Introduction
Lung cancer is the most frequently diagnosed cancer
in men and the third most commonly diagnosed cancer in women
throughout the world (1). Eighty
to eighty-five percent of lung cancer are non-small cell lung
cancer (NSCLCs) (2). The majority
of lung cancer (57%) are diagnosed at advanced stage. Despite
decades of bench and clinical research to attempt to improve
outcome for NSCLC, the 5-year overall rate remains only ~21%
(3). The existing therapies,
including radiotherapy, chemotherapy and the emerging target
therapy, are still unsatisfactory to improve outcomes for lung
cancer patients (4). It is an
urgent issue to achieve breakthrough in medical treatment on lung
cancer.
Tumor microenvironment has attracted much attention
in recent years. Macrophages are the main non-cancer cell
components in tumor microenvironment, and often described as
tumor-associated macrophages (TAMs) (5). Macrophages originate from blood
monocytes, and there are two main classes of macrophages:
classically activated macrophages (also called M1) and
alternatively activated macrophages (M2) (6). When exposed to lipopolysaccharide
(LPS) or interferon-γ (IFN-γ), macrophages are polarized into M1
macrophages; whereas interleukin-4 (IL-4) or interleukin-13 (IL-13)
exposure can polarize them into M2 macrophages (7). M1 macrophages generally exhibit
microbicidal activity and have pro-inflammatory phenotype. In
contrast, M2 macrophages are able to tune inflammatory responses
and adaptive Th2 immunity (8).
Solid tumors are often infiltrated with abundant macrophages, which
resembles M2 macrophages that can produce high amounts of
interleukin (IL)-10 and transforming growth factor-β (TGF-β),
express scavenger receptors (CD204) and mannose receptor (CD206),
and exhibit anti-inflammatory and tissue repair functions (9).
TAMs play an essential role in the complex process
for coevolution and tumorigenesis in tumor microenvironment. Lee
et al showed that IL-10 mediated M2-polarized TAMs promoted
renal cell carcinoma (RCC) growth through increasing BMP-6
expression (10). TLR4/IL-10
signaling increased the proliferation of cancer cells induced by
M2-polarized TAMs, while TLR4 siRNA or neutralizing antibodies of
TLR4 and IL-10 significantly reversed the increased growth
(11). TAMs and their secreted
cytokines (eg., IL-10, TNF-α, TGF-β) or proteases were able to
promote the migration and invasion of cancer cells in all aspects
(12,13). Previous studies revealed that
M2-polarized macrophages induced migration and invasion of human
basal carcinoma cells and prostate cancer cells (14,15).
TAMs also promoted tumor angiogenesis, proliferation and invasion
in pancreatic cancer, and hepatocellular carcinoma (16-19).
Indeed, removal of macrophages by genetic mutation, ablation of
macrophage function or inhibition of macrophage infiltration into
tumors successfully reduced tumor progression and metastasis in
human breast carcinomas, and pulmonary metastatic cells (20,21).
The establishment of macrophages contributed to progress of
research to develop therapies that specifically target macrophages.
These possible therapeutics are divided into three main aspects:
inhibition of monocyte recruitment into tumors, eradication of
resident macrophages already in tumors, and neutralization of key
molecules of TAM release, such as CCL5 and VEGF (8,22–25).
However, most of these therapeutic methods are still at the
research stage.
In addition, IL-10 may promote cancer growth by
suppressing macrophage function and enabling tumors to evade
immunosurveillance (26). Kuang
and his colleagues (27) observed
that IL-10 secreted from activated monocytes strongly induced the
expression of programmed cell death 1 (PD-L1) in an autocrine
manner. PD-L1-positive monocytes induced T cell dysfunction, as
defined by the presence of low cytotoxicity to tumor cells and a
reduction in T cell proliferation targeted therapy aiming at IL-10
or PD-L1 may be an effective method for NSCLC. Whether IL-10
secreted from TAMs could regulate the expression of PD-L1 in NSCLC
cells by paracrine way and weather BFD can interfere with the
interaction between TAMs and tumor cells is still unknown.
Bu Fei decoction (BFD) is a classical formula of
traditional Chinese medicine (TCM) in clinical setting for hundreds
of years. It consists of six herbal Chinese medicines including
Codonopsis pilosula, Schisandra chinensis, Rehmannia
glutinosa, Astragalus, Aster and Cortex Mori. BFD is a
mixture of these herbal Chinese medicines in a certain proportion.
BFD is widely used for improving lung functions, especially for the
Qi deficiency. So it is to alleviate lung cancer related
symptoms in oncology clinical context and has a good clinical
effect. The team of Xia et al (28,29)
have devoted research effort on BFD on lung tissue of pulmonary
fibrosis. They found that BFD could decrease the degree of
alveolitis and pulmonary fibrosis decreased significantly, which
was associated with reduced expression of α-SMA, MMP-9 and TIMP-1
and re-established the balance among these, then improved the
abnormal metabolism of extracellular matrix. Nevertheless, there
has been until now no systematic research to study BFD as a whole
in lung cancer. In recent years, several studies found that many
components of BFD possess antitumor effect. Research data showed
that herbal extract from Astragalus could induce dendritic cell
maturation, promote IFN-γ secretion and activate the antitumor
effect of special cytotoxic T lymphocytes (CTLs) (30). Some research data show that
Schisandrin B, one of the bioactive compounds from Schisandra
chinensis, could suppress epithelial mesenchymal transform
(EMT) in breast cancer both in vitro and in vivo,
scavenged oxygen free radicals and inhibited the expression of VEGF
(31,32). However, nothing is known on whether
BFD possesses antitumor effect or it could influence tumor
microenvironment in lung cancer. Therefore, the aim of the present
study was to investigate the effects of BFD on TAMs and NSCLC in
vitro and in vivo, to possible clarify underlying
mechanisms.
Materials and methods
Preparation of drugs
The component herbs of BFD used in this study are as
follows, Codonopsis pilosula, Schisandra chinensis, Rehmannia
glutinosa, Astragalus, Aster and Cortex Mori, at a ratio
of 3:2:1.8:4:2:2. Furthermore, Schisandra chinensis was
treated with vinegar. Radix Rehmanniae Praeparata was used
in the prescription. Aster is a honey-fried Chinese herb. The BFD
granules used in the present study were purchased from Beijing
Tcmages Pharmaceutical Co., Ltd. (Beijing, China). The quality of
formula granules was monitored by Fourier transform infrared
spectroscopy (FTIR) (Model IRPRestige-21; Shiamdzu Corp., Kyoto,
Japan). Prior to experiment, the formula granules were milled to
powders and dissolved in distilled water to get a concentration of
1 g/ml crude drug, and then filtered through a 0.22-µm
membrane (Millipore, Billerica, MA, USA). The concentrations of BFD
in the study refer to the crude drug concentrations.
Reagents
Phorbol 12-myristate 13-acetate (PMA) and 3-(4,
5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2-H-tetrazolium bromide (MTT)
was purchased from Sigma-Aldrich (St. Louis, MO, USA). Recombinant
human IL-4 was purchased from R&D Systems (Minneapolis, MN,
USA). Human Fc receptor binding inhibitor purified and anti-human
IL-10 for neutralization were purchased from eBioscience (San
Diego, CA, USA). Recombinant human IL-10 was purchased from
Peprotech (Rocky Hill, NJ, USA). Antibodies and their respective
sources were as follows: anti FITC mouse anti-human CD206 antibody
(BD Pharmingen, 551135), anti-mannose receptor antibody (Abcam,
ab64693), anti-PD-L1 antibody (Abcam, ab58810), anti-IL-10 antibody
(Abcam, ab9969).
Cell culture and differentiation
Human NSCLC cell lines A549 and NCI-H1975 were
purchased from the American Type Culture Collection (Manassas, VA,
USA). Human monocytic leukemia U937 cells (RIKEN, RCB0435) were
donated by Professor C. Wang, Peking University Health Science
Center. Cells were maintained in RPMI-1640 supplemented with 10%
fetal bovine serum (FBS) (Gibco), 100 U/ml penicillin, and 100
µg/ml streptomycin at 37°C and 5% CO2.
U937 cells were seeded at a density of
5×105 cells/well into 10-cm culture dish with RPMI-1640
containing 10% FBS and 100 ng/ml PMA. After 72 h of culture, the
cells were washed with phosphate-buffered saline (PBS, pH 7.2) and
incubated with IL-4 (20 ng/ml) for the indicated times. Then cells
were washed with PBS and incubated with serum-free 1640 for 24 h.
