Introduction
Prostate cancer (PCa) is the most frequent
malignancy and a leading cause of cancer-related deaths in American
men, accounting for an estimated 233,000 (27%) new cases and 29,480
(10%) deaths in 2014 (1–3). Androgen deprivation therapy (ADT)
provides an effective therapy for patients with advanced PCa.
Although most patients with advanced PCa show an initial response
to ADT, a significant percentage of patients invariably progress to
hormone refractory prostate cancer (HRPC) and castration-resistant
prostate cancer (CRPC) (4,5). Effective targeting treatment for
advanced CRPC and HRPC remains a critical clinical challenge. Thus,
there is a clear need for novel targeted therapeutic strategies for
the treatment of advanced PCa. siRNAs have provided us with clues
how to tackle this issue. The properties of siRNA that are
attractive for therapeutics include: i) stringent target-gene
specificity, ii) relatively low immunogenicity of siRNA and iii)
simplicity of design and testing of siRNA (6).
The HSP70 family includes at least eight members
with diverse biochemical functions, including nascent protein
folding, preventing denatured protein aggregation, and modulating
assembly/disassembly of protein complexes (7,8). The
exact role of the HSP70 family in cancer remains to be elucidated.
Firstly, the two major cytoplasmic isoforms are HSC70 and HSP72.
Generally, HSC70 is abundantly and ubiquitously expressed in
nontumor tissues, whereas HSP72 is present at relatively low levels
in the absence of stress. Secondly, HSC70 and HSP72 expression may
reduce sensitivity to HSP90 inhibitors. HSP70 as a survival tool
for cancer cells. Targets of HSP70 could be upstream and downstream
of mitochondria. However, targeting HSP70 or androgen receptor
alone can result in off-target effets, drug resistance and disease
recurrence. Combinatorial modulation of these two HSP70 isoforms
could therefore be doubly advantageous (9–15).
However, the specific and efficient delivery of
siRNA into cancer cells in vivo remains a major obstacle
(16). The scientific researchers
try to develop a technique that employs UTMD to deliver genes to
specific cells or tissues in a multi-model approach. Microbubble
contrast agents are emerging nonviral vector systems capable of
binding nucleic acids, and that may overcome some of the
limitations of current siRNA delivery systems. The use of
microbubbles as nucleic acid delivery vectors is based on the
observation that the rupture of microbubbles via UTMD results in
deposition of microbubble shell components and increased cell
membrane permeability localized to the site of
microbubble-ultrasound interaction. This phenomenon has been
exploited to deliver drug and genes both in vitro and in
vivo. We and others have recently shown that UTMD can deliver
therapeutic genes to tumors (17–21).
In the present study, we sought to use UTMD to
achieve RNAi as a strategy prostate cancer cells growth
suppression. Specifically, we tested the hypothesis that
UTMD-mediated delivery of dual targeting of HSC70 and HSP72 siRNA
will inhibits HSP90 function, induce extensive tumor-specifc
apoptosis.
Materials and methods
Cell and cell culture
The human prostate carcinoma cell line VCaP and
prostate epithelial cell RWPE-1 cell line were obtained from
American Type Culture Collection (ATCC, Manassas, VA) and cultured
accordingly. VCaP cell lines were cultured in DMEM (Invitrogen)
supplemented with 10% fetal bovine serum (Gibco), 100 U/ml
penicillin and 100 U/ml streptomycin (Invitrogen) and maintained in
a humidified atmosphere of 5% CO2, 95% air at 37°C.
RWPE-1 cells were grown in Keratinocyte serum-free medium (K-SFM)
medium (Invitrogen) supplemented with 5 ng/ml human recombinant EGF
and 0.05 mg/ml bovine pituitary extract under standard cell culture
conditions.
Experimental groups
VCaP cells were divided into the following 8 groups
for gene transfection experiment. The control group (NC); HSP72
siRNA, HSC70-siRNA, HSP72/HSC70-siRNA; UTMD+HSP72-siRNA,
UTMD+HSC70-siRNA, UTMD+HSP72/HSC70-siRNA; UTMD.
UTMD exposure protocols
A therapeutic ultrasound machine (Physioson-Basic,
Physioson Elektromedizin, Germany) was used to emit ultrasound at
the frequency of 1 MHz. The area of the ultrasound probe was 2.5
cm2. The ultrasound transducer was placed at the bottom
of plates or dishes with coupling medium on the surface of the
transducer. The adjustable sonication parameters included
ultrasound intensity, exposure time, pulse frequency, and duty
cycle.
Microbubbles (SonoVue, Bracco, Milan, Italy) were
lipid-shelled ultrasound contrast agents containing sulfur
hexafluoride gas (diameter 2.5–6.0 μm) and used at a
concentration of ∼2×108 bubbles/ml. The volumetric ratio
of microbubbles to medium dictated the choice of contrast agent
dose.
After protocol optimization with several various
settings, the following UTMD parameters were used: ultrasound
intensity, 1 W/cm2; exposure time, 60 sec; pulse
frequency, 100 Hz; duty cycle, 20%; volumetric ratio of
microbubbles: medium, 1:5.
siRNA-mediated gene knockdown
siRNAs (Shanghai GenePharma Co., Ltd.) were designed
against the open reading frame of HSP72 (HSPA1A; accession number
NM_005345) or HSC70 (HSPA8; accession number NM_006597). Two active
sequences were used for studies against HSP72 or HSC70 (designated
HSP72 or HSC70). Active sequences for HSP72 and HSC70 are as
follows: HSP72 (sense: GGACGAGUUUG AGCACAAGTT, antisense:
CUUGUGCUCAAACUCGUC CTT), HSC70 (sense: CCAAGCAGACGCAGAUCUUTT,
antisense: AAGAUCUGCGUCUGCUUGGTT), Negative control (sense:
UUCUCCGAACGUGUCACGUTT, antisense: ACGUGACACGUUCGGAGAATT). For all
experiments cells were transfected with siRNA (200 nM for single
transfections or 100 nM for combinatorial transfections) in
Opti-MEM® medium without serum according to the
Lipofectamine® 2000 protocol (Invitrogen). An equal
amount of scrambled Stealth siRNA was used as a negative control.
Six hours after the cells were transfected, the medium was replaced
with fresh culture medium. All experiments were performed 48 h
after transfection and repeated three times.
Gene transfection efficiency
Following gene transfection, the effciency of
transfection was evaluated by fuorescence microscopy and flow
cytometry. Green fuorescence was detected using inverted
fuorescence microscopy (Zeiss Axiovert S100; Carl Zeiss, Jena,
Germany). The percentage of FAM expression of the transducted VCaP
cells was quantitatively measured by flow cytometry (FACSCalibur;
BD Biosciences, Franklin Lakes, NJ, USA).
Cell viability assays
A Cell Counting Kit-8 assay (CCK-8, Dojindo
Laboratories) was used to evaluate relative cell viability. At 48 h
after siRNA transfections, RWPE-1 and VCaP cells were plated in
96-well microtiter plates at a density of 5×103 cells
per well, then the live cell count was assayed using CCK-8
according to the manufacturer’s instructions. Briefly, 10 μl
of CCK8 solution was added to each well, and the absorbance at 450
nm was measured using a Thermomax microplate reader (Molecular
Devices, Hercules, CA, USA) after 1–4 h of incubation. Relative
cell viability was calculated as a percentage of untreated control
cells.
Flow cytometry
Cells were trypsinized and resuspended in Opti-MEM
medium and plated on 6-well plates at 1,000 cells/well after
transfection with siRNA for 48 h. For apoptosis detection, cells
were stained with Annexin V and propidium iodide using the Annexin
V-FITC Apoptosis Detection kit (Invitrogen), and the percentage of
apoptotic cells was determined by flow cytometry (Beckman
Coulter).
Quantitative real-time PCR
Total RNA from cultured cells was extracted using
TRIzol reagent (Invitrogen) and reverse transcription was performed
by PrimeScript® RT Master mix Perfect real-time (Takara)
according to the manufacturer’s instructions. RNA integrity was
examined by agarose gel electrophoresis. Quantitative RT-PCR was
performed in triplicate using the SYBR-Green (Takara) method on an
Applied Biosystems Fast Sequence Detector System. Conditions for
each target were validated by standard and melting curve analyses.
