Introduction
Esophageal squamous cell carcinoma (ESCC) is one of
the most common malignant tumors worldwide and seriously threatens
human health (1). As current
methods for ESCC screening are invasive and can hardly detect ESCC
in its early stages, clinically significant and minimally invasive
tests for ESCC are necessary. Recently, the potential applications
of circulating tumor cell-associated microRNAs have been
investigated, and these miRNAs have been suggested as novel
biomarkers in plasma or serum for cancer diagnosis (2–6). A
series of miRNA expression profile studies in plasma from cancer
patients has been conducted to explore the tumor-specific miRNAs. A
research on pancreatic cancer (PC) found 37 miRNAs downregulated
and 54 miRNAs upregulated in plasma from patients with PC.
Furthermore, miR-21 was correlated with worse PC survival, whereas
let-7 was inversely correlated with survival (7). With the current development of
next-generation deep sequencing, Illumina platform-based sequencing
was performed to establish a plasma miRNA profile in nasopharyngeal
carcinoma (NPC) patients; 13 miRNAs were found to be NPC specific
(8). miRNAs may be widely used as
a clinical biomarker as miRNAs in plasma were demonstrated to be
resistant to ribonuclease digestion and temperature changes
(9,10).
Exosomes, which are small vesicles of endocytic
origin, were reported to be released with miRNAs wrapped from both
tumor and healthy cells to participate in cell-cell communication
(11–13). Rabinowits et al (14) evaluated the expression level of 12
miRNAs from lung adenocarcinoma tissues and circulating exosomes
vs. the control group; their study found that the tumor-derived
miRNA patterns and the circulating exosomal miRNAs were similar.
Taylor et al (15) detected
a greater number of circulating epithelial cell adhesion molecule
(EpCAM)-positive exosomes in ovarian cancer patients than benign
ovarian disease or controls. They also discovered that there
existed a similar level of miRNA profile between ovarian cancer
cells and exosomes, which suggest that tumor-derived exosomal miRNA
profiling could be used as a diagnosis approach. Therefore,
circulating miRNA profiles of exosomes may be applied in tumor
early diagnosis, prognosis and recurrence with a great value.
Current research has indicated that the transfer of
exosome-derived miRNAs to recipient cells is a novel mechanism in
addition to classical mechanisms, which include direct cell-cell
contact or chemical receptor-mediated events (16). The number of studies that deal with
identification of exosomal miRNA function is increasing in recent
years. Tumor cell-derived exosome-formed miRNAs have been shown to
be transferred into recipient cells and performed their biological
functions. Nouraee et al (17) demonstrated that miR-21 could be
detected in conditioned media of esophageal cancer cells. Moreover,
a significant miR-21 upregulation in esophageal cancer cells was
observed when these cells were co-cultured with fibroblasts; this
process provided the possibility for transfer of exosome-shuttling
miRNAs. Some researchers have found that co-culture with exosomes
derived from miR-10b vector stable transfected-breast cancer cells
could induce breast epithelial cell invasiveness; thus,
exosome-shuttling miRNAs may play an important role in tumor
microenvironment modulation (18).
A study on co-cultivation of leukemia cells and endothelial cells
revealed that exogenous miR-92a could enhance recipient endothelial
cell migration and tube formation, such as endogenous miR-92a, via
exosomal transport by targeting its target gene, namely, integrin
5α (19). An increase in evidence
raises the possibility that genetic information can be transferred
via exosomes between cells. Thus, we hypothesized that
exosome-shuttling miRNAs can be transferred between esophageal
cells. Furthermore, certain miRNAs may contribute to the change of
biological properties by affecting their target genes in recipient
cells.
In our previous studies, the expression profiling of
exosomal miRNAs was derived from esophageal cancer cells through
high-throughput sequencing (20);
basing on the said study, we have revealed that miR-21 was abundant
both in esophageal cancer cells and their corresponding exosomes.
However, no studies have reported on the functional research of
exosome-shuttling miR-21 in esophageal cancer. In the present
study, we explored the exosome-shuttling miR-21 involved in
esophageal cancer progression by analyzing its impact on esophageal
cancer migration and invasion. In addition, we investigated the
regulatory mechanism behind this gene, including the target gene
and downstream molecules. Case-control study was also conducted to
identify the association of exosome-shuttling miR-21 and esophageal
cancer risk.
Materials and methods
Specimen collection and ethics
statement
Seventy patients, aged 47 to 82 years, were
recruited from the First People's Hospital of Huaian, with their
consent and agreement. All patients were confirmed by pathology or
endoscopy as ESCC without preoperative radiotherapy or
chemotherapy. Plasma samples were also collected from 70 healthy
volunteers, aged between 46–84 years, with matching age and gender
to the patients. Among these patients, blood plasma samples from
three ESCC female patients, aged 58, 73 and 77, and three female
healthy volunteers, aged 60, 76 and 80, were selected for miRNA
microarray analysis. Written consents were obtained from all
subjects prior to recruitment. The study protocol was approved by
the Institutional Review board (IRB) of Southeast University
Affiliated Zhongda Hospital in Nanjing, China. The design of the
esophageal cancer study, including plasma sample collection, was
approved by the IRB.
miRNA microarray
Three pairs of plasma samples from ESCC patients and
healthy controls were collected for micro-array analysis. Agilent
Human miRNA microarray (v19.0; Agilent Technologies Inc., Santa
Clara, CA, USA) was used in the analysis. miRNA samples from six
esophageal specimens were labeled and hybridized with miRNA
Complete Labeling and Hybridization kit (Agilent Technologies)
according to the manufacturer's protocol. Signals were normalized
using Gene Spring software 11.0 (Agilent Technologies). ANOVA was
used to compare the different miRNA expressions.
Cell culture
The human esophageal cancer cell line EC9706
(National Laboratory of Molecular Oncology, Cancer Institute,
Chinese Academy of Medical Sciences and Peking Union Medical
College, Beijing, China) (21) was
maintained in RPMI-1640 medium that contains 10% fetal bovine serum
(FBS), 100 U/ml penicillin and 100 μg/ml streptomycin. FBS was
centrifuged 10,000 × g for 30 min, followed by ultracentrifugation
at 200,000 × g for 6 h to eliminate bovine-derived exosomes using a
Type 70 Ti rotor in L-80XP ultracentrifuge (Beckman Coulter, Brea,
CA, USA) (22). Cell cultures were
performed at 37°C in a 5% CO2, water-saturated
atmosphere.
Exosome purification
Culture medium from EC9706 (1×108 cells)
was collected after a 48-h culture; this medium was subjected to
differential centrifugation. Briefly, the medium was first
centrifuged at 300 × g for 10 min, 800 × g for 10 min, 1,200 × g
for 20 min, and 10,000 × g for 30 min to remove any live or dead
cells and cellular debris. Afterwards, the supernatant was
ultracentrifuged at 100,000 × g for 3 h to pellet exosomes. The
supernatant was disposed, and the exosome pellet was washed with
phosphate-buffered saline (PBS) at 100,000 × g for 2 h. All steps
were performed at 4°C. The pellet was finally re-suspended in the
appropriate buffer for future studies. The exosome levels were
determined by measuring the total protein content, which was
presented as micrograms of total protein in the exosomes. The
exosome fraction was measured for its protein content using a
Pierce BCA protein assay kit (Thermo Fisher Scientific, Wilmington,
DE, USA).
