Introduction
Lung cancer is the most commonly diagnosed and fatal
cancer, which was estimated as 1.8 million new cases and almost 1.6
million deaths occurring worldwide in 2012 (1,2).
Lung adenocarcinoma (LUAD) is major pathologic subtype of lung
cancer in non-smoking males, and in females (smokers or
non-smokers) (3,4). Based on the GLOBOCAN series, the
incidence of LUAD has significantly increased over the past few
decades in Africa, America and Asia (5). LUAD mainly occurs in the peripheral
region of the lungs, with ability to metastasize to the lymph nodes
and other organs. Previous studies have shown that several genetic
abnormalities were associated with the initiation and development
of LUAD (6–10), but the pathogenesis contributing to
biological properties of LUAD remain inconclusive.
Non-coding RNAs (ncRNAs) are a class of RNA
molecules with no protein-coding function, which are widely
expressed in organisms. ncRNAs are divided into several subtypes,
such as microRNAs (miRNAs), small nucleolar RNA (snoRNA), small
nuclear RNA (snRNA), ribosomal RNA (rRNA), transfer ribonucleic
acidtRNA (tRNA) and long non-coding RNA (lncRNA).
Recent studies indicate that ncRNAs play important
biological roles in various biological processes, such as regulate
gene expression, cell proliferation, differentiation and apoptosis
(11–15). lncRNAs, ranging from 200
nucleotides to 100 kb in length, have become an area of increased
research (12,16). lncRNAs may regulate gene expression
at the transcriptional, post-transcriptional and epigenetic levels
(17–19). Because of the stronger tissue
specificity, lncRNAs can be more effective as early diagnosis and
screening of cancer biomarkers (20). Increasing number of studies on
lncRNAs has been performed in various types of cancer, such as
lung, esophageal, gastric, hepatocellular and colorectal cancer
(21–25.) The aberrantly expressed lncRNAs, which regulate
initiation, development, invasion and metastasis of tumors have
been detected in LUAD (10,26).
The ceRNA (competing endogenous RNA) hypothesis was
proposed as a novel regulatory mechanism between non-coding RNA and
coding RNA (27). lncRNA can
interact with the miRNA by miRNA-binding sites (MREs), thereby
regulating gene expression (28).
The complex crosstalk of ceRNA network has been detected in many
different diseases (19). Zhang
et al (29) analyzed the
lncRNA profiles in 372 hepatocellular cell carcinoma (HCC) patients
from The Cancer Genome Atlas (TCGA) (http://cancergenome.nih.gov/) and NCBI GEO Omnibus
(GSE65485), and constructed an lncRNA-miRNA-mRNA network in HCC.
Based on the ceRNA hypothesis, Li et al (30) selected 361 RNA sequencing profiles
of gastric cancer (GC) patients from TCGA, and further analysis of
the associations between these RNA data and related patient
clinical information. Then, the lncRNA-miRNA-mRNA ceRNA network of
GC was constructed.
A recent study showed the differential expression of
lncRNA profile of LUAD using high-throughput microarray analysis
from 33 LUAD patients (31).
However, studies with large scale samples and microarray detection
is still rare, and the relationship between cancer specific lncRNAs
and clinical features are unclear. TCGA database is a public
platform, in which sequencing data of lncRNA, miRNA and mRNA for
LUAD can be downloaded. To improve the reliability and accuracy of
the studies, we detected lncRNAs in LUAD using data sets from TCGA
database. In the present study, the RNA sequencing data of 521 LUAD
samples and 49 adjacent non-tumor lung tissues samples were
collected from TCGA database. This is the first study to
investigate the cancer specific lncRNA from large scale sequencing
database (TCGA) and ceRNA network constructed in LUAD.
Quantificational real-time polymerase chain reaction (qRT-PCR) was
used to verify the bioinformatic analysis results in LUAD tumor
tissues and adjacent non-tumor lung tissues from 53 newly diagnosed
LUAD patients. The present study can help elaborate the function of
lncRNAs through lncRNA-miRNA-mRNA ceRNA network in LUAD.
Materials and methods
Patients and samples
This study was approved by the ethics committee of
the Zhongda Hospital Southeast University. A total of 521 LUAD
cases were obtained from the TCGA database. The exclusion criteria
were set as follows: i) histologic diagnosis is not LUAD; ii)
suffering of other malignancy except LUAD; iii) patients samples
without complete data for analysis; and iv) overall survival (OS)
>5 years. Overall, a total of 465 LUAD patients were included in
this study. lncRNA expression profiles for normal lung tissue
samples were obtained from adjacent non-tumor lung tissues (n=49).
In addition, there are 170 LUAD patients with lymph node metastasis
and 295 LUAD patients with non-lymph node metastasis. According to
the Union for International Cancer Control (UICC), well and
moderately-differentiated LUAD (stage I–II) were 359 patients,
poorly-differentiated LUAD (stage III–IV) were 106 patients. Both
RNA expression data and clinical data, including staging
information and outcome of LUAD patients downloaded from the TCGA
Data Portal, are open-access and publicly available. The present
study meets the publication guidelines provided by TCGA (http://cancergenome.nih.gov/publications/publicationguidelines).
In addition, 53 frozen tissue specimens (tumor
tissues and their paired adjacent non-cancerous tissues) from LUAD
patients collected between 2006 and 2015 were obtained from Nanjing
Chest Hospital Medical School of Southeast University. Tissues were
snap-frozen in RNAlater (Ambion, Austin, TX, USA) and stored in
liquid nitrogen immediately after surgical resection until total
RNA extraction and analysis. Samples were received with a pathology
report and a quality assessment report verifying collection of
tumor and/or adjacent non-tumor lung tissues. Informed consent
forms were obtained from all patients included in the present
study.
RNA sequence data procession and
computational analysis
The LUAD RNA expression data (level 3) of the
corresponding patients were downloaded from the TCGA Data Portal
(March 2016). The RNA sequencing raw reads (lncRNA and mRNA) were
post-processed and normalized by TCGA RNASeqV2 system. Level 3,
normalized miRNA expression data (the calculated expression for all
reads aligning to a particular miRNA per sample) were downloaded
from the TCGA Data Portal performed using Illumina HiSeq and
Illumina GA microRNA sequencing platforms (Illumina, Inc., Hayward,
CA, USA) and quantile normalized before performing analysis. No
further normalizations were applied to the lncRNA, miRNA and mRNA
expression profile data in level 3, because these data were already
normalized by TCGA. To detect the differential expression of
lncRNA, miRNA and mRNA, samples were divided into LUAD patient
tumor tissues vs. adjacent non-tumor lung tissues, lymph node
metastasis of LUAD patients vs. non-lymph node metastasis of LUAD
patients, well and moderately-differentiated LUAD (stages I–II) vs.
poorly-differentiated LUAD (stages III–IV), respectively. For
further analysis, intersection of lncRNA, miRNA and mRNA was
selected. The flow chart for bioinformatics analysis is depicted in
Fig. 1.
Functional enrichment analysis
To understand the underlying biological processes
and pathways of aberrantly expressed intersection genes, we use the
DAVID (Database for Annotation, Visualization, and Integrated
Discovery) Bioinformatics resources (http://david.abcc.ncifcrf.gov/) (32), to conduct functional enrichment
analysis, and we were only interested in GO (Gene Ontology)
biological processes and KEGG (Kyoto Encyclopedia of Genes and
Genomes) pathways at the significant level (P<0.05 and an
enrichment score of >1.5).
