Introduction
The phosphoinositide 3 kinase (PI3K)/Akt/mammalian
target of the rapamycin (mTOR) pathway regulates the cell cycle and
is associated with cellular proliferation and cancer. The
activation of PI3K phosphorylates and activates Akt, which have
numerous downstream effects including activating mTOR (1). The overactivation of this pathway
stimulates proliferation and reduces apoptosis, involved in the
pathogenesis of cancer.
Colorectal adenocarcinoma is the third most common
type of cancer globally making up ~10% of all cases (World Cancer
Report 2014). PI3K/Akt and mTOR are activated in colorectal
adenocarcinoma, and they would be important targets for treatment
of colorectal adenocarcinoma (2).
However, direct antagonizing mTOR would activate feedback loop of
PI3K/Akt/mTOR pathway and affect the anticancer effect (3,4).
Hence, a dual inhibition of Akt and mTOR may be more effective to
treat colorectal adenocarcinoma. In the present study, we chose
NVP-BEZ235, which has dual effects on PI3K and mTOR.
NVP-BEZ235 (dactolisib) is a novel dual inhibitor of
PI3K/mTOR. Its effects have been reported in tumors, such as
glioma, breast cancer and Burkitt lymphoma (5–7) and
Waldenström's macroglobulinemia (8). By targeting PI3K/mTOR, NVP-BEZ235
inhibits the cancer stem cells of prostate cancer, glioblastoma and
colorectal adenocarcinoma (9–11).
In addition, NVP-BEZ235 reduces cell proliferation in HCT-116 and
DLD-1 colorectal adenocarcinoma cells after cytotoxic therapy with
two inhibitors of ubiquitin-proteasome system,
methyl-13-hydroxy-15-oxokaurenoate (MHOK) and bortezomib (11). However, the effects of NVP-BEZ235
alone are unclear on proliferation, migration, apoptosis and
autophagy in HT-29 colorectal adenocarcinoma cells.
Cis-diamminedichloroplatinum (DDP) binds and cross-links
DNA. It is used to treat various types of cancers, such as
sarcomas, lung, ovarian and cervical cancer (12–14).
Nevertheless, effects of drug combination of NVP-BEZ235 and DDP in
human colorectal adenocarcinoma remain elusive.
Tumor cells are hallmarked by unrestricted
proliferation. Apoptosis and autophagy are involved in cancer
genesis and prevention. Apoptosis is known as programmed cell
death, and autophagy is an intracellular degradation process to
eliminate damaged macromolecules and organelles during cellular
distress. The excessive self-cannibalistic function may be
deleterious and cause cell death (15). However, it is unknown how the dual
inhibition of PI3K and mTOR affects apoptosis or autophagy in
colorectal adenocarcinoma. The activation of caspase-3 participates
in the execution-phase of cell apoptosis (16). Microtubule-associated proteins
1A/1B light chain 3B (MAP1LC3B and LC3B) and Beclin-1 are important
proteins associated with autophagy (17). Therefore, we explored the protein
expression of cleaved caspase-3, cleaved LC3B and Beclin-1 in order
to explore effects of NVP-BEZ235 on apoptosis and autophagy in
HT-29 colorectal adenocarcinoma cells. Akt is an essential
component in PI3K/Akt/mTOR pathway and is a downstream effector of
PI3K (18). mTOR is a
serine/threonine kinase that regulates cellular metabolism, growth
and proliferation (19). Akt and
mTOR could be phosphorylated by their activating kinases.
Phosphorylated Akt and mTOR are active and functional molecules
that activate downstream signals of PI3K/Akt/mTOR pathway involving
in cell cycle and proliferation (18,19).
Cyclin D1 is required for the progression of cell cycle through G1
phase (20). Therefore, we
explored the protein expression of phospho (p)-Akt, p-mTOR and
cyclin D1 in order to explore effects of NVP-BEZ235 and DPP on
proliferation and migration in HT-29 colorectal adenocarcinoma
cells.
Materials and methods
Cells and reagents
Cells
HT-29 cells (human colorectal adenocarcinoma cell
line; Cell Bank of Chinese Academy of Sciences, Shanghai, China)
were cultured in RPMI-1640 supplemented with 100 ml/l fetal bovine
serum (FBS), 100 kU/l penicillin and 100 mg/l chloramphenicol in a
cell incubator with 5% CO2 at 37°C. Cells were
subcultured after digestion by 0.25% trypsin, and cell growth was
observed under an inverse microscope.
Drug
The powder of NVP-BEZ235 (dactolisib; Selleck Inc.,
Houston, TX, USA) was dissolved in dimethylformamide (DMF;
Sinopharm, Inc., Shanghai, China) in a ratio of 5 mg to 10.6485 ml
to make 1 mM storage solution. The powder of DDP (Shanghai Macklin
Biochemical, Co., Ltd., Shanghai, China) was dissolved in dimethyl
sulfoxide (DMSO). Aliquots were stored at −80°C.
Main reagents
RPMI-1640 (Gibco, Inc., Grand Island, NY, USA);
fetal bovine serum (FBS; Gibco); typsin and antibodies (Gibco); MTT
Cell Proliferation and Toxicity kit (Biyuntian Inc., Shanghai,
China); TUNEL apoptosis detection kit (Shanghai Yisheng Inc.,
Shanghai, China); primary antibodies of cleaved LC3B, p-mTOR and
GAPDH (Abcam, Inc., Cambridge, MA, USA); primary antibodies of
cleaved caspase-3, Beclin-1, cyclin D1 and p-Akt (Cell Signaling
Technology, Inc., Danvers, MA, USA); goat anti-rabbit antibody
(Invitrogen, Inc., Grand Island, NY, USA); 4% paraformaldehyde
(Sigma-Aldrich, Inc., St. Louis, MO, USA); DAPI (Weiao, Inc.,
Shanghai, China); anti-quenching mounting medium (Sigma-Aldrich);
bovine serum albumin (BSA; Amresco, Inc., Dallas, TX, USA).
Main equipments
Fluorescence microscope (Olympus, Inc., Tokyo,
Japan); light microscope (Olympus); microplate reader (Shanghai
Kehua, Inc., Shanghai, China); table-type refrigerated centrifuge
(USTC Zonkia, Inc., Hefei, China); cell incubator (Thermo Fisher
Scientific, Inc., Waltham, MA, USA); vertical and horizontal
electrophoresis system (Liuyi, Inc., Beijing, China); electric
thermostatic drying oven (Huyue, Inc., Shangyu, China); upright
fluorescence microscope (Nikon, Inc., Tokyo, Japan).
MTT assay
MTT assay was used to detect effects of NVP-BEZ235
alone and combination of NVP-BEZ235 with DDP on proliferation of
HT-29 colorectal adenocarcinoma cells. Cells were placed in 96-well
plates (100 μl/well), and cultured for 24 h before adding
NVP-BEZ235 to make final concentrations of 0, 0.001, 0.01, 0.1, 1
and 3 μM (control group had a concentration of 0 μM; and others
were experimental groups; n=6/concentration). Cells were then
cultured continuously at an incubator with 5% CO2 at
37°C for 24 and 48 h, respectively. Cells were treated with
NVP-BEZ235 (0.1 μM), DDP (100, 300 and 1,000 μM), and NVP-BEZ235
(0.1 μM) combined with DDP (100, 300 and 1,000 μM) respectively,
and cultured for 24 h after treatment (n=5/group). A total of 20 μl
of MTT solutions (5 mg/ml) was added into each well, and continue
to culture at 37°C for 4 h. Supernatant was then discarded and 150
μl DMSO was added into each well, which was shaken for 15 min to
dissolve crystals. Absorbance was detect at 570 nm (A570) using
ELISA microplate reader, and the inhibition rate of cell
proliferation was calculated. Calculation formula: the inhibition
rate of cell proliferation (%) = (1 − absorbance of experimental
groups/absorbance of the control group) × 100%. The probability sum
method was utilized to determine whether there was synergism
combining NVP-BEZ235 and DDP (21). Formula: q = EAB/(EA + EB − EA ×
EB). EAB is the effect (for example, inhibition rate) when drug A
and drug B are combined. EA and EB are effects after drug A and
drug B treatment, respectively. If q is between 0.85 and 1.15, the
effects of drug A and B are additive. Drug A and drug B are
synergic if q>1.15, while antagonistic if q<0.85.
