Introduction
Several pre-malignant states of prostate tissues
have been previously described to indicate the progression to
prostate adenocarcinoma (prostate cancer). Perhaps the most
prominent histological deviation from normalcy is prostatic
intraepithelial neoplasia (PIN), which can manifest itself as a
low- or high-grade form (1). All
forms of PIN are characterized by the presence of intraluminal
proliferation of the secretory cells of the duct acinar system and
abnormal cytological features, including the ratio of
nuclear-to-cytoplasmic area, the size of nucleoli, and the
chromatin content (2). Another
form of pre-malignancy is accepted to be proliferative inflammatory
atrophy (PIA), which constitutes a possible link between
inflammation and the malignant transformation of prostatic tissues
(3). PIA is mainly recognized in
low-magnification microscopy by a distinct hyperchromatic
appearance of glandular components and variable acinar calibers,
and a marked presence of inflammatory cells (4). Of note, both PIN and PIA are
histologically evident lesions that are identifiable by trained
surgical pathologists. However, it is reasonable to postulate that
cell morphological changes leading to histologically abnormal
appearances of prostate glands are preceded by molecular
alterations that occur in complete absence of any cytological or
histological change. This definition is in complete agreement with
the concept of ‘field effect’ or ‘field cancerization’, two terms
that are used interchangeably in this report to reflect
contemporary research efforts. Originally introduced for renegade
cancer cells outside the margins of squamous oral cell carcinoma
(5), the updated definition
excludes cellular and histological changes and focuses on molecular
aberrations (6). Thus,
‘field-cancerized’ prostate tissues have been recently
characterized by us and others (7–10) by
genetic, epigenetic, and biochemical alterations in structurally
intact epithelial and stromal cells of histologically normal
tissues adjacent to prostate adenocarcinomas.
Along this line, we have recently described four
protein factors of prostate field effect. These include the key
transcription factor early growth response-1 (EGR-1), the lipogenic
enzyme fatty acid synthase (FASN), and the secreted growth factors
platelet-derived growth factor-A (PDGF-A) and macrophage inhibitory
cytokine-1 (MIC-1) (11–13). Our previous reports focused on
emphasizing the similarity of the expressions of these factors
between tumor tissues and their adjacent tissue areas, thereby
supporting the concept of a field effect. Field effect in the
prostate has been recognized to be of potential clinical value
(7–10), which ideally necessitates an
understanding of its underlying causative functional pathways.
Towards this goal, the specific purpose of the present study was to
explore a possible regulatory association between the transcription
factor EGR-1 and the expression of PDGF-A, MIC-1, and FASN. Our
primary focus was the analysis of this potential regulatory network
by mining extensive datasets consisting of expression levels of
EGR-1, PDGF-A, MIC-1, and FASN, in human prostate tissues. Findings
from these analyses were corroborated by ectopic control of EGR-1
and its effect on PDGF-A, MIC-1, and FASN expression in the
non-cancerous RWPE-1 human prostate epithelial cell model.
Accordingly, our data indicate that the key transcription factor
EGR-1 positively regulates PDGF-A and FASN, and negatively
regulates MIC-1. These associations provide novel insight into the
pathways underlying prostate field effect, which may lead to the
development of targeted intervention strategies preventing
progression from pre-malignancy to cancer.
Materials and methods
Tissues
The tissue cohort utilized in the present study
represents a combination of the cohorts reported in our previous
studies on prostate field effect (12,13).
These tissues were collected in agreement with all Federal, State,
and University laws, from consenting patients undergoing
prostatectomy and donating ~100–500 mg of remnant tissue for
molecular analyses. Individual cases of de-identified disease-free
tissue samples were obtained from the Cooperative Human Tissue
Network (CHTN) supported by the National Institutes of Health (NIH;
Vanderbilt University, Nashville, TN, USA). All tissues were
available as formalin-fixed and paraffin-embedded (FFPE) sections
of 5-μm thickness [processed by the Department of Pathology,
University of New Mexico Health Sciences Center (Albuquerque, NM,
USA) or provided by CHTN]. The study was approved by the
Institutional Review Board of the University of New Mexico Health
Sciences Center specifically approved the present study (#05-417).
The combined tissue cohort consisted of 14 adenocarcinomas, 16
tumor-adjacent tissues, and 9 disease-free tissues. Twelve
tumor-adjacent and tumor tissues were matched; for the missing
unmatched tissues, the quality of data was insufficient for
inclusion into the final results. The definition of the term
‘tumor-adjacent’ in our studies refers to tissue resected at a
distance of ~1 cm from the visible tumor margin. The definition of
the term ‘disease-free’ refers to prostate specimens from autopsy
cases from individuals who died due to conditions unrelated to
cancer. All tissues had been histologically reviewed previously by
the surgical pathologist E.G. Fischer (Department of Pathology,
University of New Mexico Health Sciences Center), especially to
exclude the presence of cryptic cancer cells in the tumor-adjacent
prostate tissues (12,13). The mean age of all cases utilized
was 56.1 years with a range of 26–79 years. The cancer specimens
featured Gleason scores from 6 to 9 and pathological tumor node
metastasis (TNM) stages (according to the American Joint Committee
on Cancer; https://cancerstaging.org/Pages/default.aspx) from T2c
to T3b (Table I).
 | Table IDemographics and clinical parameters
of prostate tissues, and number of images analyzed.a |
Table I
Demographics and clinical parameters
of prostate tissues, and number of images analyzed.a
| Prostate
tissues | Age (years) | TNMb | Gleason | No. of images
analyzedc |
|---|
|
|---|
| EGR-1 | MIC-1 | PDGF-A | FASN |
|---|
| Disease-free
(CHTN) | | | | | | | |
| 1 | 26 | Not applicable | Not applicable | 3 | 3 | -- | 3 |
| 2 | 43 | Not applicable | Not applicable | 3 | 3 | -- | 3 |
| 3 | 46 | Not applicable | Not applicable | 3 | -- | 3 | 4 |
| 4 | 79 | Not applicable | Not applicable | 3 | 4 | 2 | -- |
| 5 | 43 | Not applicable | Not applicable | 3 | 3 | 3 | 4 |
| 6 | 55 | Not applicable | Not applicable | 3 | 3 | 2 | 4 |
| 7 | 55 | Not applicable | Not applicable | 3 | -- | -- | 4 |
| 8 | 45 | Not applicable | Not applicable | 3 | -- | -- | 3 |
| 9 | n/ad | Not applicable | Not applicable | 3 | -- | -- | -- |
| Total | | | | 27 | 16 | 10 | 25 |
|
| Prostate
tissues | Age (years) | TNMb | Gleason | Tumor | Adjacent |
|
|
| EGR-1 | MIC-1 | PDGF-A | FASN | EGR-1 | MIC-1 | PDGF-A | FASN |
|
| Tumor and adjacent
(UNMH/CHTN)e | | | | | | | | | | | |
| 1 | 51 | n/ad | 7 (3+4) | -- | 3 | -- | -- | -- | -- | -- | -- |
| 2 | 54 | T3a | 7 (3+4) | -- | 3 | -- | -- | -- | -- | -- | -- |
| 3 (m) | 59 | T3b | 9 (4+5), 6
(3+3) | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| 4 (m) | 63 | T3a | 6 (4+3) | -- | 5 | 2 | -- | -- | 3 | 3 | -- |
| 5 (m) | 69 | T2c | 7 (4+3) | 3 | 3 | 6 | 3 | 6 | 3 | 3 | 3 |
| 6 (m) | 68 | T3b | 8 (5+3) | 3 | 4 | 3 | 3 | 3 | 3 | 3 | 3 |
| 7 (m) | 55 | T2c | 8 (3+5) | 3 | 6 | 9 | -- | 6 | 6 | -- | -- |
| 8 (m) | 57 | T3a | 7 (4+3) | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| 9 (m) | 55 | T2c | 8 (3+5) | 3 | -- | 3 | 3 | 6 | -- | 3 | 9 |
| 10 (m) | 54 | T2–T3 | 6 (3+3) | -- | -- | -- | 3 | 6 | -- | 6 | 6 |
| 11 | 54 | T2c | 6 (3+3) | -- | -- | -- | -- | 9 | -- | 5 | 9 |
| 12 (m) | 64 | T3b | 6 (3+3) | 3 | -- | 4 | -- | 9 | -- | 4 | -- |
| 13 | 62 | T2c | 6 (3+3) | -- | -- | -- | -- | 9 | 9 | 9 | 16 |
| 14 (m) | 62 | T3b | 7 (4+3) | 3 | 4 | 3 | 4 | 6 | 5 | 3 | 9 |
| 15 (m) | 44 | T2c | 6 (3+3) | 3 | -- | 3 | 4 | 5 | -- | -- | 6 |
| 16 | 58 | T2c | 9 (4+5) | -- | -- | -- | -- | 9 | -- | -- | 10 |
| 17 | 69 | T2c | 6 (3+3) | -- | -- | -- | -- | 9 | -- | -- | 12 |
| 18 (m) | 68 | T3a | 7 (3+4) | 3 | 3 | 3 | 4 | 3 | 6 | -- | 4 |
| Total | | | | 30 | 37 | 42 | 30 | 92 | 41 | 45 | 93 |
Quantitative immunofluorescence
The generation of quantitative immunofluorescence
data was reported in our previous studies on prostate field effect
(12,13). These procedures included
deparaffinization, antigen retrieval, and immunostaining using
specific primary antibodies and Alexa Fluor 633-conjugated
secondary antibodies. For reference purposes, we list here the
specific reagents, while the experimental details have been
described (12,13). The primary antibodies were:
anti-EGR-1 mouse monoclonal antibody ab54966 (at 3 μg/ml);
anti-MIC-1 goat polyclonal antibody ab39999 (at 3 μg/ml) (both from
Abcam, Cambridge, MA, USA); anti-PDGF-A rabbit polyclonal antibody
sc-7958 (at 3 μg/ml); and anti-FASN rabbit polyclonal antibody
sc20140 (H-300) (at 8 μg/ml) (both from Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA). The corresponding control antibodies to
ensure target specificity at the same concentrations were: normal
mouse IgG (GC270; EMD Millipore, Billerica, MA, USA), normal rabbit
IgG (10500C), and normal goat IgG (10200) (both from Invitrogen,
Carlsbad, CA, USA). The corresponding secondary antibodies were
Alexa Fluor 633-conjugated goat anti-mouse IgG, Alexa Fluor
633-conjugated goat anti-rabbit IgG, and Alexa Fluor-conjugated
rabbit anti-goat IgG (A21052, A21070, A21086, respectively; all
from Invitrogen). Nuclear counterstaining was performed with
4′,6-diamidino-2-phenylindole (DAPI).