Construction of lncRNA-miRNA-mRNA ceRNA
network
The lncRNA-miRNA-mRNA ceRNA network was constructed
based on the relationship among lncRNA, miRNA and mRNA. It is
established that the post-transcriptional regulation of mRNA
transcripts could be bound by single-stranded miRNAs. In the
present study, the lncRNA-miRNA-mRNA ceRNA network, which was based
on the theory that lncRNAs can have a direct interaction by
invoking the miRNA sponge to regulate the activity of mRNAs
(33). To construct ceRNA network,
the human miRNA-mRNA and miRNA-lncRNA interactions could be
downloaded from the starBase V2.0 database (http://starbase.sysu.edu.cn/) (34), then aberrantly expressed
intersection lncRNA, miRNA and mRNA with fold change >2.0 and/or
fold change <0.5 and P<0.05 were retained. The miRanda tools
(http://www.microrna.org/microrna/home.do) were used to
predict the lncRNA-miRNA interactions. Targetscan (http://www.targetscan.org/) and miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/) (35) were performed to predict the mRNAs
targeted by miRNAs. The results could be verified in starBase
database. Furthermore, in the present study the predicted miRNAs
and aberrantly expressed data of TCGA were combined to select the
intersection lncRNAs and mRNAs. To construct ceRNA network, the
miRNAs negatively regulated intersection expression of lncRNAs and
mRNAs were selected, based on the theory of ceRNA. Cytoscape v3.0
(30) was performed to construct
and visualize the lncRNA-miRNA-mRNA ceRNA network. The flow chart
for ceRNA network construction is described in Fig. 2.
The correlations between LUAD specific
lncRNAs and clinical features
According to the bioinformatics analysis and the
ceRNA network, specific lncRNAs were selected. We further analyzed
the clinical features including gender, tumor pathological stage,
TNM staging system, lymph node metastasis and patient outcome
assessment. Subsequently, to identify the specific lncRNAs with
prognostic characteristics, the univariate Cox proportional hazards
regression model was used to analyze the association between the
specific lncRNAs and the LUAD patient survival time.
Total RNA extraction and qRT-PCR
verification
We randomly selected 7 of the specific lncRNAs in
lncRNA-miRNA-mRNA ceRNA network, and validated the reliability and
validity of the results in 53 newly diagnosed LUAD patients using
qRT-PCR.
The total RNA was isolated with TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) according to the manufacture's
protocol, and RNA purity was detected by NanoDrop 2000 spectrometer
(Thermo Fisher Scientific, Waltham, MA, USA). Reverse transcription
reactions using A3500 reverse transcription system kit (Promega,
Madison, WI, USA) was conducted in two steps according to the
manufacturer's protocol. QRT-PCR was carried out to detect the
expression levels of candidate lncRNAs with the StepOnePlus
Real-Time PCR system (Applied Biosystems, Foster City, CA, USA).
QRT-PCR was then performed using A6001 GoTaq qPCR Master Mix
(Promega) according to the manufacturer's protocol. All primers
were purchased from Generay Biotech Co., Ltd. (Shanghai, China).
The Ct-value for each sample was calculated with the ΔΔCt method
(10), and fold change results
were presented as 2−ΔΔCt, where ΔΔCt =
(CtRNAs − CtGAPDH) tumor − (CtRNAs
− CtGAPDH) adjacent non-tumor tissues.
Statistical analysis
All data are presented as mean ± SD, and
statistically compared performing the paired t-test. The
significance level was set as 0.001 as default to control the false
discovery rate (FDR). Values of P<0.05 were considered
statistically significant. The statistical analyses were performed
by the SPSS 21.0.
Results
LUAD specific lncRNAs in LUAD
patients
We identified a total of 1806 lncRNAs from TCGA
database. Analysis of lncRNA expression profiles in LUAD patient
tissues (n=465) compared with adjacent non-tumor lung tissues
(n=49) identified 256 aberrantly expressed lncRNAs (absolute fold
change >2, P<0.01). Of these, 103 lncRNAs were downregulated
and 153 lncRNAs were upregulated. Based on these data, further
analysis was performed between tumor stage (lymph node metastasis)
LUAD patient tissues and adjacent non-tumor lung tissues. A total
of 148 aberrantly expressed lncRNAs were selected between stages
I–II (non-lymph node metastasis) LUAD patient tissues and adjacent
non-tumor lung tissues, 176 aberrantly expressed lncRNAs were
selected between stages III–IV (non-lymph node metastasis) LUAD
patient tissues and adjacent non-tumor lung tissues, 78 aberrantly
expressed lncRNAs were selected between stages I–II (lymph node
metastasis) LUAD patient tissues and adjacent non-tumor lung
tissues, 188 aberrantly expressed lncRNAs were selected between
stages III–IV (lymph node metastasis) LUAD patient tissues and
adjacent non-tumor lung tissues. To further enhance the data
reliability, we selected 41 aberrantly expressed lncRNAs (17
downregulated and 24 upregulated) in the intersection of the above
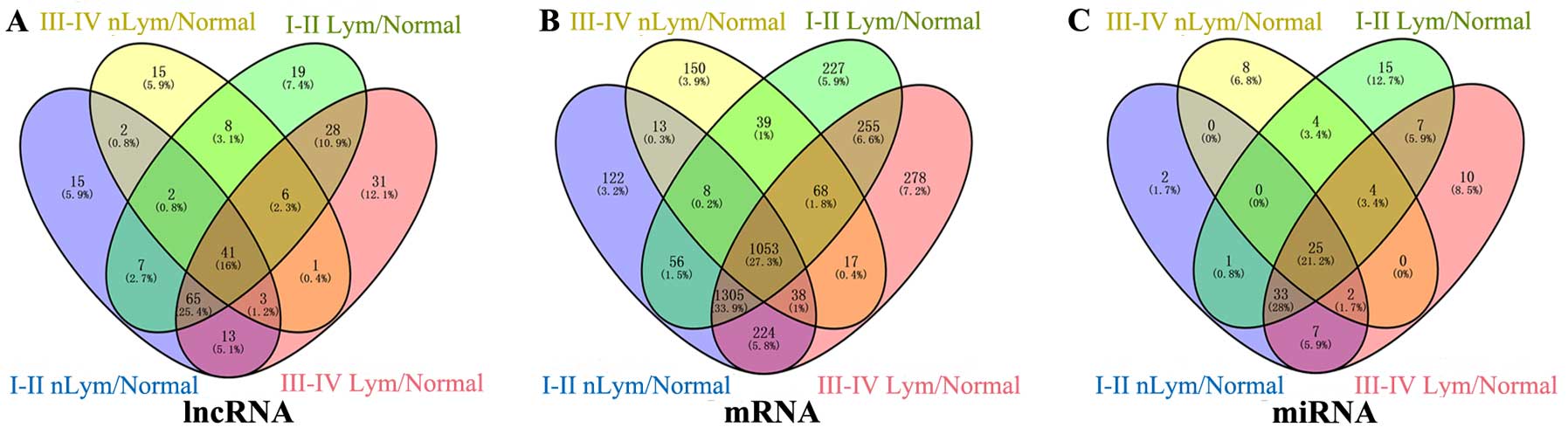
4 groups to build the lncRNA-miRNA-mRNA ceRNA network (Fig. 3A and Table I).
 | Table IAberrantly expressed intersection
lncRNAs between I–II Lym/Normal, I–II nLym/Normal, III–IV
Lym/Normal and III–IV nLym/Normala,b,c. |
Table I
Aberrantly expressed intersection
lncRNAs between I–II Lym/Normal, I–II nLym/Normal, III–IV
Lym/Normal and III–IV nLym/Normala,b,c.