Cell wound scratch assay
HT-29 colorectal adenocarcinoma cells in logarithmic
growth phase were placed on 6-well plates at a concentration of
2×105/ml and cultured in a humid cell incubator with 5%
CO2 at 37°C. Three horizontal lines were drawn at the
back of 6-well plates using marker pens, and 200 μl tips were
utilized to draw horizontal lines at the bottom of plates. The
plates were rinsed with phosphate buffer solution (PBS) 3 times to
eliminate the cells peeled off during dounding. NVP-BEZ235 was
added at concentrations of 0, 0.001, 0.01, 0.1, 1 and 3 μM (control
group had a concentration of 0 μM, and others were experimental
groups; n=12/concentration). Cells were then cultured in humid
incubator with 5% CO2 at 37°C for 24 and 48 h,
respectively. Moreover, cells were treated with NVP-BEZ235 (0.1
μM), DDP (100, 300 and 1,000 μM) and NVP-BEZ235 (0.1 μM) combined
with DDP (100, 300 and 1,000 μM) respectively, and cultured for 24
h after treatment (n=5/group). Images were taken at 0, 24 and 48 h
after adding drug. The distance of migration, and migration index
in each well were calculated using Image-Pro Plus 6.0 software
(Media Cybernetics, Inc., Rockville, MD, USA). Calculation formula:
the inhibition rate of cell migration (%) = (1 − migration distance
of experimental groups/migration distance of the control group) ×
100%.
Expression of LC3B detected by cell
immunofluorescence
HT-29 cells were cultured in 24-well plates with
coverslips at a concentration of 5×104/ml. After
culturing for 24 h, NVP-BEZ235 (1 μM) was added, and wells with no
drug added were the control group. Cells were cultured in an
incubator with 5% CO2 at 37°C for another 48 h.
Coverslips were fixed with 4% paraformaldehyde and blocked with
BSA. Anti-LC3B antibody (1:1,000; Abcam) was incubated at 4°C
overnight. After being washed with PBS, cells were incubated with
goat anti-rabbit secondary antibody conjugated with FITC (1:600;
OriGene Technologies, Inc., Beijing, China) in the dark at 37°C for
30 min. Cells were stained with DAPI for 5 min, and slides were
mounted with anti-quenching mounting medium. Green fluorescence at
520±20 nm and blue fluorescence at 460 nm from DAPI were detected.
DAPI stained both apoptotic and necrotic cells blue.
Western blot analysis
Protein expression of cleaved caspase-3, cleaved
LC3B, p-Akt, p-mTOR, cyclin D1 and Beclin-1 were detected by
western blot analysis. HT-29 cells were treated with NVP-BEZ235 at
different concentrations (0, 1 and 3 μM) for 48 h. HT-29 cells were
then treated with 1 μM NVP-BEZ235 for 0, 4 and 24 h, respectively
(n=4/group). In addition, cells were treated with NVP-BEZ235 (0.1
μM), DDP (100, 300 and 1,000 μM), and NVP-BEZ235 (0.1 μM) combined
with DDP (100, 300 and 1,000 μM) respectively, and cultured for 24
h after treatment. Protein was extracted and separated with 10%
SDS-polyacrylamide gel during eletrophoresis at 120 V. Signal was
transferred to polyvinylidene fluoride (PVDF) membrane at 100 V for
120 min. After blocking with 5% non-fat milk powder for 1 h,
anti-cleaved LC3B antibody (1:1,000; Abcam) were incubated at 4°C
overnight. The membrane was washed with Tris-buffered saline and
Tween-20 (TBST) 3 times, 10 min each time. Membrane was then
incubated with goat anti-rabbit secondary antibody labeled with
horseradish peroxidase (HRP, 1:3,000; Invitrogen) at room
temperature for 1 h. After washing the membrane, and incubating
briefly with electrochemiluminescence (ECL) solution, film was
exposed in a dark room. Experiments were repeated 3 times.
Statistical analysis
Results are demonstrated as mean ± SD. One way
analysis of variance (ANOVA) was utilized to compare differences
among 3 or more groups, followed by Bonferroni post hoc testing for
multiple comparisons. P-values ≤0.05 were regarded significant.
Figures and statistical analysis were made by GraphPad Prism 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA).
Results
NVP-BEZ235 inhibits proliferation of
HT-29 colorectal adenocarcinoma cells
MTT assay was used to evaluate effects of NVP-BEZ235
on proliferation of HT-29 cells. Cells were cultured in 96-well
plates for 24 h before adding NVP-BEZ235 to make final
concentrations of 0, 0.001, 0.01, 0.1, 1 and 3 μM, respectively.
Cells were then cultured at an incubator with 5% CO2 at
37°C for another 24 or 48 h. The inhibition rate of cell
proliferation was calculated by the formula: inhibition rate of
cell proliferation (%) = (1 − absorbance of experimental
groups/absorbance of the control group) × 100%. The inhibitory rate
of proliferation increased as the concentration of NVP-BEZ235
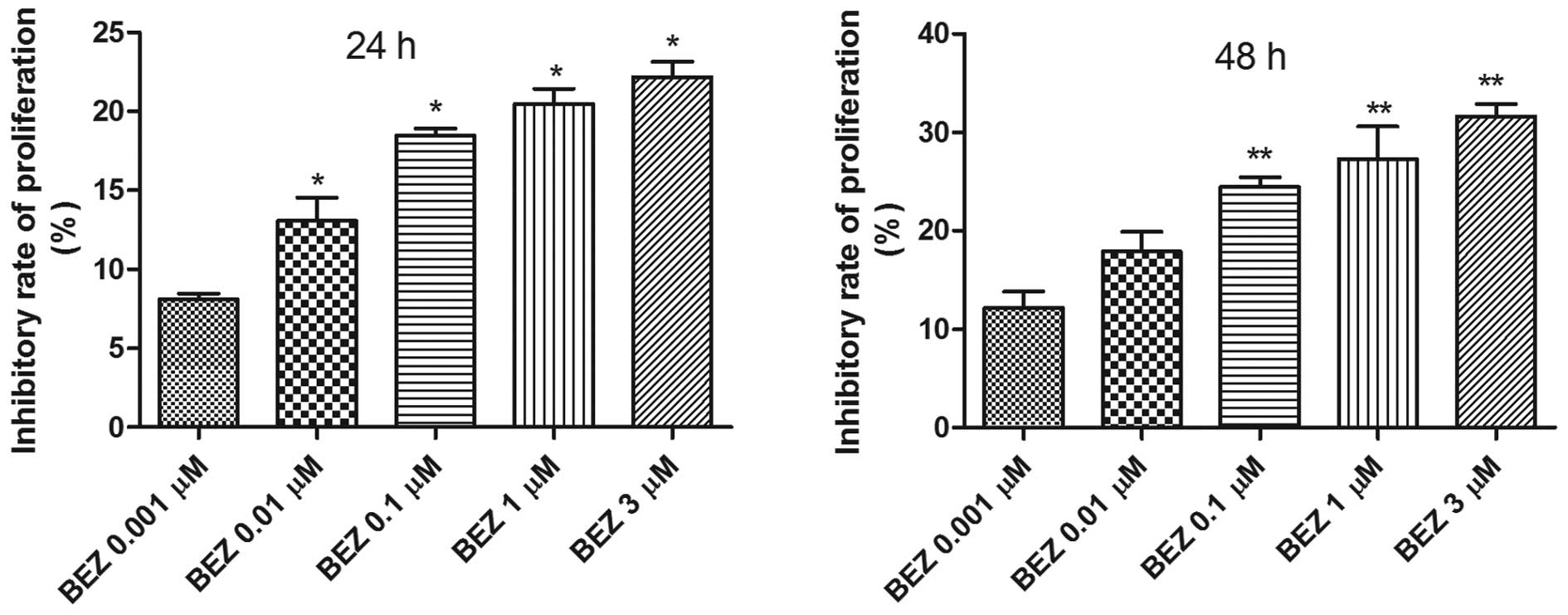
increased at both 24 h (P<0.0001) and 48 h (P=0.0002; Fig. 1). At 24 h after treatment with
NVP-BEZ235, the inhibitory rate of cell proliferation at the
concentration of 0.01 μM increased significantly compared to those
at 0.001 μM (13.1±2.1 vs. 8.1±0.5%, P<0.05). The inhibitory
rates of cell proliferation at the concentration of 0.1, 1 and 3 μM
NVP-BEZ235 were 18.5±0.5, 20.4±1.0 and 22.2±1.4%, and they
increased markedly compared to the inhibitory rate at 0.001 μM
NVP-BEZ235 (P<0.05). Similarly, the inhibitory rate of cell
proliferation at the concentration of 0.001 μM NVP-BEZ235 at 48 h
after treatment was 12.2±2.4%. The rates increased significantly as
the concentration increased to 0.1, 1 and 3 μM (24.5±1.6, 27.3±3.3
and 31.6±2.2%; P<0.05; Fig.