Quantitative assessment of fluorescence was by
spectral image acquisition and linear unmixing modes of confocal
microscopy performed at the University of New Mexico Health
Sciences Center, Fluorescence Microscopy Shared Resource Core
Facility, as described previously by us (12,13).
Of note, control tissue slides with DAPI only, secondary antibody
only, as well as unstained tissue were imaged separately to
generate specific emission spectra for nuclear staining (DAPI; 405
nm excitation, 433 nm emission), Alexa Fluor (633 nm excitation,
490 nm emission), and background autofluorescence (ditto as per
Alexa Fluor), respectively. These spectra were subjected to linear
unmixing, a process that was equally applied to all spectral images
to ensure the validity of inter-tissue comparisons. Consistent with
our previous studies (12,13), quantification was achieved by
digital imaging of the spectrally unmixed confocal images using two
data acquisition modes. i) Whole-image analysis: the total Alexa
Fluor 633 signal was ratio-normalized to the total DAPI signal to
account for the number of cells and the cell density per slide,
which tends to be different between cancerous and non-cancerous
tissues. For EGR-1, the whole-image data acquisition mode was
applied in three settings, i.e., whole-cell (no selection), nuclear
selection, and cytoplasmic selection, according to its ability to
translocate between the two cell compartments (14). ii) Region of interest (ROI)
analysis: three representative ROIs (defined as areas with robust
immunostaining) per slide were chosen and the cumulative signal
specific for Alexa Fluor 633 was determined. The ROI acquisition
mode was applied to all factors according to their typical
expression, i.e., both nuclear and cytoplasmic for EGR-1,
extranuclear for MIC-1 and PDGF-A, and cytoplasmic for FASN. The
size of ROI was identical from image to image (~80 μm2
each) and they were chosen by persons blinded to the nature of the
tissue (Mrs. Virginia Severns, Ms. Fiona Bisoffi, Ms. Suzanne
Jones) to avoid bias (Fig. 1B).
All original red signals were converted to yellow for better
visibility. In total, 488 images with associated quantitative
immunofluorescence data were available for the present analysis
(Table I).
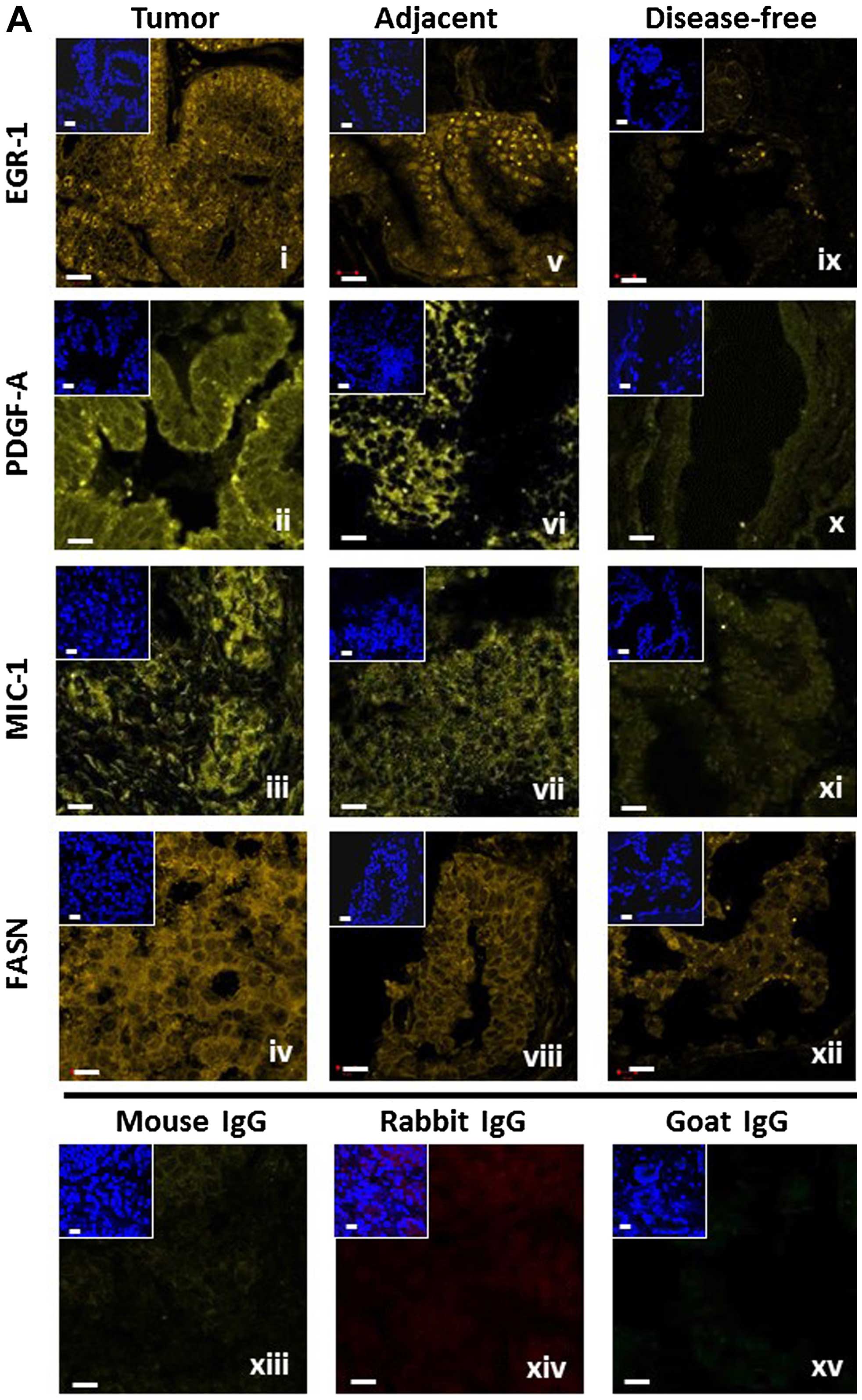 | Figure 1(A) Representative detection of
EGR-1, PDGF-A, MIC-1, and FASN by immunofluorescence in tumor
(panels i–iv), tumor-adjacent (panels v–viii), and disease-free
(panels ix–xii) human prostate tissues. Unspecific IgG of mouse,
rabbit, and goat origin were tested for absence of staining (panels
xiii–xv). Images represent Alexa Fluor 633 immunostaining (yellow
signals); the smaller insets represent corresponding nuclear
staining by DAPI (blue); white bars, 10 μm. (B) Schematic
representation of the whole-image (top) and ROI (bottom)
quantitative acquisition modes for EGR-1 fluorescence intensity.
Whole-image data acquisition includes three different settings as
defined by DAPI staining, whole-cell/no selection (panel i),
nuclear (panel ii), and cytoplasmic (panel iii), as indicated by
the bright blue shading. ROI data acquisition includes nuclear
(panel iv) and extranuclear/cytoplasmic (panel v), as indicated by
the areas designated by the randomly placed yellow rectangle frames
(~80 μm2); white bars, 10 μm. EGR-1, early growth
response-1; PDGF-A, platelet-derived growth factor-A; MIC-1,
macrophage inhibitory cytokine-1; FASN, fatty acid synthase; ROI,
region of interest. |
Computational transcription factor
binding site analysis
Computational searches for a potential transcription
factor binding site were performed using the Tfsitescan software of
the Molecular Informatics Resource for the Analysis of Gene
Expression (MIRAGE) provided by the Institute for Transcriptional
Informatics (IFTI; http://www.ifti.org/cgi-bin/ifti/Tfsitescan.pl).
