Introduction
Multiple myeloma (MM) is a hematologic malignancy of
terminally differentiated plasma cells. A most common complication
of MM is myeloma bone disease (MBD), occurring in >80% of MM
patients. Bone lesions in MM are the result of imbalance between
osteoclasts (OCs) and osteoblasts (OBs), the activation of OCs and
inhibition of OBs (1,2). Many studies have focused on the
function of OCs and found RANKL/RANK system, which became the
target of treatement (3,4). However, osteolytic lesions still
exist after the inhibition of OCs, which is associated with the
inhibition of OBs. Recent research showed many cytokines in bone
marrow (BM) microenvironment participate in the pathogenesis of
MBD, such as CCL3 (5,6), activin A (7), alpha hypoxia-inducible factor
(HIF-1α) (8), pim-2 (9), cysteine-rich protein 61 (CYR61)
(10), which may be new targets in
MBD.
CYR61 is the first cloned member of the CCN family,
an immediate early gene family consisting of six members, CTGF,
NOV, WISP-1, WISP-2 and WISP-3 (11,12).
CYR61 is known to link cell surface and extracellular matrix and
plays important roles on cell adhesion, proliferation, migration,
differentiation and angiogenesis during normal developmental and
pathophysiological processes (11). Clines et al (13) found that tumor-produced
endothelin-1 increased osteoblasts proliferation and new bone
formation by activating Wnt signaling pathway through suppression
of Wnt pathway inhibitor DKK1 in murine primary OBs culture. Si
et al (14) suggested that
tightly regulated CCN1/CYR61 expression may play an important role
in Wnt3A-induced OBs differentiation from mesenchymal stem cells.
In MC3T3-E1 osteoblasts, CYR61 upregulated BMP-2, resulting in
enhanced cell proliferation and osteoblastic differentiation
through activation of the α(v)β(3)
integrin/integrin-linked kinase/ERK signaling pathway (15). The study of Chen et al
(16) in osteoarthritis indicated
that CCN1 heightened OSM expression via α(v)β(3) receptor, FAK, c-Src, PI3K, and NF-κB
pathway in osteoblastic cells. Johnson et al (10) found that recombinant CCN1 directly
inhibited growth of MM cells in vitro, and overexpression of
CYR61 in MM cells prevented bone destruction in vivo.
However, still scarce research exists of CCN1 on OBs in MM
patients. In our study, we cultured OBs from MBD patients with
recombinant CCN1 and observed the effect of CCN1 on OBs in
vitro, then further investigated the effect of CCN1 on bone
remodeling in vivo.
Patients and methods
Study subjects
Thirty-six newly diagnosed MM patients (20 males and
16 females) were enrolled in this study. The participants were
selected as inpatients in the Hematology Department of Tianjin
Medical University General Hospital from January 2015 to November
2016 according to the International Myeloma Workgroup criteria.
According to X-ray scanning data obtained before treatment, bone
disease was graded into three stages: stage A, no osteolytic
lesions or osteoporosis alone; stage B, one to three osteolytic
lesions; stage C, more than three osteolytic lesions and/or a
pathological fracture (17). There
were 9 patients in stage A and 27 in stage B/C in our study. The
characteristics of the patients are shown in Table I. Eighteen healthy volunteers with
a median age of 48 years (range, 42–69 years) were included as
normal controls. Bone marrow aspirations were collected from all
the patients diagnosed with MM and from normal controls. This study
was approved by the Ethics Committee of the Tianjin Medical
University. Written informed consent was obtained from the patients
for the publication of this report and any accompanying images.
 | Table IProfiles of patients with MM enrolled
in this study. |
Table I
Profiles of patients with MM enrolled
in this study.
| Total no. of
patients | 36 |
| Gender | 20 M-16 F |
| Age-median
(range) | 68 (43–80) |
| Type of MM | IgG (17), IgM (7), IgA (5), light-chain (6), non-secretory (1) |
| Stage at diagnosis
(ISS) | |
| I | 6 (17%) |
| II | 8 (22%) |
| III | 22 (61%) |
| Bone disease | |
| Stage A | 9 (25%) |
| Stage B/C | 27 (75%) |
| Parameters at
baseline | |
| Creatinine
clearance >30 ml/min | 18 (50%) |
| Creatinine
clearance ≤30 ml/min | 18 (50%) |
| Hb ≥100 g/l | 7 (19%) |
| Hb <100
g/l | 29 (81%) |
| Ca >2.75
mmol/l | 7 (19%) |
| Ca ≤2.75
mmol/l | 29 (81%) |
Enzyme-linked immunosorbent assay
The CCN1 level in the bone marrow supernatant was
assessed using the CCN1 ELISA kit (R&D Systems, Inc.,
Minneapolis, MN, USA). Human paraprotein (Lambda) level in the
mouse model was detected using the Human Lambda ELISA kit (Bethyl
Laboratories, Inc., Montgomery, TX, USA). Diluted standards and
patient serum (100 µl) were added in duplicate and incubated
at 37°C for 1 h. After washing the plates 5 times, 100 µl of
antibody was added to each well and incubated at room temperature
for 1 h. Next, HRP was added to each well. After incubation at 37°C
for 30 min, the wells were washed 5 times. Then, TMB solution was
added to each well, and the samples were incubated in the dark at
room temperature for 20 min. Finally, a stop solution was added,
and the OD was read at 450 nm within 15 min.
Cell culture
The bone marrow mononuclear cells (BMMNC) were
separated using Ficoll-Hypaque density sedimentation. The BMMNCs
were cultured in Dulbecco's modified Eagle's medium/F12 medium
supplemented with 15% fetal bovine serum (Gibco, Darmstadt,
Germany), 1×10−7 mol/l dexamethasone, 0.05 g/l vitamin
C, 0.01 mol/l β-sodium glycerophosphate, 100 g/ml penicillin
(Gibco), and 100 U/ml streptomycin (Gibco). Non-adherent cells were
removed the next day, and the media were replaced every other day.