We used β-actin expression as an internal control. Expression was
normalized to β-actin and determined using the Applied Biosystems
7500. All primers were designed using Primer 5 and synthesized by
Shanghai Sunny Biotechnology Co., Ltd. The following primers were
used: HSP72 (forward, 5′-TGGAGTCCTACGCCTTCAAC-3′ reverse, 5′-AGCCAC
GAGATGACCTCTTG-3′), HSC70 (forward, 5′-CGCAAGC ATAAGAAGGACATCA-3′
reverse, 5′-GAACAGGTCAGC ATTCAGTTCT-3′), β-actin (forward,
5′-GGAGATTACTGC CCTGGCTCCTA-3′ reverse, 5′-GACTCATCGTACTCCT
GCTTGCTG-3′). Caspase-3 (forward, 5′-GAACTGGACTG TGGCATTGAGA-3′
reverse, 5′-ATGGCACAAAGCGAC TGGATGA-3′). HSP90 (forward,
5′-TCTTGGCACCACCT ACTCTTGT-3′ reverse, 5′-CATCACCGATCAACCGT
TCAGT-3′).
Western blot analysis
Prostate cells transfected with siRNA were washed
twice in ice-cold phosphate buffer solution (PBS), harvested and
then lysed using RIPA buffer (Sigma). Protein concentrations were
quantified using the BCA Protein assay kit (Thermo Scientific,
Rockford, IL, USA). Equal amounts of protein and PageRuler™
Prestained molecular weight markers (Thermo Scientific) were
separated by 8% or 10% SDS-PAGE and transferred to 0.45 μm
pore size PVDF membrane (Millipore). Membranes were blocked in TBS
with Tween-20 containing 5% skim milk (Bright Dairy) and incubated
with the primary antibody at room temperature for 1 h in blocking
buffer. Antibodies and dilutions were 1:2,000 for anti-HSP72
(C92F3A-5, Stressgen Biotech Corp.), 1:1,000 for anti-caspase-8
(1006-1, Epitomics), 1:2,000 for anti-HSP90 (2877-1, Epitomics),
1:50,000 for anti-PARP-1 (1051-1, Epitomics), 1:5,000 for
anti-HSC70, anti-caspase-3 (1776-1, 1087-1, Epitomics), 1:2,000 for
anti-cleaved caspase-3, and anti-β-actin (9664, 4970, Cell
Signaling Technology) antibodies. Washing steps after hybridization
were three times at room temperature for 10 min for all other
antibodies. Membranes were incubated with secondary HRP-conjugated
antibodies diluted 1:2,000 in blocking buffer and at room
temperature for 1 h. After three washing steps of 10 min at room
temperature, detection of the signals was performed using the
Immobilon Western Chemiluminescent HRP Substrate (Millipore).
Statistical analysis
Statistical analysis was carried out with SPSS
version 12.0 (SPSS, Inc., Chicago, IL, USA). All data are presented
as mean ± SD and analyzed by One-way ANOVA. Values were considered
statistically significant at P<0.05. Results are representative
of more than three individual experiments.
Results
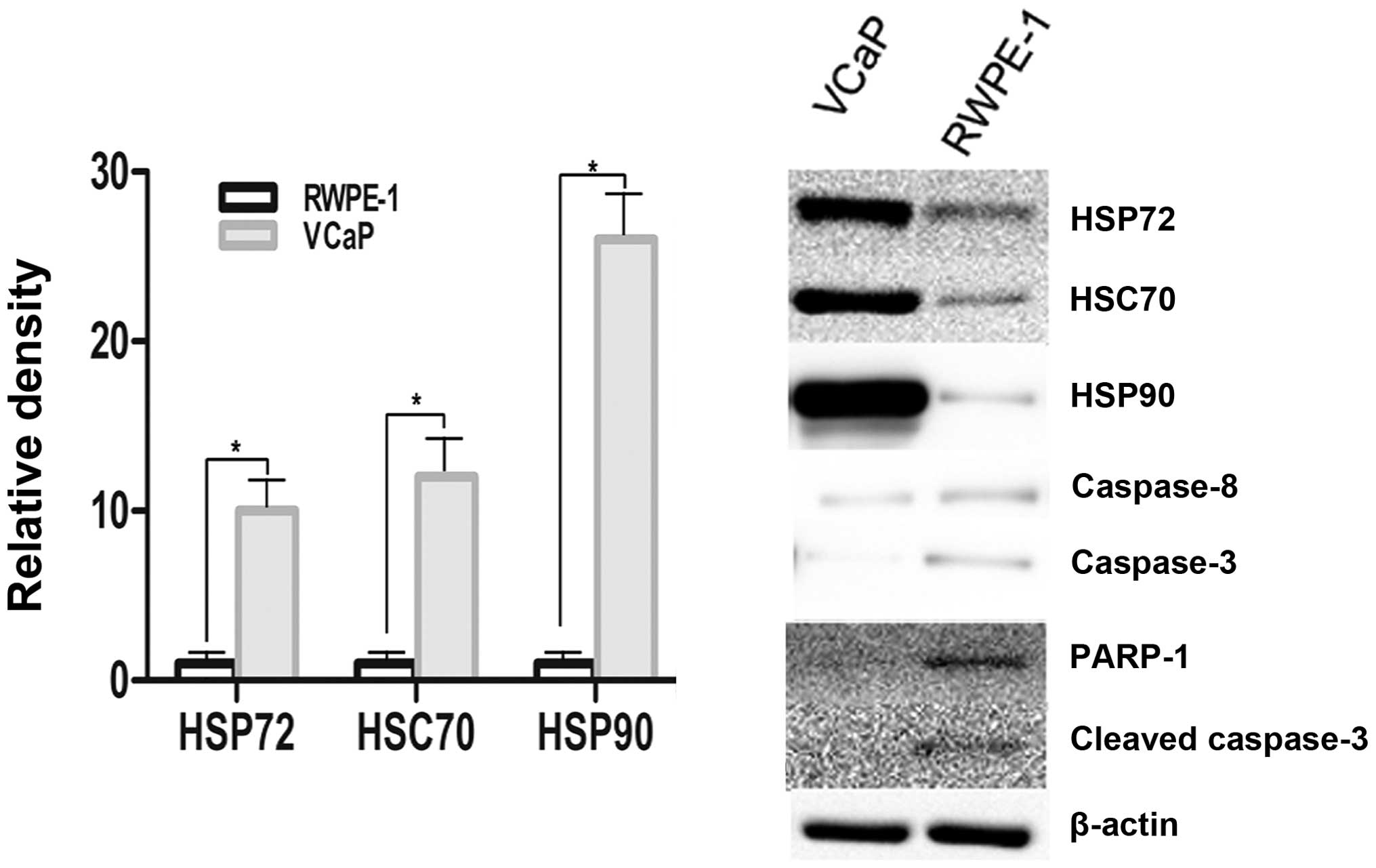
HSP72, HSC70 and HSP90 is overexpressed
in human prostate cancer cells compared with human prostate
epithelial cells
HSP70 is a critical co-chaperone for HSP90. HSP70
and HSP90 are important therapeutic biomarker of prostate cancer.
Caspase-8 is a member of a family of cysteine-requiring aspar-tate
proteases that participate in the intracellular signaling cascade
leading to apoptosis (programmed cell death) (22). Poly (ADP-ribose) polymerase (PARP)
is zinc-dependent DNA binding protein that recognizes DNA strand
breaks and is presumed to play a role in DNA repair. As a marker
for apoptosis, PARP is cleaved in vitro by many caspases
(23). We detected the cleaved
caspase-8 and p25 cleaved-form of PARP-1 (Cleaved p25). We compared
gene expression profiles of VCaP and RWPE-1 in vitro, and
found that HSP72, HSC70, and HSP90 was overexpressed in VCaP cells.
Western blot analysis revealed that VCaP cells expressed higher
levels of HSP72, HSC70 and HSP90 protein in vitro compared
with RWPE-1, which expressed undetectable levels. VCaP and RWPE-1
cells expressed very low levels of caspase-8, caspase-3, PARP-1 and
cleaved caspase-3 (Fig. 1).