1).
NVP-BEZ235 inhibits the migration of
HT-29 colorectal adenocarcinoma cells
HT-29 cells were placed on 6-well plates and
cultured with 5% CO2 at 37°C until 80% confluence. Three
horizontal lines were drawn at the back of 6-well plates using
marker pens, and 200 μl tips were used to draw horizontal lines at
the bottom of plates. NVP-BEZ235 were added at concentrations of 0,
0.001, 0.01, 0.1, 1 and 3 μM, respectively. Cells were then
cultured for another 24 or 48 h. Images were taken at 0, 24 and 48
h after adding the drug. The distance of migration and migration
index in each well were calculated by Image-Pro Plus 6.0 software.
The distance of cell migration decreased as the concentration of
NVP-BEZ235 increased at both 24 h (P<0.0001) and 48 h
(P<0.0001; Fig. 2). The cell
migration at the concentration of 0.1, 1 and 3 μM NVP-BEZ235 was
64.5±13.9, 63.7±15.6 and 42.7±14.0% of the control group at 24 h,
and they decreased markedly compared to the control group
(P<0.05; Fig. 2). In addition,
the fold change of cell migration at the concentration of 0.001,
0.01, 0.1, 1 and 3 μM NVP-BEZ235 was 80.4±10.7, 80.8±15.7,
30.1±9.0, 31.0±11.8 and 26.2±8.8% of the control group at 48 h, and
they decreased significantly compared to the control group
(P<0.05; Fig. 2).
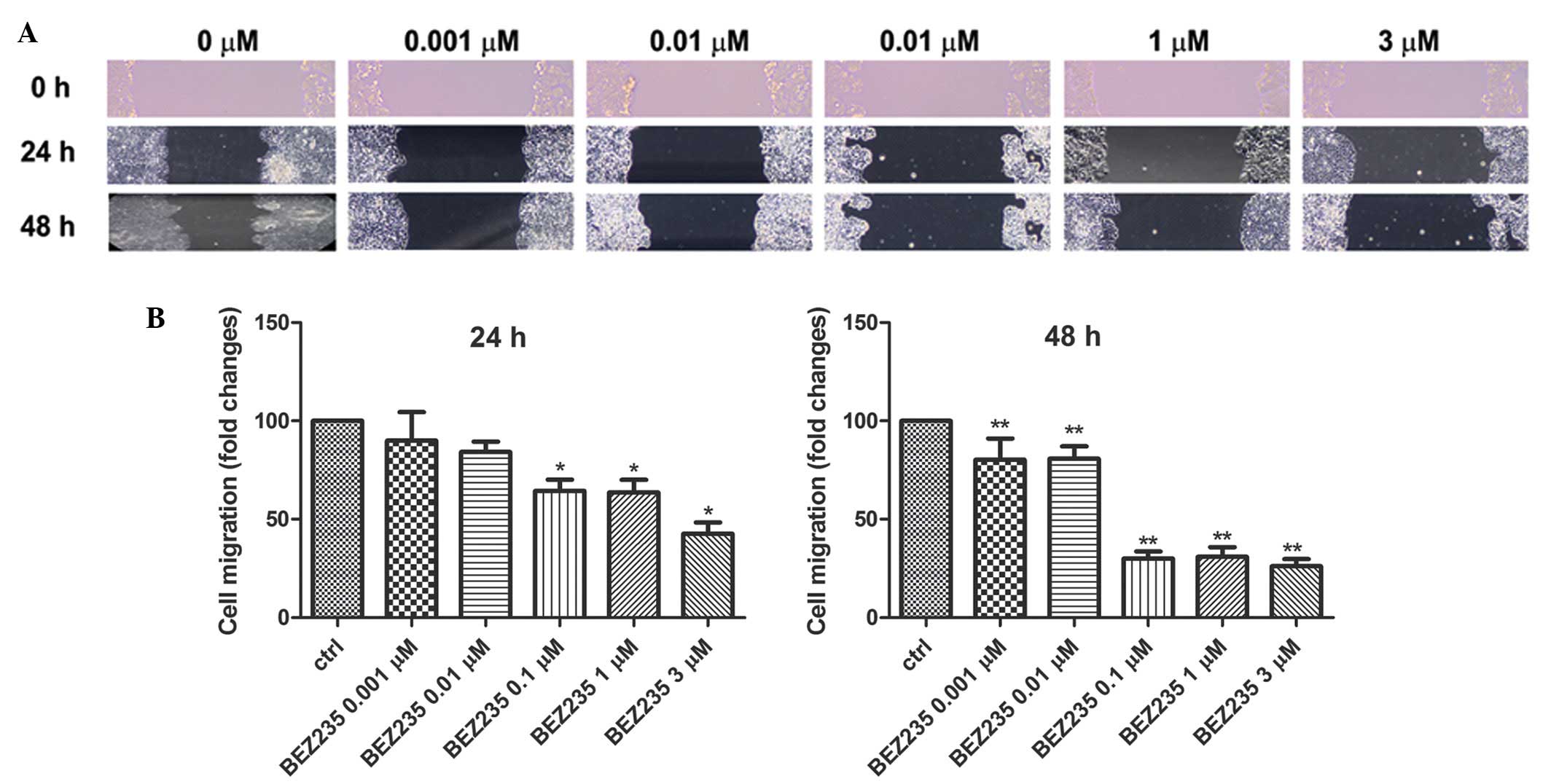 | Figure 2NVP-BEZ235 inhibits the migration of
HT-29 colon adenocarcinoma cells. HT-29 cells were cultured in
6-well plates with 5% CO2 at 37°C until 80% confluence.