Adherent BMMNCs were cultured at 37°C in an atmosphere containing
5% CO2. The OBs were counted and seeded in 24-well
plates at a plating density of 1×104
cells/cm2. Trypsin was used to detach three wells of OBs
for cell count every second day. The number of cells was used to
draw the OB growth curve. The OB doubling time (DT) was calculated
using the following formula: DT = t ×
[lg2/(lgNt − lgNo)], where t is the culturing
period (h); No is the cell density when the cells were
seeded; and Nt is the cell density when the cells were
cultured after t hours.
The OBs were divided into two groups: the blank
group and CCN1 group (OBs were cultured with CCN1: 10 and 30 ng/l).
The changes in the OB osteogenic potential and biological
characteristics were observed after intervention.
Quantitative real-time PCR
The total RNA from the OBs of each group was
extracted using the TRIzol reagent (Invitrogen). The TIANScript RT
kit (Tiangen, Beijing, China) was utilized to reverse-transcribe 1
µg of RNA. Table II
presented the primer sequences of runt-related transcription factor
2 (Runx2), β-catenin, CYR61, bone morphogenetic protein-2 (BMP2)
and GAPDH. These primer sequences were designed and synthesized by
Sangon Biotech (Shanghai, China).
| Table IIPrimer sequences. |
Table II
Primer sequences.
| Target | Sense and antisense
sequences | bp |
|---|
| Runx2 | F:
5′-GACGAGGCAAGAGTTTCACC-3′ | 106 |
| R:
5′-GGTTCCCGAGGTCCATCTAC-3′ | |
| β-catenin | F:
5′-GCTTTGTGTCGTCTTGAACG-3′ | 122 |
| R:
5′-TCAGCAATCCCTTTCTCACC-3′ | |
| CYR61 | F:
5′-GAAGCGGCTCCCTGTTTTTG-3′ | 174 |
|
R:5′-CGGGTTTCTTTCACAAGGCG-3′ | |
| BMP2 | F:
5′-TCAAGCCAAACACAAACAGC-3′ | 103 |
| R:
5′-AGCCACAATCCAGTCATTCC-3′ | |
| GAPDH | F:
5′-GCACCGTCAAGGCTGAGAAC-3′ | 100 |
| R:
5′-TGGTGAAGACGCCAGTGGA-3′ | |
The quantitative real-time PCR was performed using
the Bio-Rad iQ 5 Real-time system (Bio-Rad, Hercules, CA, USA). The
SYBR Green (Invitrogen) was used as a double-strand DNA-specific
dye. The amplification of CYR61 and Runx2 utilized 40 cycles at
95°C for 30 sec and 95°C for 5 sec with the extension at 57°C for
30 sec. The amplification of BMP2 and β-catenin both utilized 45
cycles at 95°C for 30 sec and 95°C for 5 sec with the extension at
55°C for 30 sec. GAPDH was employed as the housekeeping gene to
standardize the targeted mRNA expression. The CYR61, Runx2,
β-catenin and BMP2 levels were calculated using the
2−∆∆Ct method [(Ct, target gene Ct,
GAPDH)sample − (Ct, target gene Ct,
GAPDH)control] after normalizing the data according to
the GAPDH mRNA expression.
ALP and Von Kossa staining
The alkaline phosphatase (ALP) expression was an
early osteoblast marker detected using the ALP Staining kit
(Sigma-Aldrich, Taufkirchen, Germany). The expression was utilized
to confirm the presence of OB. Von Kossa staining was performed to
confirm that the OB synthesized and mineralized the extracellular
matrix. The mineralized nodules were found in both groups cultured
for 3 weeks. The amount of mineralized nodules in each patient was
counted and used as an indicator of OB function.
Lentiviral vector and myeloma cell
transduction
The sequence of cDNA for human CYR61 (GENE-ID 3491)
was cloned into the Ubi-MCS-SV40-EGFP-IRES-puromycin vector.
Lentiviral particles were generated by cotransfecting the
expression vector Ubi-MCS-SV40-EGFP-IRES-puromycin and ViraPower
Packaging Mix into 293T cells according to the Invitrogen ViraPower
Lentiviral Expression Systems protocol (18).
Myeloma cells (RPMI-8226) were plated at a density
of 5×104 cells per well in a 6-well plate. For each of
the following 3 days, cells were exposed to viral supernatant at
1×108 TU/ml in the presence of polybrene (5
µg/ml). Transduced RPMI-8226/CYR61 cells were used for
experiments in vivo. The efficiency of transduction was
determined using analysis of GFP expression. We detected the
expression of CYR61/CCN1 by qRT-PCR, ELISA and western blotting. In
addition, the proliferation of non-transduced RPMI-8226, transduced
with CYR61 or EV RPMI-8226 cells was measured by CCK-8 kit.
Animal model of myeloma bone disease
Female NOD/SCIDcc null mice at 5 weeks of age (the
Chinese Academy of Medical Sciences Institute of Radiation
Medicine) were acclimatized to the animal housing facility for 1
week before experiments. All of the experimental procedures on
animals were carried out with strict adherence to the rules and
guidelines for the ethical use of animals in research.
Fifteen microliter of the cell suspension
(1×106 cells per inoculum) were injected directly into
the marrow space of right tibia. The needle was inserted 3–5 mm
down the diaphysis of the tibia through the cortex of the anteria
tuberosity. The left contralateral tibia was injected with PBS
alone as an internal control (19). Ten mice were investigated for
myeloma cell infiltration and bone disease at 3, 5 and 7 weeks
after transplantation, respectively.
In the next experiment, mice were randomized into
three groups (bone disease group as a control, zoledronic acid
group and CCN1 overexpression group) of 10 mice per group. For the
control and zoledronic acid groups, RPMI-8226/EV myeloma cells were
injected into the marrow space of right tibia. In parallel,
RPMI-8226/CYR61 cells were injected to the mice of CCN1
overexpression group. From the fourth week after myeloma cell
injection, mice of zoledronic acid group were injected
subcutaneously with antiresorptive bisphosphonate zoledronic acid
(Novartis Pharma) at 100 µg/kg/dose once weekly for 4 weeks.
All the mice were sacrificed at 7 weeks after myeloma cell
inplantation.