Genomic sequences for EGR-1, PDGF-A, MIC-1, and FASN were retrieved
from the GRCh38 primary assembly of the gene database available at
the National Center for Biotechnology Information (NCBI; http://www.ncbi.nlm.nih.gov/). The specific reference
sequences and locations were: NC_000005.10, Homo sapiens
chromosome 5, location 138,465,492–138,469,315 for EGR-1;
NC_000019.10, Homo sapiens chromosome 19, location
18,386,158–18,389,176 for MIC-1; NC_000007.14, Homo sapiens
chromosome 7, location 497,258–520,123 for PDGF-A; and
NC_000017.11, Homo sapiens chromosome 17, location
82,078,338–82,098,230 for FASN. The genomic sequences were
subjected to searches for the EGR-1 recognition sequence
[GCG(G/T)GGCG] (15).
Cell culture and transfections
Non-cancerous RWPE-1 human prostate epithelial cells
were purchased from the American Type Culture Collection (Manassas,
VA, USA) and cultured in serum-free keratinocyte basal medium
containing 4,500 mg/l glucose, 0.05 mg/ml bovine pituitary extract
and 5 ng/ml recombinant epidermal growth factor (Invitrogen). Cells
were maintained at 37°C in a humidified 5% CO2
atmosphere. Trypsin-EDTA at 0.25% was used to detach the cells for
splitting and reculturing. pcDNA3.1 control and pcDNA3.1/EGR-1
plasmids were a kind gift of Dr W. Xiao (University of Science and
Technology of China, Hefei, China). pLKO.1 control and pLKO.1/EGR-1
shRNA plasmids were from Sigma (St. Louis, MO, USA). Plasmids were
propagated in E. coli strain JM109 grown in LB broth
containing 100 μg/ml ampicillin and purified using spin column
chromatography (Qiagen, Inc., Valencia, CA, USA). Transfections
were performed with 1 μg plasmid DNA in 24-well plates containing
150,000 cells/well using Lipofectamine 2000 reagent (Invitrogen)
for 48 h. Our transfection protocol yields reproducible
transfection rates of 45±5% for pairs of empty control and
cDNA-carrying plasmids (fluorescence-based assay, not shown). Cells
were snap-frozen in liquid nitrogen to preserve RNA integrity and
stored short-term at −80°C.
Quantitative reverse
transcriptase-polymerase chain reaction (qRT-PCR) and western
blotting
RNA was isolated using spin column chromatography
(Qiagen, Inc.). A total of 1–3 μg of RNA was transcribed to cDNA
using random decamers of the Retroscript™ RT Kit (Ambion/Life
Technologies, Carlsbad, CA, USA). mRNA expression was quantitated
in a CFX Connect Real-Time PCR Detection System from Bio-Rad
(Hercules, CA, USA) using the SYBR-Green PCR Master Mix and
SYBR-Green RT-PCR Reagents Kit (Applied Biosystems/Life
Technologies, Carlsbad, CA, USA) in 25-μl reactions, using 100 ng
of template cDNA and a final primer concentration of 900 nM. The
cycling parameters were 95°C for 5 min followed by 45 cycles of
94°C for 15 sec, and 51–58°C for 1 min. Primers were designed using
Primer Express software (Invitrogen) and synthesized by Integrated
DNA Technologies (Coralville, IA, USA). The following primer
sequences (5′→3′) were used: EGR-1 forward, GAGCAG CCCTACGAGCAC and
reverse, AGCGGCCAGTATAGG TGATG; MIC-1 forward,
CTACAATCCCATGGTGCTCAT and reverse, TCATATGCAGTGGCAGTCTTT; PDGF-A
forward, CGTAGGGAGTGAGGATTCTTT and reverse, GCTTCCTCGATGCTTCTCTT;
FASN forward, AGAACT TGCAGGAGTTCTGGGACA and reverse, TCCGAAGAA
GGAGGCATCAAACCT; TATA-binding protein (TBP) forward,
CACGAACCACGGCACTGATT and reverse, TTT TCTTGCTGCCAGTCTGGAC. qRT-PCR
reactions were performed in triplicate. Relative expression levels
were determined by the ΔΔCt method using TBP as normalization
control after determining that amplification efficiencies were
similar to the ones of the control transcripts.
Protein lysates were generated on ice in lysis
buffer: 25 mM Tris, 8 mM MgCl2, 1 mM DTT, 15% glycerol,
1% Triton X-100, protease inhibitor cocktail (Sigma). Insoluble
cell material was removed by centrifugation of lysates at 13,000
rpm for 10 min at 4°C. The protein concentration was determined by
Bradford assay (Sigma) against a bovine serum albumin (BSA)
standard. Total protein (80 μg) was size-separated by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE),
electro-blotted onto polyvinylidene fluoride (PVDF) membranes,
blocked with 5% milk powder in Tris-buffered saline, and probed
overnight with anti-EGR-1 and anti-β-actin primary antibodies
(sc-189 from Santa Cruz Biotechnology, Inc., Dallas, TX, USA and
A1978 from Sigma, respectively). Detection and chemiluminescent
visualization (Clarity ECL substrate; Bio-Rad) of EGR-1 and β-actin
were performed using host-matched secondary horseradish
peroxidase-conjugated antibodies (Sigma). The quantitative signal
intensity of bands was determined by densitometry using ImageJ
software (https://imagej.nih.gov/ij/).
Statistics
EGR-1, PDGF-A, MIC-1, and FASN expression levels
were represented by signal intensities (sum pixel count per area)
generated by quantitative immunofluorescence analysis (as described
above). Straightforward, yet robust statistical methods were
applied to the datasets using the Microsoft Excel software package
(Microsoft, Redmond, WA, USA). The datasets were inclusive (all
available informative images), for matched cases only, or separated
by the means. These approaches are indicated in the ‘Results’
section.
Correlations between the expressions of EGR-1 and
PDGF-A, MIC-1, and FASN were analyzed by several statistical
methods. To control for small sample size and a distribution with
infinite variance due to tissue heterogeneity (expressed as
coefficient of variation in %; reported in the text of ‘Results’),
the Wilcoxon rank-sum test (as opposed to the Student's t-test) was
used for pairs of datasets (reported in the text of ‘Results’). The
single factor analysis of variance (ANOVA) was applied for
comparisons of multiple datasets with unequal variances.
Statistical significance for the change of ratios of PDGF-A, MIC-1,
or FASN to EGR-1 in tumor-adjacent and tumor tissues as compared to
disease-free tissues was determined by the two-tailed Student's
t-test (statistical significance defined as p≤0.05; Fig. 2A and B). The datasets were further
mined for potential associations between factors by determining the
Pearson's correlation coefficient (r). The significance for these
observations was determined by first calculating the t-value of the
correlation using the equation t = r/SQRT[(1 −
r2)/(n−2)], where r is the correlation coefficient, n is
the number of samples, and n−2 is the degree of freedom. The
t-value was then used to determine the significance of r by the
two-tailed Student's t-distribution (TDIST; statistical
significance defined as p≤0.05; reported in the text, but not
shown). Statistical significance for the change of ratios of
positive to negative Pearson's correlations of PDGF-A, MIC-1, and
FASN to EGR-1 in tumor-adjacent and tumor tissues as compared to
disease-free tissues was determined by the F-test with p≤0.05
considered to be significant (Fig.
3B and D).
