
LNCaP prostate cancer cells while βE2 only attenuated the
DHT-induced cell proliferation in LAPC-4 cells without any
inhibition of DHT-induced cell proliferation in LNCaP cells.
Moreover, treatment with βE2 alone in LNCaP cells significantly
increased cell proliferation, probably via transactivation of the
mutant AR in LNCaP cells (24).
Taken together, our current results and previous reports strongly
indicate that the modulation of DHT actions by ER ligands is
receptor-ligand, receptor-isoform and cell-specific. Based on the
cell specificity of ER ligands on the modulation of DHT actions, ER
ligands are categorized to three different categories (Table II), which would be informative for
the development of ER ligands in the treatment of prostate
cancer.
| Table II.Classification of ER-ligands based on
modulation of AR activity in HAECs and LAPC-4 cells. |
Table II.
Classification of ER-ligands based on
modulation of AR activity in HAECs and LAPC-4 cells.
| Categories | ER ligands | Modulation of AR
activity
|
|---|
| HAECs | LAPC-4 cells |
|---|
| I | βE2, ICI | ↓ | ↓ |
| II | αE2, genistein | ↑/- | ↓ |
| III | DES, tamoxifen | ↓ | - |
It is noteworthy that the ER ligand specificity in
modulation of DHT actions is unparallel or unrelated to the
pharmacological categorization. For instance, ICI, a pure ER
antagonist, not only blocked the DHT-induced cell proliferation in
HAECs and LAPC-4 cells, but also inhibited cell growth by itself in
LAPC-4 cells. Although the molecular mechanisms of ICI actions
remain to be further elucidated, downregulation of AR gene
expression (33) and direct
inhibition of AR transactivational activity (7) may account, at least in part, for
these actions. Tamoxifen, a partial ER agonist/antagonist or a
selective ER modulator, completely blocked the DHT-induced cell
proliferation in HAECs, but did not affect the DHT-induced cell
proliferation and PSA expression in prostate cancer cells, which
could partially explain the ineffectiveness of tamoxifen in the
treatment of prostate cancer in clinical trials (16,34).
Surprisingly, DES, an ER agonist and an agent used effectively for
androgen deprivation therapy of prostate cancer in the clinic, did
not display any inhibitory effect on DHT-induced LAPC-4 prostate
cancer cell proliferation, whereas it completely blocked
DHT-induced cell growth in HAECs at low nanomolar concentrations.
These data suggest that the antitumor effects of DES may be mainly
mediated through the negative feedback of
hypothalamus-pituitary-gonadal axis to inhibit testosterone
biosynthesis without a direct inhibition of DHT action in the tumor
cells, and those patients treated with DES may be more susceptible
to cardiovascular side-effects (8,9) due
to its inhibition of DHT-induced endothelial cell growth.
The genomic effects of estrogens are mainly mediated
through the transactivation of ERs, ERα and ERβ in the cells.
Although the modulation of DHT effects by estrogens can be mediated
through either ERα or ERβ as previously reported (7,35),
the estrogen modulation of DHT induction of LAPC-4 cell
proliferation was most likely mediated through ERβ as supported by
previous studies (7,22) and our current demonstrations. In
the present study, we have observed that both LAPC-4 and HAEC cells
expressed high levels of ERβ mRNA and protein, while the expression
of ERα was quite low or undetectable. Moreover, knockdown of ERβ
expression using a specific siRNA largely abolished the effect of
βE2 on the inhibition of DHT-induced LAPC-4 cell proliferation.
However, an activation of ERβ by a specific ligand is not
sufficient to produce inhibition of DHT actions in LAPC-4 cells
since DPN, a specific ERβ agonist (36), did not inhibit, but slightly
potentiate DHT-induced cell proliferation in LAPC-4 cells, further
indicating the receptor-ligand specificity in the modulation of DHT
actions in this system.
The observation that both PPT, an ERα specific
agonist (38), and DPN, an ERβ
specific agonist, significantly blocked DHT-induced cell
proliferation in LNCaP, but not in LAPC-4 cells is unexpected. Like
LAPC-4 cells, LNCaP cells also mainly express ERβ while ERα
expression is quiet low or undetectable (7,22).