Exosome labeling and live-cell
fluorescence microscopy
The exosome pellet suspension was diluted to a 40
μg/ml concentration with RPMI-1640 medium by adding 5 μg/ml DiI
(Biotium, Hayward, CA, USA), which is a fluorescent dye that labels
the plasma membrane. This suspension was then incubated at 37°C for
30 min and subsequently passed through a 0.22-μM filter to remove
bacteria. The excessive dye was removed using a 300-kDa
ultrafiltration tube (Pall Corp., Washington, NY, USA) five times
at 4000 × g for 10 min centrifugation. For Live Cell Imaging System
analysis, 2×105 EC9706 and 1.5 ml complete medium
(RPMI-1640 with 10% FBS) were placed in a 35-mm glass-bottom
culture dish (Nest Biotechnology Co., Ltd., Wuxi, China). After a
24-h culture, the DiI-labeled exosomes were incubated with cells
for 2.5 h and washed once to eliminate the free exosomes. Cells
were then transferred to the cell culture chamber of the
microscope, which could provide temperature and CO2
concentration that can sustain live cells. Fluorescence emission
was collected by ×60 oil-immersion objective and passed through EM
565-nm emission filters. Fluorescence images could be recorded at
0.6 ftp speed using the UltraVIEWVoX Live Cell Imaging System
(Perkin-Elmer, Waltham, MA, USA).
Co-cultivation experiment
Co-cultivation of EC9706 cells was performed in
12-well Transwell inserts (cat. no. 3401; Corning Inc., Corning,
NY, USA). EC9706 cells that were treated as recipient cells were
pre-seeded in the lower chambers at a 1×105 cells/well
density. The following day, the EC9706 cells transfected with
miR-21 mimics or the negative controls that were treated as donor
cells were scraped off and seeded onto 0.4 μM Transwell inserts,
which were filtered to supernatants but not to the cellular
components (23).
Inhibition of exosome release
To validate whether miR-21 secretion depends on
exosome transfer, exosome release was blocked using GW4869
(sc-218578; Santa Cruz Biotechnology, Santa Cruz, CA, USA), which
is a specific chemical inhibitor for nSMase2. EC9706 cells were
pre-seeded in a 24-well plate and cultured for 12 h in complete
medium. After the incubation, the medium was switched to fresh
complete medium with different GW4869 concentrations. The cells
were collected, and exosome fraction was obtained from the culture
medium after 48-h incubation.
Transfer efficiency assay for Cy3-labeled
miR-21 mimics
EC9706 cells (3.5×105/well) treated as
donor cells were transfected with 30 nM of Cy3-labeled miR-21
mimics using Lipofectamine RNAiMAX (Invitrogen). One day after 12-h
transfection, the culture medium was disposed, and cells were
washed thrice with PBS to remove the residual transfection reagent.
Subsequently, the medium was changed to fresh complete medium.
Culture medium from donor cells was collected and centrifuged to
remove residual cells after a 24-h culture. These cells were then
added into pre-seeded recipient EC9706 cells
(2×105/well) in a 6-well plate. The recipient cells were
acquired after 3, 6 and 24 h of culture. Transfer efficiency was
calculated as fluorescent cell percentage by flow cytometry. The
miR-21 expression level in recipient cells was detected by
real-time quantitative polymerase chain reaction (RT-qPCR).
RNA extraction and quantitative reverse
transcription PCR
Total RNA was extracted from cells and exosomes
using TRIzol reagent (Invitrogen) and mirVana miRNA isolation kit
(Ambion, Austin, TX, USA) according to the manufacturer's
instructions, respectively. RNA concentration was analyzed using
NanoDrop spectrophotometer (NanoDrop ND-1000; NanoDrop
Technologies, Inc., Wilmington, DE, USA). RT-qPCR was performed
using SYBR-Green Master Mix Plus (Toyobo, Osaka, Japan). Real-time
PCR was performed in 96-well plates using Primer Express software
version 3.0 (Applied Biosystems, Foster City, CA, USA). RNU6 and
cel-miR-39 were used as the invariant control for cells and
supernatant, respectively, in miR-21 expression level analysis.
β-actin was used as the invariant control for mRNA analysis. The
sequences of PDCD4 primers were as follows: forward,
5′-TATGATGTGGAGGAGGTGGATGTGA-3′ and reverse,
5′-CCTTTCATCCAAAGGCAAAACTACAC-3′. The sequen- ces of matrix
metalloproteinases (MMP)-2 primers were as follows: forward,
5′-CTGATGGCACCCATTTACA CCT-3′ and reverse,
5′-GATCTGAGCGATGCCATCAAA-3′. The sequences of MMP-9 primers were as
follows: forward, 5′-TGGGCTACGTGACCTATGACAT-3′ and reverse,
5′-GCCCAGCCCACCTCCACTCCTC-3′. The sequences of β-actin primers were
as follows: forward, 5′-ATCCGCAAAGACC TGT-3′ and reverse,
5′-GGGTGTAACGCAACTAAG-3′. The primers used for the amplification of
miR-21, RNU6 and cel-miR-39 were purchased from Guangzhou RiboBio
Co., Ltd. (Guangzhou, China). The PCR reaction was performed at
95°C for 5 min, which was followed by 40 cycles of 95°C for 15 sec,
60°C for 30 sec, and 72°C for 30 sec. Dissociation curve was
analyzed from 60 to 99°C. Relative transcript quantities for each
miRNA were calculated using the ΔΔCt method.
Cell migration assay
Transwell migration assay was performed using a
Transwell insert that contains a polycarbonate filter with 8 μM
pore size (cat. no. 3422; Corning). Transfected recipient cells
(5×103) suspended in 150 μl serum-free RPMI-1640 were
added to the 24-well upper chamber. The chambers were placed in
24-well plates, and 600 μl RPMI-1640 that contains 10% FBS was
added to the bottom wells of the multiwell insert assembly. Cells
were incubated at 37°C for 12 h to allow cell migration through the
membrane. Migrated cells were fixed in 95% ethanol and stained with
crystal violet. Migrated cell images were captured by FSX100
(Olympus) at ×200 magnification. Cell migration was quantitated by
counting in 10 random fields on the lower membrane surface.
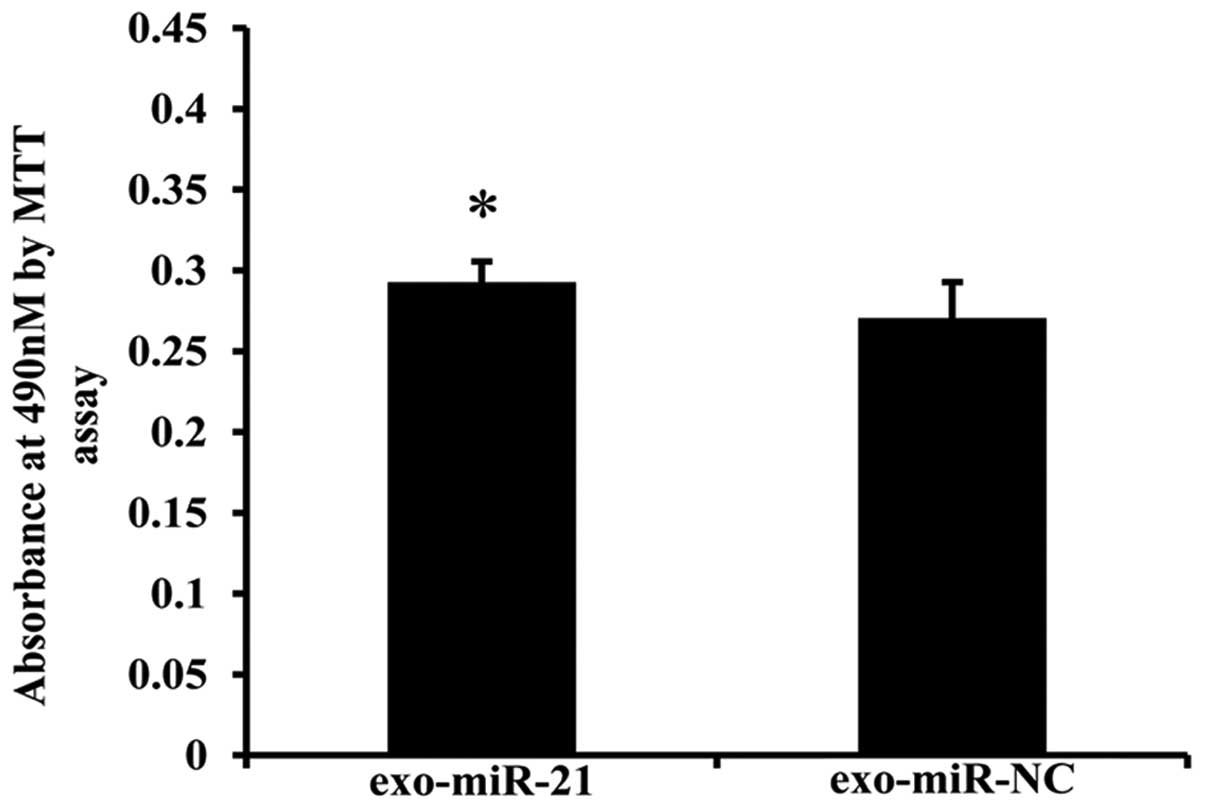
Cell invasion assay
Invasiveness of donor and recipient cells from the
co-culture model was measured by cell invasion through a Matrigel
(BD Biosciences)-coated Transwell inserts with 8 μM pore size (cat.