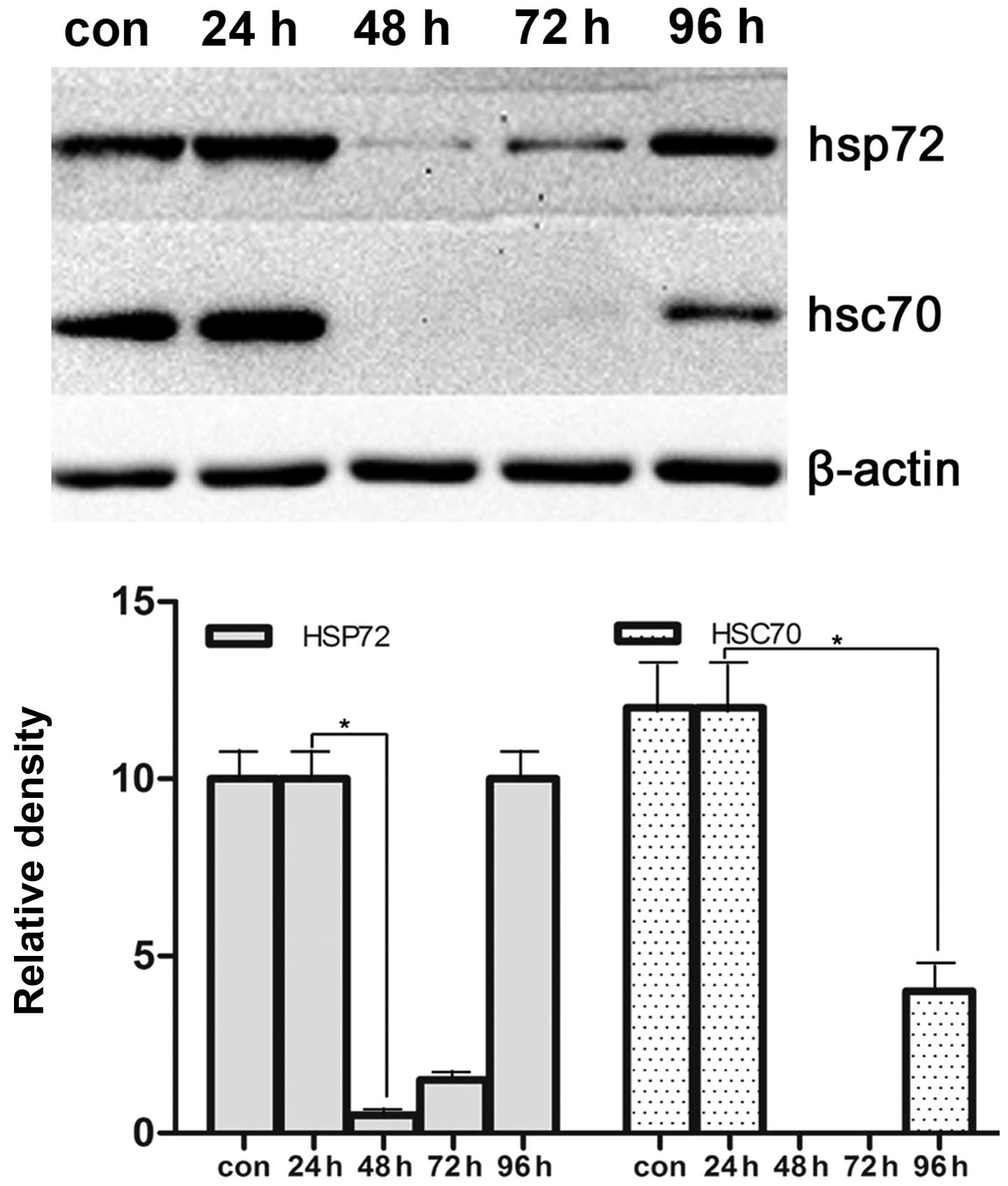
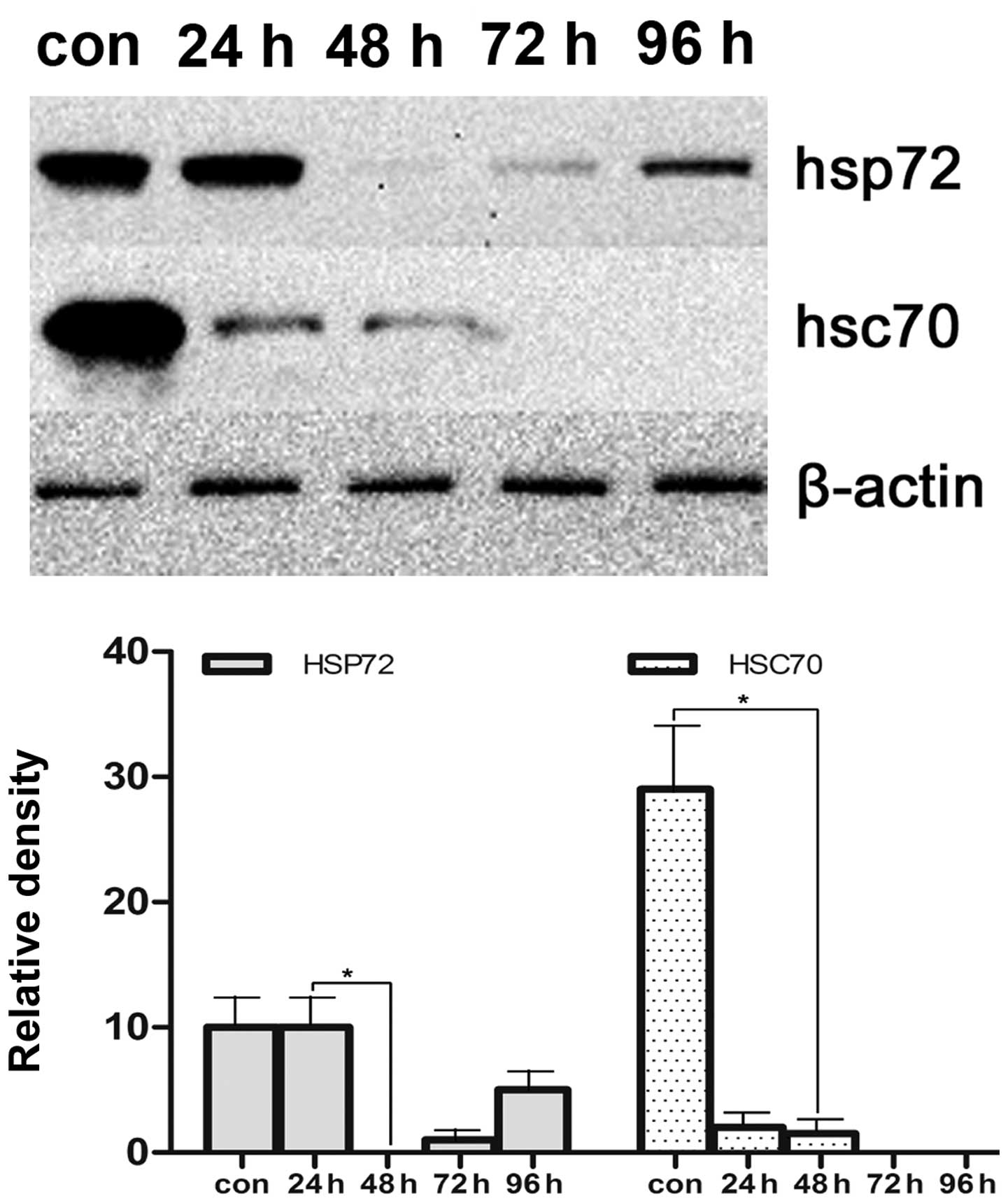
Effects of HSP70 expression after
transfection at different time points
To confirm the optimal time points in HSP70
expression in vitro after transfection, we selected 24, 48,
72, 96 h to quantify the expression of HSP70 protein in VCaP cells.
As shown in Figs. 2 and 3, optimal time points of efficacy of
silencing HSP70 gene by siRNA is 48 h. Therefore, this was used for
further investigation after transfection in vitro.