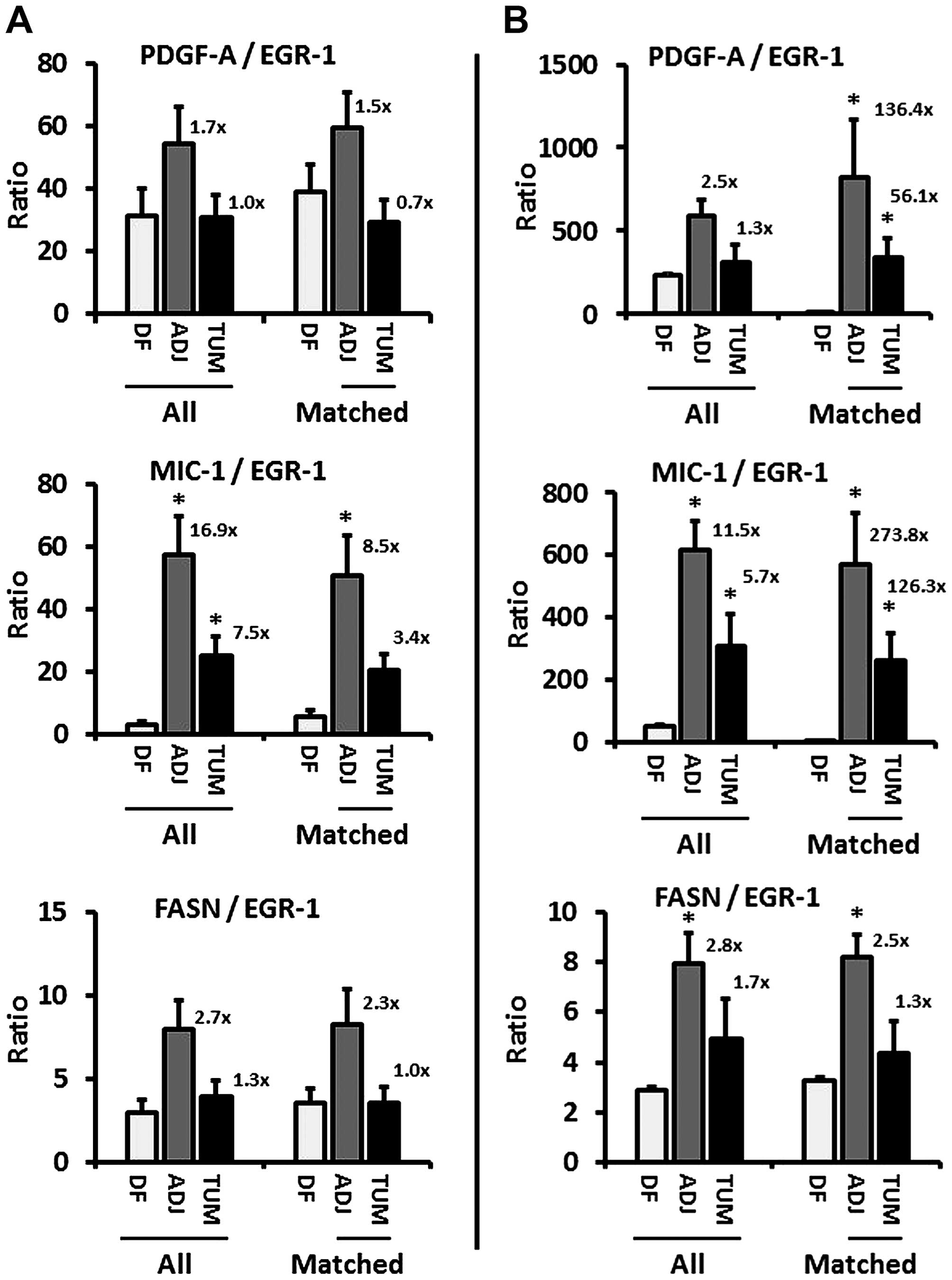 | Figure 2(A and B) Ratios of PDGF-A, MIC-1,
and FASN to EGR-1 expression (combined whole-cell, nuclear,
cytoplasmic) in disease-free (DF), tumor-adjacent (ADJ), and tumor
(TUM) tissues using images from all (left three bars) and matched
only (right three bars) cases, acquired by the whole-image and the
ROI mode, respectively. The bars represent average ratios +
standard errors. The numbers by the bars represent the fold change
in ADJ and TUM compared to DF tissues. *Statistical
significance compared to DF tissues (p≤0.05). PDGF-A,
platelet-derived growth factor-A; MIC-1, macrophage inhibitory
cytokine-1; FASN, fatty acid synthase; EGR-1, early growth
response-1; ROI, region of interest. |
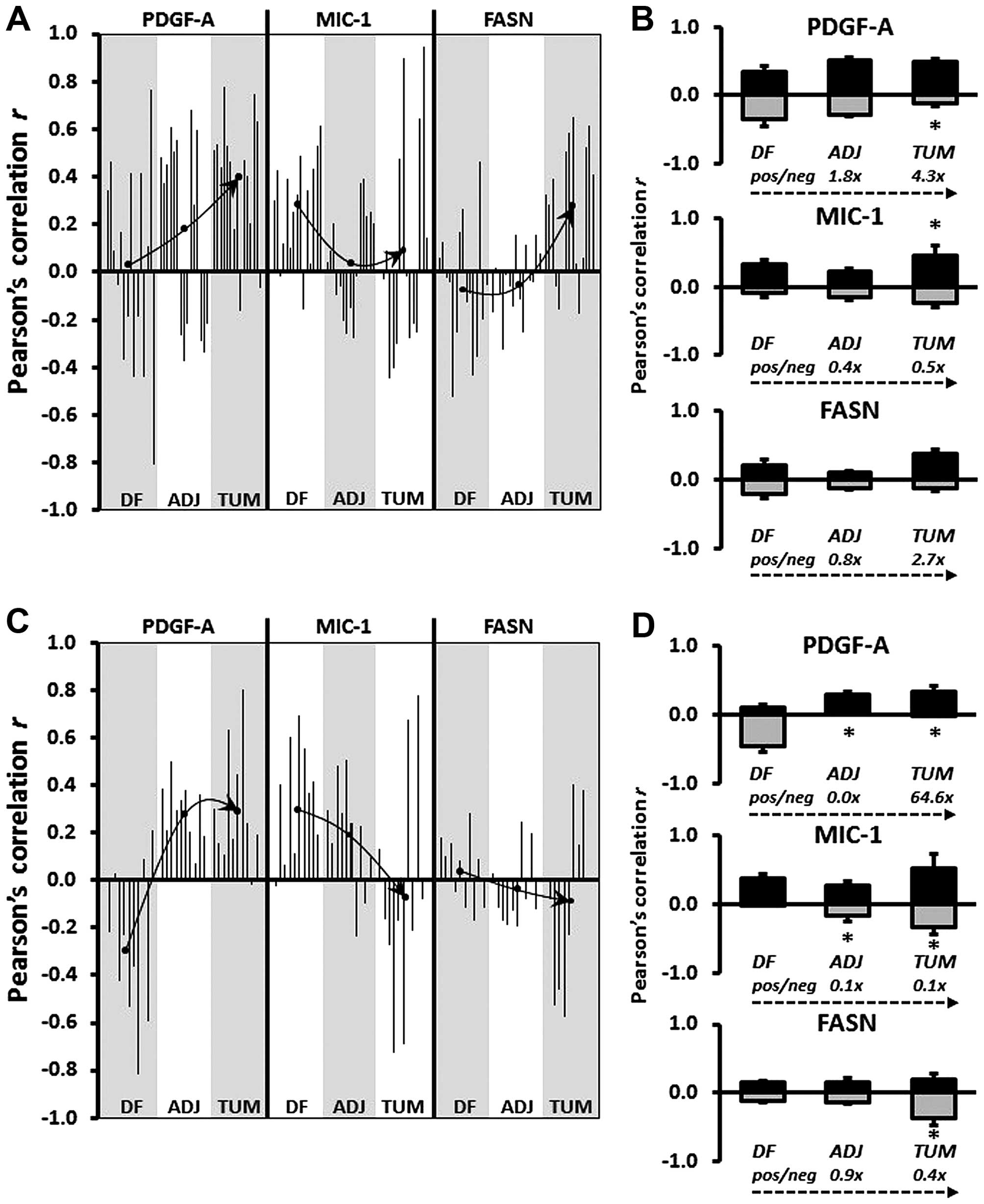 | Figure 3(A and C) Graphical representation of
Pearson's correlation (r) between EGR-1 and PDGF-A, MIC-1, and FASN
using data from digitized images acquired by the whole-image and
the ROI mode, respectively. Within each type of tissue,
disease-free (DF), tumor-adjacent (ADJ), and tumor (TUM),
correlations were determined for all matched, and for EGR-1 above
or below the median with the corresponding median-divided datasets
of PDGF-A, MIC-1, and FASN. (A) Datasets consist of whole-cell,
nuclear, and cytoplasmic EGR-1 measurements (a total of 15
correlations per factor). (B) Datasets consist of nuclear and
cytoplasmic EGR-1 measurements (a total of 12 correlations per
factor). Arrows depict the change of regulation by linking the mean
Pearson's correlations (black dots) in the different types of
tissues. (B and D) Average positive (pos; black bars) and negative
(neg; grey bars) Pearson's correlations between EGR-1 and PDGF-A,
MIC-1, and FASN in DF, ADJ, and TUM tissues acquired by the
whole-image and the ROI mode, respectively. The bars represent
average ratios + standard errors. The numbers represent the fold
change in the ratio of positive/negative r in ADJ and TUM compared
to DF tissues. *Statistical significance compared to DF
tissues (p≤0.05). EGR-1, early growth response-1; PDGF-A,
platelet-derived growth factor-A; MIC-1, macrophage inhibitory
cytokine-1; FASN, fatty acid synthase; ROI, region of interest. |
Results
Immunofluorescence detection of EGR-1,
PDGF-A, MIC-1, and FASN in human prostate tissues
We previously reported on the extent of the
individual expression of EGR-1, PDGF-A, MIC-1, and FASN to support
the concept of field effect in histologically normal prostate
tissues adjacent to histologically overt adenocarcinomas, as
compared to disease-free tissues (12,13).