Cell wound-scratch assay was performed. NVP-BEZ235 was added to
make final concentrations of 0, 0.001, 0.01, 0.1, 1 and 3 μM,
respectively. Cells were then cultured for another 24 or 48 h. (A)
Representative images are show of the distance of cell migration,
images were taken at 0, 24 and 48 h after adding the drug. (B) The
fold change of cell migration after NVP-BEZ235 treatment. The
distance of migration, and migration index in each well were
calculated by Image-Pro Plus 6.0 software. The distance of cell
migration decreased as the concentration of NVP-BEZ235 increased at
both 24 and 48 h (mean ± SD, n=6/group). *P<0.05;
**P<0.0001. Ctrl, control. |
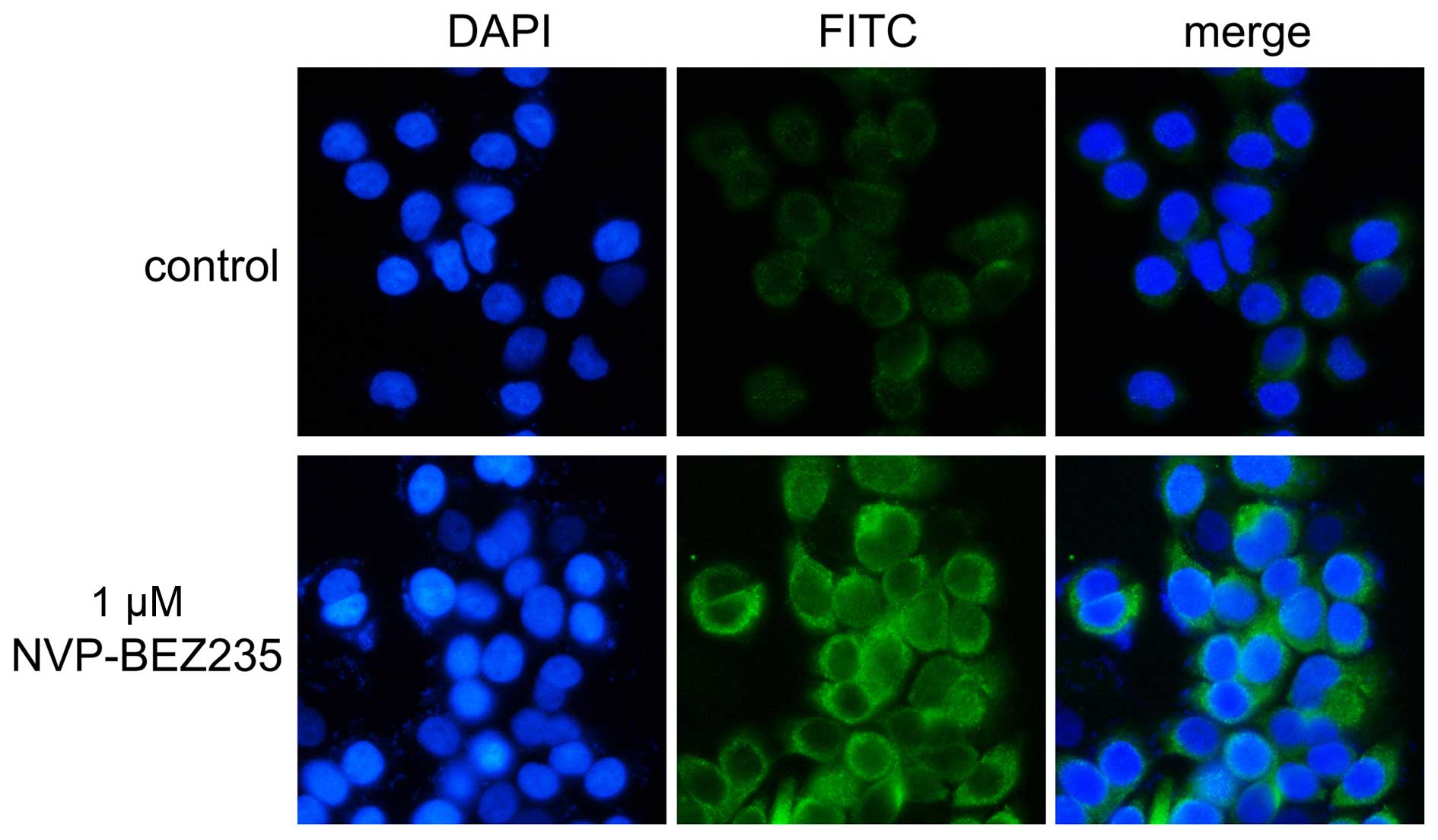
NVP-BEZ235 increases the expression of
LC3B in HT-29 colorectal adenocarcinoma cells
The culture of HT-29 cells in 24-well plates with
coverslips. After culturing for 24 h, NVP-BEZ235 (1 μM) was added,
and wells with no drug added were the control group. Cells were
cultured in an incubator with 5% CO2 at 37°C for another
48 h. Coverslips were fixed, blocked and incubated with anti-LC3B
antibody, goat anti-rabbit secondary antibody and DAPI according to
the instructions in Materials and methods. Green fluorescence
indicates the signaling of LC3B and blue fluorescence signifies the
nuclei. NVP-BEZ235 increased the expression of LC3B in HT-29 cells
as shown in Fig. 3.
Protein expression of p-Akt, p-mTOR,
cyclin D1, cleaved caspase-3, cleaved LC3B and Beclin-1 in HT-29
colorectal adenocarcinoma cells after being treated by
NVP-BEZ235
HT-29 cells were treated with NVP-BEZ235 at
different concentrations (0, 1 and 3 μM) for 48 h, and HT-29 cells
treated with 1 μM NVP-BEZ235 for 0, 4 and 24 h. Protein expression
of p-Akt, p-mTOR, cyclin D1, cleaved caspase-3, cleaved LC3B and
Beclin-1 were detected by western blot analysis. NVP-BEZ235
decreased protein expression of p-Akt (P<0.0001), p-mTOR
(P=0.0105), and cyclin D1 (P=0.0012; Fig. 4), and increased protein expression
of cleaved caspase-3 (P=0.0033), cleaved LC3B (P<0.0001), and
Beclin-1 (P=0.0318; Fig. 4) at 48
h as the concentrations of NVP-BEZ235 increased. In addition,
NVP-BEZ235 decreased protein expression of p-Akt (P=0.0292), p-mTOR
(P=0.0189), and cyclin D1 (P=0.0044; Fig. 5), and increased protein expression
of cleaved caspase-3 (P=0.0026), cleaved LC3B (P=0.0161), and
Beclin-1 (P=0.0141; Fig. 5) as the
incubation time of 1 μM NVP-BEZ235 increased.