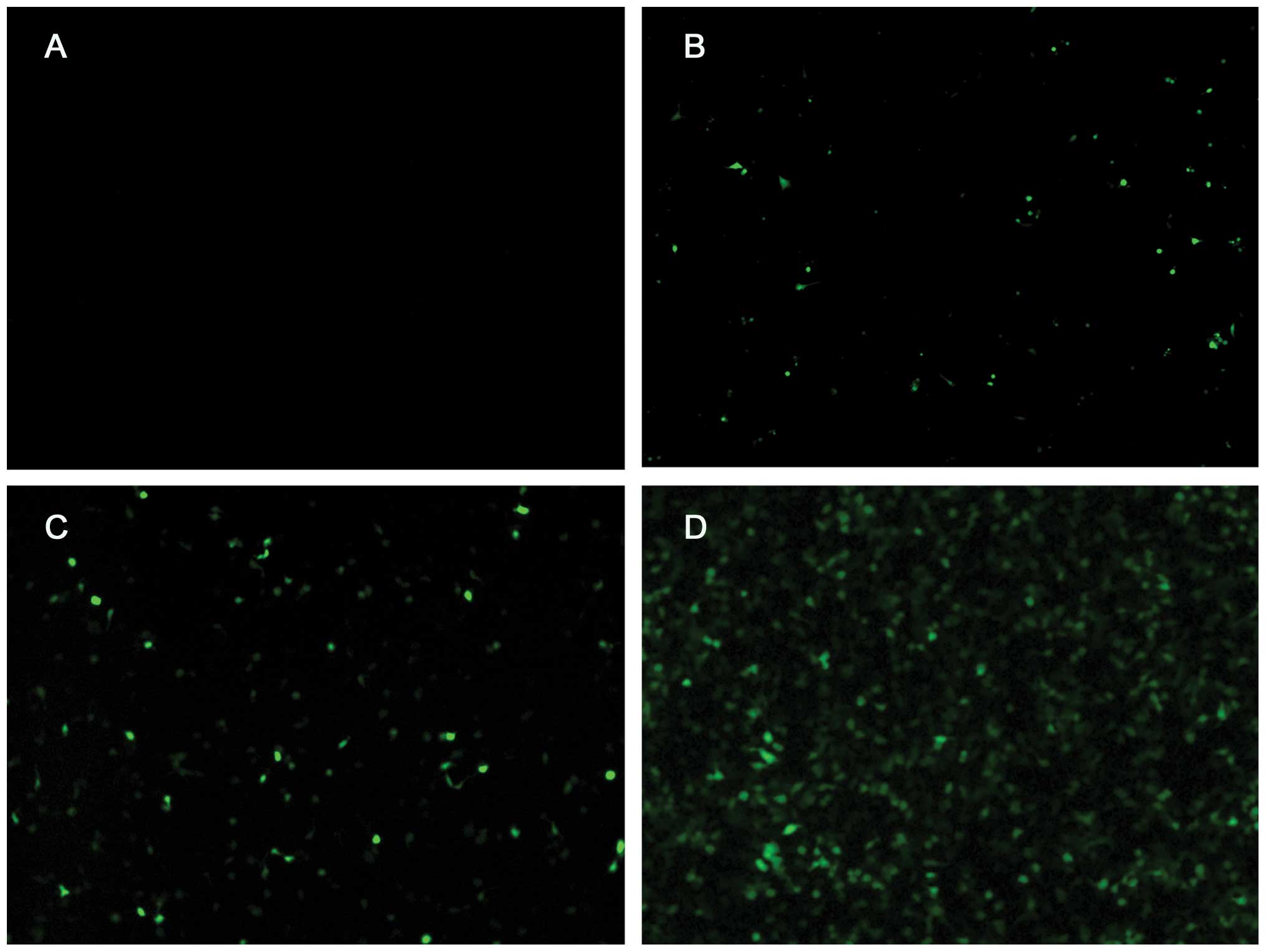
Gene transfection efficiency and
apoptosis
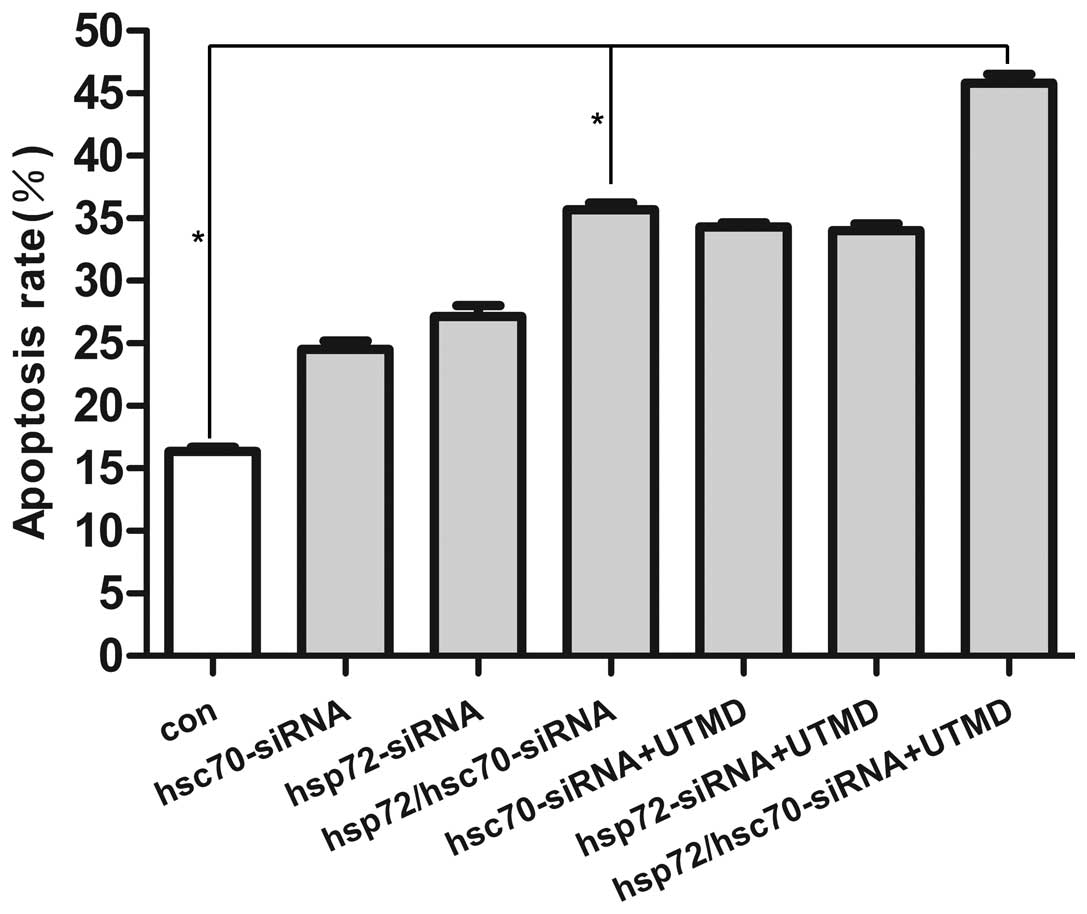
Fig. 4 shows the
UTMD+HSC70/HSP72-siRNA group had maximal transfection percentage
(73.15±0.53) and it was significantly higher than HSC70/HSP72-siRNA
group (40.37±0.65), UTMD+HSC70-siRNA group (28.31±0.60) and
UTMD+HSP72-siRNA group (24.25±0.53). In accordance with the results
of transfection analysis, the apoptosis percentage of the
UTMD+HSC70/HSP72-siRNA group was the highest (47.16%) and the other
6 groups had significant difference between each other (P<0.05).
It seems that the UTMD+HSC70/HSP72-siRNA could serve as a gene
delivery system, and silencing HSP70 and HSP90 expression induced
extensive cell apoptosis (Fig.
5).