Unlike LAPC-4 cells that express a wild-type AR, the AR in LNCaP
cells is mutated, resulting in a wide-spectrum of ligand binding to
the receptor (38). It is
therefore most likely that both PPT and DPN may bind to the mutant
AR and function as an AR antagonist to block DHT actions. This
hypothesis is currently under investigation in the laboratory.
How different ER ligands produce a differential
regulation of DHT actions in a cell-dependent manner is currently
unknown. Previous studies have clearly demonstrated that different
ER ligands led to different conformational changes in ERs (39–41),
resulting in a differential recruitment of transcriptional factors
and/or co-regulators to control the biological activity of the
cells (10,37,42,43).
This principle also applies in androgen-estrogen interaction
(7,35). In this context, our current results
suggest that based on the cell-dependent differential modulation of
androgen actions by ER ligands and the elucidation of their
molecular mechanisms, it would be possible to develop therapeutic
agents that have great effects on prostate cancer with minimal
cardiovascular side-effects. Thus, further investigation of
androgen-estrogen interaction in other endothelial and prostate
cancer cells, in animal models and eventually in clinical trials is
warranted.
It is well documented that regulation of the cell
cycle plays an essential role in cell proliferation,
differentiation, and cell death (44,45).
Cyclin A is a key regulator in cell cycle progression, especially
in the G1/S transition (45). Indeed, previous studies have shown
that cyclin A is overexpressed in prostate cancer cells (46) and tumor tissues (47). In the present study, we observed
that DHT induced cyclin A expression in LAPC-4 cells, consistent
with our previous demonstrations in LAPC-4 (6) and HAEC cells (25). Notably, this DHT-induced cyclin A
expression is also differentially modulated by ER ligands in a
manner parallel to their modulation of DHT-induced cell
proliferation, suggesting that cyclin A might be a downstream
molecular target of androgen-estrogen interaction in the control of
cell proliferation.
It is worthwhile to emphasize that αE2, a
stereoisomer of βE2, binds weakly to ER to form an αE2-ER complex
that only transiently binds to the estrogen-responsive element
(48), resulting in significantly
less feminizing effects than βE2. Compared to βE2, αE2 has no
carcinogenic effect in a mammalian model system (49), and has little effect on the
vascular smooth muscle (50).
However, αE2 can protect neuronal cells from ischemic damage as
potently as βE2 (51). Unlike
other ER ligands, we found that αE2 was able to specifically induce
growth of HAECs, while it blocked DHT-induced prostate tumor cell
proliferation and inhibited tumor growth in prostate cancer
xenograft mice (5,7,24).
Although the mechanism responsible for αE2 stimulation of HAEC
growth remains to be determined, this αE2 action could help
maintain endothelial homeostasis. Taken together, these data
suggest that αE2 is superior to other ER ligands for prostate
cancer therapy since it blocks AR-dependent prostate gene
expression, prostate tumor cell proliferation and tumor growth,
while it stimulates HAEC growth, a potential beneficial action on
protection of endothelium and on minimizing cardiovascular
side-effects of anti-androgen therapy.
In summary, using endothelial HAECs and prostate
cancer LAPC-4 and LNCaP cells as the model system, we have
demonstrated that DHT-induced cell proliferation and gene
expression are differentially modulated by ER ligands in a
cell-specific manner. Further exploration of this hormonal
interaction in other model systems and the elucidation of the
molecular mechanisms will facilitate the development of effective
therapeutic agent(s) for the prostate cancer therapy with minimal
cardiovascular side-effects.
Abbreviations:
|
DHT
|
dihydrotestosterone
|
|
DPN
|
diarylpropionitrile
|
|
ER
|
estrogen receptor
|
|
HAECs
|
human aortic endothelial cells
|
|
IMEM
|
Iscove’s modified Eagle’s medium
|
|
PPT
|
4,4′,4″-(4-Propyl-[1H]-pyrazole-1,3,5-triyl) trisphenol
|
|
PSA
|
prostate-specific antigen
|
Acknowledgements
We would like to thank Dr C. Tan for
his technical advice and Dr X. Xing for help of the manuscript
preparation. We also thank Dr C. Sawyer of Memorial Sloan-Kettering
Cancer Center for providing LAPC-4 cells and Dr Wakeling of Zeneca
Pharmaceuticals for providing ICI182780 compound. This study was
supported in part by a Grant-in-Aid from National Institutes of
Health (NIH) (UL1 RR024996) and a Grant-in-Aid from the National
Natural Science Foundation of China (no. 30873126).