To begin unraveling the functional pathways of field effect in
prostate tissues, here we analyzed the potential association
between these markers of field effect in human prostate tissues of
different histology. For this analysis, a total of 488 digitized
images from 39 individual human prostate tissue samples was
available for a comprehensive analysis (Table I). The images indicate the specific
detection of EGR-1, PDGF-A, MIC-1, and FASN by immunofluorescence
which was quantified computationally (12,13).
Representative images are shown in Fig. 1A. In general, the expressions of
EGR-1, PDGF-A, MIC-1, and FASN were highest in tumor and lowest or
absent in disease-free tissues (Fig.
1A, panels i–iv and ix–xii, respectively). Furthermore,
tumor-adjacent tissues tended to display elevated expression of all
factors (Fig. 1A, panels v–viii).
The specificity of detection was corroborated by the absent
staining with isotype-specific control antibodies (Fig. 1A, panels xiii–xv).
Quantification and association analyses
of EGR-1, PDGF-A, MIC-1, and FASN expressions in human prostate
tissues
We have previously developed sensitive
quantification methods for signals generated by immunofluorescence
in human prostate tissues [(12,13)
and the ‘Materials and methods’]. These methods include whole-image
and ROI data acquisition modalities for all investigated factors
(in the ‘Materials and methods’). Furthermore, in line with the aim
of this study to be as comprehensive as possible with respect to
associative analyses, EGR-1 expression was measured using three
specific settings for cell compartmentalization: whole-cell, as
well as nuclear and cytoplasmic separately. This is supported by an
elegant study by Mora et al (14) who showed that EGR-1 can shuttle
between these locations depending on cellular type and context.
These different types of data acquisition are shown in Fig. 1B.
While our previous reports compared the level of
expression for EGR-1, PDGF-A, MIC-1, and FASN in disease-free,
tumor-adjacent, and tumor tissues, thereby supporting the concept
of field effect (12,13), the primary objective of the present
study was to explore a potential relationship between these factors
and to determine whether that relationship changes in different
types of tissues. As expected, and typical for human tissue
studies, both the whole-image and the ROI data acquisition modes
resulted in substantial heterogeneity with respect to variation of
expression of all factors in disease-free, tumor-adjacent, and
tumor tissues. The coefficient of variations ranged from 4.7 to
39.0% in the whole-image and from 3.9 to 31.1% in the ROI
measurements.
Quantified expression data were comprehensively
analyzed for similarities, discrepancies, and associations using
straightforward, yet robust statistical methods. Of note, because
of the expected inter- and intra-tissue heterogeneity, the
identification of outliers was not meaningful and we adopted an
inclusive approach in which we did not exclude any data points. In
addition, due to different antibody affinities for their targets,
we determined that comparisons of the mean, variance, and
distribution of expression data between factors would not be good
indicators of a causative regulatory role of EGR-1 for the other
factors. In fact, group analysis by ANOVA indicated that all
expression patterns in all types of tissues were distinct from each
other (p<0.001), and individual comparisons by Wilcoxon rank-sum
test were non-informative with respect to the distinction between
induction and repression (p≤0.05) or coupled expression
(p>0.05). Consequently, we chose to analyze the change of the
ratio of either PDGF-A, MIC-1, or FASN to EGR-1 in disease-free
compared to tumor-adjacent and tumor tissues. Based on our previous
results showing that prostate tissues adjacent to adenocarcinomas
feature a field effect compared to disease-free tissues (12,13),
such a change in ratio would suggest a potential regulatory role of
EGR-1 in agreement with its proven upregulation during
tumorigenesis and cancer progression (16). Accordingly, EGR-1 expression
determined by both the whole-image and ROI acquisition modes in all
available tissues revealed an increase of all factors-to-EGR-1
ratios, up to 2.5-fold for PDGF-A, 16.9-fold for MIC-1, and
2.8-fold for FASN (Fig. 2A and B,
left bar graphs). Similarly, when analyzed for matched adjacent and
tumor tissues only (derived from the same patients, respectively),
the ratio of the other factors to EGR-1 in both acquisition modes
markedly increased, up to 136.4-fold for PDGF-A, 273.8-fold for
MIC-1, and 2.5-fold for FASN (Fig.
2A and B, right bar graphs). While this analysis does not
reveal the direction of regulation (positive or negative), the
changes do indicate a regulatory function of EGR-1 for PDGF-A,
MIC-1, and to a lesser extent for FASN.
The changes in the expression ratio of PDGF-A,
MIC-1, and to some extent FASN, prompted us to refine our
determination of a potential regulatory effect of EGR-1 on these
factors by using Pearson's correlation analysis, which is
independent of differences in antibody affinities for the different
factors. By definition, this approach included tissues from matched
cases only. To refine our analysis, we also separated all
expression data by the median and determined the correlation
between expression levels above and below median values. Similar to
the ratio analysis presented in Fig.
2, we attempted to corroborate possible regulatory effects of
EGR-1 for PDGF-A, MIC-1, and FASN expressions by comparing
Pearson's correlations between different types of tissues, i.e.,
disease-free, tumor-adjacent, and tumor tissues. Fig. 3A and C shows a graphical
representation of all possible correlations between whole-cell,
nuclear, and cytoplasmic EGR-1 and PDGF-A, MIC-1, and FASN
expression in disease-free, tumor-adjacent, and tumor tissues as
acquired by whole-image and ROI acquisition mode, respectively. In
contrast to group analyses by ANOVA or individual comparisons by
Wilcoxon rank-sum test, Pearson's correlation analyses are
indicators of positive vs. negative regulation. The significance
(average p) of the Pearson's correlation coefficients for the
whole-image acquisition mode was 0.16, 0.24, and 0.25 (with 40, 7
and 18% of all coefficients being p≤0.05) for PDGF-A, MIC-1, and
FASN, respectively. For the ROI acquisition mode, the significance
(average p) for the corresponding factors was 0.21, 0.21, and 0.25
(with 17, 23 and 7% of all coefficients being p≤0.05). Visual
inspection of the Pearson's correlation analyses in Fig. 3A and C indicates that EGR-1
positively and negatively regulates PDGF-A and MIC-1, respectively,
while the results for FASN regulation were less clear due to the
contrasting data between the two data acquisition modes. Similar to
the ratio analysis presented in Fig.
2, we attempted to corroborate possible regulatory effects of
EGR-1 for PDGF-A, MIC-1, and FASN expressions by comparing
Pearson's correlations between different types of tissues, i.e.,
disease-free, tumor-adjacent, and tumor tissues. Given the high
tissue heterogeneity, we used an inclusive approach and compared
the average of all positive and negative correlations (r>0 or
<0) for each factor in the three types of tissues. This analysis
showed a progressive positive and negative regulation of PDGF-A (up
to 64.6-fold) and MIC-1 (up to 10-fold), respectively, in
tumor-adjacent and tumor tissues compared to disease-free tissues.
Again, results for FASN were less clear with contrasting results
depending on the data acquisition mode (Fig. 3B and D). These possible regulations
were confirmed by visually linking the means of Pearson's
correlations in the different types of tissues (Fig. 3A and B).
Computational and cell experimental
analysis of EGR-1 regulation of PDGF-A, MIC-1, and FASN
The theoretical potential of the transcription
factor EGR-1 to be a regulator of PDGF-A, MIC-1, and FASN
expression was determined computationally using Tfsitescan software
applied to 1,500 bp upstream and 500 bp downstream of the
transcription initiation site on the genomic sequences of PDGF-A,
MIC-1, and FASN. Thus, a total of 2,000 bp was screened for the
presence of the EGR-1 recognition sequence [GCG(G/T)GGCG] (15). This analysis resulted in the
identification of two, one, and four recognition sequences for
PDGF-A, MIC-1, and FASN, respectively (Fig. 4A). Regulation of PDGF-A, MIC-1, and
FASN expression by EGR-1 was experimentally tested by
overexpression and suppression of EGR-1 in transient transfection
experiments using the non-cancerous RWPE-1 human prostate
epithelial cell model. The immortalized but non-cancerous RWPE-1
cells were chosen because they best represent the tissues analyzed
in this study, which are almost exclusively early-stage malignancy
and tumor-adjacent, i.e., best reflective of field effect.