| lncRNAs | Gene ID | Chromosome | Regulation | Fold−change | −Log(p) | −Log(FDRd) |
|---|
| WWC2-AS2 | 152641 | 4 | Downregulation | −3.306±0.220 | 6.757 | 5.243 |
| LINC00472 | 79940 | 6 | Downregulation | −7.018±0.423 | 7.000 | 6.490 |
| AGPAT4-IT1 | 79992 | 6 | Downregulation | −3.200±0.086 | 3.916 | 2.406 |
| CES1P1 | 51716 | 16 | Downregulation | −7.407±2.079 | 6.240 | 4.583 |
| FAM95B1 | 100133036 | 9 | Downregulation | −2.581±0.390 | 3.334 | 1.950 |
| LINC00930 | 100144604 | 15 | Downregulation | −4.124±1.099 | 5.140 | 3.480 |
| RAMP2-AS1 | 100190938 | 17 | Downregulation | −4.819±0.520 | 6.903 | 5.513 |
| BRE-AS1 | 100302650 | 2 | Downregulation | −3.922±1.397 | 4.080 | 2.534 |
| CHIAP2 | 149620 | 1 | Downregulation | −43.011±6.456 | 6.903 | 5.513 |
| LINC01105 | 150622 | 2 | Downregulation | −5.797±1.526 | 3.721 | 2.254 |
| LINC00961 | 158376 | 9 | Downregulation | −6.061±0.683 | 7.000 | 5.774 |
| LINC00908 | 284276 | 18 | Downregulation | −3.810±0.695 | 4.337 | 2.748 |
| BCRP3 | 644165 | 22 | Downregulation | −2.759±0.328 | 4.190 | 2.625 |
| MGC27382 | 149047 | 1 | Downregulation | −11.799±0.396 | 7.000 | 6.490 |
| RPL13AP17 | 399670 | 7 | Downregulation | −47.619±10.808 | 7.000 | 6.490 |
| SIGLEC16 | 400709 | 19 | Downregulation | −3.478±0.259 | 3.413 | 2.023 |
| TPTEP1 | 387590 | 22 | Downregulation | −3.200±1.353 | 4.274 | 2.525 |
| C1orf220 | 400798 | 1 | Upregulation | 4.635±0.649 | 5.078 | 3.409 |
| LINC00319 | 284836 | 21 | Upregulation | 6.750±1.475 | 4.638 | 3.027 |
| LINC00471 | 151477 | 2 | Upregulation | 2.423±0.135 | 3.423 | 2.033 |
| ZNF252P-AS1 | 286103 | 8 | Upregulation | 3.685±0.681 | 3.623 | 2.190 |
| DDX12P | 440081 | 12 | Upregulation | 4.395±1.037 | 4.232 | 2.683 |
| DGCR5 | 26220 | 22 | Upregulation | 9.073±2.637 | 5.049 | 3.384 |
| FER1L4 | 80307 | 20 | Upregulation | 21.753±9.600 | 5.199 | 3.515 |
| ANKRD36BP2 | 645784 | 2 | Upregulation | 5.205±1.082 | 3.520 | 2.109 |
| LOC100132111 | 100132111 | 1 | Upregulation | 3.048±0.504 | 3.361 | 1.984 |
| LOC100133985 | 100133985 | 2 | Upregulation | 2.888±0.318 | 3.471 | 2.069 |
| LINC00896 | 150197 | 22 | Upregulation | 5.338±0.378 | 7.000 | 6.245 |
| LOC285629 | 285629 | 5 | Upregulation | 9.730±5.681 | 7.000 | 6.245 |
| LRRC37A6P | 387646 | 10 | Upregulation | 2.860±0.538 | 3.322 | 1.954 |
| LOC399815 | 399815 | 10 | Upregulation | 8.038±1.458 | 7.000 | 6.245 |
| AFAP1-AS1 | 84740 | 4 | Upregulation | 46.210±7.498 | 7.000 | 6.245 |
| BMS1P20 | 96610 | 22 | Upregulation | 4.468±0.767 | 3.550 | 2.132 |
| PVT1 | 5820 | 8 | Upregulation | 5.335±1.037 | 5.949 | 4.199 |
| RAET1K | 646024 | 6 | Upregulation | 3.463±0.161 | 7.000 | 5.568 |
| SNHG4 | 724102 | 5 | Upregulation | 3.625±0.117 | 5.505 | 3.791 |
| UCA1 | 652995 | 19 | Upregulation | 5.445±0.844 | 6.456 | 4.695 |
| TCAM1P | 146771 | 17 | Upregulation | 4.053±0.409 | 4.205 | 2.661 |
| CECR7 | 100130418 | 22 | Upregulation | 4.055±0.821 | 4.428 | 2.850 |
| ALOX12P2 | 245 | 17 | Upregulation | 3.623±0.261 | 3.787 | 2.321 |
| UNQ6494 | 100129066 | 9 | Upregulation | 2.330±0.881 | 3.549 | 1.940 |
Functional enrichment analysis
In order to understand the functions of aberrantly
expressed genes in this study, the intersection mRNAs were analyzed
by DAVID bioinformatics resources. In total, we identified 18633
mRNAs from TCGA database. A total of 3853 aberrantly expressed
mRNAs were found between LUAD patient tissues and adjacent
non-tumor lung tissues (absolute fold change >2, P<0.01).
Based on these data, aberrantly expressed genes were further
analyzed; 2819 and 1386 aberrantly expressed mRNAs were
respectively identified between stages I–II (non-lymph/lymph node
metastasis) LUAD patient tissues and adjacent non-tumor lung
tissues, 3011 and 3238 aberrantly expressed mRNAs were,
respectively, identified between stages III–IV (non-lymph/lymph
node metastasis) LUAD patient tissues and adjacent non-tumor lung
tissues. Subsequently, 1053 mRNAs were selected from the
intersection of the above 4 groups, and divided into downregulated
and upregulated genes for further functional analysis (Fig. 3B).
It revealed enrichment of 668 GO categories and 87
KEGG categories (P-value of <0.05 and an enrichment score of
>1.5; Figs. 4 and 5). It was found that the top GO
biological process of downregulated and upregulated genes was
G-protein coupled receptor signaling pathway (GO:0007186) and
mitotic cell cycle (GO:0000278) (Fig.
4), respectively. The top 15 KEGG pathways of downregulated and
upregulated genes are also described in Fig. 5. Of these above pathways, PI3K-Akt
signaling pathway, p53 signaling pathway, MAPK signaling pathway
and Wnt signaling pathway are involved in development, invasion and
metastases of cancer. In addition, there were other cancer-related
pathways including pathways in cancer, small cell lung, pancreatic
and bladder cancer and glioma.
Prediction of miRNA targets and ceRNA
network construction
In order to establish lncRNA-miRNA-mRNA ceRNA
network, lncRNAs and mRNAs targeted by miRNAs were identified from
the above data. In the present study, in total we identified 1030
miRNAs from TCGA database. A total of 118 aberrantly expressed
miRNAs were found between LUAD patient tissues and adjacent
non-tumor lung tissues (absolute fold change >2, P<0.05).
Based on comparison of the above 4 groups, 25 miRNAs were selected
from the intersection aberrantly expressed miRNAs (Fig. 3C). Subsequently, we focused on the
targeted relationship between these 25 miRNAs and the above 41
specific intersection lncRNAs. In the lncRNA-miRNA-mRNA ceRNA
network, miRNAs interact with lncRNAs through MREs, starBase v2.0
was performed to detect the potential MREs. The results showed that
22 specific miRNAs interacted with 29 specific lncRNAs (Table II).
| Table IImiRNAs that may target LUAD specific
lncRNAs. |
Table II
miRNAs that may target LUAD specific
lncRNAs.