Analysis of bone disease (microCT)
Tibiae were fixed in formalin and scanned using a
microCT scanner (model 1172; Skyscan) at 50 kV and 200 AA with a
pixel size of 4.3 Am, an image being captured every 0.7° through
180° rotation of each bone. During scanning, the samples were
enclosed in a tightly fitting rigid plastic tube to prevent
movement, scanned images were reconstructed and analyzed using the
Skyscan Recon and Skyscan CT analysis software, respectively. The
following three-dimensional (3D) parameters were measured: total
bone volume/total volume, bone surface area/bone volume, trabecular
thickness, trabecular number, trabecular spacing and trabecular
bone volume (20).
Immunohistochemistry and histochemistry
in the mouse model
Immunohistochemistry (IHC) staining was performed
using formalin-fixed, complete decalcification was confirmed with 2
weeks of decalcification in 0.5 mol/l EDTA/0.5% paraformaldehyde in
PBS (pH 8.0) at 4°C and tibiae were then paraffin-embedded.
Longitudinal sections (5 µm) were prepared and stained with
IHC. Anti-CD138 antibody (sc-69833 Santa Cruz Biotechnology, Santa
Cruz, CA, USA; 1:200) was used. Tatrate-resistant acid phosphatase
(TRAP) staining, a well-recognized marker of osteoclasts in
vivo, was performed with an acid phosphatase kit (Sigma, St.
Louis, MO, USA). Analysis was done on an Olympus CX41 microscope
(21).
Statistical analysis
SPSS 16.0 (SPSS, Inc., Chicago, IL, USA) was used to
perform the statistical analysis. The data are expressed as the
mean ± SD or median. The unpaired t-test and one-way ANOVA analysis
were used to analyze the significance between different groups.
Non-parametric test was used when the data were of abnormal
distribution. A value of P<0.05 was considered statistically
significant.
Results
CYR61/CCN1 is elevated significantly in
all newly diagnosed MM patients, especially in patients without
bone disease (stage A)
We detected CCN1 level in BM supernatant by ELISA in
newly diagnosed patients and normal controls. The levels of CCN1 in
the patients in stage A and stage B/C were 416.90±179.78 and
299.48±160.65 pg/ml, which were both significantly higher than
normal controls [202.12±41.53 pg/ml (P=0.001 and 0.045)] (Fig. 1A). Also, the level of CCN1 in stage
A was significantly higher than stage B/C (P=0.035), indicating
that CCN1 may have a promoting role in bone remodeling.
Previous research has shown that CYR61 was not
expressed in normal plasma cells, MM cells, osteoclasts, but highly
expressed in MSCs (10). We
further detected the expression of CYR61 in OBs by real-time PCR.
The median CYR61 mRNA levels in the patients in stage A and stage
B/C were 5.45 (0.08–17.23) and 2.46 (1.68–16.48), both
significantly increased compared with normal controls (1.47,
0.01–4.76) (P<0.001 and P=0.036). Similarly, the level of stage
A was significantly higher than stage B/C (P=0.013) (Fig. 1B).
CCN1 promotes proliferation and
mineralization of OBs in vitro
The OBs from the patients in stage B/C were
incubated without or with CCN1 (10 and 30 ng/l) for 24 h in
vitro (16) and the quantity
and function of OBs were subsequently observed. Matrix formation
and mineralization were also investigated by histochemistry. ALP
and Von Kossa staining positively identified the OBs. The quantity
and function of OBs both continuously increased with increasing
CCN1 concentration (Fig. 1C–E).
The OB quantity cultured with different CCN1 concentrations (10 and
30 ng/l) were 3.01±0.49×105/ml and
3.39±1.21×105/ml, while there was significant difference
between the blank group and 30 ng/l CCN1 group (P=0.046). The
amount of mineralized nodules cultured with 30 ng/l CCN1
(14.33±5.72/HPF) was also significantly increased compared to the
blank group (9.11±0.97/HPF) (P=0.048), while there was no
significant difference in 10 ng/l CCN1 group (11.82±3.76/HPF)
(Table III). According to these
results, we found that suitable concentration of CCN1 stimulating
the OBs was 30 ng/l. Therefore, we detected the Runx2, β-catenin
and BMP2 by real-time PCR in 30 ng/l CCN1 incubation, not 10 ng/l
CCN1.
| Table IIIQuantity and function of OBs cultured
with different concentration of CCN1. |
Table III
Quantity and function of OBs cultured
with different concentration of CCN1.
| No. of cells
(105/ml) | No. of mineralized
nodules (/HPF) |
|---|
| Blank | 2.72±0.24 | 9.11±0.97 |
| CCN1 (10 ng/l) | 3.01±0.49 | 11.82±3.76 |
| CCN1 (30 ng/l) | 3.39±1.21a | 14.33± 5.72a |
Runx2 and β-catenin upregulation in OBs
after CCN1 stimulation
The mRNA expressions of Runx2, β-catenin and BMP2
were analyzed to observe OB differentiation. These expressions were
critical transcription factors of early OB differentiation
(22). The median mRNA of Runx2 in
CCN1 group (30 ng/l) was 0.9965 (range, 0.3751–9.7473), which was
significantly higher than that in the blank group (0.5421,
0.0938–1.8725) (P=0.012). The mRNA of β-catenin had a similar
result. The median mRNA of β-catenin after CCN1 stimulating
increased significantly from 0.6538 (0.2836–1.8751) to 0.7540
(0.3893–4.7142) (P=0.011). However, the mRNA of BMP2 had no
significant difference before and after CCN1 stimulation. The
median mRNA of BMP2 in the blank and CCN1 group were 1.2414
(0.0488–1.167) and 1.5390 (0.2001–1.3756) (Fig. 1F–H).