Transfections with the pcDNA3.1 and the pLKO.1 plasmids typically
resulted in 50–100-fold overexpression and suppression of EGR-1 at
the mRNA level (not shown). Modulation of EGR-1 protein expression
was verified by western blotting and resulted in ~2-fold
overexpression and suppression. Although the regulatory effects on
PDGF-A, MIC-1, and FASN were rather small, transient EGR-1
overexpression upregulated PDGF-A and FASN protein expression (up
to 2-fold) and downregulated MIC-1 protein expression (up to
3-fold), while transient EGR-1 suppression corroborated this effect
by upregulating MIC-1 protein expression (~1.5-fold), while
downregulating PDGF-A and FASN protein expression (up to 2-fold)
(Fig. 4B). These results were
accompanied by similar changes at the mRNA level, as measured by
qRT-PCR. Accordingly, transient EGR-1 overexpression upregulated
PDGF-A and FASN (up to 2-fold) and downregulated MIC-1 (up to
2-fold), while transient EGR-1 suppression corroborated this effect
by downregulating PDGF-A and FASN (up to 2.5- and 5-fold,
respectively) and by upregulating MIC-1 (up to 10-fold) (Fig. 4C). Overall, these results are in
good agreement with the observations made in the tissues.
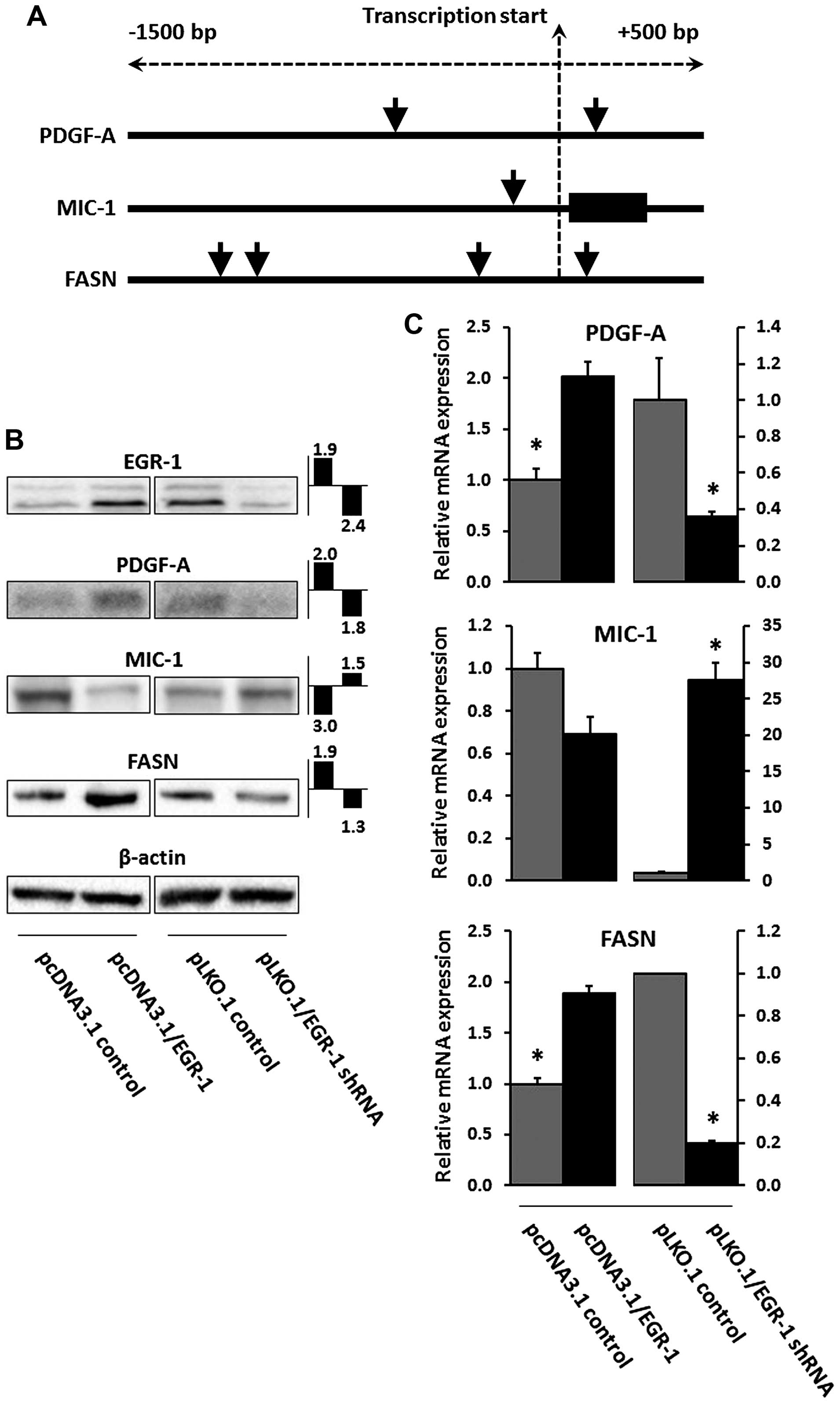 | Figure 4(A) Computational analysis of the
EGR-1 recognition sequence [GCG(G/T)GGCG] in the genomic sequence
1,500 bp upstream and 500 bp downstream of the transcription
initiation site of PDGF-A, MIC-1, and FASN. Black vertical lines
and black rectangular boxes denote genomic sequences and exons,
respectively; vertical arrow heads indicate EGR-1 recognition
sequences. (B) EGR-1, PDGF-A, MIC-1, and FASN protein expression in
RWPE-1 cells transiently transfected with pcDNA3.1/EGR-1 (EGR-1
overexpression) or pLKO.1/EGR-1 shRNA (EGR-1 suppression), and
their empty plasmid controls. Double bands in EGR-1 represent
post-translational modifications (44). The fold change difference compared
to empty plasmid control and determined by densitometry as a ratio
with β-actin signal is indicated in the small bar graphs (left bar,
EGR-1 overexpression; right bar, EGR-1 suppression). (C) Relative
mRNA expression of PDGF-A, MIC-1, and FASN in RWPE-1 cells
transiently transfected with pcDNA3.1/EGR-1 (EGR-1 overexpression)
or pLKO.1/EGR-1 shRNA (EGR-1 suppression), and their empty plasmid
controls. Bars represent averages of triplicates ± standard
deviation; *Statistical significance (p≤0.05) from
pcDNA3.1 and pLKO.1 plasmid vector control, respectively. EGR-1,
early growth response-1; PDGF-A, platelet-derived growth factor-A;
MIC-1, macrophage inhibitory cytokine-1; FASN, fatty acid
synthase. |
Discussion
The importance of field effect, or field
cancerization, in the prostate has been well-recognized as worthy
of being explored in detail for the benefit of developing clinical
applications towards a better clinical management of prostate
cancer (8–10,17).
For example, we have previously argued that prostate field effect
could be used to improve the diagnosis of prostate cancer in
false-negative biopsies (10). The
latter remains an important and continuous challenge in
confirmatory diagnosis of prostate adenocarcinoma that has
clinical, psychological, and financial implications (18–21).
Accordingly, field-cancerized tissue could increase the clinically
informative area that can be analyzed microscopically by a surgical
pathologist if histology could be combined with immunological
techniques. In this scenario, the pathologist would recognize the
presence and location of a lesion even in the absence of its visual
confirmation thereby avoiding false-negative cells, even after
repeated biopsies (22). This
possibility has prompted others to term tissues affected by
field-effect tumor-indicating normal tissue (TINT) (8). Even in the case of a positive
identification of cancer, the extent (number of positive biopsy
cores, % of tissue affected) and the grade (Gleason) may indicate a
low risk for progression and thus eligibility for active
surveillance with frequent testing for serum prostate-specific
antigen (PSA), as opposed to prostatectomy (23). It is conceivable that during active
surveillance, a recognized field effect could be monitored and
queried as an indicator of potential progression (10,24).
This would help mitigate the well-known overtreatment of prostate
cancer with surgery, which albeit performed with curative intent,
may unnecessarily decrease quality of life due to its severe
side-effects (25,26). The latter approach could also be
amenable to the assessment of pre-surgical neo-adjuvant therapeutic
interventions, for which the efficacy could be monitored during
active surveillance by established markers and parameters of field
effect (10,27). A further potential application of
field effect lies in its inclusion in the definition of surgical
margins for focal therapy, which seems to be on the rise as a form
of less invasive therapy and as more refined interventions have
developed (10,28,29).
As such, the presence of a field effect at the margin may be
indicative of elevated risk for progression or of the extent of
tumor multifocality within the prostate (10,30).
Of note, the common assumption underlying the aforementioned
potential applications of prostate field effect is that a field
exists as a consequence of the presence of a lesion. However, it is
also conceivable that field effect precedes tumor formation and
represents a truly pre-malignant status evident at the molecular
level but in absence of any histological change. In fact, the
latter view is widely accepted (8–10,17)
and defines field-cancerized prostate tissues as a temporal record
of tumorigenesis. As such, it is a source for early biomarkers and
potential targets for preventative strategies (8,10).