no. 3422; Corning). Approximately 150 μl cell suspension
(1×105 cells) in serum-free RPMI-1640 was added into the
upper well of the chamber in triplicate wells, whereas the lower
wells were filled to the top (~600 μl) with RPMI-1640 that contains
50% FBS. After 12-h incubation, the non-invasive cells were removed
with a cotton swab, and MTT stock solution was added to each
culture to obtain a final MTT concentration of 0.5 mg/ml in the
medium. Afterwards, the inserts with adherent cells on the lower
membrane surface were incubated for 4 h at 37°C. Subsequently, 500
μl 100% DMSO was added to solubilize the MTT-formazan product per
well. After thorough mixing, the absorbance was measured with a
microplate reader at a 490 nm test wavelength.
Luciferase reporter assay
The 3′-UTR segment of PDCD4 mRNA, which contains the
miR-21-binding sites, was amplified by PCR from human genomic DNA.
Subsequently, this segment was cloned into the pmiR-RB-Report
plasmid (Guangzhou RiboBio), which encodes two luciferase reporter
genes to obtain the wild-type plasmid pmiR-Report-WT-PDCD4.
Mutant-type plasmids pmiR-Report-MUT-PDCD4 were generated by
changing the binding site for miR-21 ‘AUAAGCU’ to ‘UAUUGCA’ from
the corresponding wild-type pmiR-RB-Report plasmid. EC9706 cells in
the miR-21 mimics or negative control group were transfected in
96-well plates (1×104 cells/well) with
pmiR-Report-WT-PDCD4 as well as the corresponding mutant reporter
plasmid by Lipofectamine RNAiMAX. The Renilla luciferase was
used as an internal control. Luciferase activity was measured using
the Dual-Luciferase reporter assay system (Promega) 48 h after
transfection.
Immunoblot analysis
For western blot analysis, miR-21 mimic/negative
control-transfected EC9706 cells were lysed with RIPA lysis buffer
and protease inhibitors (Beyotime Institute of Biotechnology,
Haimen, China) at 14,000 × g for 5 min at 4°C. A total of 20 μg
protein was separated on 10% SDS-PAGE gels and transferred to PVDF
membranes (0.45 μM pore size; Millipore, Billerica, MA, USA).
Membranes were blocked with non-fat milk and incubated with primary
antibodies anti-PDCD4 (1:1,000) (no. 9535S; Cell Signaling
Technology, Danvers, MA, USA), MMP-2 (1:500) (ab7033; Abcam,
Cambridge, UK), MMP-9 (1:500) (MAB3309; Millipore), and β-actin
(1:1000) (BM0627; Wuhan Boster Biological Technology, Ltd., Wuhan,
China) overnight at 4°C. Primary antibody incubation was followed
by incubation with horse-radish peroxidase-conjugated secondary
antibodies for 1 h at room temperature. The protein-antibody
complex was visualized using the enhanced chemiluminescence kit
(Thermo Fisher Scientific) and chemiluminescence image analysis
system (Tanon 5200; Tanon Science, Shanghai, China).
Results
Candidate miRNA screening by
microarray
The microarray data of plasma from esophageal cancer
patients and healthy control have been submitted to GEO website
with accession number GSE71043. Microarray analysis identified 20
miRNAs that could distinguish the plasma from ESCC patients and
healthy controls. A total of 15 miRNAs were found to be
overexpressed in plasma from ESCC patients (hsa-miR-16-5p,
hsa-miR-130a-3p, hsa-miR-15a-5p, hsa-miR-144-3p, hsa-miR-19b-3p,
hsa-miR-5196 -5p, hsa-miR-25a-3p,
hsa-miR-1914-3p,hsa-miR-93-5p,hsa-miR-107,hsa-miR-3911,
hsa-miR-21-5p, hsa-let-7d-3p, hsa-let7i-5p and hsa-miR-1290). By
contrast, hsa-miR-1238-3p, hsa-miR-6069, hsa-miR-191-3p,
hsa-miR-4665-3p and hsa-miR-937-5p were found to be downregulated
(Table I). Cluster analysis, based
on the differentially expressed miRNAs, successfully separated the
plasma samples from ESCC patients and healthy controls (Fig. 1).
 | Table IDifferential expression of miRNAs in
plasma from ESCC patients (g1) vs. healthy controls (g2) by
microarray. |
Table I
Differential expression of miRNAs in
plasma from ESCC patients (g1) vs. healthy controls (g2) by
microarray.
| Systematic name of
miRNA | P-value
(<0.05) | Fold change
(g1/g2) | Expression level of
g1 compared with g2 |
|---|
| hsa-let-7d-3p | 0.000632 | 3.55 | Upregulation |
| hsa-let-7i-5p | 0.028094 | 3.484 | Upregulation |
| hsa-miR-107 | 0.038254 | 9.687 | Upregulation |
|
hsa-miR-1238-3p | 0.026845 | 0.112 | Downregulation |
| hsa-miR-1290 | 0.029073 | 3.307 | Upregulation |
|
hsa-miR-130a-3p | 0.049716 | 104.736 | Upregulation |
| hsa-miR-144-3p | 0.008005 | 36.17 | Upregulation |
| hsa-miR-15a-5p | 0.014182 | 38.202 | Upregulation |
| hsa-miR-16-5p | 0.016302 | 111.321 | Upregulation |
| hsa-miR-191-3p | 0.029836 | 0.166 | Downregulation |
|
hsa-miR-1914-3p | 0.01045 | 10.139 | Upregulation |
| hsa-miR-19b-3p | 0.030333 | 13.434 | Upregulation |
| hsa-miR-21-5p | 0.046401 | 6.779 | Upregulation |
| hsa-miR-25-3p | 0.038363 | 11.525 | Upregulation |
| hsa-miR-3911 | 0.000374 | 8.137 | Upregulation |
|
hsa-miR-4665-3p | 0.005431 | 0.263 | Downregulation |
|
hsa-miR-5196-5p | 0.037794 | 11.583 | Upregulation |
| hsa-miR-6069 | 0.042763 | 0.136 | Downregulation |
| hsa-miR-93-5p | 0.028228 | 9.745 | Upregulation |
| hsa-miR-937-5p | 0.048971 | 0.319 | Downregulation |
Movement trail of EC9706 cell-derived
exosomes
To examine whether exosomes derived from EC9706
cells can be accepted by recipient cells, we labeled exosomes with
DiI dye (red fluorescence) as described in Materials and methods.
When DiI-labeled exosomes were added to the culture medium, red
fluorescence was traced in recipient EC9706 cells using live-cell
fluorescence microscopy. The exosome sizes were below diffraction
limit; thus, DiI-labeled exosomes on glass were represented as
small dots in wide-field fluorescence images (24). We observed an internalization of
DiI-labeled exosomes from the extracellular environment to the
cytoplasm of recipient EC9706 cells (Fig. 2). These studies indicated that
EC9706 cell-derived exosomes could be accepted by recipient
cells.