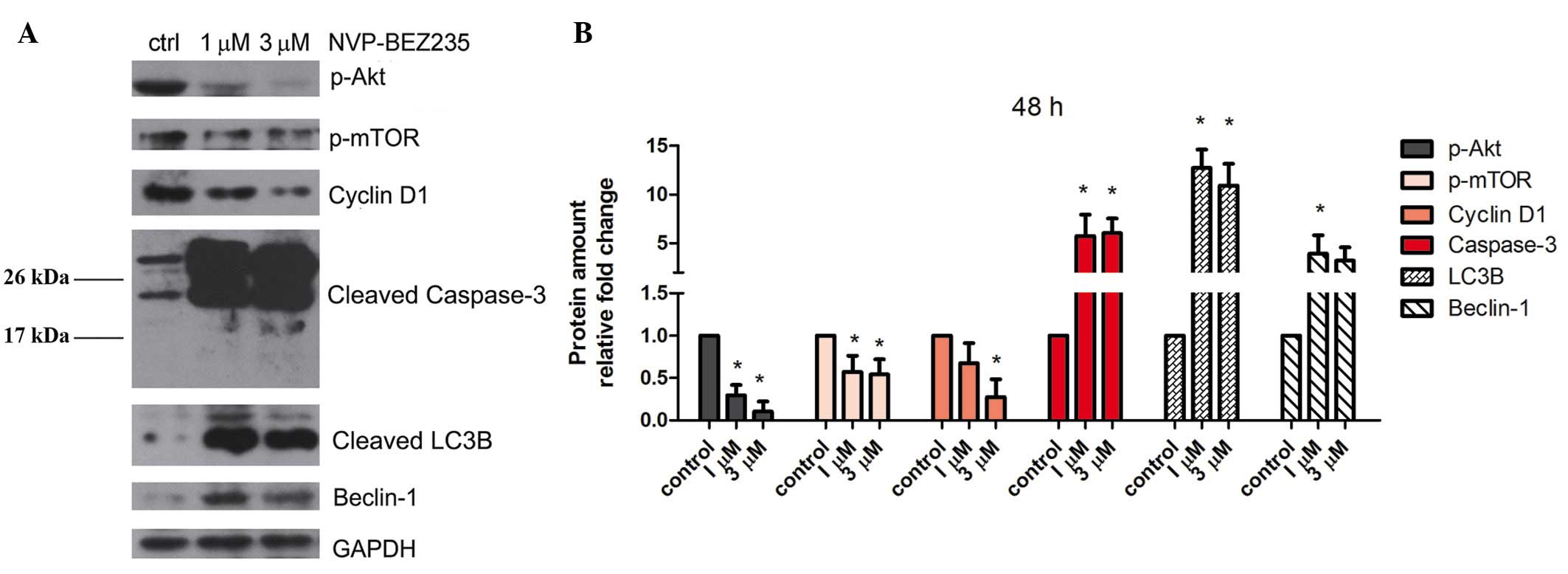 | Figure 4Protein expression of p-Akt, p-mTOR,
cyclin D1, cleaved caspase-3, cleaved LC3B and Beclin-1 in HT-29
colon adenocarcinoma cells after treatment with NVP-BEZ235 at
different concentrations. HT-29 cells were treated with NVP-BEZ235
at different concentrations (0, 1 and 3 μM) for 48 h. (A) Western
blot results demonstrated protein expression of cleaved caspase-3,
cleaved LC3B, p-Akt, p-mTOR, cyclin D1 and Beclin-1 after treatment
with 0, 1 and 3 μM NVP-BEZ235. (B) Quantification of relative fold
change of blot density compared to control group (mean ± SD,
n=4/group). NVP-BEZ235 decreased protein expression of p-Akt,
p-mTOR, and cyclin D1 and increased protein expression of cleaved
caspase-3, cleaved LC3B and Beclin-1 at 48 h as the concentrations
of NVP-BEZ235 increased. *P<0.05. Ctrl, control. |
 | Figure 5Protein expression of p-Akt, p-mTOR,
cyclin D1, cleaved caspase-3, cleaved LC3B and Beclin-1 in HT-29
colon adenocarcinoma cells after treatment with NVP-BEZ235 at
different time-points. HT-29 cells were treated with 1 μM
NVP-BEZ235 for 0, 4 and 24 h. (A) Western blot results demonstrated
protein expression of cleaved caspase-3, cleaved LC3B, p-Akt,
p-mTOR, cyclin D1 and Beclin-1 after treatment with 1 μM NVP-BEZ235
for 0, 4 and 24 h. (B) Quantification of relative fold change of
blot density compared to control group (mean ± SD, n=3/group).
NVP-BEZ235 decreased protein expression of p-Akt, p-mTOR, and
cyclin D1, and increased protein expression of cleaved caspase-3,
cleaved LC3B and Beclin-1 as the incubation time of 1 μM NVP-BEZ235
increased. *P<0.05. Ctrl, control. |
Synergic effects of NVP-BEZ235 and DDP in
cell proliferation, migration and apoptosis
Cells were cultured in 96-well plates for 24 h
before adding NVP-BEZ235 (0.1 μM), DDP (100, 300 and 1,000 μM), and
NVP-BEZ235 (0.1 μM) combined with DDP (100, 300 and 1,000 μM),
respectively. Cells were then cultured for another 24 h. MTT assay
was utilized to detect cell proliferation. We revealed that
NVP-BEZ235 and DDP had synergic effects in inhibition of cell
proliferation when the concentration of DDP was 100 and 300 mM
(Fig. 6 and Table I). Then, we chose 300 mM DDP to
combine with NVP-BEZ235, and detected cell migration and expression
of proteins involved in cell proliferation and apoptosis by cell
wound scratch assay and western blot analysis, respectively.
NVP-BEZ235 and DDP had synergic effects in inhibition of cell
migration (Fig. 7 and Table II). Western blot results
demonstrated that the expression of proteins associated with cell
cycle and proliferation (p-Akt and cyclin D1) decreased in drug
combination group compared to the single treatment groups. The
expression of protein involved in apoptosis (cleaved caspase-3) was
higher in drug combination group compared to the NVP-BEZ235 single
treatment group (Fig. 8).
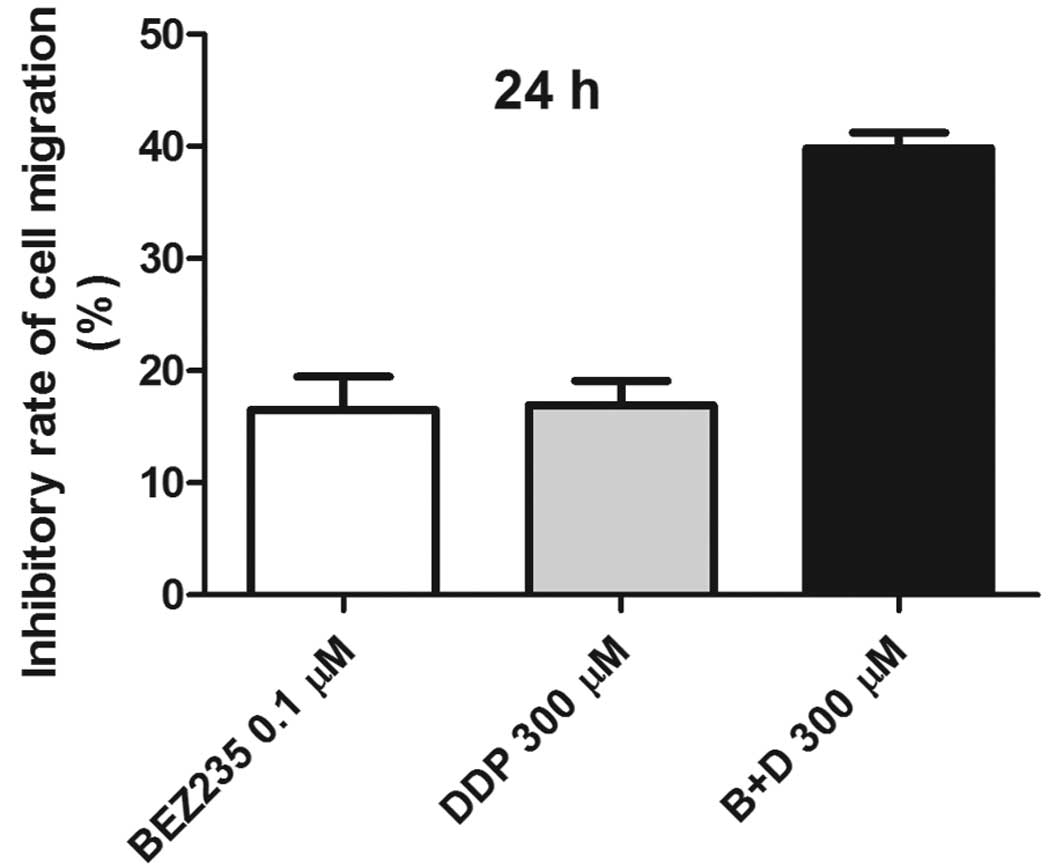
 | Figure 6Synergic effects of NVP-BEZ235 and DDP
on cell proliferation. Cells were treated with NVP-BEZ235 (0.1 μM),
DDP (100, 300 and 1,000 μM), and NVP-BEZ235 (0.1 μM) combined with
DDP (100, 300 and 1,000 μM), respectively, and cultured for 24 h
after treatment. MTT assay was used to detect cell proliferation
and migration. NVP-BEZ235 and DDP had synergic effects in
inhibition of cell proliferation when the concentration of DPP was
100 and 300 mM (Table I). B,
BEZ235: NVP-BEZ235. D, DDP:
cis-diamminedichloroplatinum. |
 | Table ISynergic effects of NVP-BEZ235 and
DDP on cell proliferation at 24 h after treatment. |
Table I
Synergic effects of NVP-BEZ235 and
DDP on cell proliferation at 24 h after treatment.