After 24-h serum starvation, cells were collected and used as
M2-polarized TAMs, and the medium was collected and prepared as
M2-polarized TAM-conditioned medium (TAM-CM). The supernatants and
cells were collected and stored at −80°C until use.
Morphological characterization and flow
cytometry
Morphological characterization of U937 cells and
PMA-IL-4-treated U937 cells was acquired by phase-contrast
microscopy. Cells were washed with PBS and resuspended in PBS.
Cells were then incubated in Clear Back (human Fc receptor blocking
reagent) (MBL, Nagoya, Japan) for 20 min followed by incubation
with FITC-mouse anti-human CD206 (BD Biosciences) for 60 min on ice
in the dark. After the final washing step, labeled cells were
analyzed by flow cytometry (BD Accuri C6).
Cell viability assay
5-diphenyltetrazolium bromide (MTT) assay was used
to measure cell viability. Briefly, cells were plated in 96-well
culture plates at the density of 5–10×103 per well in
complete medium. After 24-h incubation, cells were treated with BFD
(0–80 mg/ml) in serum-free medium or TAM-CM for 24 h. Then the
cells were incubated with 100 µl 0.5 mg/ml MTT at 37°C for 4
h, and the precipitate was dissolved in 150 µl
dimethylsulfoxide (DMSO). After shaking for 10 min, the optical
density at 570 nm was measured and the IC50 value was
calculated on the non-linear regression fit method by Graphpad
Prism 5.0 software (San Diego, CA, USA).
Transwell assay
Cell migration and invasion were performed in
24-well plates using transwell chambers (Corning Inc., Corning, NY,
USA) fitted with a polyethylene terephthalate filter membrane with
8-µm pores. Cells (5×104) were seeded in
serum-free medium in the upper chamber (normal chamber for
migration assay and matrigel-coated chamber for invasion assay).
The lower chamber was filled with medium containing 10 or 20% FBS.
After incubating for 24 h at 37°C, cells in the upper chamber were
carefully removed with a cotton swab, and the cells that had
traversed to reverse face of the membrane were cleaned and fixed in
methanol, stained with crystal violet. Images were taken with a
microscope and cells were counted under 5 randomly chosen
fields.
RNA extraction and quantitative real-time
PCR
Total RNA was extracted from cells using TRIzol
reagent according to the manufacturer's instructions (Invitrogen
Life Technologies). The concentration and quality of the extracted
RNA were measured with a NanoDrop 2000 (Thermo Fisher Scientific,
Wilmington, DE, USA). The first-strand cDNA was generated using the
TransScript first-strand cDNA synthesis supermix (Transgen,
Beijing, China) according to the manufacturer's instructions.
Primers designed for PCR were synthesized by Sangon Biotech Co.,
Ltd. (Shanghai, China) and are shown below. The RT-PCR assay was
performed using SYBR green qPCR supermix (Applied Biosystems Life
Technologies, Foster City, CA, USA) in an ABI PRISM 7500 sequence
detection system (Applied Biosystems Life Technologies). The PCR
specific primers were 5′-GAC TTT AAG GGT TAC CTG GGT TG-3′
(forward) and 5′-TCA CAT GCG CCT TGA TGT CTG-3′ (reverse) for
IL-10, 5′-GGA CAA GCA GTG ACC ATC AAG-3′ (forward) and 5′-CCC AGA
ATT ACC AAG TGA GTC CT-3′ (reverse) for PD-L1, 5′-GGT GAA GGT CGG
TGT GAA CG-3′ (forward) and 5′-CTC GCT CCT GGA AGA TGG TG-3′
(reverse) for GADPH. The conditions as shown below: 55°C for 10
min, followed by 40 cycles of 95°C for 30 sec, 55–59°C 30 sec and
72°C for 42 sec. The fold changes of each gene were calculated
using the ∆∆Ct (cycle threshold) method, and gene expression levels
were normalized by GADPH.
Western blotting
The cells were plated in dishes at a density of
5×105 cells/ml, which were treated with BFD (0–80 mg/ml)
in serum-free medium or TAM-CM for 48 h. After the treatment, the
cells were washed in PBS and lysed with RIPA lysis buffer (CWBio,
Beijing, China). Total protein of tumor tissues was extracted as
above. Protein concentrations were determined using the BCA protein
assay kit (Thermo Fisher Scientific, Rockford, IL, USA). The
samples corresponding to 20 µg of protein were boiled for
8–10 min, resolved on an 8–15% denatured SDS polyacrylamide gel,
and transferred onto a PVDF membrane (Millipore, Bedford, MA, USA).
After blocking non-specific binding sites for 1 h using 5% skim
milk, the membranes were incubated with specific antibodies
overnight at 4°C. The membranes were washed with TBST and incubated
with a peroxidase-conjugated secondary antibody for 1 h.
Visualization of the protein bands was accomplished using an
Immobilon Western Chemiluminescent HRPS substrate (Millipore).
ImageJ software was used to calculate the expression of each
protein, which was normalized by β-actin.
Regulation of PD-L1 expression
In order to verify whether it was IL-10 in TAMs that
regulated the expression of PD-L1 in NSCLC cells. TAM-CM with
neutralizing mAbs against IL-10 (eBioscience) or recombinant human
(rh) cytokines IL-10 was used instead of TAM-CM in the culture
system, and PD-L1 expression was detected by western blotting as
mentioned above. The level of IL-10 in culture supernatants was
detected by ELISA, and concrete implementation method was in
accordance with manufacturers specifications.
Determination of antitumor effect in nude
mice
Female BALB/c nude mice (6–8-week-old) were obtained
from Peking University Health Science Center (Beijing, China). The
animals were kept under a specific pathogen-free (SPF) condition
with a 12-h light/dark cycle and freely accessed autoclaved
standard food and water. The animal experiment protocol was
approved by Peking University Animals Research Committee and
conducted in accordance with European community guide lines. Human
NSCLC cell lines A549 (3×106) or H1975
(3×106) cells were suspended in 0.25 ml of PBS mixed 1:1
with Matrigel (BD Biosciences) and subcutaneously inoculated into
the right flank of nude mice.
The daily dosage for nude mice was obtained based on
the daily dosage for humans at 0.58 g/kg (equivalent to a crude
drug dose of 1.23 g/kg) in clinical. After calculation using the
human-mouse transfer formula, the dosage of BFD for nude mice was
determined to be 30 and 60 g/kg, which were 2 and 4 times the
anticipated clinical dosage, respectively. The human-mouse transfer
formula: Animal dose = Human dose ×
(HKm/AKm), where HKm and
AKm represent the Km factor of human (Human
Km: 37) and mouse (Mouse Km: 3) (33).
Tumor growth was measured with tumor volume by the
following formula: tumor volume (mm3) =1/2 × length ×
width2. When the tumors reached a size range of 50–100
mm3, the mice were randomly divided into 3 groups
(n=8–10): control, BFD (30 g/kg) and BFD (60 g/kg). All the
compounds were administered orally twice a day for consecutive 21
days, and then the tumor volume and body weight of the mice were
measured every 3 days during the administration period.
At the end of study, blood was gained from the
eyeballs of mice, and their tumors were removed and weighed after
the animals were sacrificed. Tumor tissues were fixed in formalin
for immunohistochemistry analysis or stored at −80°C for protein
election.
Statistical analysis
The data were presented as the means ± SE of at
least three separate experiments. All comparisons were analyzed
with Student's two-tailed t-test, with p<0.05 considered
statistically significant.
Results
Successful induction of M2-polarized
TAMs
As shown in Fig.
1A, compared with native cells, PMA-IL-4 treated U937 cells
attached to the culture dish and developed elongated
macrophage-like projections. Western blotting and flow cytometry
results (Fig. 1B and C) showed
that the mannose receptor CD206 was significantly higher than
native cells (p<0.05). The mRNA and protein expression levels of
IL-10 were obviously increased in PMA-IL-4 treated U937 cells
(p<0.05) (Fig. 1C–E). The above
results suggested that IL-4 (20 ng/ml) and PMA (100 ng/ml)
successfully induced the alteration of classical macrophages to
M2-polarized TAMs.