Extracellular miR-21 can be transfected
to recipient cells by EC9706 cell-derived exosomes
To determine whether miR-21 can be shuttled by donor
EC9706 cell-derived exosomes, Cy3-labeled miR-21 mimics were
transfected to donor EC9706 cells. In addition, culture medium from
donor cells was added to recipient EC9706 cells. After the donor
EC9706 cells were transfected for 48 h, the miR-21 expression level
in donor cells was increased with a 71.87-fold change compared with
the negative control group; thus, the synthetic miR-21 has been
transfected successfully. Flow cytometric analysis showed that the
transfer efficiency of Cy3-labeled miR-21 mimics into recipient
cells was 60.3, 82.6 and 85.0% after 3, 6 and 24 h of culture,
respectively. Therefore, the Cy3-labeled miR-21 was transferred by
the recipient cells (Fig. 3).
Fig. 4 shows that recipient EC9706
cells treated with culture medium from donor EC9706 cells were
fluorescently labeled under fluorescence microscope. RT-qPCR
results showed that the expression level of miR-21 in miR-21
mimics-transfected group was higher than the negative control
group, with an average fold change of 1.11, 1.45 and 1.67 in
recipient cells (Fig. 5A) and the
expression levels were 7.64, 3.15, 1.69 in culture supernatant
(Fig. 5B) after 3, 6 and 24 h of
culture, respectively. The results showed that miR-21 was suggested
to be secreted from donor EC9706 cells and was delivered into
recipient EC9706 cells via exosomes.
nSMase inhibits the release of EC9706
cell derive-exosomes
Recent reports show that miRNAs can be incorporated
into exosomes and released via a ceramide-dependent pathway
(25). Ceramide is a kind of fatty
acid molecule whose biosynthesis is regulated by nSMase2, which is
known to hydrolyze sphingomyelins to generate ceramides and trigger
the budding of exosomes (26). In
the present study, to evaluate whether the miR-21 secretion depends
on exosome transfer, we treated EC9706 cells with GW4869, which is
an nSMase2 inhibitor, and examined the miR-21 expression level in
donor cells and their culture medium. As a result of this
treatment, the miR-21 expression levels in extracellular (Fig. 6A) and intra-cellular conditions
(Fig. 6B) were dose-dependent at
various GW4869 concentrations. The extracellular miR-21 expression
in exosomes was reduced when the GW4869 concentration was
increased, whereas the miR-21 cellular expression was increased
after incubation with GW4869. These findings were based on the
comparison with the controls. These data showed that the GW4869
treatment reduced the expression level of exosome-shuttling
miR-21.
Exosome-shuttling miR-21 promotes
recipient cell migration
Cell migration assay was detected using an 8 μM pore
transwell as described in Materials and methods. We tested the
potential migration capacity of miR-21-transfected donor EC9706
cell-derived exosomes on the recipient cells through the
co-cultivation model. Results showed that co-culture with
miR-21-transfected donor EC9706 cells enhanced the migration of
recipient cells relative to the miR-NC-transfected group with an
average fold change of 1.54 (Fig.
7). These data suggest that exosomes, derived from donor EC9706
cells with enforced miR-21 expression, can be transferred to and
affect the migration of recipient cells.
Exosome-shuttling miR-21 promotes
recipient cell invasion
In the invasiveness assay, cell co-cultivation was
performed using an 8 μM pore Transwell with Matrigel-coated inserts
as described in Materials and methods. We tested the potential
biological function of miR-21-transfected donor EC9706 cell-derived
exosomes on the recipient cells through co-cultivation model.
Results showed that co-culture with miR-21-transfected donor EC9706
cells enhanced the invasiveness of recipient cells compared with
the miR-NC-transfected group with an average fold change of 1.08
(Fig. 8). Our results showed that
EC9706-derived exosome-shuttling miR-21 can be transferred to and
affect the invasive ability of recipient cells.
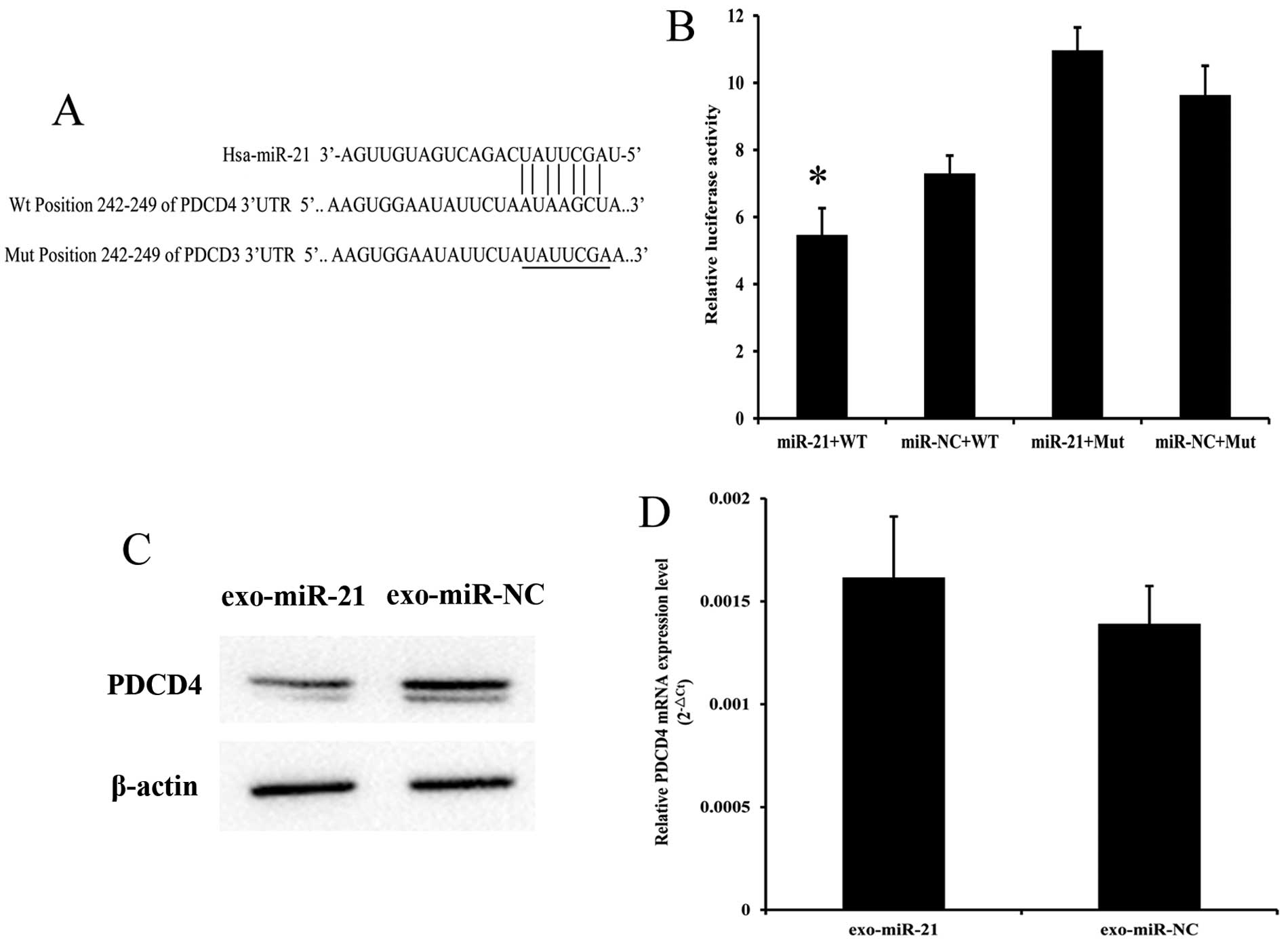
Exosome-shuttling miR-21 represses PDCD4
protein expression by binding to 3′-UTR
To determine whether donor cell-derived
exosome-shuttling miRNA exerts regulatory action on its target gene
in recipient cells, we examined the expression of PDCD4, which is a
predicted target gene for miR-21 (27–29).
As predicted by TargetScan, the 3′-UTR regions from PDCD4 contain
the miR-21 binding site(s) with a seven-mer seed match (Fig. 9A). Luciferase reporter assay was
used to identify whether PDCD4 is a direct target gene of miR-21.