| BEZ235
0.1 μM | DDP
100 μM | DDP
300 μM | DDP
1,000 μM | B+D
100 μM | B+D
300 μM | B+D
1,000 μM |
|---|
| EA (BEZ235 0.1
μM) | 0.215551 | | | | | | |
| EB (DDP μM) | | 0.079619 | 0.164428 | 0.631103 | | | |
| EAB (B+D) | | | | | 0.358899 | 0.596705 | 0.659406 |
|
q[EAB/(EA+EB−EA*EB)] | | | | | 1.290965a | 1.731903a | 0.927933 |
| Table IISynergic effects of NVP-BEZ235 and
DDP on cell migration at 24 h after treatment. |
Table II
Synergic effects of NVP-BEZ235 and
DDP on cell migration at 24 h after treatment.
| BEZ235
1 μM | DDP
300 μM | B+D
300 μM |
|---|
| EA (BEZ235 1
μM) | 0.165 | | |
| EB (DDP μM) | | 0.169 | |
| EAB (B+D) | | | 0.398 |
|
q[EAB/(EA+EB−EA*EB)] | | | 1.30a |
Discussion
We have demonstrated that NVP-BEZ235 inhibited the
proliferation and migration, and induced apoptosis and autophagy of
HT-29 human colorectal adenocarcinoma cells. NVP-BEZ235 decreased
the protein expression of p-Akt, p-mTOR and cyclin D1, and
increased cleaved caspase-3, cleaved LC3B and Beclin-1 in
colorectal adenocarcinoma cells in a dose- and time-dependent
manner. In addition, NVP-BEZ235 and DDP displayed synergic effects
in cell proliferation, migration and apoptosis at certain
concentrations of DDP.
The PI3K/Akt/mTOR pathway is an important
intracellular signaling pathway that associated with cellular
quiescence, proliferation and cancer. The activation of PI3K and
Akt could activate CREB and mTOR, inhibit p27 and localize FOXO in
the cytoplasm (1,22). In contrast, many factors inhibit
the pathway, such as PTEN, GSK3B and HB9 (1,23,24).
In addition, the PI3K/Akt/mTOR pathway promotes growth and
proliferation of adult stem cells, especially neural stem cells
(1). The pathway is also a
necessary component in neural long-term potentiation (25). Colorectal adenocarcinoma is one of
the most common cancers worldwide with high mortality. The
PI3K/Akt/mTOR pathway induces cell growth and tumor proliferation,
and plays a significant role in colorectal adenocarcinoma.
NVP-BEZ235 is a novel and orally available dual
inhibitor of PI3K/mTOR. It displays antitumor effects in tumors
such as glioma, breast cancer and Burkitt lymphoma (5–7).
NVP-BEZ235 was also reported to be effective in treating
Waldenström's macroglobulinemia (8). The PI3K/Akt pathway plays an
important role in prostate cancer progenitors and NVP-BEZ235
inhibits the cancer stem-like cells of prostate cancer (9,26).
NVP-BEZ235 inhibits the cancer stem cells of glioblastoma (10,27).
Inhibition of the PI3K/Akt/mTOR pathway by NVP-BEZ235 also
suppresses the proliferation of colorectal adenocarcinoma stem
cells with reduced stemness indicated by expressions of CD133 and
Lgr5 (28). NVP-BEZ235 was found
to reduce cell proliferation and induce apoptosis in 3-D cultured
HCT-116 and DLD-1 colorectal adenocarcinoma cells after cytotoxic
therapy with two inhibitors of ubiquitin-proteasome system, MHOK
and bortezomib (11). DDP was the
first member of platinum-containing anticancer drugs. The platinum
complexes react in vivo, bind to and cause DNA crosslinking,
ultimately triggering cell apoptosis (29). However, it is unclear how
NVP-BEZ235 alone affects proliferation, migration, apoptosis and
autophagy in HT-29 human colorectal adenocarcinoma cells, and what
the anticancer effects would be if NVP-BEZ235 and DDP are combined
in treating colorectal cancer.
We revealed that NVP-BEZ235 inhibited proliferation
and migration, and induced apoptosis of human colorectal
adenocarcinoma cells in the present study. This can be explained by
the dose- and time-dependent decrease in protein expression of
p-Akt, p-mTOR and cyclin D1, and increase in cleaved caspase-3
after NVP-BEZ235 treatment as shown in the study. A number of
factors enhance the PI3K/Akt pathway, such as insulin-like growth
factor (IGF-1) (1), insulin
(19), sonic hedgehog homolog
(SHH) and epidermal growth factor (EGF) (20). The overactivation of PI3K/Akt/mTOR
pathway reduces apoptosis and stimulates proliferation, which
involves in the pathogenesis of cancer. Rapamycin, an mTOR
inhibitor, induces apoptosis of human osteosarcoma MG63 cells
(30). Quinazolinone chalcone
derivative inhibits the PI3K/Akt/mTOR signaling pathway and induces
mitochondrial-dependent apoptosis in human colorectal
adenocarcinoma HCT-116 cells (31). We revealed for the first time that
NVP-BEZ235, a dual inhibitor of PI3K and mTOR, inhibited
proliferation and induced apoptosis of HT-29 human colorectal
adenocarcinoma cells.
Akt is an essential component in PI3K/Akt/mTOR
pathway, and is a downstream effector of PI3K. Once correctly
positioned at the membrane via binding of PIP3, Akt can be
phosphorylated by its activating kinases and mTOR complex 2.
Activated Akt can then go on to activate or deactivate its numerous
substrates via its kinase activity, such as mTOR (18). mTOR is a serine/threonine kinase,
and it belongs to PI3K-related kinases family. It regulates
cellular metabolism, growth and proliferation, and is a target for
a number of mTOR inhibitors, including NVP-BEZ235 (19). Cyclin D is a member of cyclin
protein family involving in regulating cell cycle progression.
Cyclin D1 is required for the progression of cell cycle through G1
phase (20). During the G1 phase,
it is rapidly synthesized and accumulates in the nucleus, and is
then degraded when the cell enters S phase. Caspase-3 is a key
member of the cysteine-aspartic acid protease family, and
sequential activation of caspases plays a central role in the
execution-phase of cell apoptosis (16). Therefore, NVP-BEZ235, a dual
inhibitor of PI3K and mTOR, inhibited proliferation and migration,
and induced apoptosis of human colorectal adenocarcinoma cells by
downregulating p-Akt, p-mTOR and cyclin D1 and upregulating cleaved
caspase-3.