References
|
1.
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2.
|
Djulbegovic M, Beyth RJ, Neuberger MM,
Stoffs TL, Vieweg J, Djulbegovic B and Dahm P: Screening for
prostate cancer: systematic review and meta-analysis of randomised
controlled trials. BMJ. 341:c45432010. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Ginzburg S and Albertsen PC: The timing
and extent of androgen deprivation therapy for prostate cancer:
weighing the clinical evidence. Endocrinol Metab Clin North Am.
40:615–623. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Huggins C and Hodges CV: Studies on
prostatic cancer I. The effect of castration, of estrogen and of
androgen injection on serum phosphatases in metastatic carcinoma of
the prostate. Cancer Res. 1:293–297. 1941.
|
|
5.
|
Dowling AJ and Tannock IF: Systemic
treatment for prostate cancer. Cancer Treat Rev. 24:283–301. 1998.
View Article : Google Scholar
|
|
6.
|
Qiao Y, Zhang ZK, Cai LQ, Tan C,
Imperato-McGinley JL and Zhu YS: 17alpha-estradiol inhibits LAPC-4
prostatic tumor cell proliferation in cell cultures and tumor
growth in xenograft animals. Prostate. 67:1719–1728. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Zhu YS, Cai LQ, Huang Y, Fish J, Wang L,
Zhang ZK and Imperato-McGinley JL: Receptor isoform and
ligand-specific modulation of dihydrotestosterone-induced prostate
specific antigen gene expression and prostate tumor cell growth by
estrogens. J Androl. 26:500–508. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
The Coronary Drug Project: Findings
leading to discontinuation of the 2.5-mg day estrogen group. The
coronary Drug Project Research Group. JAMA. 226:652–657. 1973.
View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
The Veterans Administration Co-operative
Urological Research Group: Treatment and survival of patients with
cancer of the prostate. Surg Gynecol Obstet. 124:1011–1017.
1967.PubMed/NCBI
|
|
10.
|
Ling S, Komesaroff P and Sudhir K:
Cellular mechanisms underlying the cardiovascular actions of
oestrogens. Clin Sci (Lond). 111:107–118. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Ouyang P, Michos ED and Karas RH: Hormone
replacement therapy and the cardiovascular system lessons learned
and unanswered questions. J Am Coll Cardiol. 47:1741–1753. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Ross R: Atherosclerosis - an inflammatory
disease. N Engl J Med. 340:115–126. 1999. View Article : Google Scholar
|
|
13.
|
Moolman JA: Unravelling the
cardioprotective mechanism of action of estrogens. Cardiovasc Res.
69:777–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Leung YK, Mak P, Hassan S and Ho SM:
Estrogen receptor (ER)-beta isoforms: a key to understanding
ER-beta signaling. Proc Natl Acad Sci USA. 103:13162–13167. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Nilsson S and Gustafsson JÅ: Estrogen
receptors: therapies targeted to receptor subtypes. Clin Pharmacol
Ther. 89:44–55. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Matthews J and Gustafsson JA: Estrogen
signaling: a subtle balance between ER alpha and ER beta. Mol
Interv. 3:281–292. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Sotoca AM, van den Berg H, Vervoort J, et
al: Influence of cellular ERalpha/ERbeta ratio on the
ERalpha-agonist induced proliferation of human T47D breast cancer
cells. Toxicol Sci. 105:303–311. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Stein S, Zoltick B, Peacock T, et al:
Phase II trial of toremifene in androgen-independent prostate
cancer: a Penn cancer clinical trials group trial. Am J Clin Oncol.
24:283–285. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Babiker FA, De Windt LJ, van Eickels M,
Grohe C, Meyer R and Doevendans PA: Estrogenic hormone action in
the heart: regulatory network and function. Cardiovasc Res.