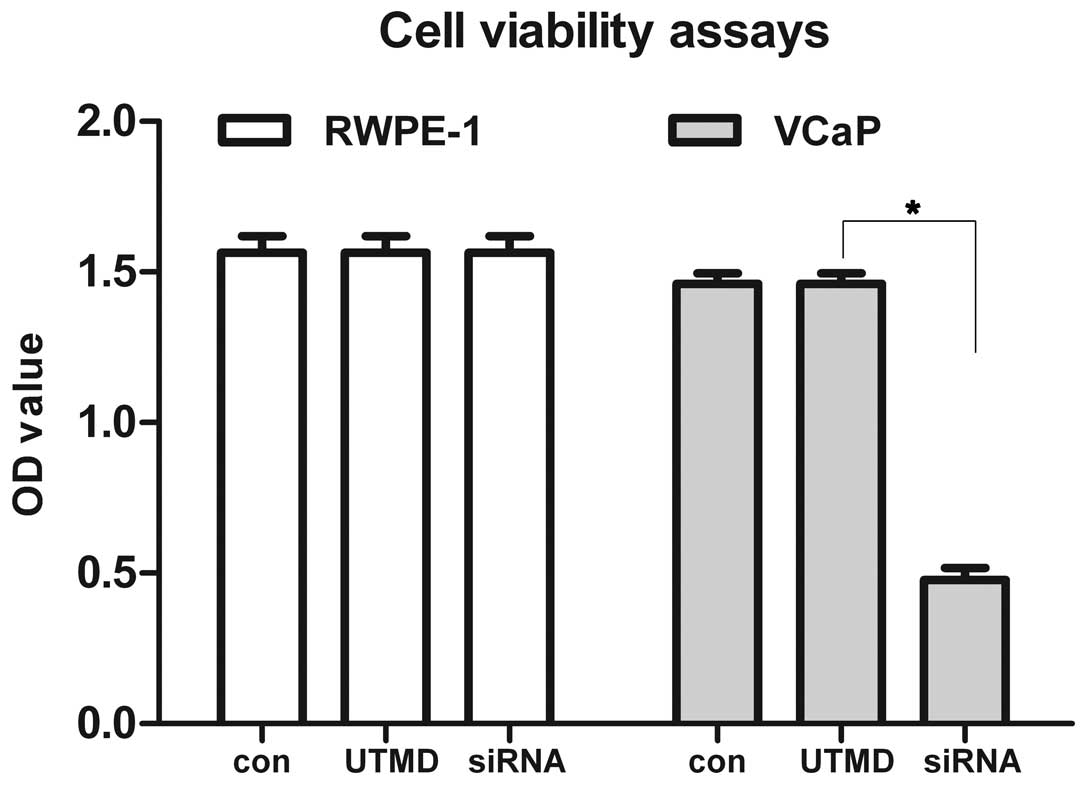
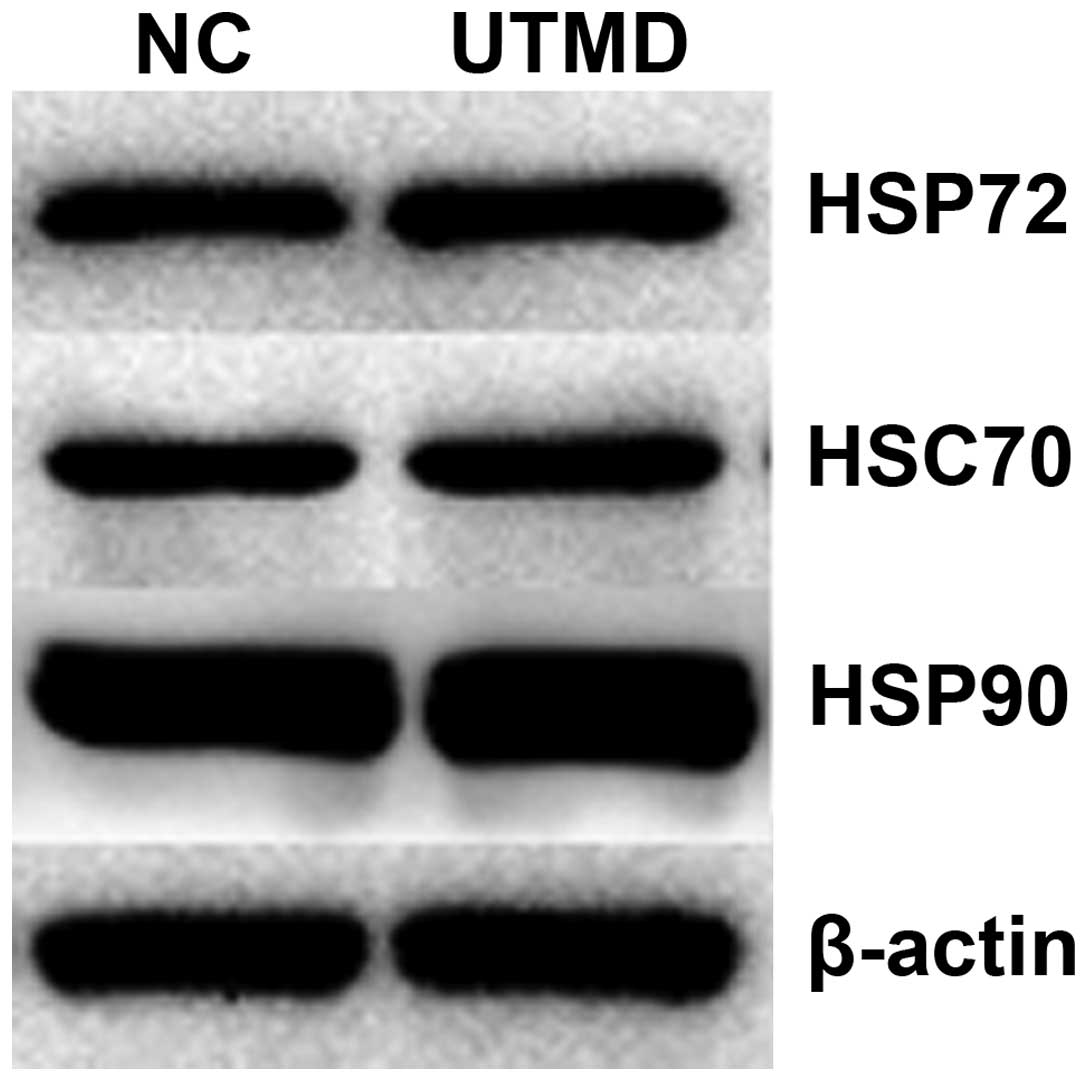
UTMD is a safe and noninvasive
approach
In order to investigate the safety of UTMD, cellular
viability was monitored using the Cell Counting Kit-8 assay for 48
h. The cellular viability of VCaP cells transfected with HSP72 and
HSC70 siRNA was delayed compared with that of the control groups
after UTMD. No significant difference was found in the cellular
viability between the RWPE-1 and VCaP of the control group and the
two group treated with UTMD (Fig.
6).