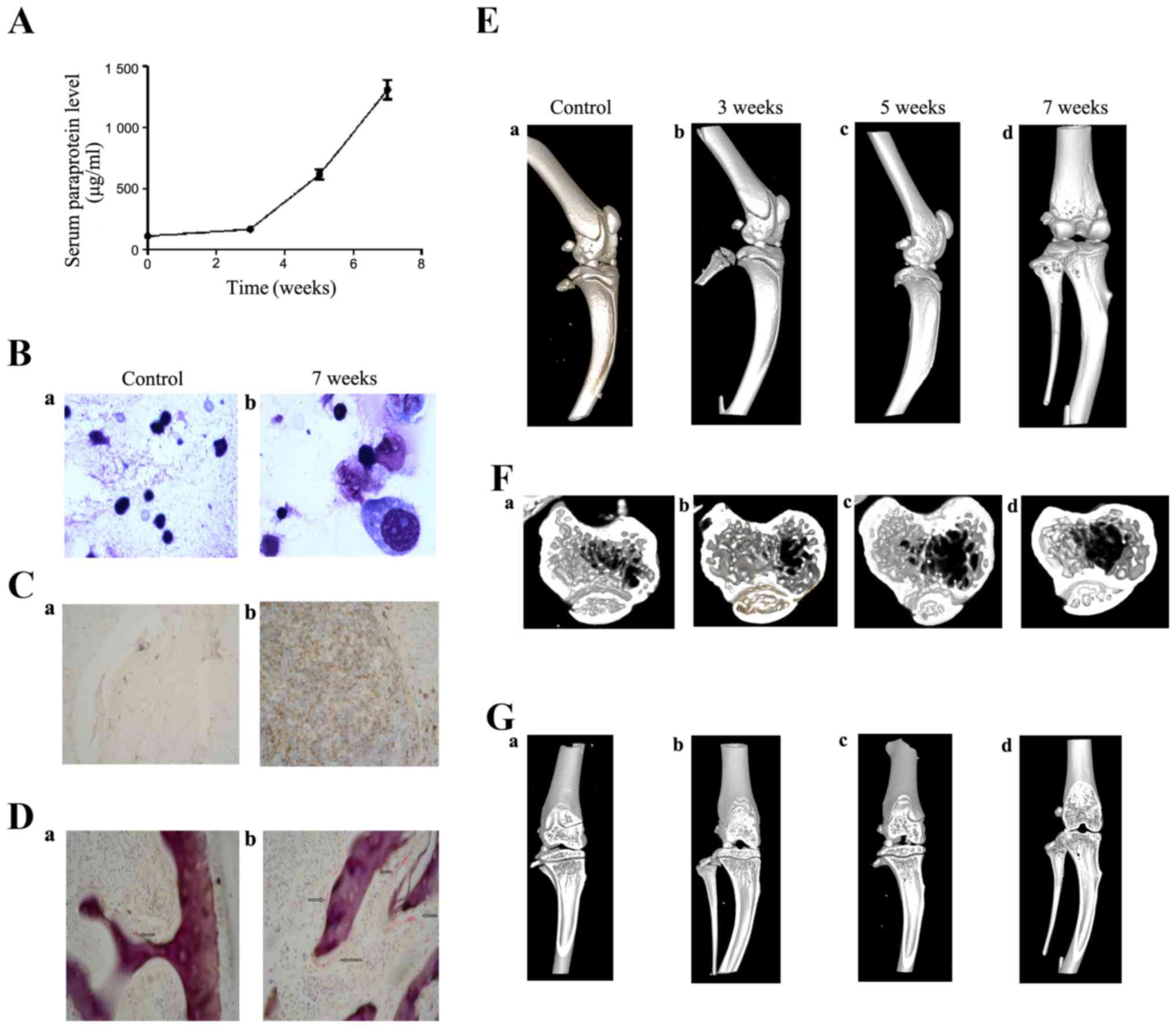
Bone disease of MM animal model
In order to evaluate the myeloma animal model, we
investigated the serum human monoclonal Igλ level by ELISA at 3, 5
and 7 weeks after myeloma cells injection. Compared with 3 weeks,
there was a corresponding increase in Igλ level. The serum human
Igλ level increased from 164.6 to 611.93 (5 weeks) and 1308.9
µg/ml (7 weeks) (Fig.
2A).
Myeloma cells in mouse bone marrow were observed by
H&E staining and immunohistochemistry (CD138) at 7 weeks. The
results showed myeloma cells in tibiae (Fig. 2B) by morphology and positive CD138
expression (Fig. 2C), which
comfirmed that human myeloma cells were injected directly into the
bone marrow of the model mice.
Furthermore, we observed bone disease by microCT at
3, 5 and 7 weeks and TRAP staining in BM at 7 weeks. TRAP staining,
which revealed increased number of osteoclasts in tibiae, was
stronger in the mouse model (Fig.
2D–b) compared with controls (Fig.
2D–a). We observed the lateral, coronal and sagittal scan of
mouse tibiae and analyzed several three-dimensional (3D) parameters
to assess the bone disease. The results showed that bone damage
existed in images from 5 weeks and became worse at 7 weeks after
injection (Fig. 2E–G). Table IV shows the three-dimensional (3D)
parameters. The total bone volume/total volume at 5 and 7 weeks
significantly increased compared with controls (P=0.041 and 0.017).
Also, trabecular bone volume at 5 and 7 weeks were also
significantly higher than controls (P=0.027 and 0.004). While other
parameters had no significant changes.
| Table IVThe three-dimensional (3D) parameters
in myeloma bone disease mouse model. |
Table IV
The three-dimensional (3D) parameters
in myeloma bone disease mouse model.