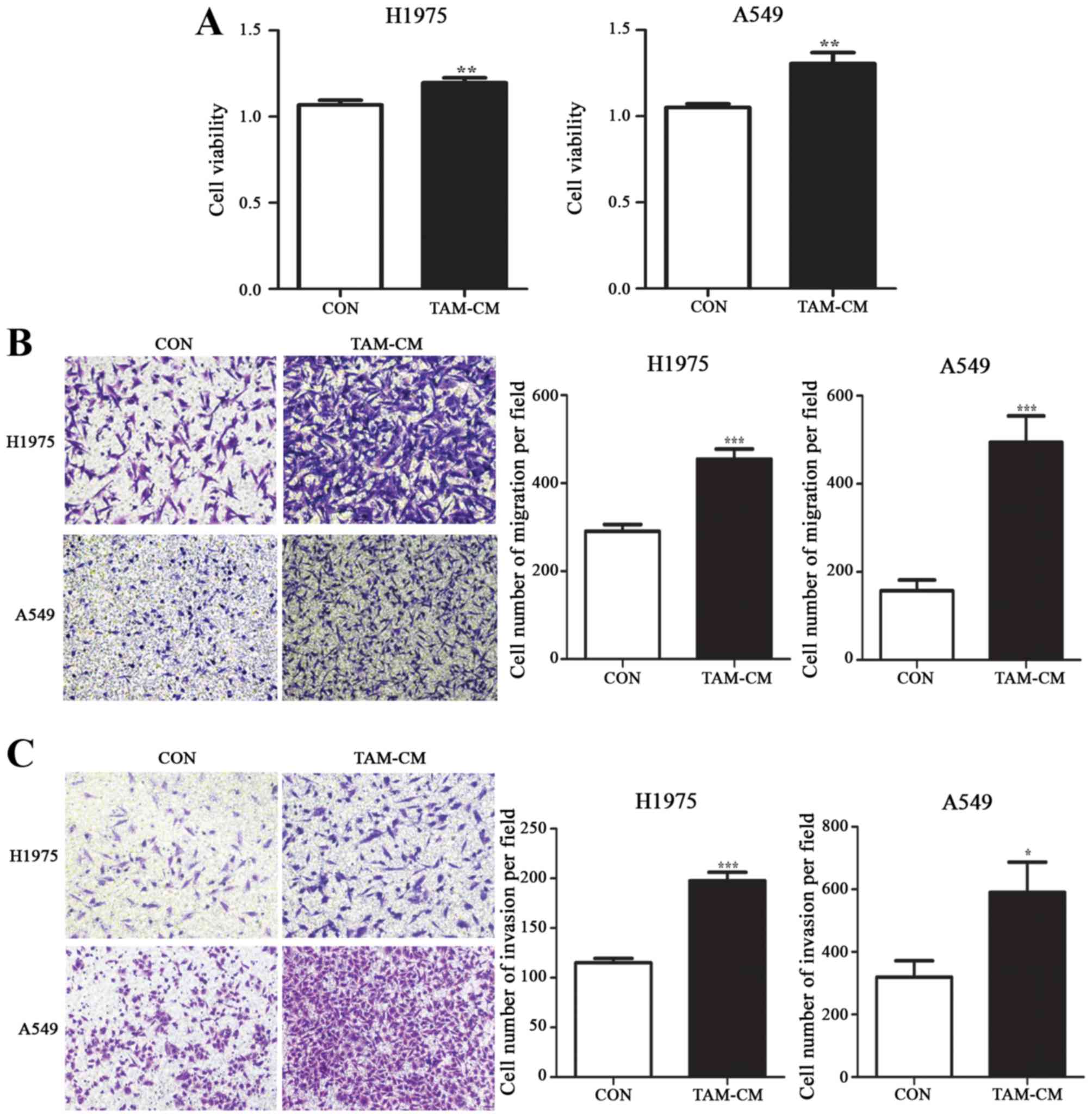
Coculture with M2-polarized TAMs
increased the proliferation, migration and invasion of NSCLC
cells
The coculture system between TAMs and NSCLC cells
was established by culturing cancer cells in M2-polarized
TAM-conditioned medium (TAM-CM). We next examined the effects of
TAMs on NSCLC cell proliferation, migration and invasion. Compared
with routine medium, coculture with TAMs significantly increased
the cell viability in both H1975 and A549 cells. Cell survival
rates were increased by 11.9% in H1975 and 24.3% in A549, p<0.01
(Fig. 2A).
TAMs significantly promoted more H1975 and A549 cell
migration to the lower chamber. Compared with the control group,
the number of migrated cells in TAM-CM was 454.5±23.49 and
494.1±60.18 in H1975 and A549 cells, respectively; whereas the
corresponding cell number was 290.9±15.2 and 157.4±24.22, in the
control group (p<0.001) (Fig.
2B).
The invasive ability of cancer cells in TAM-CM
underwent similar alterations as the migratory ability. The number
of invaded cells in TAM-CM was 197.6±8.617 and 590.3±96.52 in H1975
and A549 cells, compared to the cells in routine medium, with the
invaded cell number of 115.0±4.402 and 319.6±52.36, respectively
(p<0.05) (Fig. 2C).
Dose-dependent suppression of NSCLC cell
proliferation, migration and invasion by BFD in TAM conditioned
medium
NSCLC cell proliferation was reduced by BFD in a
dose-dependent manner, especially in H1975 cells with an
IC50 of 22.84 mg/ml in routine medium and 49.36 mg/ml in
TAM-CM. The inhibition of A549 cells was relatively weak, the
IC50 was 75.80 mg/ml in routine medium and 73.62 mg/ml
in TAM-CM (Fig. 3A).
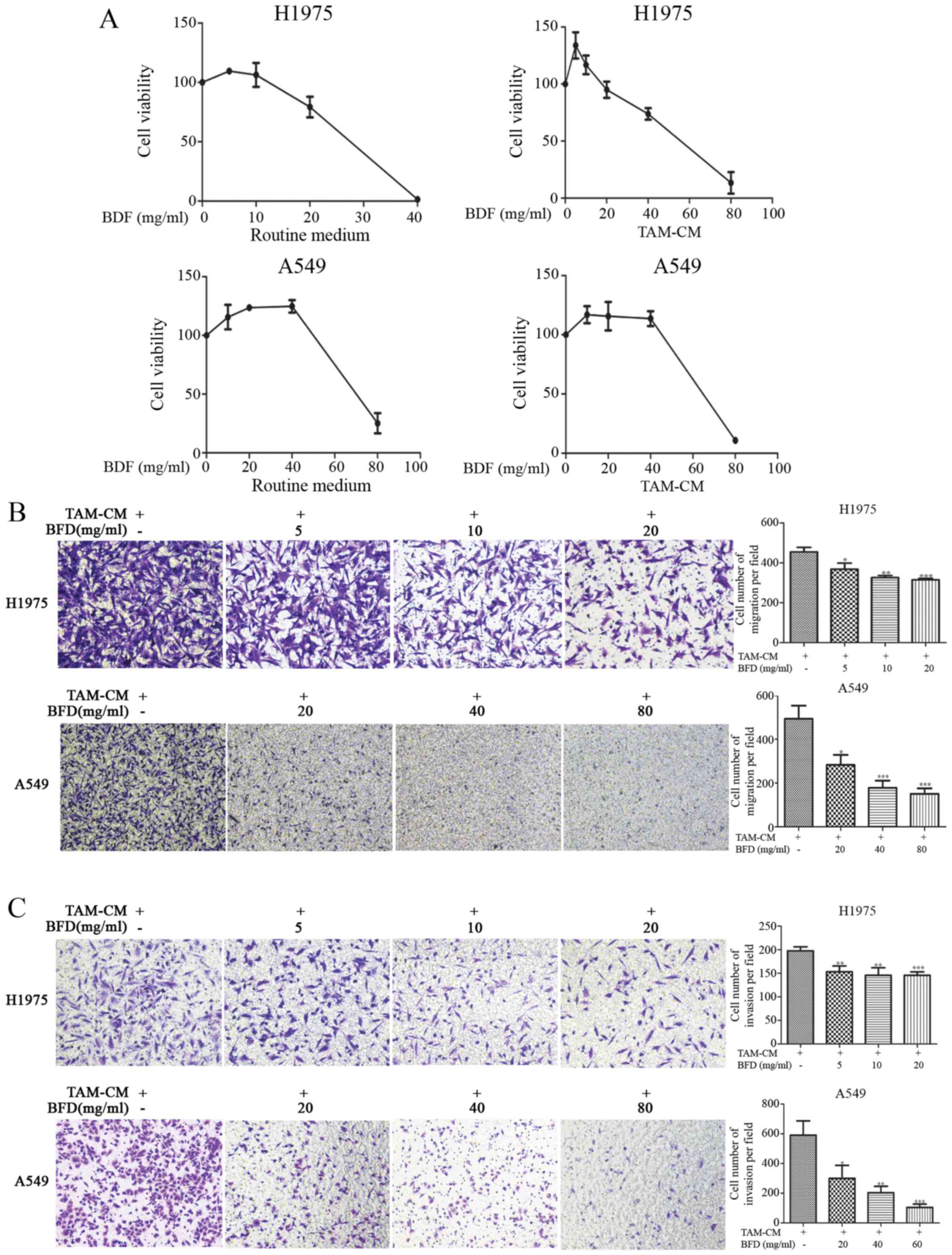 | Figure 3Dose-dependent suppression of NSCLC
cell proliferation, migration and invasion by BFD. (A) Cell
proliferation was reduced by BFD, H1975: IC50, 22.84
mg/ml (routine medium), IC50, 49.36 mg/ml (TAM-CM);
A549: IC50, 75.80 mg/ml (routine 1640 medium),
IC50, 73.62 mg/ml (TAM-CM). (B) The numbers of migrated
cell permeating septum were reduced by BFD. (C) The invasive
ability of cells was decreased by BFD, *p<0.05,
**p<0.01, ***p<0.001 vs TAM-CM. |
The number of migrated cells was reduced gradually
BFD dose-dependently, and there was a significant difference in
comparison to TAMs. The number of migrated H1975 cells was
368.0±30.65, 326.0±11.45 and 139.5±34.23 with BFD at dose of 5, 10
and 20 mg/ml, respectively, p<0.05. While for A549 cells, the
number was 283.0±45.82, 178.1±33.06 and 150.7±25.48 with BFD at
dose of 20, 40 and 80 mg/ml, respectively, p<0.05 (Fig. 3B).