| lncRNAs | miRNAs |
|---|
| LINC00961 | hsa-miR-127-5p,
hsa-miR-96-5p |
| SIGLEC16 | hsa-miR-127-5p |
| ALOX12P2 | hsa-miR-133a-3p,
hsa-miR-30a-3p, hsa-miR-486-5p |
| BCRP3 |
hsa-miR-135b-5p |
| LINC01105 | hsa-miR-135b-5p,
hsa-miR-143-5p, hsa-miR-182-5p, hsa-miR-708-5p, hsa-miR-96-5p |
| FER1L4 | hsa-miR-139-3p,
hsa-miR-30c-2-3p |
| LINC00319 | hsa-miR-139-3p |
| LINC00896 | hsa-miR-139-3p,
hsa-miR-30c-2-3p |
| DDX12P |
hsa-miR-139-5p, |
| TCAM1P | hsa-miR-139-5p,
hsa-miR-221-5p, hsa-miR-30c-2-3p |
| TPTEP1 | hsa-miR-142-3p,
hsa-miR-708-5p |
| FAM95B1 | hsa-miR-143-5p,
hsa-miR-9-5p |
| LINC00472 | hsa-miR-143-5p,
hsa-miR-96-5p |
| LINC00930 |
hsa-miR-143-5p, |
| RAMP2-AS1 | hsa-miR-143-5p |
| CHIAP2 | hsa-miR-182-5p,
hsa-miR-708-3p, |
| LOC399815 | hsa-miR-30a-3p,
hsa-miR-3614-5p, hsa-miR-378a-3p, hsa-miR-378c |
| LRRC37A6P | hsa-miR-30a-3p,
hsa-miR-338-5p, hsa-miR-378a-3p, hsa-miR-378c |
| BMS1P20 | hsa-miR-30c-2-3p,
hsa-miR-378a-3p, hsa-miR-378c |
| LOC285629 |
hsa-miR-30c-2-3p, |
| MGC27382 | hsa-miR-33a-5p |
| AFAP1-AS1 | hsa-miR-3614-5p,
hsa-miR-378a-3p, hsa-miR-378c, hsa-miR-451a |
| SNHG4 |
hsa-miR-3614-5p |
| UCA1 |
hsa-miR-3614-5p |
| CECR7 | hsa-miR-378a-3p,
hsa-miR-378c |
| RAET1K | hsa-miR-378a-3p,
hsa-miR-378c |
| PVT1 | hsa-miR-378c, |
| UNQ6494 | hsa-miR-378c |
| DGCR5 | hsa-miR-486-5p |
In the next step, mRNA targeted by miRNAs was
performed. Based on the miRNAs described in Table II, TargetScan and miRTarBase were
employed to predict miRNA-targeted mRNA. We identified 19 specific
miRNAs related to the 72 intersection mRNAs (Table III). Some of their targets are
cancer-associated genes, such as BDNF, CACNB4, DKK2, EPAS1, FIGF,
RAD51, THBS2 and SLC2A1.
| Table IIImiRNAs targeting LUAD-specific
mRNAs. |
Table III
miRNAs targeting LUAD-specific
mRNAs.
| miRNAs | mRNAs |
|---|
| hsa-miR-127-5p | ATOH8, PKNOX2,
SYNPO2L |
|
hsa-miR-135b-5p | ADARB2, ARC, NTNG1,
PCYT1B, PGR, PRIMA1, SCN2B |
| hsa-miR-139-3p | SOX11 |
| hsa-miR-139-5p | MEX3A, NRK,
TRIM9 |
| hsa-miR-143-5p | FAT3, TPPP |
| hsa-miR-182-5p | AATK, ANGPTL1,
BDNF, CACNB4, DKK2, EPAS1, FAM107A, FAT3, FIGF, KCNMB2, LDB3, NRN1,
ST6GALNAC3, ZFP36 |
| hsa-miR-210-3p | HIF3A |
| hsa-miR-221-5p | CACNA1E, NAT8L,
RAD51 |
| hsa-miR-30a-3p | KCNE4 |
|
hsa-miR-30c-2-3p | INHBE, MCHR1, MNX1,
NAT8L, PACSIN1 |
| hsa-miR-338-5p | ONECUT2 |
| hsa-miR-33a-5p | AFF3, SLC24A4 |
|
hsa-miR-3607-3p | CDH13, NECAB1,
TNXB |
|
hsa-miR-3614-5p | ELAVL2, EPN3,
FAM155B, GAP43, THBS2, UBE2QL1 |
|
hsa-miR-378a-3p | SLC2A1, SULF1 |
| hsa-miR-378c | SLC2A1, SULF1 |
| hsa-miR-708-3p | AFF2, AFF3, MDGA1,
CNTFR, GPM6A, MASP1, MDGA1 |
| hsa-miR-9-5p | CNTFR, GABRB2,
NTNG1, SCN2B, SHC3, TMTC1, TSPAN18 |
| hsa-miR-96-5p | AHNAK, CACNB4,
COL13A1, FREM1, HBEGF, LDB3, LRRC4, NR4A3, RIMS4, SCNN1G, SLC1A1,
SOX5, TLL1 |
Based on the above data (Tables II and III), the lncRNA-miRNA-mRNA ceRNA
network was constructed. The ceRNA network was plotted with
Cytoscape 3.0. Fig. 6, shows 29
lncRNAs, 24 miRNAs and 72 mRNAs that are involved in the proposed
ceRNA network.
The correlations between LUAD specific
lncRNAs and clinical features
The 29 specific lncRNAs from the ceRNA network were
further analyzed according to clinical features including gender,
race, tumor pathological stage, TNM staging system, lymph node
metastasis, and patient outcome assessment at diagnosis in TCGA
database. There were 21 LUAD-specific lncRNAs, the expression
levels of which were significantly aberrantly expressed in clinical
feature comparisons (P<0.05; Table
IV). Six lncRNAs (BCRP3, ALOX12P2, TCAM1P, UCA1, LOC399815 and
SNHG4) were aberrantly expressed in gender, 2 lncRNAs (LINC00896
and SNHG4) were aberrantly expressed in race, 7 lncRNAs (BCRP3,
LINC01105, AFAP1-AS1, SNHG4, CECR7, RAET1K and PVT1) were
aberrantly expressed in tumor pathological stage, 5 lncRNAs
(LINC01105, TPTEP1, LINC00472, RAMP2-AS1 and DDX12P) were
aberrantly expressed in TNM staging system, 3 lncRNAs (BCRP3,
LINC00472 and LINC00930) were aberrantly expressed in lymph node
metastasis, and 8 lncRNAs (LINC00961, TPTEP1, LINC00472, LINC00930,
MGC27382, FER1L4, LINC00896 and BMS1P20) were aberrantly expressed
in patient outcome assessment.
| Table IVThe correlations between LUAD
specific lncRNAs from ceRNA network and clinical features. |
Table IV
The correlations between LUAD
specific lncRNAs from ceRNA network and clinical features.
| Comparisons | Downregulated | Upregulated |
|---|
| Gender (female vs.
male) | BCRP3 | ALOX12P2, TCAM1P,
UCA1, LOC399815, SNHG4 |
| Race (White vs.
Asian) | | LINC00896,
SNHG4 |
| Tumor pathological
stage (I–II vs. III–IV) | BCRP3,
LINC01105 | AFAP1-AS1, SNHG4,
CECR7, RAET1K, PVT1 |
| TNM staging system
(T1 + T2 vs. T3 + T4) | LINC01105, TPTEP1,
LINC00472, RAMP2-AS1 | DDX12P |
| Lymph node
metastasis (no vs. yes) | BCRP3, LINC00472,
LINC00930 | |
| Patient outcome
assessment (dead vs. alive) | LINC00961, TPTEP1,
LINC00472, LINC00930, MGC27382 | FER1L4, LINC00896,
BMS1P20 |
In order to identify the 29 specific lncRNAs with
prognostic characteristics, these lncRNAs in the TCGA database were
profiled with the univariate Cox proportional hazards regression
model and 5 specific lncRNAs were found significantly associated
with OS (log-rank P<0.05). While the remaining 5 (BCRP3,
LINC00472, CHIAP2, BMS1P20 and UNQ6494) were positively correlated
with OS (Fig. 7).