Results revealed that miR-21 overexpression in EC9706 cells
significantly reduced luciferase activity of pmiR-Report-WT-PDCD4
plasmid with a 25.1% reduction. However, this process proceeded
without changes in the lucif-erase activity of
pmiR-Report-MUT-PDCD4. These results indicated that miR-21 directly
binds the 3′-UTR of PDCD4 genes (Fig.
9B). We performed western blot analysis to detect the PDCD4
protein expression level in recipient EC9706 cells after a 24 h
co-culture. Fig. 9C shows that
exosomes from miR-21-transfected EC9706 cells significantly
downregulated PDCD4 protein expression with a 22.27% reduction in
recipient cells under the co-cultivation model. RT-qPCR was
performed to detect the RNA levels of PDCD4. Results showed that no
statistically significant change in PDCD4 mRNA expression existed
between the miR-21 mimics and the negative control group
(P>0.05; Fig. 9D). These
results suggest that miR-21 secreted by donor EC9706 cells can be
effectively uptaken by recipient EC9706 cells and regulate PDCD4
expression at a post-transcriptional level in the recipient
cells.
miR-21 overexpression increases MMP-2 and
MMP-9 expression
Expression of MMP-2 and MMP-9 involved in metastasis
was tested to investigate whether exosome-shuttling miR-21
regulates esophageal cancer cell migration and invasion. RT-qPCR
was used to determine the MMP-2 mRNA and MMP-9 mRNA expression
levels in recipient cells after co-cultivation. Compared with the
miR-negative control group, the MMP-2 mRNA and MMP-9 mRNA
expression levels showed an 11.24- and 1.76-fold increase,
respectively, in miR-21 mimics group (Fig. 10A and B). Western blot analysis
was used to determine the expression levels of MMP-2 and MMP-9
proteins in recipient cells after co-cultivation. Compared with the
miR-negative control group, the expression levels of MMP-2 and
MMP-9 proteins showed a 13.84- and 5.83-fold increase,
respectively, in miR-21 mimics group (Fig. 10C and D). These data suggest that
exosome-shuttling miR-21 inhibits target PDCD4 expression at
translational level and plays an important role in the JNK
downstream signaling pathway involved in tumor migration and
invasion.
Case-control study on the association
between exosome-shuttling miR-21 and esophageal cancer
incidence
The smoking status, alcohol use and family history
of ESCC patients and healthy controls are shown in Table II. The average ages of cancer
patients and controls were not significantly different. The
difference was statistically significant in the distribution of
smoking, alcohol use, and cancer family history between patients
and healthy controls. Paired t-test was used to assess the
differences between the plasma recruited from esophageal cancer
patients and healthy controls. Table
III shows that the relative expression of miR-21 was
upregulated, with a fold change of 2.95, in the patients' plasma
when compared with plasma from healthy controls. Furthermore,
conditional logistic regression analysis showed that a
significantly increased risk for esophageal cancer was associated
with increased miR-21 expression (OR, 1.107; 95% CI, 1.012–1.21;
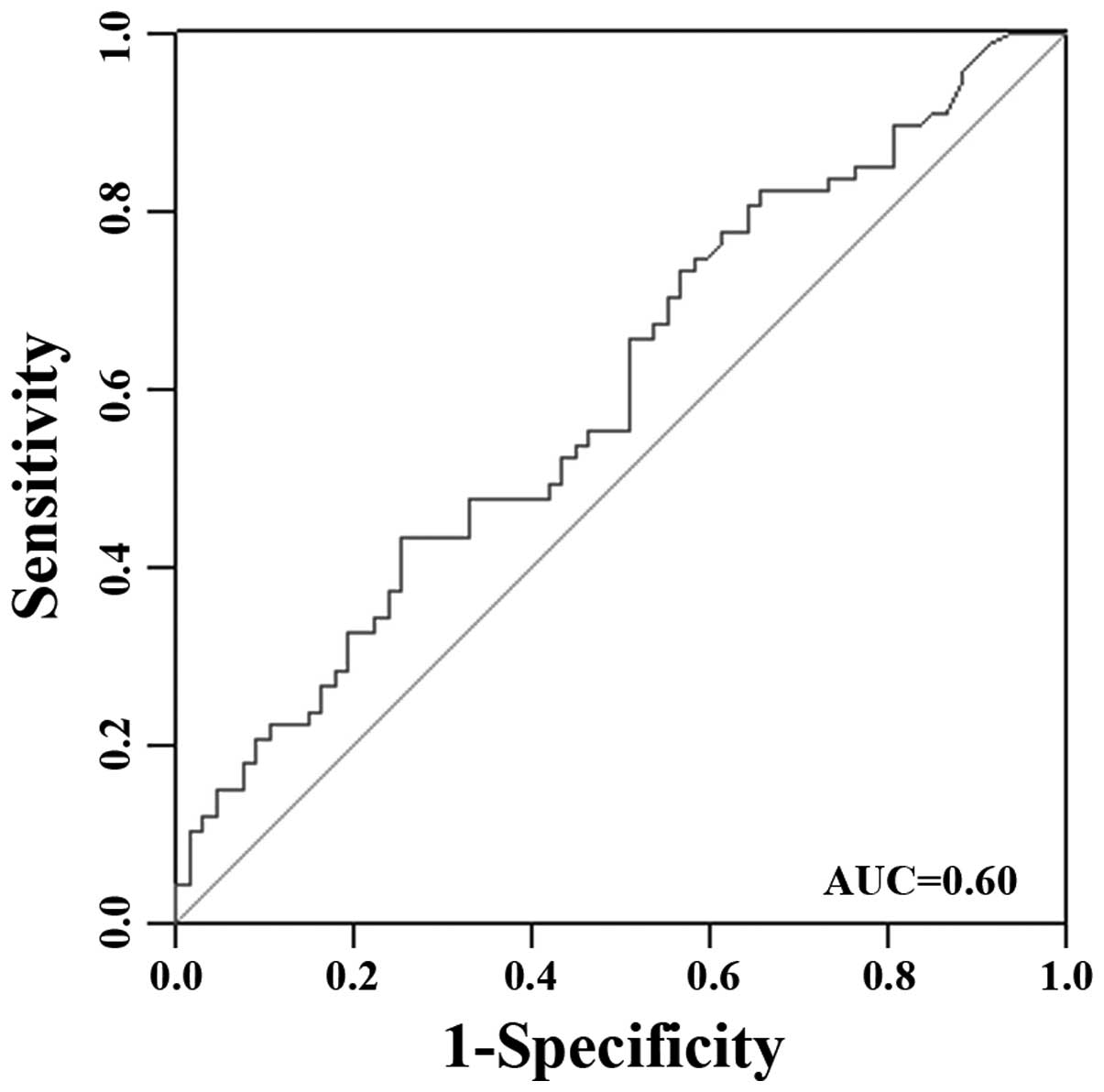
P=0.026). Receiver operating characteristic (ROC) curve was
generated to see the power of exosome-shuttling miR-21 to
distinguish ESCC patients from healthy controls. The area under the
curve was used as a measurement for level of discrimination. As
shown in Fig. 11, circulating
miR-21 discrimination of esophageal cancer from healthy person was
noted by an area-under-the-curve (AUC) value of 0.60.
| Table IICharacteristics of ESCC patients and
healthy controls. |
Table II
Characteristics of ESCC patients and
healthy controls.
| Variables | No. of ESCC
patients (n=67) | No. of healthy
controls (n=67) |
|---|
| Age (years) | 61.82±8.14 | 63.03±9.66 |
| Smoking
indexb |
| Never | 32a | 48 |
| Everc |
| <400 | 8 | 5 |
| ≥400 | 27 | 14 |
| Alcohol
consumption |
| Never | 33a | 48 |
| Everd | 34 | 19 |
| Family history |
| No | 58a | 67 |
| Yes | 9 | 0 |
| Table IIIRelative expression of miR-21 in
plasma from ESCC patients and healthy controls. |
Table III
Relative expression of miR-21 in
plasma from ESCC patients and healthy controls.