53:709–719. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Rossouw JE, Anderson GL, Prentice RL, et
al: Risks and benefits of estrogen plus progestin in healthy
postmenopausal women: principal results from the Women’s Health
Initiative randomized controlled trial. JAMA. 288:321–333.
2002.PubMed/NCBI
|
|
21.
|
Zhu Y, Bian Z, Lu P, et al: Abnormal
vascular function and hypertension in mice deficient in estrogen
receptor beta. Science. 295:505–508. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Lau KM, LaSpina M, Long J and Ho SM:
Expression of estrogen receptor (ER)-alpha and ER-beta in normal
and malignant prostatic epithelial cells: regulation by methylation
and involvement in growth regulation. Cancer Res. 60:3175–3182.
2000.
|
|
23.
|
Nakajima Y, Akaogi K, Suzuki T, et al:
Estrogen regulates tumor growth through a nonclassical pathway that
includes the transcription factors ERβ and KLF5. Sci Signal.
4:ra222011.PubMed/NCBI
|
|
24.
|
Qiao Y, Wang L, Cai LQ, Tan C,
Imperato-McGinley J and Zhu YS: Inhibition of aberrant androgen
receptor induction of prostate specific antigen gene expression,
cell proliferation and tumor growth by 17α-estradiol in prostate
cancer. J Urol. 185:305–314. 2011.PubMed/NCBI
|
|
25.
|
Cai J, Hong Y, Weng C, Tan C,
Imperato-McGinley JL and Zhu YS: Androgen stimulates endothelial
cell proliferation via an androgen receptor-VEGF/cyclin A mediated
mechanism. Am J Physiol Heart Circ Physiol. 300:H1210–H1221. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Arnold JT, Liu X, Allen JD, Le H, McFann
KK and Blackman MR: Androgen receptor or estrogen receptor-beta
blockade alters DHEA-, DHT-, and E(2)-induced proliferation and PSA
production in human prostate cancer cells. Prostate. 67:1152–1162.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Zhu YS and Pfaff DW: Differential
regulation of AP-1 DNA binding activity in rat hypothalamus and
pituitary by estrogen. Mol Brain Res. 55:115–125. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Martin MB, Angeloni SV, Garcia-Morales P,
Sholler PF, Castro-Galache MD, Ferragut JA and Saceda M: Regulation
of estrogen receptor-alpha expression in MCF-7 cells by taxol. J
Endocrinol. 180:487–496. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Collins L, Mohammed N, Ahmad T and Basaria
S: Androgen deprivation therapy for prostate cancer: implications
for cardiometabolic clinical care. J Endocrinol Invest. 35:332–339.
2012.PubMed/NCBI
|
|
30.
|
Sieveking DP, Lim P, Chow RW, et al: A
sex-specific role for androgens in angiogenesis. J Exp Med.
207:345–352. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Panet-Raymond V, Gottlieb B, Beitel LK,
Pinsky L and Trifiro MA: Interactions between androgen and estrogen
receptors and the effects on their transactivational properties.
Mol Cell Endocrinol. 167:139–150. 2000. View Article : Google Scholar
|
|
32.
|
Sumanasekera WK, Sumanasekera GU,
Mattingly KA, Dougherty SM, Keynton RS and Klinge CM: Estradiol and
dihydrotestosterone regulate endothelial cell barrier function
after hypergravity-induced alterations in MAPK activity. Am J
Physiol Cell Physiol. 293:C566–C573. 2007. View Article : Google Scholar
|
|
33.
|
Bhattacharyya RS, Krishnan AV, Swami S and
Feldman D: Fulvestrant (ICI 182,780) down-regulates androgen
receptor expression and diminishes androgenic responses in LNCaP
human prostate cancer cells. Mol Cancer Ther. 5:1539–1549. 2006.
View Article : Google Scholar
|
|
34.
|
Bergan RC, Reed E, Myers CE, et al: A
phase II study of high-dose tamoxifen in patients with
hormone-refractory prostate cancer. Clin Cancer Res. 5:2366–2373.