Pertinent to all applications of field effect is the
knowledge of the molecular markers and pathways that are
characteristic for it. We and others have previously compiled lists
of molecular markers reported in the scientific literature
(7–10), but for most of these factors the
etiology remains unknown. For markers of field effect to be of best
use, either as indicators or as targets, it is important to begin
identifying distinct cellular and molecular events and pathways
that underlie the formation of a field. Towards this goal, in this
report we have established a link between four protein factors of
prostate field effect, which were originally identified
individually or deduced from the literature. We had identified the
key transcription factor EGR-1, the divergent member of the
transforming growth factor-β (TGF-β) MIC-1, and the lipogenic
oncogene FASN as being elevated in prostate tissues 1 cm from the
visible tumor margin (11). While
our original study was microarray-based and thus RNA-specific, we
subsequently confirmed EGR-1, MIC-1, FASN, and PDGF-A protein
upregulation in field-cancerized human prostate tissues (12,13).
EGR-1 is a central regulator of many molecular
pathways and acts divergently according to the cell context
(31). While in other types of
tissues, it may function primarily as a tumor suppressor, it
ultimately assumes, with some ambiguity, a tumor-promoting role in
prostate cancer development and progression (16,32,33).
The role of the secreted factor PDGF-A in prostate cancer is
well-established. It is one of four isoforms that binds as a dimer
to the tyrosine kinase receptors PDGFRα and β. PDGF-A stimulates
growth, survival, and motility of various cell types and when
hyperactivated, promotes prostate cancer development and
progression through paracrine and autocrine actions (34,35).
Equally established in prostate cancer development and progression
is FASN, which has been termed a metabolic oncogene and is the
target of ongoing efforts to develop specific inhibitors of its
lipogenic activity promoting tumor cell proliferation through lipid
biosynthesis and post-translational protein modification (36,37).
The role of MIC-1 is less clear and is reported as both a cancer
promoter and suppressor (38,39).
Originally discovered in macrophages (40), it may promote a pro-tumorigenic
environment when secreted by prostate cancer cells by suppressing
the anticancer activity of immune cells (41).
It is conceivable that the concerted actions of
MIC-1, PDGF-A, and FASN can lead to the formation of molecularly
altered fields through autocrine stimulation of hyperproliferative
cell foci prone to further genetic and biochemical change towards
transformation, which is congruent with the definition of a
pre-malignant field effect. However, the possibility of
cross-regulatory influences of these actions remain unknown. Since
EGR-1 is a pleiotropic transcription factor, we hypothesized that
it could regulate MIC-1, PDGF-A, and FASN. The present study aimed
at testing this possibility through comprehensive association
analyses using quantitative immunofluorescence expression data
generated in human prostate tissues. EGR-1 has been previously
shown to induce many target genes, including PDGF-A in the LAPC4
cell model of prostate cancer after ectopic overexpression of EGR-1
(42). Similarly, MIC-1 seems to
be positively regulated by EGR-1 in the LNCaP prostate cancer cell
model (43). In contrast, there is
a lack of information for a potential regulatory function of EGR-1
for FASN in prostate cells or tissues, although our computational
analysis of genomic DNA up- and downstream of the transcription
initiation site indicates multiple EGR-1 recognition sequences. Our
own ectopic EGR-1 overexpression and suppression data in RWPE-1
cells confirms a positive regulation of PDGF-A, but resulted in a
negative regulation of MIC-1. An obvious reason for this
discrepancy is that RWPE-1 represents a non-cancerous
pre-malignant, as opposed to an advanced cancer cell model, such as
LNCaP (43). At the experimental
level, the use of reporter constructs for MIC-1 activity (43) vs. qRT-PCR using specific primers
may also have contributed to differences in the result. More
importantly however, our in vitro findings are supported by
our extensive in situ association studies in human tissues
which are based on factor correlations and their changes from
disease-free to tumor-adjacent to histologically abnormal tissues,
thereby confirming the presence of a field effect. In fact, using
two data acquisition modes our data show a positive association
between EGR-1 and both PDGF-A and FASN, which in turn support a
positive regulation. In contrast, our results suggest a negative
regulation of MIC-1 by EGR-1, which seemingly contradicts our
observation that both are upregulated in tumor-adjacent and
cancerous prostate tissues when compared to disease-free controls
(12). While the latter justifies
the inclusion of MIC-1 in the present study, this discrepancy
indicates a more complex regulatory network and warrants further
investigations using functional approaches in systems that reflect
the complexity of human tissues.
In summary, three principal conclusions can be drawn
from our findings. First, immunohistochemistry and
immunofluorescence are techniques usually employed towards
qualitative assessment of protein expression and localization in
cells and tissues in a static manner. However, we show here that
using sophisticated quantitation methods, such as spectral image
acquisition, linear unmixing, and digital imaging developed in our
previous reports (12,13), can deliver meaningful indications
of molecular associations in a physiologically relevant in
situ environment, even in the presence of high heterogeneity. A
related issue is the use of ROIs in quantitation. ROIs are often
used to compensate for inequalities of cell composition. Although
our data show good congruency between the whole-image and ROI
approaches for the most part, it also cautions for care with
respect to the number of ROIs and their random and blinded
placement. Second, our study prompts for caution when comparing
molecular association data generated in cell models with data
stemming from tissues. Although it can be argued that tissue
studies are static and compromised by sample heterogeneity, they
can provide meaningful indications of molecular regulations when
coupled with sophisticated data acquisition. Also, tissues are
physiologically relevant, reflect better the complexity of cellular
and molecular pathways influenced by the environment, and can guide
confirmatory studies in cell models. Third, we propose EGR-1 to be
a key regulator of prostate field effect through induction of
pro-proliferative and pro-metabolic (PDGF-A and FASN, respectively)
and suppression of pro-apoptotic (MIC-1) factors. This is supported
in particular by our comparative data between disease-free and
tumor-adjacent tissues (field effect). Admittedly, while the
positive regulation of PDGF-A and FASN by EGR-1 can be easily
acknowledged, its regulatory function for MIC-1 seems less clear
due to its concomitant upregulation in tumor-adjacent tissues
(13). However, it is important to
note that these findings are not in disagreement, as MIC-1
regulation has been discussed to be complex (38,39).
This may be reflected in a complex in situ environment, such
as tissues, where many other factors may also exert their
regulatory effect. Future studies are warranted to test the exact
mechanisms of direct and/or indirect regulation under physiological
conditions, such as in animal models. Because it is widely accepted
that field effect represents a pre-malignant state, such knowledge
may help develop targeted intervention strategies preventing
progression to cancer.
Acknowledgements
We thank the following individuals at the New Mexico
Health Sciences Center, Department of Pathology and Hospital:
Trisha Fleet for procuring prostate tissues through patient
consent; Myra Zucker, Cathy Martinez, and Kari Rigg for skillfully
preparing prostate tissue sections; the surgical pathologist Dr
E.G. Fischer for the histological review of all prostate tissues
utilized in this study. We acknowledge Kerry Wiles from the
CHTN-Western Division at Vanderbilt University Medical Center
(Nashville, TN, USA) for the successful procurement of prostate
tissues and annotated reports. We are grateful to Genevieve
Phillips and Dr Rebecca Lee from the University of New Mexico and
Cancer Center, Fluorescence Microscopy Shared Resource for
excellence assistance and technical input for generating the images
by spectral imaging and linear unmixing. We thank Ms. Virginia
Severns, Ms. Fiona Bisoffi, and Ms. Suzanne Jones for the unbiased
placing of the ROI boxes for signal quantitation in the tissue
images. The departmental offices and staff of the University of New
Mexico, Department of Biochemistry and Molecular Biology, Office of
Medical Student Affairs, and the Schmid College of Science and
Technology, Chapman University are acknowledged for administrative
support. This study was supported by NIH grant RR0164880, NIH grant
R03CA136030-02, Prostate Cancer Research Program grant
W81XWH-15-1-0056 from the Department of Defense (to Dr M. Bisoffi),
University of New Mexico Cancer Center Support grant NIH/NCI
P30CA118110, grants from the Chapman University Office of
Undergraduate Research (to Miss K. Gabriel and Miss E. Frisch), and
a generous gift from Melinda and Edward Subia of Orange County, CA,
USA.
Abbreviations:
|
EGR-1
|
early growth response-1
|
|
FASN
|
fatty acid synthase
|
|
MIC-1
|
macrophage inhibitory cytokine-1
|
|
PDGF-A
|
platelet-derived growth factor-A
|
References
|
1
|
Epstein JI: Mimickers of prostatic
intraepithelial neoplasia. Int J Surg Pathol. 18(Suppl): 142S–148S.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Montironi R, Mazzucchelli R, Algaba F and
Lopez-Beltran A: Morphological identification of the patterns of
prostatic intraepithelial neoplasia and their importance. J Clin
Pathol. 53:655–665. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Marzo AM, Platz EA, Sutcliffe S, Xu J,
Grönberg H, Drake CG, Nakai Y, Isaacs WB and Nelson WG:
Inflammation in prostate carcinogenesis. Nat Rev Cancer. 7:256–269.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
De Marzo AM, Marchi VL, Epstein JI and
Nelson WG: Proliferative inflammatory atrophy of the prostate:
Implications for prostatic carcinogenesis. Am J Pathol.