qRT-PCR verification
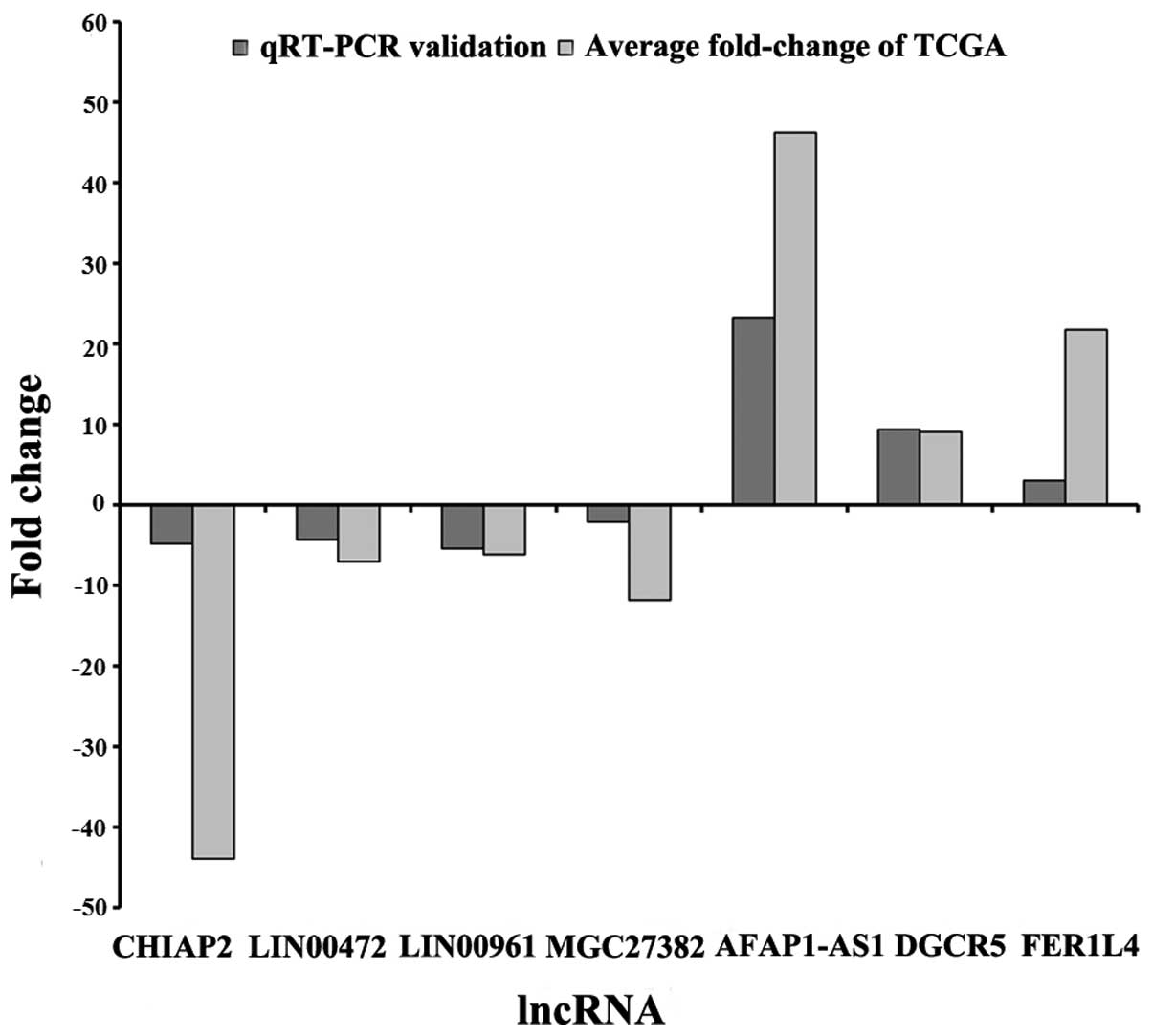
Finally, we randomly selected 7 specific lncRNAs
(AFAP1-AS1, CHIAP2, DGCR5, FER1L4, LINC00472, LINC00961 and
MGC27382) to validate the reliability and validity of the above
analysis results. We applied the paired t-test to assess the
differences between the LUAD tumor tissues and the adjacent
non-tumor lung tissues. The results showed that CHIAP2, LINC00472,
LINC00961 and MGC27382 were downregulated in LUAD tumor tissues
when compared with adjacent non-tumor lung tissues, while
AFAP1-AS1, CHIAP2 and FER1L4 were upregulated in LUAD tumor tissues
(Fig. 8 and Table V). The results from the qRT-PCR
validation in 53 newly diagnosed LUAD patients were consistent with
the above bioinformatics results (Table I). Then, we analyzed the 7 lncRNAs
with clinical features. The result showed that AFAP1-AS1 was
significantly related with tumor pathological stage, and LINC00472
was significantly related with lymph node metastasis (Fig. 9). The clinical relevance of the 2
lncRNAs was substantially identical with the results of
bioinformatics analysis. The above results revealed that our
bioinformatics analysis was credible.
| Table VRelative expression of 7 lncRNAs in
the LUAD tumor tissues and the adjacent non-tumor lung tissues. |
Table V
Relative expression of 7 lncRNAs in
the LUAD tumor tissues and the adjacent non-tumor lung tissues.
| lncRNAs | Tissues | ΔCT
(mean ± SD) | ΔΔCT
(mean ± SD) |
2−ΔΔCt | P-valuea |
|---|
| CHIAP2 | C | 11.492±3.274 | 2.267±3.834 | 0.206 | 0.000b |
| N | 9.225±2.448 | | | |
| LINC00472 | C | 10.463±2.038 | 0.275±3.143 | 0.826 | 0.002b |
| N | 10.188±3.091 | | | |
| LINC00961 | C | 11.361±2.027 | 2.109±2.917 | 0.232 | 0.000b |
| N | 9.253±2.372 | | | |
| MGC27382 | C | 8.419±2.559 | 2.443±3.059 | 0.184 | 0.000b |
| N | 5.977±2.397 | | | |
| AFAP1-AS1 | C | 9.143±3.779 | −4.539±4.813 | 23.244 | 0.000b |
| N | 13.682±4.450 | | | |
| DGCR5 | C | 13.504±2.427 | −3.227±4.183 | 9.365 | 0.000b |
| N | 16.669±3.591 | | | |
| FER1L4 | C | 7.887±1.983 | −1.585±2.840 | 2.999 | 0.000b |
| N | 9.472±1.994 | | | |
Discussion
Lung cancer is current the neoplasia with the
highest global incidence and mortality (36,37).
LUAD, a cancer that originates in lung glandular cells, is the most
frequent type of lung cancer (38). It has been reported that LUAD is
steadily rising in both men and women replacing lung squamous cell
carcinoma as the most common pathological type of lung cancer in
Korea (39). While major
improvements in diagnosis, surgical skills, and medical treatments
of lung cancer have been applied over the past few years, the
average 5-year survival rate remains ~10% (40), mainly because diagnosis cannot find
all early stage of the disease. Although several investigative
methods are available to support diagnosis and staging of LUAD,
such as biopsy sampling, mediastinoscopy, and bronchoscopy, these
methods are time-consuming, which may lead to delays in early
treatment (41). Therefore, to
improve this situation, the identification of cancer-related genes
and the exact regulatory mechanism of LUAD initiation and
development have received increasing attention.
Aberrant expression of lncRNAs has been widely
observed in various diseases, and studies have shown that the
dysregulated lncRNAs emerge as key roles in vital biological
functions of cancers (42,43). However, only a few studies have
described lncRNA profiles in LUAD by microarray analysis (31). Song et al (44) have constructed lncRNA-mRNA network
by dysregulated lncRNAs and mRNAs based on ceRNA hypothesis. To
date, only a few studies represent the interaction between lncRNA
and mRNA, or lncRNA and miRNA in LUAD, the results of which showed
that lncRNAs can function as a key part of ceRNA network, but such
ceRNA network is still poorly described (45–48).
The ceRNA hypothesis has been proposed as a novel regulatory
mechanism functioning through miRNA competition (27,49).
With further research of ceRNA crosstalk, studies showed that ceRNA
genes were mediated by miRNAs, which interacted with lncRNAs, in
the complex ceRNA network (50).
In the present study, we identified lncRNAs, miRNAs
and mRNAs from TCGA database, which were related with TNM stage and
lymph node metastasis in LUAD. Then, we constructed the
lncRNA-miRNA-mRNA ceRNA network with TNM stage and lymph node
metastasis-related LUAD-specific lncRNAs, miRNAs and mRNAs, which
provides integrated biological views of the ceRNA network. The
constructed LUAD-related lncRNA-associated ceRNA network provides
important hints for detecting the key RNAs of ceRNA-mediated gene
regulatory network in the initiation and development of LUAD. Next,
we selected the cancer specific key lncRNAs from ceRNA network and
investigated their distributions in different LUAD clinical
features and their associations with overall survival on the basis
of RNA sequencing profile of 465 LUAD tissues and 49 adjacent
non-tumor lung tissues. We randomly selected 7 specific lncRNAs
(AFAP1-AS1, CHIAP2, DGCR5, FER1L4, LINC00472, LINC00961 and
MGC27382) and analyzed their expression levels in the 53 newly
diagnosed tumor tissues of LUAD patients and adjacent non-tumor
lung tissues using qRT-PCR.