1999.PubMed/NCBI
|
|
35.
|
Zhu YS and Imperato-McGinley JL:
5alpha-reductase isozymes and androgen actions in the prostate. Ann
NY Acad Sci. 1155:43–56. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Harrington WR, Sheng S, Barnett DH, Petz
LN, Katzenellenbogen JA and Katzenellenbogen BS: Activities of
estrogen receptor alpha- and beta-selective ligands at diverse
estrogen responsive gene sites mediating transactivation or
transrepression. Mol Cell Endocrinol. 206:13–22. 2003. View Article : Google Scholar
|
|
37.
|
Kraichely DM, Sun J, Katzenellenbogen JA
and Katzenellenbogen BS: Conformational changes and coactivator
recruitment by novel ligands for estrogen receptor-alpha and
estrogen receptor-beta: correlations with biological character and
distinct differences among SRC coactivator family members.
Endocrinology. 141:3534–3545. 2000.
|
|
38.
|
Veldscholte J, Ris-Stalpers C, Kuiper GG,
et al: A mutation in the ligand binding domain of the androgen
receptor of human LNCaP cells affects steroid binding
characteristics and response to anti-androgens. Biochem Biophys Res
Commun. 173:534–540. 1990. View Article : Google Scholar
|
|
39.
|
Brzozowski AM, Pike AC, Dauter Z, et al:
Molecular basis of agonism and antagonism in the oestrogen
receptor. Nature. 389:753–758. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Dai SY, Burris TP, Dodge JA, et al: Unique
ligand binding patterns between estrogen receptor alpha and beta
revealed by hydrogen-deuterium exchange. Biochemistry.
48:9668–9676. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Pike AC, Brzozowski AM, Hubbard RE, et al:
Structure of the ligand-binding domain of oestrogen receptor β in
the presence of a partial agonist and a full antagonist. EMBO J.
18:4608–4618. 1999.
|
|
42.
|
Margeat E, Bourdoncle A, Margueron R,
Poujol N, Cavailles V and Royer C: Ligands differentially modulate
the protein interactions of the human estrogen receptors alpha and
beta. J Mol Biol. 326:77–92. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Shang YF and Brown M: Molecular
determinants for the tissue specificity of SERMs. Science.
295:2465–2468. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Galderisi U, Jori FP and Giordano A: Cell
cycle regulation and neural differentiation. Oncogene.
22:5208–5219. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Pestell RG, Albanese C, Reutens AT, Segall
JE, Lee RJ and Arnold A: The cyclins and cyclin-dependent kinase
inhibitors in hormonal regulation of proliferation and
differentiation. Endocr Rev. 20:501–534. 1999.PubMed/NCBI
|
|
46.
|
Yasmeen A, Berdel WE, Serve H and
Muller-Tidow C: E- and A-type cyclins as markers for cancer
diagnosis and prognosis. Expert Rev Mol Diagn. 3:617–633. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Wegiel B, Bjartell A, Tuomela J, et al:
Multiple cellular mechanisms related to cyclin A1 in prostate
cancer invasion and metastasis. J Natl Cancer Inst. 100:1022–1036.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Ko YJ and Balk SP: Targeting steroid
hormone receptor pathways in the treatment of hormone dependent
cancers. Curr Pharm Biotechnol. 5:459–470. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49.
|
Li JJ, Li SA, Oberley TD and Parsons JA:
Carcinogenic activities of various steroidal and nonsteroidal
estrogens in the hamster kidney: relation to hormonal activity and
cell proliferation. Cancer Res. 55:4347–4351. 1995.PubMed/NCBI
|
|
50.
|
Freay AD, Curtis SW, Korach KS and Rubanyi
GM: Mechanism of vascular smooth muscle relaxation by estrogen in
depolarized rat and mouse aorta. Role of nuclear estrogen receptor
and Ca2+ uptake. Circ Res. 81:242–248. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
51.
|
Dykens JA, Moos WH and Howell N:
Development of 17alpha-estradiol as a neuroprotective therapeutic
agent: Rationale and results from a phase I clinical study. Ann NY
Acad Sci. 1052:116–35. 2005. View Article : Google Scholar : PubMed/NCBI
|