| Total bone
volume/total volume | Bone surface
area/bone volume | Trabecular
thickness | Trabecular
number | Trabecular
spacing | Trabecular bone
volume (mm3) |
|---|
| Controls | 0.5224±0.06294 |
24.5472±7.14071 | 0.0879±0.02992 | 5.4937±0.6331 | 0.0962±0.00779 | 1.533-0.20391 |
| 3 weeks | 0.4959±0.01369 |
23.8545±2.32456 | 0.0844±0.00797 | 5.9045±0.41117 | 0.0855±0.00359 | 1.3563±0.0498 |
| 5 weeks |
0.3650±0.06294a |
28.1016±1.16317 | 0.0713±0.00301 | 6.4713±0.27484 | 0.0835±0.00603 |
1.1042±0.07738a |
| 7 weeks |
0.3218±0.06155b |
33.4926±8.32048 | 0.0620±0.01372 | 5.799±0.618 | 0.1118±0.03037 |
0.7912±0.12501b |
CCN1 effect on bone remodeling in MBD
mouse model
To evaluate the efficacy of CCN1 on bone remodeling,
we established a CCN1 overexpression model of MM, in which
transduced RPMI-8226/CYR61 cells were injected directly into the
marrow tibial cavity of mice. Moreover, we used RPMI-8226/EV as
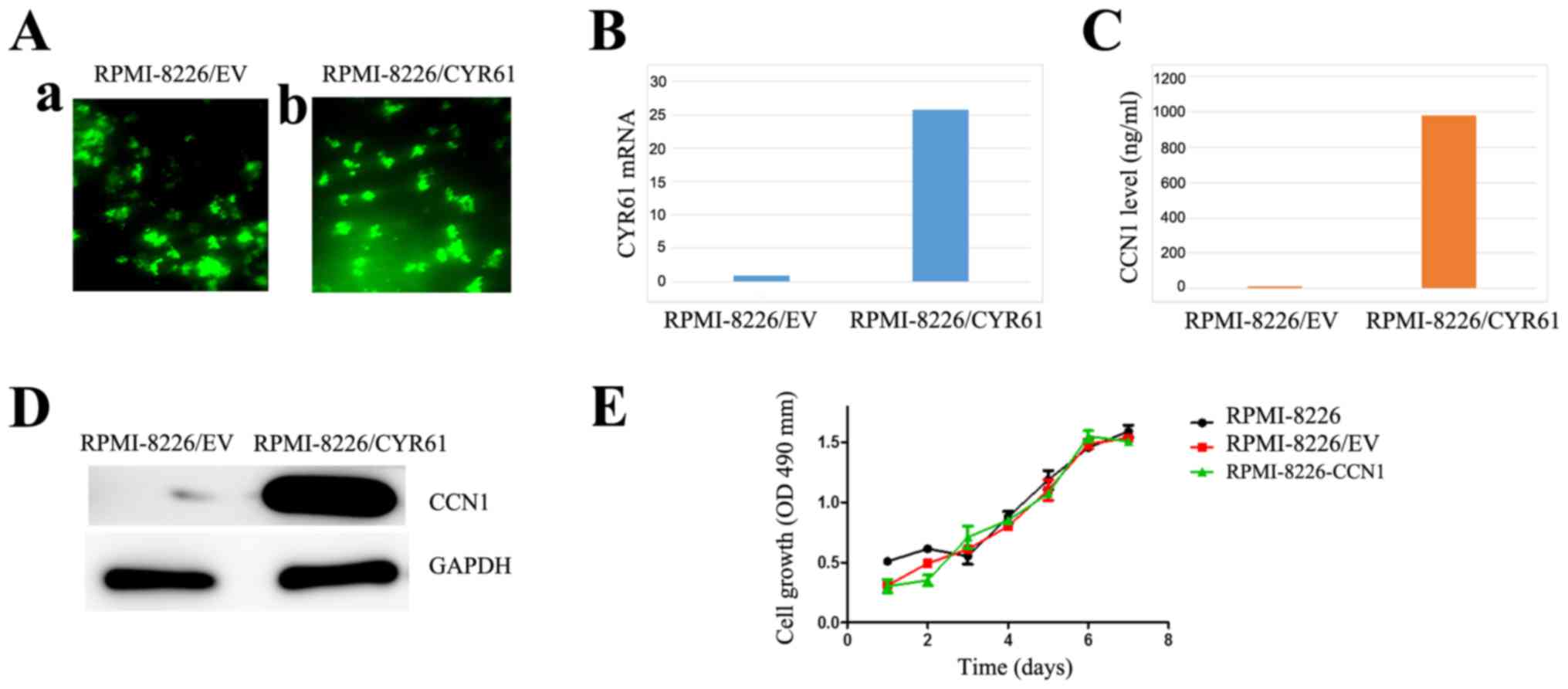
negative control and zoledronic acid as positive control. Compared
with RPMI-8226/EV, the transduced RPMI-8226/CYR61 cells expressed
CCN1 by RT-PCR, ELISA and western blotting, but the proliferation
of the cells was not affected by transduction (Fig. 3).
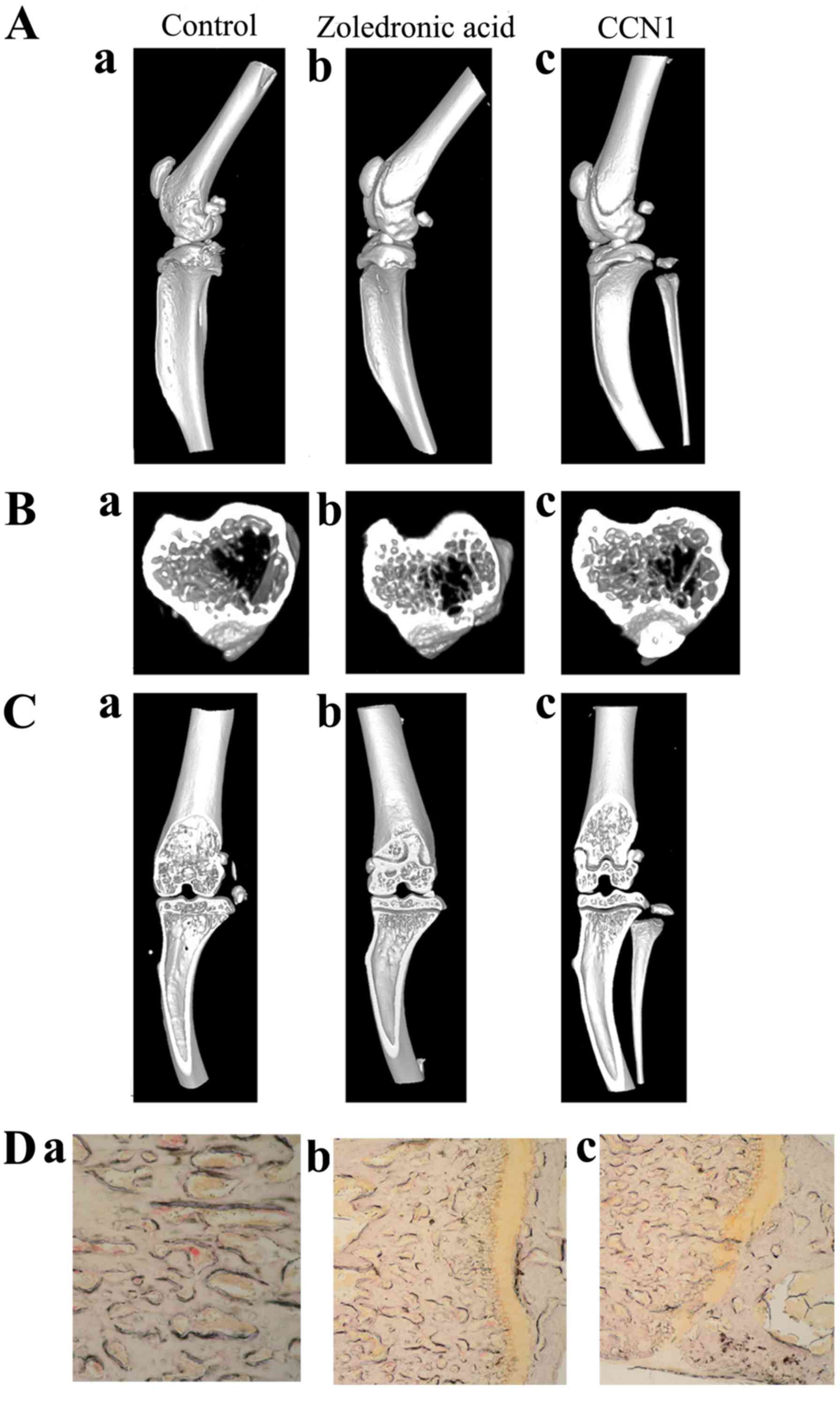
Radiographic analysis of the bones showed that bones
exhibited decreased resorption in CCN1 overexpression group and
zoledronic acid group, while severe resorption in negative control
group (Fig. 4A–C). TRAP staining
was negative in CCN1 overexpression group and zoledronic acid
group, compared with control (Fig.
4D). Table V shows that total
bone volume/total volume and trabecular bone volume in both CCN1
overexpression group and zoledronic acid group were significantly
increased compared with controls. Furthermore, trabecular bone
volume in CCN1 overexpression group was (1.7539±0.16949), which was
significantly higher than that in zoledronic acid group
(1.2839±0.077) (P=0.012).
| Table VThe three-dimensional (3D) parameters
in different groups. |
Table V
The three-dimensional (3D) parameters
in different groups.
| Total bone
volume/total volume | Bone surface
area/bone volume | Trabecular
thickness | Trabecular
number | Trabecular
spacing | Trabecular bone
volume (mm3) |
|---|
| Controls | 0.2759±0.10325 |
43.1139±12.52269 | 0.0492±0.0148 | 5.5365±0.42373 | 0.1321±0.02678 | 0.7219±0.122 |
| Zoledronic
acid |
0.4848±0.030a | 26.9035±1.582 | 0.0745±0.004 | 6.5056±0.014 | 0.0792±0.005 |
1.2839±0.077a |
| CCN1
overexpression |
0.4541±0.03383b |
25.5417±1.19068 | 0.0784±0.00367 | 5.4492±0.36592 | 0.0921±0.0078 |
1.7539±0.16949b,c |
Disscusion
MM is a malignancy of terminally differentiated
plasma cells and produces bone disease in 80% of patients (23,24).
How to cure the bone disease of MM patients is still a challenge
for the researchers and physicians. Recent studies indicated many
cytokines in BM microenvironment play a role in the pathogenesis of