The invasive ability of cancer cells was also
decreased BFD dose-dependently. Compared with the TAM-CM, the
number of invaded cells for H1975 cells was 153.1±12.44,
145.8±15.99 and 145.7±7.276 with BFD at dose of 5, 10 and 20 mg/ml,
respectively, p<0.01. For A549 cells, the number was 300.0±87.8,
203.8±42.42 and 105.1±21.71 with BFD at dose of 20, 40 and 80
mg/ml, respectively, p<0.05 (Fig.
3C).
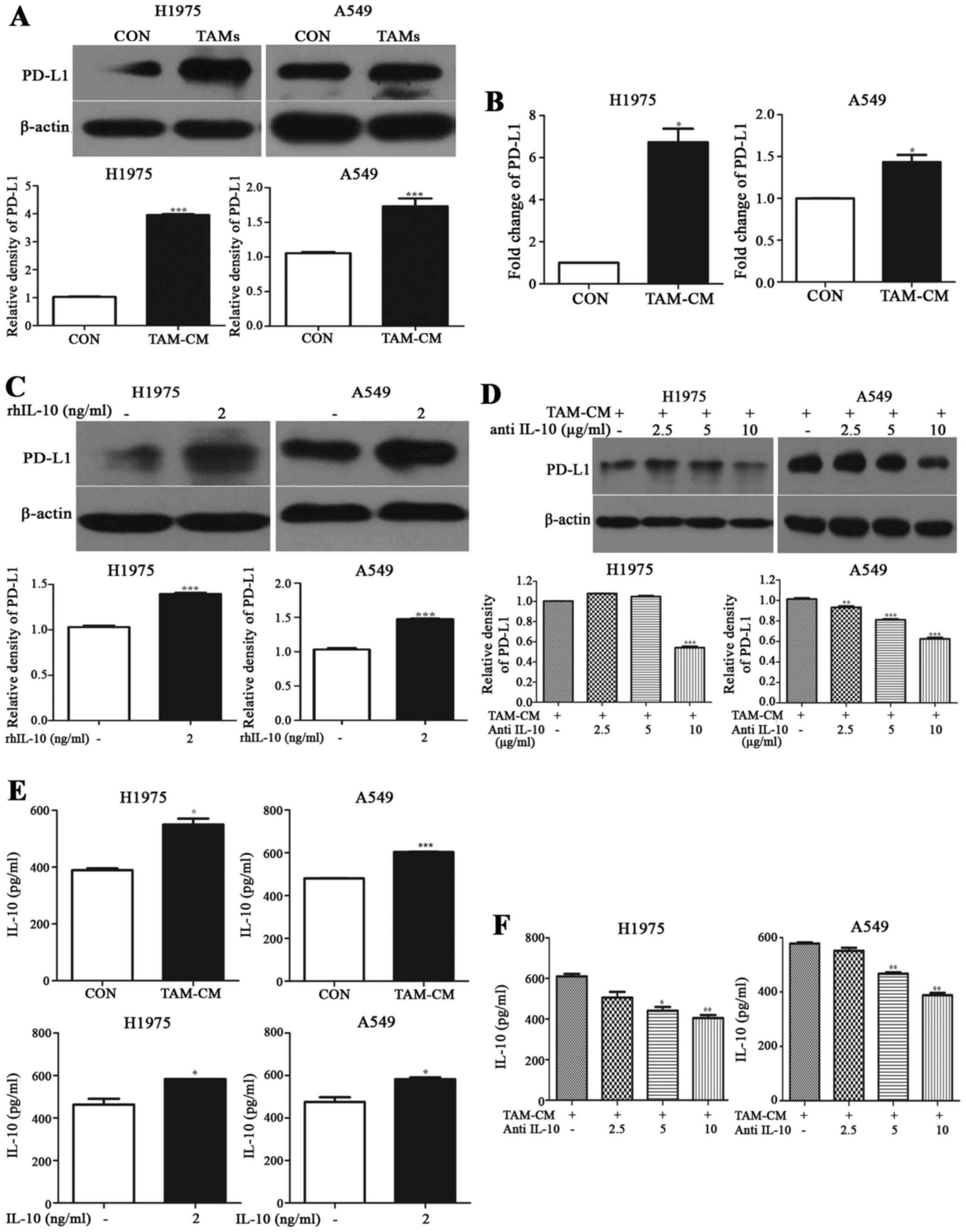
IL-10 secreted from TAMs promotes PD-L1
expression in NSCLC cells
Real-time PCR and western blotting showed that the
mRNA and protein expression of PD-L1 were higher in cancer cells
cocultured with M2-polarized TAMs than those in monoculture
(Fig. 4A and B). Compared with the
control group, the protein expression of PD-L1 in cancer cells
cocultured with TAMs was increased ~3-fold in H1975 and 0.7-fold in
A549, respectively (p<0.001). mRNA level was increased 6.7-fold
in H1975 and 1.432-fold in A549, in comparison with those of the
control group (p<0.05).
We tested the expression of PD-L1 in cancer cells in
rhIL-10 or TAM-CM with neutralizing mAbs against IL-10 instead of
TAM-CM. As shown in Fig. 4C,
compared with the control group, rhIL-10 promoted PD-L1 expression
on A549 and H1975 cells like TAMs, and the difference was
statistically significant (p<0.01). While TAM-CM with anti-IL-10
mAb gradually weakened the upregulation of PD-L1 on A549 and H1975
cells (p<0.05, Fig. 4D).
The level of IL-10 in TAM coculture system was
higher than cancer cells in monoculture. The level of IL-10 was
increased by 41.27% (p<0.05) in H1975 and 25.68% (p<0.001) in
A549 coculture system, respectively. It was increased by 25.87%
(p<0.05) in H1975 and 22.48% (p<0.05) in A549, respectively
(Fig. 4E).
The level of IL-10 in TAM-CM with anti-IL-10
coculture system was greatly lower than the TAM-CM group. The level
of IL-10 in H1975 treated with anti-IL-10 at a dose of 2.5, 5 and
10 µg/ml showed a 17.12 (p>0.05), 27.74 (p<0.05) and
33.62% (p<0.01) decrease, respectively. Similarly, IL-10 level
in A549 decreased by 4.60 (p>0.05), 19.10 (p<0.01) and 32.85%
(p<0.01) using 2.5, 5 and 10 µg/ml anti-IL-10 treatment
(Fig. 4F). It was established that
IL-10 secreted by TAMs induced PD-L1 expression in NSCLC cells.