Based on the RNA sequence data from TCGA database,
we divided LUAD patients into 4 groups with TNM stage and lymph
node metastasis as condition, and they were compared with adjacent
non-tumor lung tissues, respectively. Then, we selected the
intersection RNAs. In the next step, we investigated bioinformatics
of aberrantly expressed mRNAs, and the front enriched functional
annotation of GO and KEGG pathway might be the potential function
of lncRNA. The results of GO biological process terms showed that
the aberrantly expressed genes mainly concentrate on cellular
functions, metabolism and immune functions. Based on KEGG pathway
analysis, a few cancer-related pathways were detected, including
p53 signaling pathway, MAPK signaling pathway and Wnt signaling
pathway. Wu et al (51)
merged two datasets to find novel target genes and pathways to
explain the pathogenicity of LUAD, and they found that PPM1D and
GADD45B may regulate LUAD progression through p53 signaling
pathway. Wang et al (52)
established a co-culture system of A549 cells and bone
marrow-derived cells (HS-5) to investigate the molecular mechanism
of BMP9 in LUAD and the bone metastatic microenvironment. Then, he
found BMP9 can inhibit the growth and migration of LUAD A549 cells
through the MAPK/ERK and NF-κB pathways. It was also reported that
Wnt signaling pathways were related with LUAD (53–55).
Growing body of evidence has presented that lncRNAs
function as a crucial component of ceRNA network by modulating
other RNA transcripts (19,30,33,56–58).
For example, HOTAIR may act as an endogenous sink by binding
miR-331-3p, thereby abolishing the miRNA-induced inhibitory
activity on the HER2 3′-UTR and increasing an additional level of
post-transcriptional regulation (59). Hence, the potential connection
lncRNA, miRNA and mRNA may exist in initiation and development of
LUAD. In the present study, we constructed the lncRNA-miRNA-mRNA
ceRNA network to reveal a novel ceRNA regulatory network in LUAD.
We found some cancer special lncRNAs, such as FER1L4 (60), MGC27382 (61), UCA1 (25), AFAP1-AS1 (62), were also reported in cancers acting
as potential diagnosis and prognostic biomarkers. In addition, we
found THBS2 (63), RAD51 (64), SLC2A1 (65), BDNF (66), in the ceRNA network were also
related with initiation and development of cancers. In the present
study, we analyzed the LUAD-specific lncRNA indirectly related mRNA
signal pathways involved in ceRNA network. The results of pathway
analysis showed that there were 9 pathways related with cancer.
Therefore, our results suggested that the key lncRNAs may play an
important role in initiation and development of LUAD and the cancer
genes related pathways.
With respect to the associations between 29 cancer
specific lncRNAs from ceRNA network and clinical features,
including gender, tumor pathological stage, TNM staging system,
lymph node metastasis and patient outcome assessment, we found that
21 lncRNAs were related to clinical features. Eight of these 21
lncRNAs were reported to be associated with cancer. For example,
UCA1 was reported to be an indicator of early gastric cancer
(67). However, there is no report
on the association between LUAD and the above lncRNAs features.
Then we analyzed the associations between 29 cancer-specific
lncRNAs and the patient survival, and we found that 5 lncRNAs were
related to LUAD OS.
Finally, 7 cancer specific lncRNAs were randomly
selected to verify the expressions of specific lncRNAs and
credibility of bioinformatics analysis using qRT-PCR. The
expression data from TCGA and verification result of 53 newly
diagnosed LUAD patients were 100% in agreement. Then, we performed
the correlation analysis between the 7 cancer specific lncRNAs and
clinical features. The results showed that AFAP1-AS1 and LINC00472
were significantly correlated with tumor pathological stage and
lymph node metastasis, respectively. These above results revealed
that our bioinformatics analysis is credible.
Based on these above LUAD OS results, only 1 of
these 5 LUAD-OS related lncRNAs (LINC00472) was aberrantly
expressed in clinical feature. Therefore, lncRNAs that aberrantly
expressed in clinical feature comparisons may not be necessarily
related with OS (29). Among these
5 lncRNAs only LINC00472 has been reported in the survival of
carcinoma and the other lncRNAs (BCRP3, CHIAP2, BMS1P20 and
UNQ6494) have not been reported (68). Furthermore, a recent study also
found that high LINC00472 expression could significantly reduce
risk of relapse and death in breast cancer patients, and LINC00472
could also suppress breast cancer cell proliferation and migration
(69). It implies that LINC00472
may play an important role in LUAD.
It has been reported that upregulated AFAP1-AS1 was
associated with the poor prognosis of non-small cell lung cancer
(NSCLC) patients (70).
Furthermore, AFAP1-AS1 was also reported to be associated with
prognosis, cell proliferation and invasion of other cancers
(62,71,72).
Combined with the present study (Tables I, IV, V
and Figs. 6Figure 7–8), showing that the expression of
AFAP1-AS1 and LINC00472 were dysregulated in LUAD from TCGA
database and qRT-PCR verification, and they were related with tumor
pathological stage, and lymph node metastasis and OS, respectively,
it reveals that AFAP1-AS1 and LINC00472 may be potential biomarkers
in LUAD.
In the present study, we identified the
LUAD-specific lncRNAs from hundreds of candidate lncRNAs detected
from large scale samples in TCGA database, and revealed aberrant
expression profiles of cancer specific lncRNAs under different
clinical features. The aberrantly expressed key lncRNAs, which were
identified in LUAD, may shed light on sensitive biomarkers in LUAD.
Importantly, we have constructed the lncRNA-miRNA-mRNA ceRNA
network to clarify the unknown ceRNA regulatory network in LUAD. In
addition, qRT-PCR verification was used for the reliability and
validity of expression of key lncRNAs and bioinformatics analysis.
Our findings provide novel insight into better understanding of
lncRNA-related ceRNA network in LUAD and potential biomarkers for
diagnosis and prognosis.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (81472939, 81172618 and
81673132), the Qing Lan Project, the 333 Project of Jiangsu
Province, the Liu Da Ren Cai Gao Feng Project of Jiangsu Province
and the Fundamental Research Funds for the central universities and
innovative Research Project for postgraduates in Colleges of
Jiangsu Province. We thank Mr. Donglin Cheng for his technical
assistance.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu L, Wu S, Yang Y, Cai J, Zhu X, Wu J,
Li M and Guan H: SOSTDC1 is down-regulated in non-small cell lung
cancer and contributes to cancer cell proliferation. Cell Biosci.
6:242016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li X, Shi Y, Yin Z, Xue X and Zhou B: An
eight-miRNA signature as a potential biomarker for predicting
survival in lung adenocarcinoma. J Transl Med. 12:1592014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008 GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
6
|
Wang Y, Zhao H, Gao X, Wei F, Zhang X, Su
Y, Wang C, Li H and Ren X: Identification of a three-miRNA
signature as a blood-borne diagnostic marker for early diagnosis of
lung adenocarcinoma. Oncotarget. 7:26070–26086. 2016.PubMed/NCBI
|
|
7
|
Schneider F, Derrick V, Davison JM,
Strollo D, Incharoen P and Dacic S: Morphological and molecular
approach to synchronous non-small cell lung carcinomas: impact on
staging. Mod Pathol. 29:735–742. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qu J, Li M, An J, Zhao B, Zhong W, Gu Q,
Cao L, Yang H and Hu C: MicroRNA-33b inhibits lung adenocarcinoma
cell growth, invasion, and epithelial-mesenchymal transition by
suppressing Wnt/β-catenin/ZEB1 signaling. Int J Oncol.