| Group | miRNA | Mean ± SD of
ΔCta | ΔΔCta |
2−ΔΔCt | OR | 95% CI | P-value |
|---|
| Caseb,c | miR-21 | −2.39±4.03 | −1.56 | 2.95 | 1.107 | 1.012–1.21 | 0.026 |
| Control | miR-21 | −0.83±3.86 | | | | | |
Discussion
Emerging evidence suggests that tumor
microenvironment plays an important role in tumor occurrence and
development (30). Cell-cell
communication is an indispensably dynamic mechanism, which results
in normal cellular activities and tissue homeostasis maintenance
(31). The mechanisms of cell-cell
communication were as follows: i) integral membrane
protein-mediated direct cell-cell contact; ii) indirect contact via
the extracellular matrix; and iii) circulating miRNA via exosomes
or extracellular miRNA such as the Ago2-binding miRNA in the
extracellular microenvironment (19). Recently, the roles of tumor
cell-derived exosomes in cell-cell communication of tumor
microenvironment are gradually revealed. Research has shown that
exosomes could be internalized into the cells through endocytosis
but not incorporated into the plasma membrane (24). We applied a cell culture chamber of
the 3D live-cell imaging system to observe the exosome transfer
process. Our results showed that DiI-labeled exosomes could be
taken up from the extracellular environment to the cytoplasm by
EC9706 cells. Studies have indicated that microvesicles released
from the endosomal component or fused with the plasma membrane can
directly affect target cells by receptor-mediated interaction or by
transferring biological information molecules, such as miRNAs
(32). Studies have proposed that
the transfer of exosome-derived unique miRNA to recipient cells is
an alternative mechanism that involves direct cell-cell contact or
chemical receptor-mediated events, which allow gene-based
communication between cells (16).
miRNAs found in the macrophage-derived microvesicles were shuttled
not only to monocytic cells but also to other cell lineages. Thus,
macrophage-derived microvesicles could communicate with various
cell types, which include allogeneic cells or cell types that are
different from the donor cell; this finding signifies a widespread
impact (33). Shimbo et al
(34) found that synthetic miR-143
transfected into bone-marrow-derived mesenchymal stem cells (MSCs)
was enveloped in exosomes. MSC-derived exosome-formed miR-143 could
be transferred to osteosar-coma cells and could affect osteosarcoma
cell migration. A study on breast cancer showed that miR-223, which
was within the exosomes released by macrophages, could promote
breast cancer cell invasion via the Mefc-β-catenin pathway under a
co-culture system (23). As a
result, miRNA may be secreted and delivered into recipient cells
via exosomes after donor cell transfection. These secreted miRNAs
may alter the cellular functions of the recipient cells by
modulating the expression of their target genes. This process may
play an important role in mediating tumor microenvironment.
miR-21, which is considered an oncomiR, is
demonstrated to be overexpressed in numerous tumor tissues, cells,
or body fluids from tumor patients (35–37).
Many studies have been conducted on the function of miR-21 in tumor
occurrence and development. These studies showed that miR-21 could
promote cell growth, proliferation, invasion and metastasis of
various tumor cells by targeting PTEN, PDCD4, RECK, FASL and TIMP3
(38–40). Hiyoshi et al (41) found that the inhibition of
microRNA-21 could lead to a reduction of the proliferation and
invasion in esophageal squamous cell carcinoma cell lines by
increasing PDCD4 protein expression without changing the PDCD4 mRNA
level by binding to the PDCD4-3′ untranslated region. This finding
suggests that microRNA-21 targets PDCD4 at the post-transcriptional
level and regulates cell proliferation and invasion in esophageal
squamous cell carcinoma. In the present study, we transfected donor
EC9706 cells with Cy3-labeled miR-21 and showed the effective
transfer of miR-21 from donor cells to recipient cells by flow
cytometry and RT-qPCR. nSMase2 was shown to hydrolyze
sphingomyelins to generate ceramides and trigger exosome budding
(23,42). In a study on nSMase2 and
tumor-derived exosomes, it was found that the expression level of
nSMase2 was higher in cancer cells than in non-cancer cells, and
the secretion level of exosomes was correlated with the expression
level of nSMase2 (43). In the
present study, we used GW4869, which is an nSMase2 inhibitor, to
confirm that miR-21 secretion is an exosome-dependent process.
Results showed that with the inhibition of exosome secretion, the
miR-21 expression level in culture supernatant gradually decreased.
The abovementioned finding demonstrates that miR-21 could be
shuttled between cells via exosomes, which provides the possibility
for the function research of miR-21 in tumor cell-cell
communication.
Subsequently, we constructed a Transwell co-culture
system to imitate the tumor microenvironment and demonstrated that
exosome-shuttling miR-21 promoted the migration and invasion of
recipient EC9706 cells through Transwell insert assay. PDCD4, a
tumor suppressor gene, was demonstrated to have low expression in
various tumor tissues and cells (44,45).
PDCD4, which was predicted to be a miR-21 target gene by
TargetScan, has been reported to be a key regulator of tumor
proliferation, apoptosis, adhesion, migration, invasion and
metastasis (28–30,40,46).
Tumor invasion is a multistep process in which cell motility is
coupled with proteolysis and involves interaction of cells with
extracellular matrix (ECM). MMP-2 and MMP-9 are released as latent
enzymes, which are capable of degradation of ECM involved in the
invasion process. Studies showed that these enzymes were involved
in cell migration through the removal of adhesion sites, cleavage
of cell-cell or cell-matrix receptors, and release of
chemoattractants from ECM (47,48).
The invasiveness of U87MG glioblastoma cells was inhibited after
the cells were treated with miR-21 inhibitor, which was mediated by
an increase in PDCD4 expression and reduction of MMP-2 protein
expression (49). Zhu et al
(50) revealed that inhibition of
miR-21 expression repressed cell migration and invasion through the
miR-21-PDCD4-AP-1 feedback loop, which is involved in several key
downstream signaling pathway molecules, such as phosphorylated
c-Jun, MMP-2 and MMP-9. Another research on hepatocellular
carcinoma found that PDCD4 overexpression suppressed JNK activity;
moreover, JNK inhibition resulted in the suppression of the
phosphorylated eIF4E, which may influence the invasion via
regulation of MMP expression (51,52).
In the present study, we demonstrated that exosome-shuttling miR-21
from donor cells downregulated PDCD4 and increased the downstream
signal levels of MMP-2 and MMP-9 in recipient cells. Our results
suggested that the modulation of PDCD4 expression and downstream
signaling regulation by exosome-mediated signal transfer may
provide a novel mechanism of tumor progression.
Several studies have found that exosomal miRNAs
could be secreted into the plasma and may be potential biomarkers
in the diagnosis of different cancer types. A study on exosomal
miRNAs from ovarian cancer showed that the levels of eight
microRNAs, such as miR-21, miR-141 and miR-200a, were similar
between cellular and exosomal miRNAs, with the correlation
coefficients distributed from 0.71 to 0.90. Thus, tumor exosomal
miRNAs could be used as surrogate diagnostic markers for biopsy
profiling (15). Another study on
serum exosomes from esophageal cancer showed that miR-21 was not
detected in the serum that remained after exosomes were extracted;
this result suggested that exosomes from tumor cells are a major
source of serum circulating miR-21 in esophageal cancer patients
(43). Ogata-Kawata et al
(53) found that the amount of
seven serum exosomal miRNAs (i.e., let-7a, miR-1229, miR-1246,
miR-150, miR-21, miR-223 and miR-23a) from the serum of TNM stage I
colorectal cancer patients were significantly higher than in
healthy controls; this finding suggested that serum exosomal miRNAs
may serve as significant biomarkers for the early detection of
primary colorectal cancer (53).