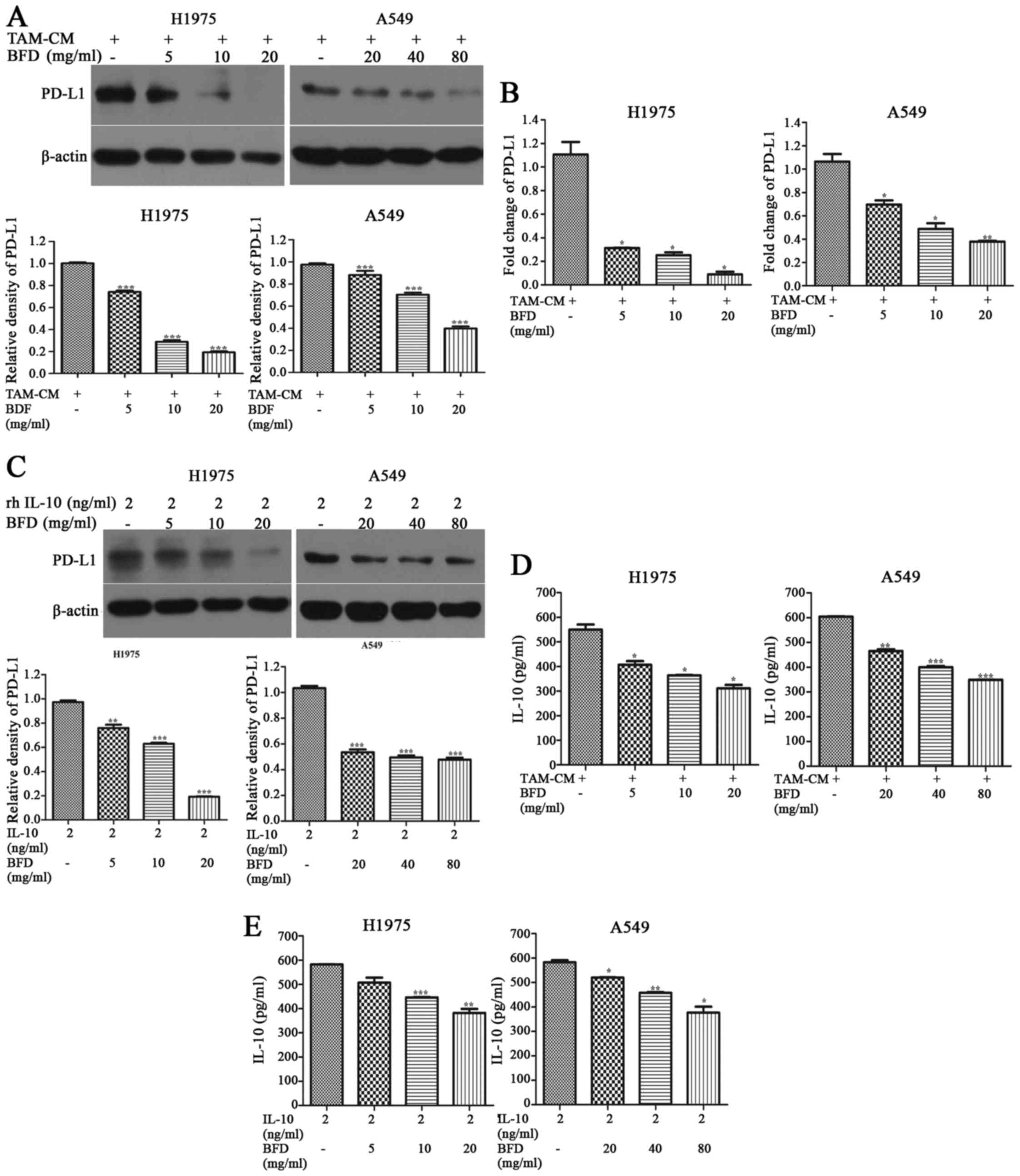
BFD reduces the expression of IL-10 and
PD-L1 in vitro
After cancer cells were treated with different doses
of BFD, the protein and mRNA expression of PD-L1 in NSCLC cells
cocultured with TAMs was gradually inhibited in comparison with the
TAM-CM group. The protein expression of PD-L1 in H1975 treated with
BFD at a dose of 5, 10 and 20 mg/ml showed a 25.92 (p<0.001),
71.32 (p<0.001) and 80.74% (p<0.001) decrease, respectively.
Similarly, PD-L1 protein in A549 decreased by 28.64 (p<0.01),
36.49 (p<0.01) and 70.57% (p<0.001) using 20, 40 and 80 mg/ml
BFD treatment (Fig. 5A). PD-L1
mRNA expression levels in H1975 treated with BFD at a dose of 5, 10
and 20 mg/ml showed a 71.84 (p<0.05), 77.27 (p<0.05) and
91.97% (p<0.05) decrease, respectively. Similarly, PD-L1 mRNA
expression in A549 was decreased by 34.71 (p<0.05), 54.30
(p<0.05) and 64.47% (p<0.01) using 20, 40, and 80 mg/ml BFD
treatment (Fig. 5B).
PD-L1 in NSCLC cells induced by rhIL-10 was also
inhibited BFD dose-dependently. Compared with the rhIL-10 group,
PD-L1 in H1975 treated with BFD at a dose of 5, 10 and 20 mg/ml
showed a 21.99 (p<0.001), 35.36 (p<0.001) and 80.28%
(p<0.001) decrease, respectively. Similarly, PD-L1 in A549
decreased by 48.26 (p<0.001), 53.45 (p<0.001) and 55.16%
(p<0.001) using 20, 40 and 80 mg/ml BFD treatment (Fig. 5C).
IL-10 in TAM coculture system was decreased by BFD
dose-dependently. Compared with the TAM-CM group, IL-10 in H1975
treated with BFD at a dose of 5, 10 and 20 mg/ml showed a 26.01
(p<0.05), 33.89 (p<0.05) and 43.42% (p<0.05) decrease,
respectively. Similarly, IL-10 in A549 decreased by 22.98
(p<0.01), 33.76 (p<0.001) and 42.21% (p<0.001) using 20,
40 and 80 mg/ml BFD (Fig. 5D).
BFD decreased IL-10 in NSCLC culture system induced
rhIL-10 dose-dependently. Compared with the rhIL-10 group, IL-10 in
H1975 treated with BFD at a dose of 5, 10 and 20 mg/ml showed a
12.93 (p>0.05), 23.46 (p<0.001) and 34.51% (p<0.01)
decrease, respectively. IL-10 in A549 decreased by 11.2
(p<0.05), 21.44 (p<0.01) and 35.38% (p<0.05) using 20, 40
and 80 mg/ml BFD (Fig. 5E).
BFD suppresses A549 and H1975 tumor
growth in xenograft mice
We evaluated the anticancer effect of BFD on female
nude mice bearing A549 or H1975 tumors. Comparing BFD groups with
the control group, there was no significant intergroup difference
in body weight during the therapy (p>0.05) (Fig. 6A). In H1975 xenograft mice, BFD (30
g/kg) group decreased tumor volume and tumor weight to some extent,
there was a statistical difference in comparison to the control
group (p<0.05); BFD (60 g/kg) group decreased tumor volume and
tumor weight to a great extent, and the difference was
statistically significant (p<0.05) (Fig. 6B). In A549 xenograft mice, tumor
volume and tumor weight slightly decreased by 30 g/kg BFD, there
was no statistical difference (p>0.05). While tumor volume and
tumor weight greatly decreased with 60 g/kg BFD compared with the
control group (p<0.05) (Fig.
6C).