47:2141–2152. 2015.PubMed/NCBI
|
|
9
|
Murayama T, Nakaoku T, Enari M, Nishimura
T, Tominaga K, Nakata A, Tojo A, Sugano S, Kohno T and Gotoh N:
Oncogenic fusion gene CD74-NRG1 confers cancer stem cell-like
properties in lung cancer through a IGF2 autocrine/paracrine
circuit. Cancer Res. 76:974–983. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang G, Chen H and Liu J: The long
noncoding RNA LINC01207 promotes proliferation of lung
adenocarcinoma. Am J Cancer Res. 5:3162–3173. 2015.PubMed/NCBI
|
|
11
|
Sana J, Faltejskova P, Svoboda M and Slaby
O: Novel classes of non-coding RNAs and cancer. J Transl Med.
10:1032012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Takahashi K, Yan I, Haga H and Patel T:
Long noncoding RNA in liver diseases. Hepatology. 60:744–753. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Braconi C, Valeri N, Kogure T, Gasparini
P, Huang N, Nuovo GJ, Terracciano L, Croce CM and Patel T:
Expression and functional role of a transcribed noncoding RNA with
an ultraconserved element in hepatocellular carcinoma. Proc Natl
Acad Sci USA. 108:786–791. 2011. View Article : Google Scholar :
|
|
14
|
Jiang F, Zhou X and Huang J: Long
non-coding RNA-ROR mediates the reprogramming in cardiac
hypertrophy. PLoS One. 11:e01527672016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang L, Qi H, Xiao Y, Li C, Wang Y, Guo
T, Liu Z and Liu Q: Integrated analysis of noncoding RNAs and mRNAs
reveals their potential roles in the biological activities of the
growth hormone receptor. Growth Horm IGF Res. 29:11–20. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Voellenkle C, Garcia-Manteiga JM, Pedrotti
S, Perfetti A, De Toma I, Da Silva D, Maimone B, Greco S, Fasanaro
P, Creo P, et al: Implication of long noncoding RNAs in the
endothelial cell response to hypoxia revealed by RNA-sequencing.
Sci Rep. 6:241412016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yoon JH, Abdelmohsen K and Gorospe M:
Posttranscriptional gene regulation by long noncoding RNA. J Mol
Biol. 425:3723–3730. 2013. View Article : Google Scholar
|
|
19
|
Zhou M, Wang X, Shi H, Cheng L, Wang Z,
Zhao H, Yang L and Sun J: Characterization of long non-coding
RNA-associated ceRNA network to reveal potential prognostic lncRNA
biomarkers in human ovarian cancer. Oncotarget. 7:12598–12611.
2016.PubMed/NCBI
|
|
20
|
Hung T and Chang HY: Long noncoding RNA in
genome regulation: Prospects and mechanisms. RNA Biol. 7:582–585.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi X, Ma C, Zhu Q, Yuan D, Sun M, Gu X,
Wu G, Lv T and Song Y: Upregulation of long intergenic noncoding
RNA 00673 promotes tumor proliferation via LSD1 interaction and
repression of NCALD in non-small-cell lung cancer. Oncotarget.
7:25558–25575. 2016.PubMed/NCBI
|
|
22
|
Wang PL, Liu B, Xia Y, Pan CF, Ma T and
Chen YJ: Long non-coding RNA-low expression in tumor inhibits the
invasion and metastasis of esophageal squamous cell carcinoma by
regulating p53 expression. Mol Med Rep. 13:3074–3082.
2016.PubMed/NCBI
|
|
23
|
Shang C, Guo Y, Zhang J and Huang B:
Silence of long noncoding RNA UCA1 inhibits malignant proliferation
and chemotherapy resistance to adriamycin in gastric cancer. Cancer
Chemother Pharmacol. 77:1061–1067. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang D, Cao C, Liu L and Wu D:
Up-regulation of LncRNA SNHG20 predicts poor prognosis in
hepatocellular carcinoma. J Cancer. 7:608–617. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bian Z, Jin L, Zhang J, Yin Y, Quan C, Hu
Y, Feng Y, Liu H, Fei B, Mao Y, et al: LncRNA-UCA1 enhances cell
proliferation and 5-fluorouracil resistance in colorectal cancer by
inhibiting miR-204–5p. Sci Rep. 6:238922016. View Article : Google Scholar
|
|
26
|
Sun R, Qin C, Jiang B, Fang S, Pan X, Peng
L, Liu Z, Li W, Li Y and Li G: Down-regulation of MALAT1 inhibits
cervical cancer cell invasion and metastasis by inhibition of
epithelial-mesenchymal transition. Mol Biosyst. 12:952–962. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pilyugin M and Irminger-Finger I: Long
non-coding RNA and microRNAs might act in regulating the expression
of BARD1 mRNAs. Int J Biochem Cell Biol. 54:356–367. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang J, Fan D, Jian Z, Chen GG and Lai
PB: Cancer specific long noncoding RNAs show differential
expression patterns and competing endogenous RNA potential in
hepatocellular carcinoma. PLoS One. 10:e01410422015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li CY, Liang GY, Yao WZ, Sui J, Shen X,
Zhang YQ, Peng H, Hong WW, Ye YC, Zhang ZY, et al: Integrated
analysis of long non-coding RNA competing interactions reveals the
potential role in progression of human gastric cancer. Int J Oncol.
48:1965–1976. 2016.PubMed/NCBI
|
|
31
|
Chen J, Hu L, Chen J, Pan Q, Ding H, Xu G,
Zhu P, Wen X, Huang K and Wang Y: Detection and analysis of Wnt
pathway related lncRNAs expression profile in lung adenocarcinoma.
Pathol Oncol Res. 22:609–615. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
McCarter JP, Mitreva MD, Martin J, Dante
M, Wylie T, Rao U, Pape D, Bowers Y, Theising B, Murphy CV, et al:
Analysis and functional classification of transcripts from the
nematode Meloidogyne incognita. Genome Biol. 4:R262003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guo LL, Song CH, Wang P, Dai LP, Zhang JY
and Wang KJ: Competing endogenous RNA networks and gastric cancer.
World J Gastroenterol. 21:11680–11687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42(D1): D92–D97. 2014. View Article : Google Scholar
|
|
35
|
Hsu SD, Tseng YT, Shrestha S, Lin YL,
Khaleel A, Chou CH, Chu CF, Huang HY, Lin CM, Ho SY, et al:
miRTarBase update 2014. An information resource for experimentally
validated miRNA-target interactions. Nucleic Acids Res. 42(D1):
D78–D85. 2014. View Article : Google Scholar
|
|
36
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Meza R, Meernik C, Jeon J and Cote ML:
Lung cancer incidence trends by gender, race and histology in the
United States, 1973–2010. PLoS One. 10:e01213232015. View Article : Google Scholar
|
|
38
|
Kerr KM: Pulmonary adenocarcinomas:
Classification and reporting. Histopathology. 54:12–27. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park JY and Jang SH: Epidemiology of lung
cancer in Korea: Recent trends. Tuberc Respir Dis (Seoul).
79:58–69. 2016. View Article : Google Scholar
|
|
40
|
Ortea I, Rodríguez-Ariza A, Chicano-Gálvez
E, Arenas Vacas MS and Jurado Gámez B: Discovery of potential
protein biomarkers of lung adenocarcinoma in bronchoalveolar lavage
fluid by SWATH MS data-independent acquisition and targeted data
extraction. J Proteomics. 138:106–114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Navani N, Nankivell M, Lawrence DR, Lock
S, Makker H, Baldwin DR, Stephens RJ, Parmar MK, Spiro SG, Morris
S, et al; Lung-BOOST trial investigators. Lung cancer diagnosis and
staging with endobronchial ultrasound-guided transbronchial needle
aspiration compared with conventional approaches: An open-label,
pragmatic, randomised controlled trial. Lancet Respir Med.