malignant diseases (25,26). CYR61, which encodes the secreted
protein CCN1, plays an important role on bone remodeling in
physiological and pathological processes. We found that CCN1 levels
in BM supernatant and OBs both increased in the newly diagnosed MM
patients, especially in the patients without bone disease,
indicating CCN1 maybe play a protective role in MBD.
In order to observe the effect of CCN1, we used two
concentrations of CCN1 (10 and 30 ng/ml) to stimulate OBs, the
results showed that the quantity and mineralization ability of OBs
increased significantly at 30 ng/ml concentration. Furthermore, we
detected the mRNA levels of Runx2, β-catenin and BMP2 to evaluate
the differentiation of OBs. As known, the canonical Wnt/β-catenin
pathway is an important pathway regulating the differentiation of
OBs (27,28). BMP2 and Runx2 also take part in the
osteogenetic process. Some studies have identified that Runx2 is
one of major target gene of BMP2, which is a key regulator of
osteoblast differentiation and bone development (29–33).
In our study, the mRNA levels of β-catenin and Runx2 increased
after the CCN1 stimulation, while the BMP2 had no significant
change. Gaur et al (34)
suggested that canonical Wnt signaling promoted osteogenesis by
directly stimulating Runx2 gene expression. Therefore, the CCN1
stimulated the growth and mineralization of OBs in MBD patients
mainly via Wnt signaling pathway.
It is well known that MM is a heterogeneous
malignant disease, featured by various subtypes and different
outcomes (35). It also determines
that the characteristics of myeloma cells in different patients are
diverse. Because myeloma cells react with OBs in different ways
(36,37), the OBs in different patients maybe
exhibit diverse performances. Our results in real-time PCR are
presented as median, which perhaps support this hypothesis.
In order to investigate the role of CCN1 in bone
remodeling in vivo, we tried to build a mouse model of MBD.
There are several kinds of MBD models, including 5T2MM model
(38), MOPC35.BM model (39) and transgenic mouse models
(Myc/Bcl-XL mice) (40). However,
these models are made by injection or transgene of mouse myeloma
cells, which can not recapitulate the human condition of BM
microenvironment and bone disease. Considering the availability,
technology and cost, we injected the human myeloma cells
(RPMI-8226) to mice, similar to Labrinidis et al (19). We detected the myeloma cells in
mouse bone marrow by H&E staining and immunochemistry staining
(CD138), which indicated the human myeloma cells grew in the mice.
Furthermore, we observed the bone lesions by microCT and TRAP
staining to identify the bone disease.
In this mouse model, overexpressed CCN1 in engrafted
RPMI-8226 cells resulted in reduced bone disease, suggesting that
CCN1 plays a positive effect on progression of bone lesions. Our
results also indicated that the effect of CCN1 may be better than
zoledronic acid, a first line treatment of MBD (41).