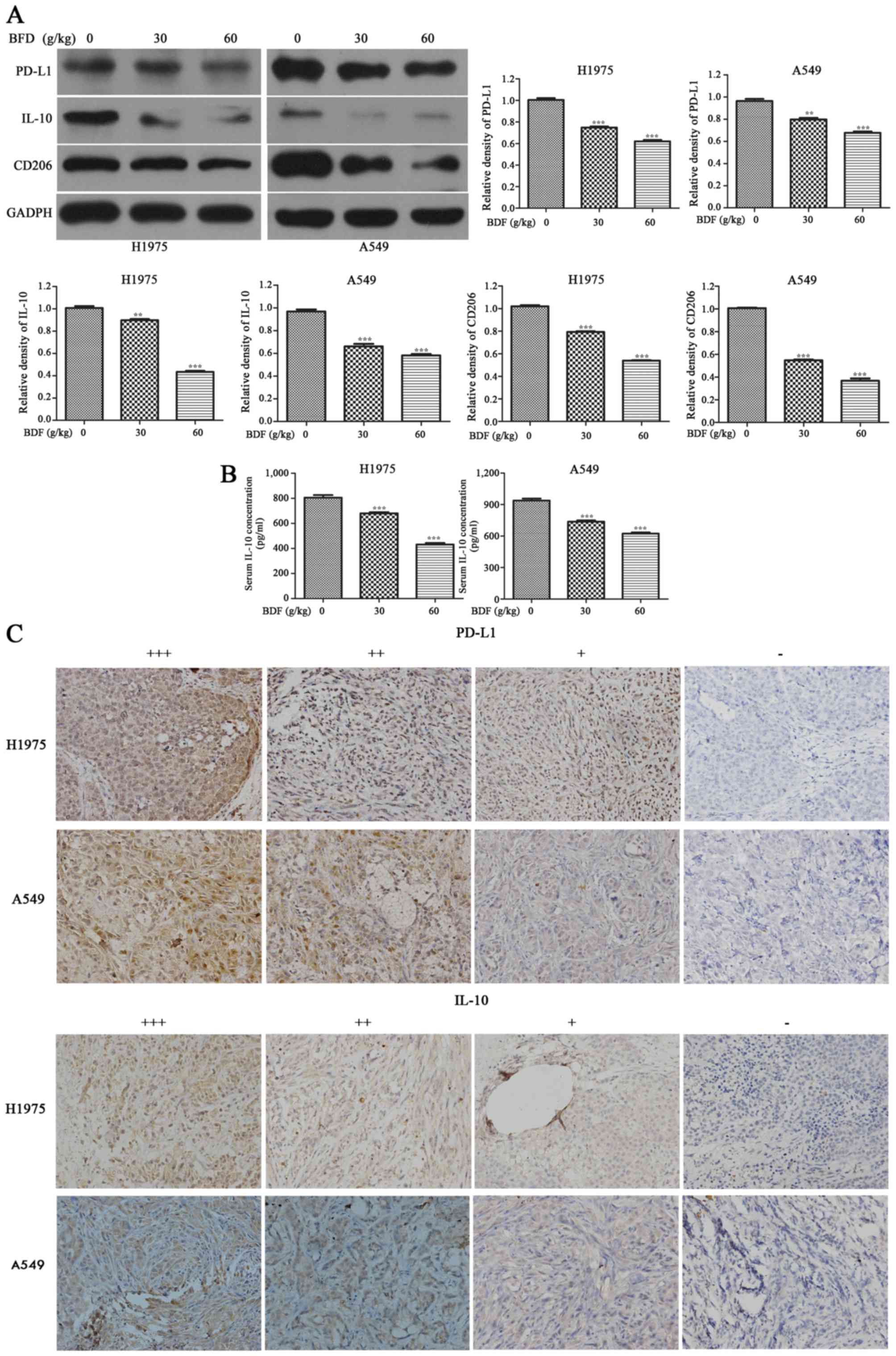
BFD reduces PD-L1, IL-10 and CD206
protein expression in vivo
BFD resulted in a downregulation of PD-L1, IL-10 and
CD206 in A549 and H1975 tumor tissue dose-dependently. As shown in
Fig. 7A, compared with the control
group, BFD 30 and 60 g/kg resulted in a downregulation of PD-L1 in
H1975 tumor tissue by 25.59 (p<0.001) and 38.39% (p<0.001),
respectively. While for A549 tumor tissue, PD-L1 was decreased by
17.24 (p<0.01) in BFD 30 and 29.79% (p<0.001) in BFD 60 g/kg.
IL-10 in H1975 tumor tissue was decreased by 51.66 (p<0.001) in
BFD 30 g/kg and 62.44% (p<0.001) in BFD 60 g/kg. While for A549
tumor tissue, BFD 30 and 60 g/kg also resulted in a significant
decrease in IL-10 by 28.87 (p<0.01) and 39.50% (p<0.01),
respectively. BFD 30 and 60 g/kg resulted in a downregulation of
CD206 in H1975 tumor tissue by 22.25 (p<0.001) and 47.21%
(p<0.001), respectively. While for A549, CD206 was decreased by
43.7% (p<0.001) in BFD 30 g/kg and 60.82% (p<0.001) in BFD 60
g/kg.
The serum level of IL-10 was significantly lower BFD
dose-dependently, compared with the control group (p<0.001)
(Fig. 7B). Using IHC staining,
PD-L1 and IL-10 were analyzed in all tumor specimens.
PD-L1-positive cells were evenly scattered throughout the
specimens, it was located primarily in the cytoplasm and membrane
of tumor cells. IL-10 was also generally expressed in the cytoplasm
of tumor cells. The above was consistent with the study of Geng
et al (34). Representative
IHC staining is shown in Fig.
7C.
Discussion
IL-10 and PD-L1 are two major immunosuppressor
proteins associated with tumorigenesis and tumor progression. Tumor
associated macrophages represent key regulators of the complex
interplay between the immune system and cancer (35). As a classic compound of TCM, BFD is
widely used to alleviate lung cancer related symptoms by improving
lung functions and enhancing the body immunity. In our study,
conditioned medium from TAMs promoted the expression of PD-L1 in
NSCLC cells by secreting IL-10. Furthermore, IL-10 and PD-L1 was
decreased by BFD in vitro and in vivo.
M2-polarized TAMs were established in vitro.
Taniguchi et al (7) used
PMA and IL-4 to treat U937 cells and obtained M2-polarized TAMs. We
treated U937 cells as described above. M2-polarized TAMs were
successfully induced and confirmed (Fig. 1). Tumor associated macrophage
conditioned medium (TAM-CM) was collected and extracted, as
described by Liu et al and Chen et al (11,36).
In the present study, TAMs significantly promoted
the proliferation, migration and invasion of NSCLC cells, H1975 and
A549, as expected (Fig. 2). The
results suggested that TAMs provided a suitable microenvironment
for tumor progression. Some cytokines may exist in the
microenvironment of TAMs probably to promote tumor growth and
metastasis. Previous studies also revealed that M2-polarized
macrophages induced tumor proliferation, migration and invasion in
hepatocellular carcinoma, human basal carcinoma cells, pancreatic
cancer and prostate cancer cells (9,10,17).
When NSCLC cells were treated with different doses of BFD, the
abilities of cell proliferation, migration and invasion induced by
TAMs were gradually decreased, and there was a significant
difference in comparison to the TAM group (Fig. 3). Thus, we presumed BFD probably
inhibited some target cytokine or protein secreted by TAMs, causing
change of the tumor microenvironment and interruption of the
interaction between TAMs and cancer cells. The next key work is to
find target cytokines or protein associated with TAMs and BFD.
Immunosuppressive cytokines (e.g., IL-10 and
TGF-β) or proteases secreted by TAMs probably play an important
role in tumor progression. IL-10 may tune inflammatory responses
and adaptive Th2 immunity, which strongly inhibits Th1 cytokines
(IFN-γ) to exert antitumor effect, and the tumor progression is
accelerated (37–39). As another important
immunosuppressive protein, PD-L1 is a co-inhibitory molecule
expressed generally on antigen-presenting cells and tumor cells
(40). By binding to its receptor
PD-1, which is mainly expressed in activated T cells, PD-L1 induces
apoptosis, anergy, unresponsiveness, and exhaustion of T cells
(41,42). We detected that the expression of
PD-L1 was significantly increased in NSCLC cells when cocultured
with TAMs (p<0.001) (Fig. 4A and
B). Prior research has shown that PD-L1 expression was
upregulated in HCC cells by macrophages. They also found a
significant correlation between PD-L1 expression and TAM
infiltration in 63 HCC tissues samples (43). Also, if we suppose some cytokines
released from TAMs induced the overexpression of PD-L1 in cancer
cells and activated relevant signal pathway. In order to find the
target cytokine in TAM-CM, TAM-CM was replaced with rhIL-10 or
TAM-CM with neutralizing mAbs against IL-10 in the culture system.
The results showed that the expression of PD-L1 in cancer cells was
related to the level of IL-10 in the culture system (Fig. 4C and D). It was probably IL-10
secreted by TAMs that regulated the expression of PD-L1 in tumor
cells. Some studies showed that PD-L1 expression in tumor
infiltrative macrophages could be upregulated through modulation of
IL-10 autocrine/paracrine signaling in gliomas and HCC, leading to
macrophages with immunosuppressive phenotype (27,44).
We found the correlation between IL-10 secreted by TAMs and PD-L1
in NSCLC cells. Geng and colleagues (34) indicated that tumor-associated PD-L1
expression was correlated with IL-10 products in 40 pancreatic
carcinoma samples. Moreover, they also found PD-L1 expression was
significantly associated with poor tumor differentiation
(p<0.01) and advanced tumor stage (p<0.01). Kuang et
al (27) found that IL-10
regulated the PD-L1 expression in macrophages of hepatoma tissue,
which was similar to our results. To the best of our knowledge,
this study is the first to demonstrate the association between
PD-L1 in cancer cells and IL-10 secreted by TAMs in NSCLC.