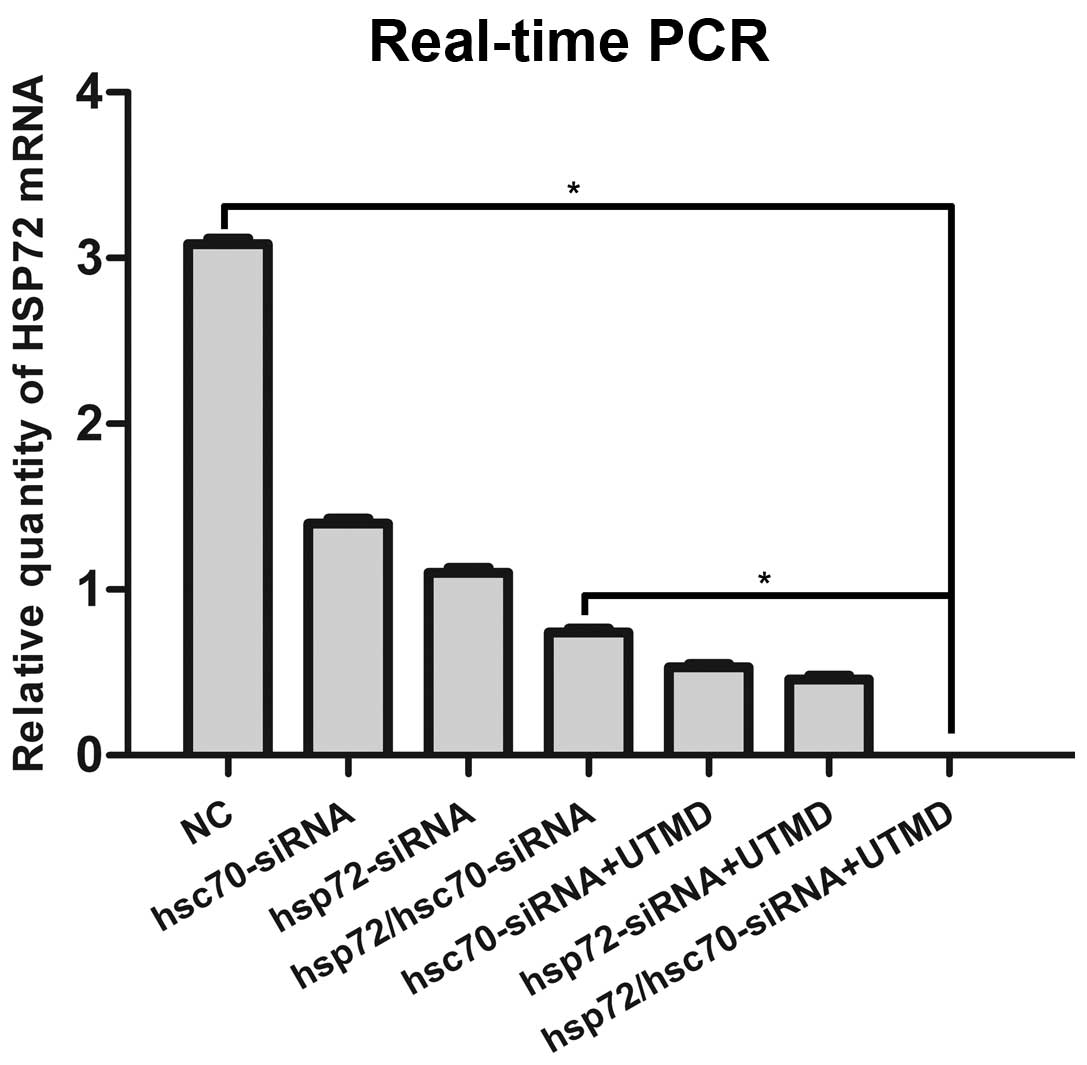
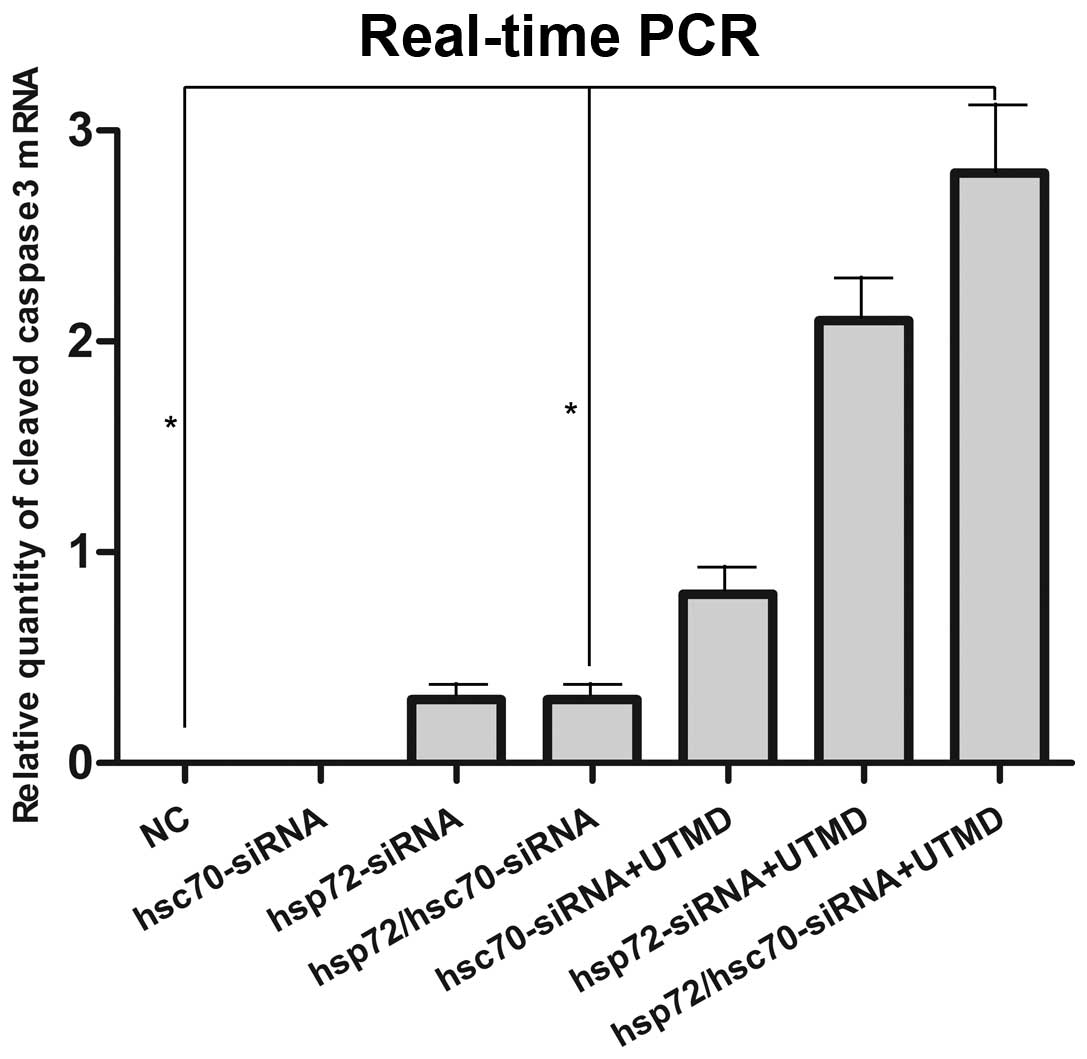
To investigate the molecular mechanisms by which the
combination of UTMD and HSP70-siRNA restrains PCa in the context of
HRPC, we processed a set of relevant experiment in vitro.
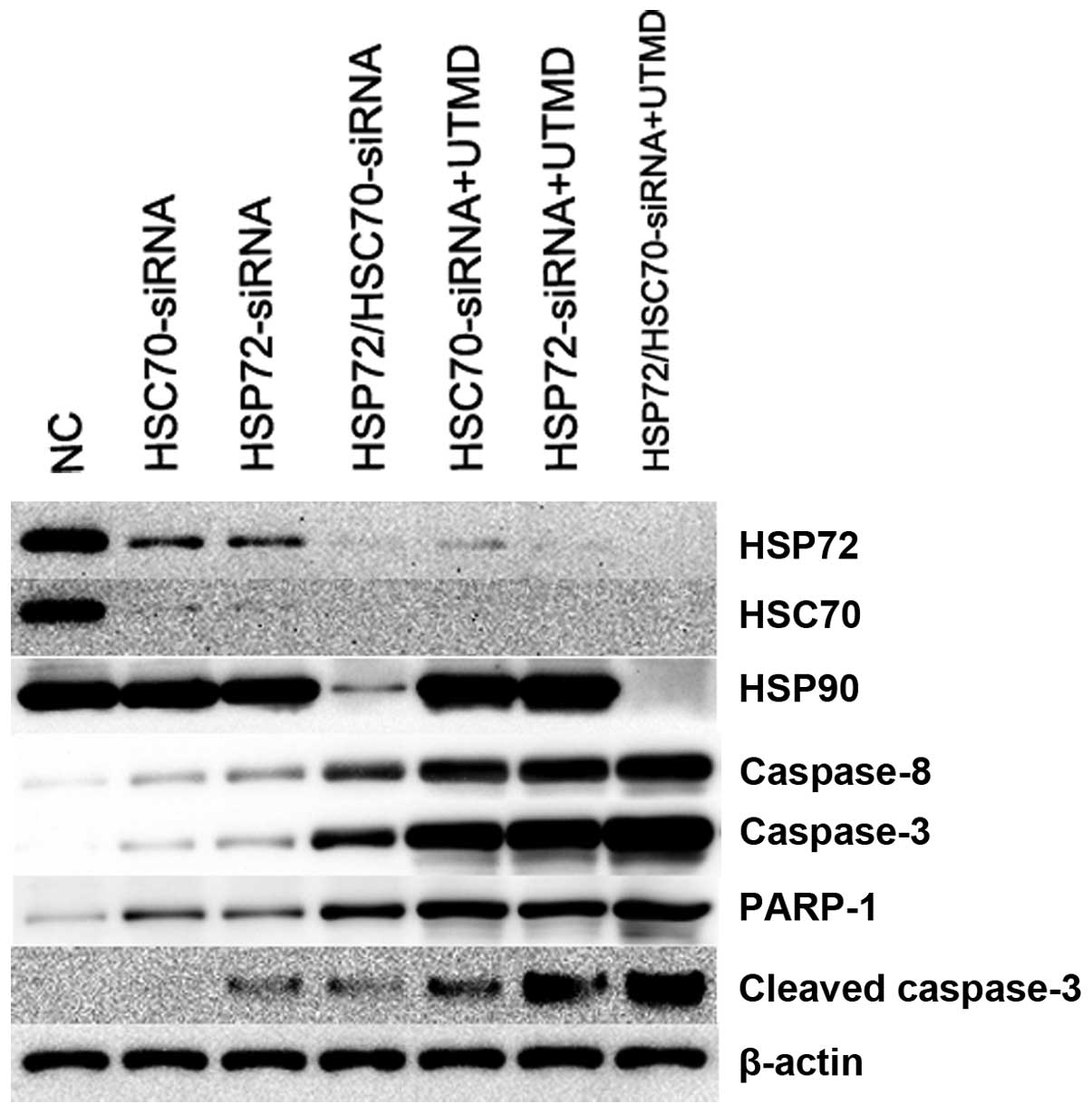
RT-qPCR revealed 3.1-fold higher levels of HSP72, HSC70 and HSP90
mRNA in NC control compared with HSP72/HSC70-siRNA treatment (Figs.