In conclusion, CCN1 is overexpressed in BM
supernatant and OB in newly diagnosed myeloma patients, especially
in the patients without bone disease, which could stimulate the
proliferation and differentiation of OBs in vitro and
contribute to bone remodeling in vivo. However, we still
need to deeply investigate the signaling pathway of CCN1 on OBs to
explore its mechanism.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 81570106 and 81400088,
81600093), the anticancer major special project of Tianjin (grant
no. 12ZCDZSY18000), the Tianjin Municipal Natural Science
Foundation (grant nos. 14JCYBJC25400 and 15JCYBJC24300), Tianjin
Health and Family Planning Commision (grant no. 15KG150) and the
Science and Technology Foundation of Tianjin Municipal Health
Bureau (grant nos. 2011kz115 and 2014KZ120).
References
|
1
|
Anderson KC: Oncogenomics to target
myeloma in the bone marrow microenvironment. Clin Cancer Res.
17:1225–1233. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yaccoby S: Osteoblastogenesis and tumor
growth in myeloma. Leuk Lymphoma. 51:213–220. 2010. View Article : Google Scholar :
|
|
3
|
Vallet S, Smith MR and Raje N: Novel
bone-targeted strategies in oncology. Clin Cancer Res.
16:4084–4093. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boyce BF and Xing L: The RANKL/RANK/OPG
pathway. Curr Osteoporos Rep. 5:98–104. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vallet S, Pozzi S, Patel K, Vaghela N,
Fulciniti MT, Veiby P, Hideshima T, Santo L, Cirstea D, Scadden DT,
et al: A novel role for CCL3 (MIP-1α) in myeloma-induced bone
disease via osteocalcin downregulation and inhibition of osteoblast
function. Leukemia. 25:1174–1181. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fu R, Liu H, Zhao S, Wang Y, Li L, Gao S,
Ruan E, Wang G, Wang H, Song J, et al: Osteoblast inhibition by
chemokine cytokine ligand3 in myeloma-induced bone disease. Cancer
Cell Int. 12:1322014. View Article : Google Scholar
|
|
7
|
Scullen T, Santo L, Vallet S, Fulciniti M,
Eda H, Cirstea D, Patel K, Nemani N, Yee A, Mahindra A, et al:
Lenalidomide in combination with an activin A-neutralizing
antibody: Preclinical rationale for a novel anti-myeloma strategy.
Leukemia. 27:1715–1721. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kristensen IB, Christensen JH, Lyng MB,
Møller MB, Pedersen L, Rasmussen LM, Ditzel HJ and Abildgaard N:
Hepatocyte growth factor pathway upregulation in the bone marrow
microenvironment in multiple myeloma is associated with lytic bone
disease. Br J Haematol. 161:373–382. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hiasa M, Teramachi J, Oda A, Amachi R,
Harada T, Nakamura S, Miki H, Fujii S, Kagawa K, Watanabe K, et al:
Pim-2 kinase is an important target of treatment for tumor
progression and bone loss in myeloma. Leukemia. 29:207–217. 2015.
View Article : Google Scholar
|
|
10
|
Johnson SK, Stewart JP, Bam R, Qu P,
Barlogie B, van Rhee F, Shaughnessy JD Jr, Epstein J and Yaccoby S:
CYR61/CCN1 overexpression in the myeloma microenvironment is
associated with superior survival and reduced bone disease. Blood.
124:2051–2060. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Perbal B: CCN proteins: Multifunctional
signalling regulators. Lancet. 363:62–64. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kireeva ML, Mo FE, Yang GP and Lau LF:
Cyr61, a product of a growth factor-inducible immediate-early gene,
promotes cell proliferation, migration, and adhesion. Mol Cell
Biol. 16:1326–1334. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Clines GA, Mohammad KS, Bao Y, Stephens
OW, Suva LJ, Shaughnessy JD Jr, Fox JW, Chirgwin JM and Guise TA:
Dickkopf homolog 1 mediates endothelin-1-stimulated new bone
formation. Mol Endocrinol. 21:486–498. 2007. View Article : Google Scholar :
|
|
14
|
Si W, Kang Q, Luu HH, Park JK, Luo Q, Song
WX, Jiang W, Luo X, Li X, Yin H, et al: CCN1/Cyr61 is regulated by
the canonical Wnt signal and plays an important role in
Wnt3A-induced osteoblast differentiation of mesenchymal stem cells.
Mol Cell Biol. 26:2955–2964. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Su JL, Chiou J, Tang CH, Zhao M, Tsai CH,
Chen PS, Chang YW, Chien MH, Peng CY, Hsiao M, et al: CYR61
regulates BMP-2-dependent osteoblast differentiation through alpha
v beta 3 integrin/ILK/ERK pathway. J Biol Chem. 285:31325–31326.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen CY, Su CM, Huang YL, Tsai CH, Fuh LJ
and Tang CH: CCN1 induces oncostatin M production in osteoblasts
via integrin-dependent signal pathways. PLoS One. 9:e1066322014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Terpos E, de la Fuente J, Szydlo R,
Hatjiharissi E, Viniou N, Meletis J, Yataganas X, Goldman JM and
Rahemtulla A: Tartrate-resistant acid phosphatase isoform 5b: A
novel serum marker for monitoring bone disease in multiple myeloma.
Int J Cancer. 106:455–457. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li X, Pennisi A and Yaccoby S: Role of
decorin in the antimyeloma effects of osteoblasts. Blood.
112:159–168. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Labrinidis A, Diamond P, Martin S, Hay S,
Liapis V, Zinonos I, Sims NA, Atkins GJ, Vincent C, Ponomarev V, et
al: Apo2L/TRAIL inhibits tumor growth and bone destruction in a
murine model of multiple myeloma. Clin Cancer Res. 15:1998–2009.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schwarzer R, Nickel N, Godau J, Willie BM,
Duda GN, Schwarzer R, Cirovic B, Leutz A, Manz R, Bogen B, et al:
Notch pathway inhibition controls myeloma bone disease in the
murine MOPC315.BM model. Blood Cancer J. 4:e2172014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kristensen IB, Christensen JH, Lyng MB,
Møller MB, Pedersen L, Rasmussen LM, Ditzel HJ and Abildgaard N:
Expression of osteoblast and osteoclast regulatory genes in the
bone marrow microenvironment in multiple myeloma: Only
up-regulation of Wnt inhibitors SFRP3 and DKK1 is associated with
lytic bone disease. Leuk Lymphoma. 55:911–919. 2014. View Article : Google Scholar
|
|
22
|
Silvestris F, Cafforio P, De Matteo M,
Calvani N, Frassanito MA and Dammacco F: Negative regulation of the
osteoblast function in multiple myeloma through the repressor gene
E4BP4 activated by malignant plasma cells. Clin Cancer Res.