When NSCLC cells were treated with different doses
of BFD, the expression of PD-L1 and IL-10 induced by TAMs was
gradually inhibited in vitro, and the difference was
statistically significant (Fig.
5). In addition, the administration of BFD resulted in a
reduction in the expression of PD-L1, IL-10 and CD206 in tumor
tissues (Fig. 7A). Overexpression
of PD-L1 has been reported to correlate with poor prognosis in a
number of human cancers, including breast cancer, kidney cancer,
and NSCLCs (45). Blocking PD-L1
with a specific antibody improved specific T cell immunity,
suggesting that PD-L1 limits the capacity of T cells to eliminate
tumor cells (46). The
concentration of IL-10 in serum of xenograft mice was significantly
decreased BFD dose-dependently (Fig.
7B). High IL-10 level in serum or tumors was associated with
worse survival in lung cancer patients (47). High level of IL-10 secreted from
TAMs was correlated with late stage (stage II, III and IV), lymph
node metastases, pleural invasion, lymphovascular invasion and poor
differentiation in NSCLC patients (39). Consistent with our in vitro
results, these results in vivo suggested that BFD
successfully interrupted the interaction between tumor cells and
TAMs by inhibiting the expression of IL-10 and PD-L1.
In our study, we constructed BFD with six kinds of
single Chinese herbs in different proportions. Thus, it is highly
probable that one or more of these compounds inhibited the
activities of PD-L1 and IL-10. For example, as one of BFD
components, Milkvetch root is a good immunomodulator (48–50).
Milkvetch root suppresses the production of oxygen free radicals to
reduce damage to the biomembrane (51). It also could regulate T-lymphocyte
function, inhibit NF-κB activation, and control clinical symptoms
in patients with lung cancer. We focused on the BFD regulation of
immunosuppression in this investigation to explain its antitumor
mechanism. Its antitumor effect is probably through downregulating
the expression of PD-L1 and IL-10 in tumor environment of TAMs.
Our results found that BFD groups had no influence
on body weight during the whole therapy (Fig. 6), but its long-term administration
still needs further study. Besides, as a classic compound of TCM,
BFD is probably a multi-targeted antitumor drug, proteomics may be
helpful to find more target proteins of the drug action. Going
further into the antitumor mechanism with proteomics is of great
significance in the future. However, the validation by western
blotting is required subsequently to proteomics. Pharmaceutical
analysis in BFD extract is required in future study. In order to
make our research comprehensive, we are currently conducting
related clinical trials of BFD.
In conclusion, the authors showed that TAMs play an
important role in tumor progression of NSCLC, which was associated
with tumor proliferation, migration, invasion and
immunosuppression. Moreover, the antitumor mechanism of BFD is
related to interruption of the link between TAMs and cancer cells
by inhibiting the expression of IL-10 and PD-L1 in vitro and
in vivo. Our results demonstrated BFD's potential as a novel
treatment for NSCLC.
Acknowledgments
The authors would like to acknowledge Huirong Ding
for his help with flow cytometry, Bin Dong for her help with IHC
staining and Guo An for her help with animal experiment at the
Central Laboratory of Peking University Cancer Hospital and
Institute.
Glossary
Abbreviations
Abbreviations:
|
TCM
|
traditional Chinese medicine
|
|
BFD
|
Bu-Fei Decoction
|
|
NSCLC
|
non-small cell lung cancer
|
|
TAMs
|
tumor associated macrophages
|
|
TAM-CM
|
tumor associated
macrophage-conditioned medium
|
|
IL-10
|
interleukin 10
|
|
PD-L1
|
programmed death ligand-1
|
|
PMA
|
phorbol 12-myristate 13-acetate
|
|
IL-4
|
interleukin 4
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Howlader N, Noone AM, Krapcho M, Miler D,
Bishop K, Altekruse SF, Kosary CL, Yu M, Ruhl J, Tatalovich Z, et
al: SEER Cancer Statistics Review, 1975–2013. National Cancer
Institute; Bethesda, MD: http://seer.cancer.gov/csr/1975_2013/urisimpleseer.cancer.gov/csr/1975_2013/(based
on November 2015 SEER data submission, posted to the SEER web site,
April 2016).
|
|
3
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gardiner RE, Jahangeer S, Forde P, Ariffin
AB, Bird B, Soden D and Hinchion J: Low immunogenicity in non-small
cell lung cancer; do new developments and novel treatments have a
role? Cancer Metastasis Rev. 34:129–144. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Geissmann F, Manz MG, Jung S, Sieweke MH,
Merad M and Ley K: Development of monocytes, macrophages, and
dendritic cells. Science. 327:656–661. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Solinas G, Germano G, Mantovani A and
Allavena P: Tumor-associated macrophages (TAM) as major players of
the cancer-related inflammation. J Leukoc Biol. 86:1065–1073. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taniguchi K, Hikiji H, Okinaga T,
Hashidate-Yoshida T, Shindou H, Ariyoshi W, Shimizu T, Tominaga K
and Nishihara T: Essential role of lysophosphatidylcholine
acyltransferase 3 in the induction of macrophage polarization in
PMA-treated U937 cells. J Cell Biochem. 116:2840–2848. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Coffelt SB, Hughes R and Lewis CE:
Tumor-associated macrophages: Effectors of angiogenesis and tumor
progression. Biochim Biophys Acta. 1796:11–18. 2009.PubMed/NCBI
|
|
9
|
Zwadlo G, Voegeli R, Schulze Osthoff K and
Sorg C: A monoclonal antibody to a novel differentiation antigen on
human macrophages associated with the down-regulatory phase of the
inflammatory process. Exp Cell Biol. 55:295–304. 1987.PubMed/NCBI
|
|
10
|
Lee JH, Lee GT, Woo SH, Ha YS, Kwon SJ,
Kim WJ and Kim IY: BMP-6 in renal cell carcinoma promotes tumor
proliferation through IL-10-dependent M2 polarization of
tumor-associated macrophages. Cancer Res. 73:3604–3614. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu C-Y, Xu J-Y, Shi X-Y, Huang W, Ruan
T-Y, Xie P and Ding J-L: M2-polarized tumor-associated macrophages
promoted epithelial-mesenchymal transition in pancreatic cancer
cells, partially through TLR4/IL-10 signaling pathway. Lab Invest.
93:844–854. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang R, Zhang J, Chen S, Lu M, Luo X, Yao
S, Liu S, Qin Y and Chen H: Tumor-associated macrophages provide a
suitable microenvironment for non-small lung cancer invasion and
progression. Lung Cancer. 74:188–196. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tjiu JW, Chen JS, Shun CT, Lin SJ, Liao
YH, Chu CY, Tsai TF, Chiu HC, Dai YS, Inoue H, et al:
Tumor-associated macrophage-induced invasion and angiogenesis of
human basal cell carcinoma cells by cyclooxygenase-2 induction. J
Invest Dermatol. 129:1016–1025. 2009. View Article : Google Scholar
|
|
14
|
Li CC, Hou YC, Yeh CL and Yeh SL: Effects
of eicosapentaenoic acid and docosahexaenoic acid on prostate
cancer cell migration and invasion induced by tumor-associated
macrophages. PLoS One. 9:e996302014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Condeelis J and Pollard JW: Macrophages:
Obligate partners for tumor cell migration, invasion, and
metastasis. Cell. 124:263–266. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kurahara H, Shinchi H, Mataki Y, Maemura
K, Noma H, Kubo F, Sakoda M, Ueno S, Natsugoe S and Takao S:
Significance of M2-polarized tumor-associated macrophage in
pancreatic cancer. J Surg Res. 167:e211–e219. 2011. View Article : Google Scholar
|
|
17
|
Mellor AL and Munn DH: Creating immune
privilege: Active local suppression that benefits friends, but
protects foes. Nat Rev Immunol. 8:74–80. 2008. View Article : Google Scholar
|
|
18
|
Fan Q-M, Jing Y-Y, Yu GF, Kou XR, Ye F,
Gao L, Li R, Zhao QD, Yang Y, Lu ZH, et al: Tumor-associated
macrophages promote cancer stem cell-like properties via
transforming growth factor-beta1-induced epithelial-mesenchymal
transition in hepatocellular carcinoma. Cancer Lett. 352:160–168.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pollard JW: Tumour-educated macrophages
promote tumour progression and metastasis. Nat Rev Cancer. 4:71–78.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin EY, Nguyen AV, Russell RG and Pollard
JW: Colony-stimulating factor 1 promotes progression of mammary
tumors to malignancy. J Exp Med. 193:727–740. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Laoui D, Van Overmeire E, De Baetselier P,
Van Ginderachter JA and Raes G: Functional relationship between
tumor-associated macrophages and macrophage colony-stimulating
factor as contributors to cancer progression. Front Immunol.