7-Figs. 10). The group of UTMD combined
with HSP72/HSC70-siRNA expressed undetectable levels of HSP72,
HSC70 and HSP90 mRNA. This is consistent with the results of
western blotting in VCaP cells (Figs.
11 and 12). To better
understand the biological processes that underlie what appears to
be HSP70, HSP90, and cleaved caspase-3 in VCaP cells, we next
sought to define the mechanism by which HSP70 regulates cleaved
caspase-3 expression. We found that the caspase-3, caspase-8,
PARP-1, and cleaved caspase-3 gene was the most significantly
enriched in the group of UTMD combined HSP72/HSC70-siRNA (Figs. 10 and 11). Therefore, we tested the ability of
the combination of UTMD with HSP72/HSC70-siRNA to induce tumor
specific apoptosis.
Discussion
Silencing of gene expression by siRNA is rapidly
becoming a powerful tool against cancer, efficient delivery of
siRNA into tumor cells remains a key obstacle (24). Although local surgery, radiation,
or hormonal ablation provide initial response at earlystages of the
disease, tumor cells often develop resistance and relapse. Thus,
the identification of new therapeutic targets for prostate cancer
is of critical importance. Recent studies further revealed that
≈2/3 of human prostate cancers expressed high level of HSP72 and
HSC70, associating with recurrence, development of
castration-resistance, and poor survival (25,26).
These studies provide the first hints that HSP72 and HSC70 may have
a critical role in prostate cancer development and therapeutic
resistance. HSP90 is an exciting therapeutic target in cancer
because inhibition of this single protein causes the simultaneous
degradation of multiple oncoproteins and combinatorial blockade of
numerous oncogenic pathways. HSP70 molecular chaperones are of
interest when considering modulation of HSP90 (27). HSP70 is the major therapeutic
target in advanced PCa (28–32).
However, targeting HSP70 or androgen receptor alone can result in
off-target effects, drug resistance and disease recurrence
(30,33,34).
Therefore, simultaneous targeting of a multi-modal approach or
multiple pathways in targeted delivery could in principle be an
effective approach to treating prostate cancer. With respect to the
functional role of HSP70 isoforms in the HSP90 chaperone, the data
we presented here demonstrate that silencing HSC70 or HSP72
individually has little effect on HSC70 and HSP72, has no effect on
HSP90 protein level of VCaP prostate cancer cells, while dual
targeting of HSC70 and HSP72 inhibit the cellular chaperone
activity of HSP90, and has good silencing effect on HSC70 and
HSP72, thus induce tumor-specifc apoptosis. Additionally, UTMD
combined with simultaneous targeting of HSC70 and HSP72 may further
enhance silencing efficacy of the expression of HSP72, HSC70, HSP90
protein and mRNA. In agreement with our findings, Powers et
al (34) also found that dual
targeting of HSC70 and HSP72 silence HSP90 protein and induce
extensive tumor-specifc apoptosis in human HCT116 colon and A2780
ovarian cells.
In the present study, we have shown by flow
cytometry that silencing HSC70 or HSP72 individually has no effect
on apoptosis. Using combinatorial siRNA approach, we revealed that
dual silencing of HSC70 and HSP72 considerably increased the
apoptotic efficacy by 20% compare with control group, UTMD may
further enhance extensive tumor-specific apoptosis by 11% (Fig. 5). Our apoptosis results were in
accord with the study of Rérole et al (35). In order to confirm further the
results of flow cytometry, we selected several major apoptotic
markers. Caspase-3, caspase-8, PARP-1 and cleaved caspase-3 are the
main executor of apoptosis and considered to be key hallmarks of
apoptosis (36,37). Our data on apoptosis is in near
agreement with the expression of major apoptotic markers
(caspase-8, caspase-3, PARP-1, cleaved caspase-3) of western
blotting. Of note, the safety and noninvasiveness of the
combinational targeting strengthen our confidence further. UTMD is
a noninvasive physical gene delivery technology by sonoporation.
There is no significant difference of cell viability in the UTMD
group and normal control group, indicating that UTMD cells without
any apparent toxicity, but also UTMD can promote effective uptake
of siRNA, and enhance the specificity of siRNA to silence HSP70 and
HSP90. Therefore, we may promote the efficacy and specificity of
siRNA delivered by UTMD. UTMD may improve the accuracy of target
genes. Moreover, these data indicate that this is a promising
strategy for inducing tumor-specific apoptosis in vitro. The
safety of the combinational targeting in vivo need to be
explored in subsequent experiments.
The most important attribute of UTMD combination
with siRNA is their specific, efficient delivery, which leads to
concentration in the target tumor and avoidance of distribution to
irrelevant normal tissues. Specific delivery into the intended
target cells will likely reduce the dose required for antitumor
activity as well as limit toxicity.
An exciting implication of our research is the
attractiveness of combinatorial targeting of HSC70 and HSP72 as an
alternative means to achieve HSP90 inhibition, with the added
advantage of avoiding the anti-apoptotic effects of HSP70 isoform
induction that limit the use of current pharmacologic inhibitors.
Noteworthy, UTMD may promote the effect of silencing HSP90. The
approach of UTMD combined siRNA was used here as a tool to silence
the expression of HSC70, HSP72 and HSP90. It is possible that UTMD
combined with RNA interference may be developed as a potential
therapeutic approach.
Our findings also revealed further details about the
relationship between UTMD and siRNA, especially in the context of
HRPC. The reasons why UTMD combined with dual targeting of HSC70
and HSP72 improve the efficacy of silencing are: Firstly, UTMD
promote the efficient delivery and cell uptake of siRNA. Secondly,
UTMD also may activate the active region of HSP72, HSC70, HSP90 or
increase crosstalk between distinct signaling pathways. Thirdly,
UTMD combined with dual targeting of siRNA might active multiple
signal transduction pathways of apoptosis and heat shock protein.
Fourthly, UTMD improve the permeability and ATP energy of
mitochondria. Further work is required to identify the molecular
and cellular mechanisms whereby UTMD promote dual targeting of
siRNA induce extensive tumor-specifc apoptosis in vitro and
in vivo from the results presented here.
In conclusion, UTMD-mediated delivery of dual
targeting of siRNA is a promising strategy for treatment of
prostate cancer in vitro that could prove more effective and
specific than other siRNA delivery strategies. The other main
finding of this study is the potential superiority for HSP70 and
HSP90 as biomarker targeting in PCa (33,38).