One study of pancreatic adenocarcinoma (PC) found that serum
exosomal miR-17-5p and miR-21 were significantly elevated in the
advanced stage and metastatic PC patients; moreover, the diagnostic
sensitivity and specificity were 72.7 and 92.6% for miR-17-5p and
95.5 and 81.5% for miR-21, respectively (54). In the present study, microarray
analysis of plasma from esophageal cancer patients and healthy
controls identified 20 miRNAs that could distinguish the plasma
from ESCC patients and healthy controls. Among these miRNAs,
miR-25-3p, miR-93-5p, miR-21-5p and miR-1290 were upregulated in
cancer tissues and plasma/serum of various cancer patients and were
confirmed to be oncogenes. miR-16-5p, miR-130a-3p, miR-15a-5p,
miR-144-3p, miR-19b-3p, miR-107 and let-7i-5p may be tumor
promotors or suppressors in different tumors with their
dysregulated expression level. Few studies have focused on
miR-5196-5p, miR-1914-3p, miR-3911, let-7d-3p, miR-1238-3p,
miR-6069, miR-191-3p, miR-4665-3p and miR-937-5p. The results of
the present study indicated that miR-21 is upregulated
significantly in the plasma of esophageal cancer patients. In
addition, a significant association was identified between miR-21
and esophageal cancer risk. These findings suggest that increased
exosome-shuttling miR-21 in plasma might be a candidate biomarker
for esophageal cancer diagnosis and high-risk population
screening.
In summary, we have recognized the uptake process of
exosomes by esophageal cancer cells and found that Cy3-labeled
miR-21 mimics could be transferred between esophageal cancer cells
by exosomes. Furthermore, our results revealed that miR-21 mimics
could affect migration and invasion of recipient cells partly via
modulation of its target gene PDCD4 and its downstream-signaling
molecules, MMP-2 and MMP-9 by using the cell co-culture system. As
a novel mechanism of cell-cell communication, the transfer of
exosome-shuttling miR-21 is in addition to the classical mechanism
and the effective delivery of exosome-shuttling miR-21 in tumor
microenvironment may affect the status of esophageal cancer cells
consequently to promote the recurrence and distant metastasis of
esophageal cancer, which suggests that exosome-shuttling miR-21
could be a potential biomarker in esophageal cancer diagnosis.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (nos. 81172747, 81573108 and
81573191), the Natural Science Foundation of Jiangsu Province,
China (no. BK2010407) and the New Century Excellent Talents in
University from Ministry of Education, China (NCET-13-0124).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
2
|
Wang KK and Sampliner RE; Practice
Parameters Committee of the American College of Gastroenterology.
Updated guidelines 2008 for the diagnosis, surveillance and therapy
of Barrett's esophagus. Am J Gastroenterol. 103:788–797. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kosaka N, Iguchi H and Ochiya T:
Circulating microRNA in body fluid: A new potential biomarker for
cancer diagnosis and prognosis. Cancer Sci. 101:2087–2092. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cazzoli R, Buttitta F, Di Nicola M,
Malatesta S, Marchetti A, Rom WN and Pass HI: microRNAs derived
from circulating exosomes as noninvasive biomarkers for screening
and diagnosing lung cancer. J Thorac Oncol. 8:1156–1162. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Manterola L, Guruceaga E, Gállego
Pérez-Larraya J, González-Huarriz M, Jauregui P, Tejada S,
Diez-Valle R, Segura V, Samprón N, Barrena C, et al: A small
noncoding RNA signature found in exosomes of GBM patient serum as a
diagnostic tool. Neuro Oncol. 16:520–527. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu R, Liao J, Yang M, Shi Y, Peng Y, Wang
Y, Pan E, Guo W, Pu Y and Yin L: Circulating miR-155 expression in
plasma: A potential biomarker for early diagnosis of esophageal
cancer in humans. J Toxicol Environ Health A. 75:1154–1162. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ali S, Almhanna K, Chen W, Philip PA and
Sarkar FH: Differentially expressed miRNAs in the plasma may
provide a molecular signature for aggressive pancreatic cancer. Am
J Transl Res. 3:28–47. 2010.PubMed/NCBI
|
|
8
|
Wang HY, Yan LX, Shao Q, Fu S, Zhang ZC,
Ye W, Zeng YX and Shao JY: Profiling plasma microRNA in
nasopharyngeal carcinoma with deep sequencing. Clin Chem.
60:773–782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng L, Sharples RA, Scicluna BJ and Hill
AF: Exosomes provide a protective and enriched source of miRNA for
biomarker profiling compared to intracellular and cell-free blood.
J Extracell Vesicles. 3:32014.
|
|
10
|
Ge Q, Zhou Y, Lu J, Bai Y, Xie X and Lu Z:
miRNA in plasma exosome is stable under different storage
conditions. Molecules. 19:1568–1575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Salido-Guadarrama I, Romero-Cordoba S,
Peralta-Zaragoza O, Hidalgo-Miranda A and Rodríguez-Dorantes M:
MicroRNAs transported by exosomes in body fluids as mediators of
intercellular communication in cancer. Onco Targets Ther.
7:1327–1338. 2014.PubMed/NCBI
|
|
12
|
Ramachandran S and Palanisamy V:
Horizontal transfer of RNAs: Exosomes as mediators of intercellular
communication. Wiley Interdiscip Rev RNA. 3:286–293. 2012.
View Article : Google Scholar :
|
|
13
|
Kosaka N, Yoshioka Y, Hagiwara K, Tominaga
N, Katsuda T and Ochiya T: Trash or treasure: Extracellular
microRNAs and cell-to-cell communication. Front Genet. 4:1732013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rabinowits G, Gerçel-Taylor C, Day JM,
Taylor DD and Kloecker GH: Exosomal microRNA: A diagnostic marker
for lung cancer. Clin Lung Cancer. 10:42–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tadokoro H, Umezu T, Ohyashiki K, Hirano T
and Ohyashiki JH: Exosomes derived from hypoxic leukemia cells
enhance tube formation in endothelial cells. J Biol Chem.
288:34343–34351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nouraee N, Van Roosbroeck K, Vasei M,
Semnani S, Samaei NM, Naghshvar F, Omidi AA, Calin GA and Mowla SJ:
Expression, tissue distribution and function of miR-21 in
esophageal squamous cell carcinoma. PLoS One. 8:e730092013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Singh R, Pochampally R, Watabe K, Lu Z and
Mo YY: Exosome-mediated transfer of miR-10b promotes cell invasion
in breast cancer. Mol Cancer. 13:2562014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Umezu T, Ohyashiki K, Kuroda M and
Ohyashiki JH: Leukemia cell to endothelial cell communication via
exosomal miRNAs. Oncogene. 32:2747–2755. 2013. View Article : Google Scholar
|
|
20
|
Liao J, Liu R, Yin L and Pu Y: Expression
profiling of exosomal miRNAs derived from human esophageal cancer
cells by Solexa high-throughput sequencing. Int J Mol Sci.
15:15530–15551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han Y, Wei F, Xu X, Cai Y, Chen B, Wang J,
Xia S, Hu H, Huang X, Han Y, et al: Establishment and comparative
genomic hybridization analysis of human esophageal carcinomas cell
line EC9706. Zhonghua Yi Xue Yi Chuan Xue Za Zhi. 19:455–457.
2002.(In Chinese). PubMed/NCBI
|
|
22
|
Ji H, Erfani N, Tauro BJ, Kapp EA, Zhu HJ,
Moritz RL, Lim JW and Simpson RJ: Difference gel electrophoresis
analysis of Ras-transformed fibroblast cell-derived exosomes.
Electrophoresis. 29:2660–2671. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang M, Chen J, Su F, Yu B, Su F, Lin L,
Liu Y, Huang JD and Song E: Microvesicles secreted by macrophages
shuttle invasion-potentiating microRNAs into breast cancer cells.
Mol Cancer. 10:1172011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian T, Wang Y, Wang H, Zhu Z and Xiao Z:
Visualizing of the cellular uptake and intracellular trafficking of
exosomes by live-cell microscopy. J Cell Biochem. 111:488–496.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kosaka N, Iguchi H, Yoshioka Y, Takeshita
F, Matsuki Y and Ochiya T: Secretory mechanisms and intercellular
transfer of microRNAs in living cells. J Biol Chem.