14:6081–6091. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kristinsson SY, Minter AR, Korde N, Tan E
and Landgren O: Bone disease in multiple myeloma and precursor
disease: Novel diagnostic approaches and implications on clinical
management. Expert Rev Mol Diagn. 11:593–603. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Giuliani N, Rizzoli V and Roodman GD:
Multiple myeloma bone disease: Pathophysiology of osteoblast
inhibition. Blood. 108:3992–3996. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Olechnowicz SW and Edwards CM:
Contributions of the host microenvironment to cancer-induced bone
disease. Cancer Res. 74:1625–1631. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Andrews SW, Kabrah S, May JE, Donaldson C
and Morse HR: Multiple myeloma: The bone marrow microenvironment
and its relation to treatment. Br J Biomed Sci. 70:110–120. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Glass DA II, Bialek P, Ahn JD, Starbuck M,
Patel MS, Clevers H, Taketo MM, Long F, McMahon AP, Lang RA, et al:
Canonical Wnt signaling in differentiated osteoblasts controls
osteoclast differentiation. Dev Cell. 8:751–764. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hill TP, Später D, Taketo MM, Birchmeier W
and Hartmann C: Canonical Wnt/β-catenin signaling prevents
osteoblasts from differentiating into chondrocytes. Dev Cell.
8:727–738. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nishio Y, Dong Y, Paris M, O'Keefe RJ,
Schwarz EM and Drissi H: Runx2-mediated regulation of the zinc
finger Osterix/Sp7 gene. Gene. 372:62–70. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bae JS, Gutierrez S, Narla R, Pratap J,
Devados R, van Wijnen AJ, Stein JL, Stein GS, Lian JB and Javed A:
Reconstitution of Runx2/Cbfa1-null cells identifies a requirement
for BMP2 signaling through a Runx2 functional domain during
osteoblast differentiation. J Cell Biochem. 100:434–449. 2007.
View Article : Google Scholar
|
|
31
|
Maruyama Z, Yoshida CA, Furuichi T,
Amizuka N, Ito M, Fukuyama R, Miyazaki T, Kitaura H, Nakamura K,
Fujita T, et al: Runx2 determines bone maturity and turnover rate
in postnatal bone development and is involved in bone loss in
estrogen deficiency. Dev Dyn. 236:1876–1890. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jeon EJ, Lee KY, Choi NS, Lee MH, Kim HN,
Jin YH, Ryoo HM, Choi JY, Yoshida M, Nishino N, et al: Bone
morphogenetic protein-2 stimulates Runx2 acetylation. J Biol Chem.
281:16502–16511. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang S, Wei D, Wang D, Phimphilai M,
Krebsbach PH and Franceschi RT: In vitro and in vivo synergistic
interactions between the Runx2/Cbfa1 transcription factor and bone
morphogenetic protein-2 in stimulating osteoblast differentiation.
J Bone Miner Res. 18:705–715. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gaur T, Lengner CJ, Hovhannisyan H, Bhat
RA, Bodine PV, Komm BS, Javed A, van Wijnen AJ, Stein JL, Stein GS,
et al: Canonical WNT signaling promotes osteogenesis by directly
stimulating Runx2 gene expression. J Biol Chem. 280:33132–33140.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Szalat R and Munshi NC: Genomic
heterogeneity in multiple myeloma. Curr Opin Genet Dev. 30:56–65.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Colucci S, Brunetti G, Oranger A, Mori G,
Sardone F, Specchia G, Rinaldi E, Curci P, Liso V, Passeri G, et
al: Myeloma cells suppress osteoblasts through sclerostin
secretion. Blood Cancer J. 1:e272011. View Article : Google Scholar
|
|
37
|
Giuliani N, Colla S, Morandi F, Lazzaretti
M, Sala R, Bonomini S, Grano M, Colucci S, Svaldi M and Rizzoli V:
Myeloma cells block RUNX2/CBFA1 activity in human bone marrow
osteoblast progenitors and inhibit osteoblast formation and
differentiation. Blood. 106:2472–2483. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vanderkerken K, De Raeve H, Goes E, Van
Meirvenne S, Radl J, Van Riet I, Thielemans K and Van Camp B: Organ
involvement and phenotypic adhesion profile of 5T2 and 5T33 myeloma
cells in the C57BL/KaLwRij mouse. Br J Cancer. 76:451–460. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hofgaard PO, Jodal HC, Bommert K, Huard B,
Caers J, Carlsen H, Schwarzer R, Schünemann N, Jundt F, Lindeberg
MM, et al: A novel mouse model for multiple myeloma (MOPC315.BM)
that allows noninvasive spatiotemporal detection of osteolytic
disease. PLoS One. 7:e518922012. View Article : Google Scholar
|
|
40
|
Cheung WC, Kim JS, Linden M, Peng L, Van
Ness B, Polakiewicz RD and Janz S: Novel targeted deregulation of
c-Myc cooperates with Bcl-X (L) to cause plasma cell neoplasms in
mice. J Clin Invest. 113:1763–1773. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Morgan GJ, Davies FE, Gregory WM, Cocks K,
Bell SE, Szubert AJ, Navarro-Coy N, Drayson MT, Owen RG, Feyler S,
et al National Cancer Research Institute Haematological Oncology
Clinical Study Group: First-line treatment with zoledronic acid as
compared with clodronic acid in multiple myeloma (MRC Myeloma IX):
A randomised controlled trial. Lancet. 376:1989–1999. 2010.
View Article : Google Scholar : PubMed/NCBI
|