Our data establish that UTMD-directed simultaneous targeting of
siRNA therapeutics is minimally invasive, and effective at
suppressing tumor growth, and induce extensive tumor-specifc
apoptosis, and these data might provide insights into further
optimization and the multi-model delivery strategy is versatile in
specific gene silencing of targeted therapeutics of cancer.
Abbreviations:
|
UTMD
|
ultrasound-targeted microbubble
destruction;
|
|
siRNA
|
small interfering RNA;
|
|
HRPC
|
hormone refractory prostate
cancer;
|
|
RT-PCR
|
reverse transcriptase-polymerase chain
reaction
|
Acknowledgements
We thank members of the Guo Hui Fu
Laboratory for helpful discussions. We thank Dr J.J. Yu for
technical assistance. This study was supported by the National
Natural Science Foundation of China; grant no. 81171352 and
81271596 (to L. Du), no. 81201097 (to M. Bai), and the Natural
Science Foundation of Shanghai no. 12ZR1424800 (to M. Bai) is
gratefully acknowledged.
References
|
1.
|
Siegel R, Ma J, Zou Z, et al: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar
|
|
2.
|
DeSantis C, Naishadham D and Jemal A:
Cancer statistics for African Americans, 2013. CA Cancer J Clin.
63:151–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011. View Article : Google Scholar
|
|
4.
|
Bitting RL and Armstrong AJ: Targeting the
PI3K/Akt/mTOR pathway in castration-resistant prostate cancer.
Endocr Relat Cancer. 20:R83–R99. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Garcia JA and Rini BI:
Castration-resistant prostate cancer: Many treatments, many
options, many challenges ahead. Cancer. 118:2583–2593. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
McNamara JO II, Andrechek ER, Wang Y, et
al: Cell type-specific delivery of siRNAs with aptamer-siRNA
chimeras. Nat Biotechnol. 24:1005–1015. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Daugaard M, Rohde M and Jäättelä M: The
heat shock protein 70 family: highly homologous proteins with
overlapping and distinct functions. FEBS Lett. 581:3702–3710. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Zuiderweg ER, Bertelsen EB, Rousaki A, et
al: Allostery in the Hsp70 chaperone proteins. Top Curr Chem.
328:99–153. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Davenport EL, Zeisig A, Aronson LI, et al:
Targeting heat shock protein 72 enhances Hsp90 inhibitor-induced
apoptosis in myeloma. Leukemia. 24:1804–1807. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Powers MV, Jones K, Barillari C, et al:
Targeting HSP70: the second potentially druggable heat shock
protein and molecular chaperone? Cell Cycle. 9:1542–1550. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Meng L, Hunt C, Yaglom JA, et al: Heat
shock protein Hsp72 plays role in Her2-induced mammary
tumorigenesis. Oncogene. 30:2836–2845. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Stangl S, Gehrmann M, Riegger J, et al:
Targeting membrane heat-shock protein 70 (Hsp70) on tumors by
cmHsp70.1 antibody. Proc Natl Acad Sci USA. 108:733–738. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Evans CG, Chang L and Gestwicki JE: Heat
shock protein 70 (hsp70) as an emerging drug target. J Med Chem.
53:4585–4602. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Balaburski GM, Leu JI, Beeharry N, et al:
A modified HSP70 inhibitor shows broad activity as an anticancer
agent. Mol Cancer Res. 11:219–229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Rérole AL, Gobbo J, De Thonel A, et al:
Peptides and aptamers targeting HSP70: a novel approach for
anticancer chemotherapy. Cancer Res. 71:484–495. 2011.
|
|
16.
|
Yao YD, Sun TM, Huang SY, et al: Targeted
delivery of PLK1-siRNA by ScFv suppresses HER2+ breast
cancer growth and metastasis. Sci Transl Med.
4:130ra482012.PubMed/NCBI
|
|
17.
|
Li HL, Zheng XZ, Wang HP, et al:
Ultrasound-targeted micro-bubble destruction enhances AAV-mediated
gene transfection in human RPE cells in vitro and rat retina in
vivo. Gene Ther. 16:1146–1153. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Xie W, Liu S, Su H, et al: Ultrasound
microbubbles enhance recombinant adeno-associated virus vector
delivery to retinal ganglion cells in vivo. Acad Radiol.
17:1242–1248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Zheng X, Du L, Wang H and Gu Q: A novel
approach to attenuate proliferative vitreoretinopathy using
ultrasound-targeted microbubble destruction and recombinant
adeno-associated virus-mediated RNA interference targeting
transforming growth factor-β2 and platelet-derived growth factor-B.
J Gene Med. 14:339–347. 2012.PubMed/NCBI
|
|
20.
|
Suzuki R, Oda Y, Utoguchi N and Maruyama
K: Progress in the development of ultrasound-mediated gene delivery
systems utilizing nano- and microbubbles. J Control Release.
149:36–41. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Tachibana K, Uchida T, Ogawa K, et al:
Induction of cell-membrane porosity by ultrasound. Lancet.
353:14091999. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Boldin MP, Goncharov TM, Goltsev YV and
Wallach D: Involvement of MACH, a novel MORT1/FADD-interacting
protease, in Fas/APO-1- and TNF receptor-induced cell death. Cell.
85:803–815. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Lazebnik YA, Kaufmann SH, Desnoyers S, et
al: Cleavage of poly(ADP-ribose) polymerase by a proteinase with
properties like ICE. Nature. 371:346–347. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Pai SI, Lin YY, Macaes B, et al: Prospects
of RNA interference therapy for cancer. Gene Ther. 13:464–477.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Daneshmand S, Quek ML, Lin E, et al:
Glucose-regulated protein GRP78 is up-regulated in prostate cancer
and correlates with recurrence and survival. Hum Pathol.
38:1547–1552. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Pootrakul L, Datar RH, Shi SR, et al:
Expression of stress response protein Grp78 is associated with the
development of castration-resistant prostate cancer. Clin Cancer
Res. 12:5987–5993. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Whitesell L and Lindquist SL: HSP90 and
the chaperoning of cancer. Nat Rev Cancer. 5:761–772. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Lebret T, Watson RW and Fitzpatrick JM:
Heat shock proteins: their role in urological tumors. J Urol.
169:338–346. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Garrido C, Schmitt E, Candé C, et al:
HSP27 and HSP70: potentially oncogenic apoptosis inhibitors. Cell
Cycle. 2:579–584. 2003. View Article : Google Scholar
|
|
30.
|
McConnell JR and McAlpine SR: Heat shock
proteins 27, 40, and 70 as combinational and dual therapeutic
cancer targets. Bioorg Med Chem Lett. 23:1923–1928. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Alaiya AA, Oppermann M, Langridge J, et
al: Identification of proteins in human prostate tumor material by
two-dimensional gel electrophoresis and mass spectrometry. Cell Mol
Life Sci. 58:307–311. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Mayer MP: Hsp70 chaperone dynamics and
molecular mechanism. Trends Biochem Sci. 38:507–514. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Lee E, Madar A, David G, et al: Inhibition
of androgen receptor and β-catenin activity in prostate cancer.
Proc Natl Acad Sci USA. 110:15710–15715. 2013.
|
|
34.
|
Powers MV, Clarke PA and Workman P: Dual
targeting of HSC70 and HSP72 inhibits HSP90 function and induces
tumor-specific apoptosis. Cancer Cell. 14:250–262. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Rérole AL, Jego G and Garrido C: Hsp70:
anti-apoptotic and tumorigenic protein. Methods Mol Biol.
787:205–230. 2011.
|
|
36.
|
Earnshaw WC, Martins LM and Kaufmann SH:
Mammalian caspases: structure, activation, substrates, and
functions during apoptosis. Annu Rev Biochem. 68:383–424. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Tang NY, Huang YT, Yu CS, et al: Phenethyl
isothiocyanate (PEITC) promotes G2/M phase arrest via p53
expression and induces apoptosis through caspase- and
mitochondria-dependent signaling pathways in human prostate cancer
DU 145 cells. Anticancer Res. 31:1691–1702. 2011.
|
|
38.
|
Goloudina AR, Demidov ON and Garrido C:
Inhibition of HSP70: a challenging anti-cancer strategy. Cancer
Lett. 325:117–124. 2012. View Article : Google Scholar : PubMed/NCBI
|