5:4892014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Robinson SC, Scott KA, Wilson JL, Thompson
RG, Proudfoot AE and Balkwill FR: A chemokine receptor antagonist
inhibits experimental breast tumor growth. Cancer Res.
63:8360–8365. 2003.PubMed/NCBI
|
|
23
|
Whitehurst B, Flister MJ, Bagaitkar J,
Volk L, Bivens CM, Pickett B, Castro-Rivera E, Brekken RA, Gerard
RD and Ran S: Anti-VEGF-A therapy reduces lymphatic vessel density
and expression of VEGFR-3 in an orthotopic breast tumor model. Int
J Cancer. 121:2181–2191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dineen SP, Lynn KD, Holloway SE, Miller
AF, Sullivan JP, Shames DS, Beck AW, Barnett CC, Fleming JB and
Brekken RA: Vascular endothelial growth factor receptor 2 mediates
macrophage infiltration into orthotopic pancreatic tumors in mice.
Cancer Res. 68:4340–4346. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rauh MJ, Sly LM, Kalesnikoff J, Hughes MR,
Cao LP, Lam V and Krystal G: The role of SHIP1 in macrophage
programming and activation. Biochem Soc Trans. 32:785–788. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Torisu-Itakura H, Lee JH, Huynh Y, Ye X,
Essner R and Morton DL: Monocyte-derived IL-10 expression predicts
prognosis of stage IV melanoma patients. J Immunother. 30:831–838.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kuang DM, Zhao Q, Peng C, Xu J, Zhang JP,
Wu C and Zheng L: Activated monocytes in peritumoral stroma of
hepatocellular carcinoma foster immune privilege and disease
progression through PD-L1. J Exp Med. 206:1327–1337. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xia Y, Wang Y, Yang J and Song K: Study on
effects of Bufei Decoction on α-SMA in lung tissue of pulmonary
fibrosis rats. Zhonghua Zhongyiyao Xuekan. 23:1916–1918. 2014.In
Chinese.
|
|
29
|
Zhang H, Xia YL, Yang J and Song K: Study
on effects of Bufei Decoction on MMP-9 and TIMP-1 in lung tissue of
pulmonary fibrosis rats. Zhonghua Zhongyiyao Xuekan. 29:2673–2679.
2011.
|
|
30
|
Du X, Du X, Zhang K, Wang J, Guo G and
Wang R: The clinical trials to observe the effect of Astragalus
injection combine with chemotherapy in patients with malignant
tumor. Shanghai Med Pharm J. 2:162010.
|
|
31
|
Chiu PY, Lam PY, Yan CW and Ko KM:
Schisandrin B protects against solar irradiation-induced oxidative
injury in BJ human fibroblasts. Fitoterapia. 82:682–691. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chi D and Huang Q: The effect of Fructus
schisandrae B element to vascular endothelial growth factor in
human glioma. PhD thesis. Southern Medical University; Guangzhou:
2008, In Chinese.
|
|
33
|
Reagan-Shaw S, Nihal M and Ahmad N: Dose
translation from animal to human studies revisited. FASEB J.
22:659–661. 2008. View Article : Google Scholar
|
|
34
|
Geng L, Huang D, Liu J, Qian Y, Deng J, Li
D, Hu Z, Zhang J, Jiang G and Zheng S: B7-H1 up-regulated
expression in human pancreatic carcinoma tissue associates with
tumor progression. J Cancer Res Clin Oncol. 134:1021–1027. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ostuni R, Kratochvill F, Murray PJ and
Natoli G: Macrophages and cancer: From mechanisms to therapeutic
implications. Trends Immunol. 36:229–239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen C, Shen Y, Qu QX, Chen XQ, Zhang XG
and Huang JA: Induced expression of B7-H3 on the lung cancer cells
and macrophages suppresses T-cell mediating anti-tumor immune
response. Exp Cell Res. 319:96–102. 2013. View Article : Google Scholar
|
|
37
|
Lei H, Ju DW, Yu Y, Tao Q, Chen G, Gu S,
Hamada H and Cao X: Induction of potent antitumor response by
vaccination with tumor lysate-pulsed macrophages engineered to
secrete macrophage colony-stimulating factor and interferon-γ. Gene
Ther. 7:707–713. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Salazar-Onfray F: Interleukin-10: A
cytokine used by tumors to escape immunosurveillance. Med Oncol.
16:86–94. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang R, Lu M, Zhang J, Chen S, Luo X, Qin
Y and Chen H: Increased IL-10 mRNA expression in tumor-associated
macrophage correlated with late stage of lung cancer. J Exp Clin
Cancer Res. 30:622011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee SJ, Jang BC, Lee SW, Yang YI, Suh SI,
Park YM, Oh S, Shin JG, Yao S, Chen L, et al: Interferon regulatory
factor-1 is prerequisite to the constitutive expression and
IFN-gamma-induced upregulation of B7-H1 (CD274). FEBS Lett.
580:755–762. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dong H, Strome SE, Salomao DR, Tamura H,
Hirano F, Flies DB, Roche PC, Lu J, Zhu G, Tamada K, et al:
Tumor-associated B7-H1 promotes T-cell apoptosis: A potential
mechanism of immune evasion. Nat Med. 8:793–800. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Barber DL, Wherry EJ, Masopust D, Zhu B,
Allison JP, Sharpe AH, Freeman GJ and Ahmed R: Restoring function
in exhausted CD8 T cells during chronic viral infection. Nature.
439:682–687. 2006. View Article : Google Scholar
|
|
43
|
Chen J, Li G, Meng H, Fan Y, Song Y, Wang
S, Zhu F, Guo C, Zhang L and Shi Y: Upregulation of B7-H1
expression is associated with macrophage infiltration in
hepatocellular carcinomas. Cancer Immunol Immunother. 61:101–108.
2012. View Article : Google Scholar
|
|
44
|
Bloch O, Crane CA, Kaur R, Safaee M,
Rutkowski MJ and Parsa AT: Gliomas promote immunosuppression
through induction of B7-H1 expression in tumor-associated
macrophages. Clin Cancer Res. 19:3165–3175. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Velcheti V, Schalper KA, Carvajal DE,
Anagnostou VK, Syrigos KN, Sznol M, Herbst RS, Gettinger SN, Chen
LP and Rimm DL: Programmed death ligand-1 expression in non-small
cell lung cancer. Lab Invest. 94:107–116. 2014. View Article : Google Scholar
|
|
46
|
Chanmee T, Ontong P, Konno K and Itano N:
Tumor-associated macrophages as major players in the tumor
microenvironment. Cancers (Basel). 6:1670–1690. 2014. View Article : Google Scholar
|
|
47
|
Hatanaka H, Abe Y, Kamiya T, Morino F,
Nagata J, Tokunaga T, Oshika Y, Suemizu H, Kijima H, Tsuchida T, et
al: Clinical implications of interleukin (IL)-10 induced by
non-small-cell lung cancer. Ann Oncol. 11:815–819. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jin M, Zhao K, Huang Q and Shang P:
Structural features and biological activities of the
polysaccharides from Astragalus membranaceus. Int J Biol Macromol.
64:257–266. 2014. View Article : Google Scholar
|
|
49
|
Li W, Sun YN, Yan XT, Yang SY, Song SB,
Lee YM and Kim YH: NF-κB inhibitory activity of sucrose fatty acid
esters and related constituents from Astragalus membranaceus. J
Agric Food Chem. 61:7081–7088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chen X, May B, Di YM, Zhang AL, Lu C, Xue
CC and Lin L: Oral Chinese herbal medicine combined with
pharmacotherapy for stable COPD: A systematic review of effect on
BODE index and six minute walk test. PLoS One. 9:e918302014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang T, Sun Y, Jin L, Xu Y, Wang L, Ren T
and Wang K: Enhancement of non-specific immune response in sea
cucumber (Apostichopus japonicus) by Astragalus membranaceus and
its polysaccharides. Fish Shellfish Immunol. 27:757–762. 2009.
View Article : Google Scholar : PubMed/NCBI
|