3:282–289. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Harries LW: Long non-coding RNAs and human
disease. Biochem Soc Trans. 40:902–906. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Song C, Zhang J, Liu Y, Pan H, Qi HP, Cao
YG, Zhao JM, Li S, Guo J, Sun HL, et al: Construction and analysis
of cardiac hypertrophy-associated lncRNA-mRNA network based on
competitive endogenous RNA reveal functional lncRNAs in cardiac
hypertrophy. Oncotarget. 7:10827–10840. 2016.PubMed/NCBI
|
|
45
|
Peters T, Hermans-Beijnsberger S, Beqqali
A, Bitsch N, Nakagawa S, Prasanth KV, de Windt LJ, van Oort RJ,
Heymans S and Schroen B: Long non-coding RNA malat-1 is dispensable
during pressure overload-induced cardiac remodeling and failure in
mice. PLoS One. 11:e01502362016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou KI, Parisien M, Dai Q, Liu N,
Diatchenko L, Sachleben JR and Pan T: N6-Methyladenosine
modification in a long noncoding RNA hairpin predisposes its
conformation to protein binding. J Mol Biol. 428(5 Pt A): 822–833.
2016. View Article : Google Scholar
|
|
47
|
Wei G: Bioinformatics analysis of microRNA
comprehensive regulatory network in congenital microtia. Int J
Pediatr Otorhinolaryngol. 79:1727–1731. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu S, Song L, Zeng S and Zhang L:
MALAT1-miR-124-RBG2 axis is involved in growth and invasion of
HR-HPV-positive cervical cancer cells. Tumour Biol. 37:633–640.
2016. View Article : Google Scholar
|
|
49
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Karreth FA and Pandolfi PP: ceRNA
cross-talk in cancer: When ce-bling rivalries go awry. Cancer
Discov. 3:1113–1121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wu X, Zang W, Cui S and Wang M:
Bioinformatics analysis of two microarray gene-expression data sets
to select lung adenocarcinoma marker genes. Eur Rev Med Pharmacol
Sci. 16:1582–1587. 2012.PubMed/NCBI
|
|
52
|
Wang J, Weng Y, Zhang M, Li Y, Fan M, Guo
Y, Sun Y, Li W and Shi Q: BMP9 inhibits the growth and migration of
lung adenocarcinoma A549 cells in a bone marrow stromal
cell-derived microenvironment through the MAPK/ERK and NF-κB
pathways. Oncol Rep. 36:410–418. 2016.PubMed/NCBI
|
|
53
|
Sunaga N, Kohno T, Kolligs FT, Fearon ER,
Saito R and Yokota J: Constitutive activation of the Wnt signaling
pathway by CTNNB1 (beta-catenin) mutations in a subset of human
lung adenocarcinoma. Genes Chromosomes Cancer. 30:316–321. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jiang HL, Jiang LM and Han WD:
Wnt/beta-catenin signaling pathway in lung cancer stem cells is a
potential target for the development of novel anticancer drugs. J
BUON. 20:1094–1100. 2015.PubMed/NCBI
|
|
55
|
Qi W, Chen J, Cheng X, Huang J, Xiang T,
Li Q, Long H and Zhu B: Targeting the Wnt-regulatory protein
CTNNBIP1 by microRNA-214 enhances the stemness and self-renewal of
cancer stem-like cells in lung adenocarcinomas. Stem Cells.
33:3423–3436. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hu Y, Tian H, Xu J and Fang JY: Roles of
competing endogenous RNAs in gastric cancer. Brief Funct Genomics.
15:266–273. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wu Q, Guo L, Jiang F, Li L, Li Z and Chen
F: Analysis of the miRNA-mRNA-lncRNA networks in ER+ and
ER− breast cancer cell lines. J Cell Mol Med.
19:2874–2887. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Song X, Cao G, Jing L, Lin S, Wang X,
Zhang J, Wang M, Liu W and Lv C: Analysing the relationship between
lncRNA and protein-coding gene and the role of lncRNA as ceRNA in
pulmonary fibrosis. J Cell Mol Med. 18:991–1003. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:922014.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu Z, Shao Y, Tan L, Shi H, Chen S and
Guo J: Clinical significance of the low expression of FER1L4 in
gastric cancer patients. Tumour Biol. 35:9613–9617. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kim ST, Sohn I, Do IG, Jang J, Kim SH,
Jung IH, Park JO, Park YS, Talasaz A, Lee J, et al: Transcriptome
analysis of CD133-positive stem cells and prognostic value of
survivin in colorectal cancer. Cancer Genomics Proteomics.
11:259–266. 2014.PubMed/NCBI
|
|
62
|
Zhang JY, Weng MZ, Song FB, Xu YG, Liu Q,
Wu JY, Qin J, Jin T and Xu JM: Long noncoding RNA AFAP1-AS1
indicates a poor prognosis of hepatocellular carcinoma and promotes
cell proliferation and invasion via upregulation of the RhoA/Rac2
signaling. Int J Oncol. 48:1590–1598. 2016.PubMed/NCBI
|
|
63
|
Tian ZQ, Li ZH, Wen SW, Zhang YF, Li Y,
Cheng JG and Wang GY: Identification of commonly dysregulated genes
in non-small-cell lung cancer by integrated analysis of microarray
data and qRT-PCR validation. Lung. 193:583–592. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Jiang Y, Verbiest T, Devery AM, Bokobza
SM, Weber AM, Leszczynska KB, Hammond EM and Ryan AJ: Hypoxia
potentiates the radiation-sensitizing effect of olaparib in human
non-small cell lung cancer xenografts by contextual synthetic
lethality. Int J Radiat Oncol Biol Phys. 95:772–781. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tang Y, Cao K, Wang Q, Chen J, Liu R, Wang
S, Zhou J and Xie H: Silencing of CerS6 increases the invasion and
glycolysis of melanoma WM35, WM451 and SK28 cell lines via
increased GLUT1-induced downregulation of WNT5A. Oncol Rep.
35:2907–2915. 2016.PubMed/NCBI
|
|
66
|
Correa DD, Satagopan J, Cheung K, Arora
AK, Kryza-Lacombe M, Xu Y, Karimi S, Lyo J, DeAngelis LM and Orlow
I: COMT, BDNF, and DTNBP1 polymorphisms and cognitive functions in
patients with brain tumors. Neuro Oncol. Apr 18–2016.pii: now057
(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Gao J, Cao R and Mu H: Long non-coding RNA
UCA1 may be a novel diagnostic and predictive biomarker in plasma
for early gastric cancer. Int J Clin Exp Pathol. 8:12936–12942.
2015.
|
|
68
|
Shen Y, Wang Z, Loo LW, Ni Y, Jia W, Fei
P, Risch HA, Katsaros D and Yu H: LINC00472 expression is regulated
by promoter methylation and associated with disease-free survival
in patients with grade 2 breast cancer. Breast Cancer Res Treat.
154:473–482. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Shen Y, Katsaros D, Loo LW, Hernandez BY,
Chong C, Canuto EM, Biglia N, Lu L, Risch H, Chu WM, et al:
Prognostic and predictive values of long non-coding RNA LINC00472
in breast cancer. Oncotarget. 6:8579–8592. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Deng J, Liang Y, Liu C, He S and Wang S:
The up-regulation of long non-coding RNA AFAP1-AS1 is associated
with the poor prognosis of NSCLC patients. Biomed Pharmacother.
75:8–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Lu X, Zhou C, Li R, Liang Z, Zhai W, Zhao
L and Zhang S: Critical role for the long non-coding RNA AFAP1-AS1
in the proliferation and metastasis of hepatocellular carcinoma.
Tumour Biol. 7:9699–9707. 2016. View Article : Google Scholar
|
|
72
|
Yang F, Lyu S, Dong S, Liu Y, Zhang X and
Wang O: Expression profile analysis of long noncoding RNA in
HER-2-enriched subtype breast cancer by next-generation sequencing
and bioinformatics. Onco Targets Ther. 9:761–772. 2016. View Article : Google Scholar : PubMed/NCBI
|