285:17442–17452. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kosaka N, Iguchi H, Yoshioka Y, Hagiwara
K, Takeshita F and Ochiya T: Competitive interactions of cancer
cells and normal cells via secretory microRNAs. J Biol Chem.
287:1397–1405. 2012. View Article : Google Scholar :
|
|
27
|
Allgayer H: Pdcd4, a colon cancer
prognostic that is regulated by a microRNA. Crit Rev Oncol Hematol.
73:185–191. 2010. View Article : Google Scholar
|
|
28
|
Li L, Zhou L, Li Y, Lin S and Tomuleasa C:
MicroRNA-21 stimulates gastric cancer growth and invasion by
inhibiting the tumor suppressor effects of programmed cell death
protein 4 and phosphatase and tensin homolog. J BUON. 19:228–236.
2014.PubMed/NCBI
|
|
29
|
Liu T, Liu Q, Zheng S, Gao X, Lu M, Yang
C, Dai F, Sheyhidin I and Lu X: MicroRNA-21 promotes cell growth
and migration by targeting programmed cell death 4 gene in Kazakh's
esophageal squamous cell carcinoma. Dis Markers. 2014:2328372014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kahlert C and Kalluri R: Exosomes in tumor
microenvironment influence cancer progression and metastasis. J Mol
Med Berl. 91:431–437. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee JK, Park SR, Jung BK, Jeon YK, Lee YS,
Kim MK, Kim YG, Jang JY and Kim CW: Exosomes derived from
mesenchymal stem cells suppress angiogenesis by down-regulating
VEGF expression in breast cancer cells. PLoS One. 8:e842562013.
View Article : Google Scholar
|
|
32
|
Tetta C, Ghigo E, Silengo L, Deregibus MC
and Camussi G: Extracellular vesicles as an emerging mechanism of
cell-to-cell communication. Endocrine. 44:11–19. 2013. View Article : Google Scholar :
|
|
33
|
Ismail N, Wang Y, Dakhlallah D, Moldovan
L, Agarwal K, Batte K, Shah P, Wisler J, Eubank TD, Tridandapani S,
et al: Macrophage microvesicles induce macrophage differentiation
and miR-223 transfer. Blood. 121:984–995. 2013. View Article : Google Scholar :
|
|
34
|
Shimbo K, Miyaki S, Ishitobi H, Kato Y,
Kubo T, Shimose S and Ochi M: Exosome-formed synthetic microRNA-143
is transferred to osteosarcoma cells and inhibits their migration.
Biochem Biophys Res Commun. 445:381–387. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao J, Zhang Q, Xu J, Guo L and Li X:
Clinical significance of serum miR-21 in breast cancer compared
with CA153 and CEA. Chin J Cancer Res. 25:743–748. 2013.
|
|
36
|
Gao W, Xu J, Liu L, Shen H, Zeng H and Shu
Y: A systematic-analysis of predicted miR-21 targets identifies a
signature for lung cancer. Biomed Pharmacother. 66:21–28. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Selcuklu SD, Donoghue MTA and Spillane C:
miR-21 as a key regulator of oncogenic processes. Biochem Soc
Trans. 37:918–925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang N, Zhang CQ, He JH, Duan XF, Wang YY,
Ji X, Zang WQ, Li M, Ma YY, Wang T, et al: MiR-21 down-regulation
suppresses cell growth, invasion and induces cell apoptosis by
targeting FASL, TIMP3, and RECK genes in esophageal carcinoma. Dig
Dis Sci. 58:1863–1870. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lou Y, Cui Z, Wang F, Yang X and Qian J:
miR-21 down-regulation promotes apoptosis and inhibits invasion and
migration abilities of OVCAR3 cells. Clin Invest Med.
34:E2812011.PubMed/NCBI
|
|
40
|
Xu LF, Wu ZP, Chen Y, Zhu QS, Hamidi S and
Navab R: MicroRNA-21 (miR-21) regulates cellular proliferation,
invasion, migration, and apoptosis by targeting PTEN, RECK and
Bcl-2 in lung squamous carcinoma, Gejiu City, China. PLoS One.
9:e1036982014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hiyoshi Y, Kamohara H, Karashima R, Sato
N, Imamura Y, Nagai Y, Yoshida N, Toyama E, Hayashi N, Watanabe M,
et al: MicroRNA-21 regulates the proliferation and invasion in
esophageal squamous cell carcinoma. Clin Cancer Res. 15:1915–1922.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tanaka Y, Kamohara H, Kinoshita K,
Kurashige J, Ishimoto T, Iwatsuki M, Watanabe M and Baba H:
Clinical impact of serum exosomal microRNA-21 as a clinical
biomarker in human esophageal squamous cell carcinoma. Cancer.
119:1159–1167. 2013. View Article : Google Scholar
|
|
43
|
Kosaka N, Iguchi H, Hagiwara K, Yoshioka
Y, Takeshita F and Ochiya T: Neutral sphingomyelinase 2
(nSMase2)-dependent exosomal transfer of angiogenic microRNAs
regulate cancer cell metastasis. J Biol Chem. 288:10849–10859.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ma G, Zhang H, Dong M, Zheng X, Ozaki I,
Matsuhashi S and Guo K: Downregulation of programmed cell death 4
(PDCD4) in tumorigenesis and progression of human digestive tract
cancers. Tumour Biol. 34:3879–3885. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang D, Guo S, Han SY, Xu N, Guo JY and
Sun Q: Distinct roles of different fragments of PDCD4 in regulating
the metastatic behavior of B16 melanoma cells. Int J Oncol.
42:1725–1733. 2013.PubMed/NCBI
|
|
46
|
Lankat-Buttgereit B and Göke R: The tumour
suppressor Pdcd4: Recent advances in the elucidation of function
and regulation. Biol Cell. 101:309–317. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang YY, Chen B and Ding YQ:
Metastasis-associated factors facilitating the progression of
colorectal cancer. Asian Pac J Cancer Prev. 13:2437–2444. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yadav L, Puri N, Rastogi V, Satpute P,
Ahmad R and Kaur G: Matrix metalloproteinases and cancer - roles in
threat and therapy. Asian Pac J Cancer Prev. 15:1085–1091. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dong CG, Wu WK, Feng SY, Wang XJ, Shao JF
and Qiao J: Co-inhibition of microRNA-10b and microRNA-21 exerts
synergistic inhibition on the proliferation and invasion of human
glioma cells. Int J Oncol. 41:1005–1012. 2012.PubMed/NCBI
|
|
50
|
Zhu Q, Wang Z, Hu Y, Li J, Li X, Zhou L
and Huang Y: miR-21 promotes migration and invasion by the
miR-21-PDCD4-AP-1 feedback loop in human hepatocellular carcinoma.
Oncol Rep. 27:1660–1668. 2012.PubMed/NCBI
|
|
51
|
Jiang Y, Zhang SH, Han GQ and Qin CY:
Interaction of Pdcd4 with eIF4E inhibits the metastatic potential
of hepatocellular carcinoma. Biomed Pharmacother. 64:424–429. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Khosravi S, Tam KJ, Ardekani GS, Martinka
M, McElwee KJ and Ong CJ: eIF4E is an adverse prognostic marker of
melanoma patient survival by increasing melanoma cell invasion. J
Invest Dermatol. 135:1358–1367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ogata-Kawata H, Izumiya M, Kurioka D,
Honma Y, Yamada Y, Furuta K, Gunji T, Ohta H, Okamoto H, Sonoda H,
et al: Circulating exosomal microRNAs as biomarkers of colon
cancer. PLoS One. 9:e929212014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Que R, Ding G, Chen J and Cao L: Analysis
of serum exosomal microRNAs and clinicopathologic features of
patients with pancreatic adenocarcinoma. World J Surg Oncol.
11:2192013. View Article : Google Scholar : PubMed/NCBI
|