We also demonstrated that NVP-BEZ235 increased
protein expression of autophagy markers in human colorectal
adenocarcinoma cells in a dose- and time-dependent manner.
Autophagy is an intracellular degradation process for eliminating
damaged organelles and macromolecules during enhanced cellular
distress. However, the self-cannibalistic function may be
deleterious and lead to cell death (15). The effectiveness of combination of
a selective inhibitor of epidermal growth factor receptor (EGFR)
and a poly(ADP-ribose) polymerase (PARP) inhibitor has been shown
to depend on the enhanced autophagy in ovarian cancer A2780
xenografts (32). Moreover,
cabergoline-mediated prolactinoma shrinkage is associated with the
inhibition of mTOR pathway and the induced autophagy-dependent cell
death (33). Oroxylin A also
induces autophagy in human malignant glioma cells via inhibiting
the activation of Akt and Erk, and the phosphorylation of mTOR and
STAT3 (34). We have shown for the
first time that PI3K/mTOR dual inhibitor NVP-BEZ235 enhanced
autophagy in human colorectal adenocarcinoma cells.
LC3B and Beclin-1 are essential components in the
process of autophagy. LC3B is a subunit of neuronal
microtubule-associated proteins 1A and 1B. LC3B activates extrinsic
apoptosis during cigarette smoking-induced emphysema through
interactions with Caveolin-1 and Fas (35). LC3B-II and Beclin-1 were reported
to be increased during reperfusion in fibrillated mouse hearts
(36). Beclin-1 affects every
major step in autophagic pathways, from autophagosome formation to
the maturation of autophagosome/endosome (15). Many of these effects are mediated
through the activation of specific Beclin 1-binding proteins,
including autophagic inducers and autophagic inhibitors. Cofactors,
such as Bif-1, high mobility group box (HMGB) 1 and survivin,
promote formation of Beclin 1-Vps34-Vps15 core complexes and induce
autophagy, whereas the BH3 domain of Beclin-1 is inhibited by Bcl-2
or Bcl-XL (15). Therefore, the
enhanced autophagy after NVP-BEZ235 treatment may be deleterious to
colorectal cancer, and is another explanation of the decreased
proliferation and migration of colorectal cancer cells after
NVP-BEZ235 treatment.
DPP cross-links DNA and interferes with cell
division by mitosis. Most notable changes in DNA damage are the
1,2-intrastrand cross-links with purine bases. Apoptosis induced by
DDP in human colon cancer cells depends on the mitochondrial
serine-protease Omi/Htra2 (37).
The damaged DNA elicits DNA repair mechanisms, and it activates
apoptosis if repair is impossible. In the present study, we
unveiled that NVP-BEZ235 and DDP had synergic effects in inducing
apoptosis in human colorectal adenocarcinoma cells. Apoptosis was
greatly induced after the drug combination. It is possible that the
greatly increased apoptosis and DNA changes after NVP-BEZ235
treatment exceeds cellular DNA repair mechanisms. The harmful DNA
material and molecules released during NVP-BEZ235-induced apoptosis
may subsequently activate apoptosis of more colorectal
adenocarcinoma cells. This may result in a viscous loop,
aggravating the apoptotic-inducing effects. In addition, we
unveiled that NVP-BEZ235 and DDP had synergic effects in inhibiting
cell proliferation and migration in colorectal adenocarcinoma
cells. It was reported that DPP affected mitosis by interacting
with cellular proteins, particularly HMG domain proteins (38). The synergic effects of NVP-BEZ235
and DDP in inducing apoptosis may also cause synergy in inhibiting
the proliferation and migration of tumor cells. Our research
findings provide rationales in combining NVP-BEZ235 and DDP during
chemotherapy treating colorectal cancer, in order to enhance the
anticancer effects.
In conclusion, we have demonstrated novel data
suggesting that NVP-BEZ235, a dual inhibitor of PI3K/mTOR,
inhibited the proliferation and migration, and induced apoptosis
and autophagy of HT-29 human colorectal adenocarcinoma cells. The
anticancer effects of NVP-BEZ235 are enhanced after being combined
with DDP in treating colorectal adenocarcinoma. Although future
research is needed to shed light on more underlying cellular and
molecular mechanisms, NVP-BEZ235 may serve as a promising therapy
for colorectal cancer.
References
|
1
|
Peltier J, O'Neill A and Schaffer DV:
PI3K/Akt and CREB regulate adult neural hippocampal progenitor
proliferation and differentiation. Dev Neurobiol. 67:1348–1361.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang G, Wang F, Ding W, Wang J, Jing R, Li
H, Wang X, Wang Y, Ju S and Wang H: APRIL induces tumorigenesis and
metastasis of colorectal cancer cells via activation of the
PI3K/Akt pathway. PLoS One. 8:e552982013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
O'Reilly KE, Rojo F, She QB, Solit D,
Mills GB, Smith D, Lane H, Hofmann F, Hicklin DJ, Ludwig DL, et al:
mTOR inhibition induces upstream receptor tyrosine kinase signaling
and activates Akt. Cancer Res. 66:1500–1508. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tabernero J, Rojo F, Calvo E, Burris H,
Judson I, Hazell K, Martinelli E, Ramon y Cajal S, Jones S, Vidal
L, et al: Dose- and schedule-dependent inhibition of the mammalian
target of rapamycin pathway with everolimus: A phase I tumor
pharmacodynamic study in patients with advanced solid tumors. J
Clin Oncol. 26:1603–1610. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu TJ, Koul D, LaFortune T, Tiao N, Shen
RJ, Maira SM, Garcia-Echevrria C and Yung WK: NVP-BEZ235, a novel
dual phosphatidylinositol 3-kinase/mammalian target of rapamycin
inhibitor, elicits multifaceted antitumor activities in human
gliomas. Mol Cancer Ther. 8:2204–2210. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ji Y, Di W, Yang Q, Lu Z, Cai W and Wu J:
Inhibition of autophagy increases proliferation inhibition and
apoptosis induced by the PI3K/mTOR inhibitor NVP-BEZ235 in breast
bancer bells. Clin Lab. 61:1043–1051. 2015.
|
|
7
|
Li C, Xin P, Xiao H, Zheng Y, Huang Y and
Zhu X: The dual PI3K/mTOR inhibitor NVP-BEZ235 inhibits
proliferation and induces apoptosis of Burkitt lymphoma cells.
Cancer Cell Int. 15:652015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sacco A, Roccaro A and Ghobrial IM: Role
of dual PI3/Akt and mTOR inhibition in Waldenstrom's
Macroglobulinemia. Oncotarget. 1:578–582. 2010. View Article : Google Scholar
|
|
9
|
Dubrovska A, Elliott J, Salamone RJ, Kim
S, Aimone LJ, Walker JR, Watson J, Sauveur-Michel M,
Garcia-Echeverria C, Cho CY, et al: Combination therapy targeting
both tumor-initiating and differentiated cell populations in
prostate carcinoma. Clin Cancer Res. 16:5692–5702. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang WJ, Long LM, Yang N, Zhang QQ, Ji WJ,
Zhao JH, Qin ZH, Wang Z, Chen G and Liang ZQ: NVP-BEZ235, a novel
dual PI3K/mTOR inhibitor, enhances the radiosensitivity of human
glioma stem cells in vitro. Acta Pharmacol Sin. 34:681–690. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hernlund E, Olofsson MH, Fayad W, Fryknäs
M, Lesiak-Mieczkowska K, Zhang X, Brnjic S, Schmidt V, D'Arcy P,
Sjöblom T, et al: The phosphoinositide 3-kinase/mammalian target of
rapamycin inhibitor NVP-BEZ235 is effective in inhibiting regrowth
of tumour cells after cytotoxic therapy. Eur J Cancer. 48:396–406.