155:1985–1992. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Slaughter DP, Southwick HW and Smejkal W:
Field cancerization in oral stratified squamous epithelium;
clinical implications of multicentric origin. Cancer. 6:963–968.
1953. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Braakhuis BJ, Tabor MP, Kummer JA, Leemans
CR and Brakenhoff RH: A genetic explanation of Slaughter's concept
of field cancerization: Evidence and clinical implications. Cancer
Res. 63:1727–1730. 2003.PubMed/NCBI
|
|
7
|
Dakubo GD, Jakupciak JP, Birch-Machin MA
and Parr RL: Clinical implications and utility of field
cancerization. Cancer Cell Int. 7:22007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Halin S, Hammarsten P, Adamo H, Wikström P
and Bergh A: Tumor indicating normal tissue could be a new source
of diagnostic and prognostic markers for prostate cancer. Expert
Opin Med Diagn. 5:37–47. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nonn L, Ananthanarayanan V and Gann PH:
Evidence for field cancerization of the prostate. Prostate.
69:1470–1479. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Trujillo KA, Jones AC, Griffith JK and
Bisoffi M: Markers of field cancerization: Proposed clinical
applications in prostate biopsies. Prostate Cancer.
2012:3028942012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haaland CM, Heaphy CM, Butler KS, Fischer
EG, Griffith JK and Bisoffi M: Differential gene expression in
tumor adjacent histologically normal prostatic tissue indicates
field cancerization. Int J Oncol. 35:537–546. 2009.PubMed/NCBI
|
|
12
|
Jones AC, Antillon KS, Jenkins SM, Janos
SN, Overton HN, Shoshan DS, Fischer EG, Trujillo KA and Bisoffi M:
Prostate field cancerization: Deregulated expression of macrophage
inhibitory cytokine 1 (MIC-1) and platelet derived growth factor A
(PDGF-A) in tumor adjacent tissue. PLoS One. 10:e01193142015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jones AC, Trujillo KA, Phillips GK, Fleet
TM, Murton JK, Severns V, Shah SK, Davis MS, Smith AY, Griffith JK,
et al: Early growth response 1 and fatty acid synthase expression
is altered in tumor adjacent prostate tissue and indicates field
cancerization. Prostate. 72:1159–1170. 2012. View Article : Google Scholar
|
|
14
|
Mora GR, Olivier KR, Cheville JC, Mitchell
RF Jr, Lingle WL and Tindall DJ: The cytoskeleton differentially
localizes the early growth response gene-1 protein in cancer and
benign cells of the prostate. Mol Cancer Res. 2:115–128.
2004.PubMed/NCBI
|
|
15
|
Pagel JI and Deindl E: Disease progression
mediated by egr-1 associated signaling in response to oxidative
stress. Int J Mol Sci. 13:13104–13117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gitenay D and Baron VT: Is EGR1 a
potential target for prostate cancer therapy? Future Oncol.
5:993–1003. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Walia G, Pienta KJ, Simons JW and Soule
HR: The 19th annual Prostate Cancer Foundation scientific retreat.
Cancer Res. 73:4988–4991. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Delongchamps NB and Haas GP: Saturation
biopsies for prostate cancer: Current uses and future prospects.
Nat Rev Urol. 6:645–652. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eichler K, Hempel S, Wilby J, Myers L,
Bachmann LM and Kleijnen J: Diagnostic value of systematic biopsy
methods in the investigation of prostate cancer: A systematic
review. J Urol. 175:1605–1612. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Presti JC Jr: Prostate biopsy strategies.
Nat Clin Pract Urol. 4:505–511. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rabbani F, Stroumbakis N, Kava BR, Cookson
MS and Fair WR: Incidence and clinical significance of
false-negative sextant prostate biopsies. J Urol. 159:1247–1250.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Patel AR and Jones JS: Optimal biopsy
strategies for the diagnosis and staging of prostate cancer. Curr
Opin Urol. 19:232–237. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pomerantz M: Active surveillance:
Pathologic and clinical variables associated with outcome. Surg
Pathol Clin. 8:581–585. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mazzucchelli R, Galosi AB, Santoni M,
Lopez-Beltran A, Scarpelli M, Cheng L and Montironi R: Role of the
pathologist in active surveillance for prostate cancer. Anal Quant
Cytopathol Histpathol. 37:65–68. 2015.PubMed/NCBI
|
|
25
|
Bellardita L, Valdagni R, van den Bergh R,
Randsdorp H, Repetto C, Venderbos LD, Lane JA and Korfage IJ: How
does active surveillance for prostate cancer affect quality of
life? A systematic review. Eur Urol. 67:637–645. 2015. View Article : Google Scholar
|
|
26
|
Kwon O and Hong S: Active surveillance and
surgery in localized prostate cancer. Minerva Urol Nefrol.
66:175–187. 2014.PubMed/NCBI
|
|
27
|
Lou DY and Fong L: Neoadjuvant therapy for
localized prostate cancer: Examining mechanism of action and
efficacy within the tumor. Urol Oncol. 34:182–192. 2016. View Article : Google Scholar
|
|
28
|
Lindner U, Lawrentschuk N, Schatloff O,
Trachtenberg J and Lindner A: Evolution from active surveillance to
focal therapy in the management of prostate cancer. Future Oncol.
7:775–787. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marshall S and Taneja S: Focal therapy for
prostate cancer: The current status. Prostate Int. 3:35–41. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Andreoiu M and Cheng L: Multifocal
prostate cancer: Biologic, prognostic, and therapeutic
implications. Hum Pathol. 41:781–793. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pagel JI and Deindl E: Early growth
response 1 - a transcription factor in the crossfire of signal
transduction cascades. Indian J Biochem Biophys. 48:226–235.
2011.PubMed/NCBI
|
|
32
|
Adamson E, de Belle I, Mittal S, Wang Y,
Hayakawa J, Korkmaz K, O'Hagan D, McClelland M and Mercola D: Egr1
signaling in prostate cancer. Cancer Biol Ther. 2:617–622. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Adamson ED and Mercola D: Egr1
transcription factor: Multiple roles in prostate tumor cell growth
and survival. Tumour Biol. 23:93–102. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Heldin CH: Autocrine PDGF stimulation in
malignancies. Ups J Med Sci. 117:83–91. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Heldin CH: Targeting the PDGF signaling
pathway in tumor treatment. Cell Commun Signal. 11:972013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Baron A, Migita T, Tang D and Loda M:
Fatty acid synthase: A metabolic oncogene in prostate cancer? J
Cell Biochem. 91:47–53. 2004. View Article : Google Scholar
|
|
37
|
Zadra G, Photopoulos C and Loda M: The fat
side of prostate cancer. Biochim Biophys Acta. 1831.1518–1532.
2013.
|
|
38
|
Husaini Y, Qiu MR, Lockwood GP, Luo XW,
Shang P, Kuffner T, Tsai VW, Jiang L, Russell PJ, Brown DA, et al:
Macrophage inhibitory cytokine-1 (MIC-1/GDF15) slows cancer
development but increases metastases in TRAMP prostate cancer prone
mice. PLoS One. 7:e438332012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vaňhara P, Hampl A, Kozubík A and Souček
K: Growth/differentiation factor-15: Prostate cancer suppressor or
promoter? Prostate Cancer Prostatic Dis. 15:320–328. 2012.
View Article : Google Scholar
|
|
40
|
Bootcov MR, Bauskin AR, Valenzuela SM,
Moore AG, Bansal M, He XY, Zhang HP, Donnellan M, Mahler S, Pryor
K, et al: MIC-1, a novel macrophage inhibitory cytokine, is a
divergent member of the TGF-beta superfamily. Proc Natl Acad Sci
USA. 94:11514–11519. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Karan D, Holzbeierlein J and Thrasher JB:
Macrophage inhibitory cytokine-1: Possible bridge molecule of
inflammation and prostate cancer. Cancer Res. 69:2–5. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Svaren J, Ehrig T, Abdulkadir SA,
Ehrengruber MU, Watson MA and Milbrandt J: EGR1 target genes in
prostate carcinoma cells identified by microarray analysis. J Biol
Chem. 275:38524–38531. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shim M and Eling TE: Protein kinase
C-dependent regulation of NAG-1/placental bone morphogenic
protein/MIC-1 expression in LNCaP prostate carcinoma cells. J Biol
Chem. 280:18636–18642. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mora GR, Olivier KR, Mitchell RF Jr,
Jenkins RB and Tindall DJ: Regulation of expression of the early
growth response gene-1 (EGR-1) in malignant and benign cells of the
prostate. Prostate. 63:198–207. 2005. View Article : Google Scholar
|