2012. View Article : Google Scholar
|
|
12
|
Li CH, Cai L, Chen XS, Meng QW and Sui GJ:
DDP-sensitivity-related genes in 10 lung cancer cell lines.
Zhonghua Zhong Liu Za Zhi. 30:418–421. 2008.(In Chinese).
PubMed/NCBI
|
|
13
|
Shen Y, Ren M, Shi Y, Zhang Y and Cai Y:
Octreotide enhances the sensitivity of the SKOV3/DDP ovarian cancer
cell line to cisplatin chemotherapy in vitro. Exp Ther Med.
2:1171–1176. 2011.
|
|
14
|
Weng Y, Wang Y, Shi Y, Zhou W, Wang H and
Wang C: TLR9 expression and its role in chemosensitivity to DDP in
human cervical cancer cells in vitro. J Huazhong Univ Sci Technolog
Med Sci. 31:550–554. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cakir E, Yilmaz A, Demirag F, Oguztuzun S,
Sahin S, Yazici UE and Aydin M: Prognostic significance of
micropapillary pattern in lung adenocarcinoma and expression of
apoptosis-related markers: Caspase-3, bcl-2, and p53. APMIS.
119:574–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cherra SJ III, Kulich SM, Uechi G,
Balasubramani M, Mountzouris J, Day BW and Chu CT: Regulation of
the autophagy protein LC3 by phosphorylation. J Cell Biol.
190:533–539. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sarbassov DD, Guertin DA, Ali SM and
Sabatini DM: Phosphorylation and regulation of Akt/PKB by the
rictor-mTOR complex. Science. 307:1098–1101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pópulo H, Lopes JM and Soares P: The mTOR
signalling pathway in human cancer. Int J Mol Sci. 13:1886–1918.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shimura T, Sasatani M, Kamiya K, Kawai H,
Inaba Y and Kunugita N: Mitochondrial reactive oxygen species
perturb AKT/cyclin D1 cell cycle signaling via oxidative
inactivation of PP2A in lowdose irradiated human fibroblasts.
Oncotarget. 7:3559–3570. 2015.PubMed/NCBI
|
|
21
|
Jin ZJ: About the evaluation of drug
combination. Acta Pharmacol Sin. 25:146–147. 2004.PubMed/NCBI
|
|
22
|
Rafalski VA and Brunet A: Energy
metabolism in adult neural stem cell fate. Prog Neurobiol.
93:182–203. 2011. View Article : Google Scholar
|
|
23
|
Wyatt LA, Filbin MT and Keirstead HS: PTEN
inhibition enhances neurite outgrowth in human embryonic stem
cell-derived neuronal progenitor cells. J Comp Neurol.
522:2741–2755. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ojeda L, Gao J, Hooten KG, Wang E,
Thonhoff JR, Dunn TJ, Gao T and Wu P: Critical role of
PI3K/Akt/GSK3β in moto-neuron specification from human neural stem
cells in response to FGF2 and EGF. PLoS One. 6:e234142011.
View Article : Google Scholar
|
|
25
|
Sui L, Wang J and Li BM: Role of the
phosphoinositide 3-kinase-Akt-mammalian target of the rapamycin
signaling pathway in long-term potentiation and trace fear
conditioning memory in rat medial prefrontal cortex. Learn Mem.
15:762–776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dubrovska A, Kim S, Salamone RJ, Walker
JR, Maira SM, García-Echeverría C, Schultz PG and Reddy VA: The
role of PTEN/Akt/PI3K signaling in the maintenance and viability of
prostate cancer stem-like cell populations. Proc Natl Acad Sci USA.
106:268–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kanai R, Wakimoto H, Martuza RL and Rabkin
SD: A novel oncolytic herpes simplex virus that synergizes with
phosphoinositide 3-kinase/Akt pathway inhibitors to target
glioblastoma stem cells. Clin Cancer Res. 17:3686–3696. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen J, Shao R, Li F, Monteiro M, Liu JP,
Xu ZP and Gu W: PI3K/Akt/mTOR pathway dual inhibitor BEZ235
suppresses the stemness of colon cancer stem cells. Clin Exp
Pharmacol Physiol. 42:1317–1326. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Apps MG, Choi EH and Wheate NJ: The
state-of-play and future of platinum drugs. Endocr Relat Cancer.
22:R219–R233. 2015.PubMed/NCBI
|
|
30
|
Horie R, Nakamura O, Yamagami Y, Mori M,
Nishimura H, Fukuoka N and Yamamoto T: Apoptosis and antitumor
effects induced by the combination of an mTOR inhibitor and an
autophagy inhibitor in human osteosarcoma MG63 cells. Int J Oncol.
48:37–44. 2016.
|
|
31
|
Wani ZA, Guru SK, Rao AV, Sharma S,
Mahajan G, Behl A, Kumar A, Sharma PR, Kamal A, Bhushan S, et al: A
novel quinazolinone chalcone derivative induces mitochondrial
dependent apoptosis and inhibits PI3K/Akt/mTOR signaling pathway in
human colon cancer HCT-116 cells. Food Chem Toxicol. 87:1–11. 2016.
View Article : Google Scholar
|
|
32
|
Sui H, Shi C, Yan Z and Li H: Combination
of erlotinib and a PARP inhibitor inhibits growth of A2780 tumor
xenografts due to increased autophagy. Drug Des Devel Ther.
9:3183–3190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin SJ, Leng ZG, Guo YH, Cai L, Cai Y, Li
N, Shang HB, Le WD, Zhao WG and Wu ZB: Suppression of mTOR pathway
and induction of autophagy-dependent cell death by cabergoline.
Oncotarget. 6:39329–39341. 2015.PubMed/NCBI
|
|
34
|
Zou M, Hu C, You Q, Zhang A, Wang X and
Guo Q: Oroxylin A induces autophagy in human malignant glioma cells
via the mTOR-STAT3-Notch signaling pathway. Mol Carcinog.
54:1363–1375. 2015. View
Article : Google Scholar
|
|
35
|
Chen ZH, Lam HC, Jin Y, Kim HP, Cao J, Lee
SJ, Ifedigbo E, Parameswaran H, Ryter SW and Choi AM: Autophagy
protein microtubule-associated protein 1 light chain-3B (LC3B)
activates extrinsic apoptosis during cigarette smoke-induced
emphysema. Proc Natl Acad Sci USA. 107:18880–18885. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meyer G, Czompa A, Reboul C, Csepanyi E,
Czegledi A, Bak I, Balla G, Balla J, Tosaki A and Lekli I: The
cellular autophagy markers Beclin-1 and LC3B-II are increased
during reperfusion in fibrillated mouse hearts. Curr Pharm Des.
19:6912–6918. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pruefer FG, Lizarraga F, Maldonado V and
Melendez-Zajgla J: Participation of Omi Htra2 serine-protease
activity in the apoptosis induced by cisplatin on SW480 colon
cancer cells. J Chemother. 20:348–354. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cohen SM, Mikata Y, He Q and Lippard SJ:
HMG-domain protein recognition of cisplatin 1,2-intrastrand d(GpG)
cross-links in purine-rich sequence contexts. Biochemistry.
39:11771–11776. 2000. View Article : Google Scholar : PubMed/NCBI
|