Introduction
Telomeres are linear structures that are located at
the end of eukaryotic chromosomes performing important functions
during chromosome integration and stability (1,2). In
somatic cells, which lack telomerase, the length of a telomere is
constantly being shortened by the process of cell replication.
Telomerase, a ribonucleoprotein enzyme that is comprised of
telomerase reverse transcriptase (hTERT) and an RNA subunit (TERC)
in addition to other proteins that control telomerase activity to
prevent telomere shortening (3,4). It
has been confirmed that hTERT overexpression plays a key role in
the initiation of unlimited proliferation during oncogenesis and
that introducing hTERT into epithelial cells and fibroblasts
conferred infinite replicative potential (5–7).
Moreover, overexpressing hTERT has been shown to be associated with
advanced invasive tumor progression and worse prognoses (8–10).
Currently available data demonstrate that >90% of malignant
tumors express high levels of telomerase activity, while normal and
benign tissues do not with the exception of lymphoid cells,
hematopoietic cells and germ cells, which possess proliferative
potential (11,12). These data indicate that telomerase
might be useful as a molecular marker of tumors in a wide range of
applications.
A primary phenotype of cancer is aneuploidy
(13–16). Aneuploidy refers to any deviation
from a normal number of chromosomes that results from a
whole-chromosome gain or loss in a cell. Aneuploidy is generally
caused by mitotic errors that occur as a result of a deficiency in
one of the mechanisms, such as the spindle assembly checkpoint
(SAC) (17–19), that are involved in ensuring that
chromosomes segregate correctly. In humans, aneuploidy is harmful
because it can cause some congenital malformation syndromes,
spontaneous abortion and stillbirth. More than 90% of solid tumors
and 50% of blood tumors are aneuploid (20). Although aneuploidy is observed in
most human cancers, the functional roles of aneuploidy during tumor
formation remain poorly understood. In mice and humans, a majority
of autosomal trisomies and monosomies are detrimental to organismal
survival (21), and it has been
demonstrated that aneuploidy blocks cellular proliferation,
spontaneous immortalization and survival (22). In contrast, because of its passive
effects on cell immortalization and viability, aneuploidy is a
hallmark of cancer cells (13). A
recent bioinformatics analysis showed that the number of genes that
can prompt tumorigenesis is higher than was previously thought and
that aneuploidy may contribute to the development of the complexity
of tumor genomics that affect multiple tumor-driving genes
(23). Genetic studies performed
using mouse models have provided experimentally based evidence
showing that mutations in genes that regulate chromosome
segregation initiate carcinogenesis by inducing the loss of
heterozygosity (24). However, the
influence of this loss on tumorigenesis is context-dependent, and
aneuploidy was found to repress carcinogenesis in some mouse
strains (25). Studies performed
in yeast have identified genes that affect cell survival in the
presence of aneuploidy. The survival factors associated with
aneuploidy in human cells in addition to their functions in cancer
formation remain to be determined. It was previously shown that the
survival of aneuploid cells is limited by ATM/p53-dependent DNA
damage checkpoints (26).
Telomerase was found to function as an aneuploidy-associated
survival factor that abrogated aneuploidy-induced DNA damage
responses, premature senescence and cell depletion by alleviating
telomere replication stress (27).
Telomerase-targeted therapies for tumors is
receiving an increasing amount of attention, but substantial
progress toward a clinical treatment involving telomerase has not
been achieved. Azido-deoxythymidine (AZT) is a type of nucleoside
analogue that is approved by the Food and Drug Administration to
treat acquired immune deficiency syndrome (AIDS). AZT inhibits DNA
double-strand formation by destroying the RNA reverse
transcriptional ability of the virus, which results in the loss of
the replication template and a consequential decrease in the
replication rate (28–31). AZT has been shown to inhibit
telomerase activity in a variety of cancer cells, including colon,
human brain glioma and liver cancer cells (32–34).
It is clear that telomerase and aneuploidy play
important roles in tumor development. In the present study, we
demonstrate that telomerase activity and the expression of hTERT
were higher in aneuploid than in euploid cells. In addition, we
found that the telomerase inhibitor AZT inhibited cell
proliferation and induced apoptosis and cell cycle arrest in both
aneuploid and euploid cells via the downregulation of hTERT gene
expression.
Materials and methods
Cell lines and lentiviral
transduction
The human colorectal cancer cell line HCT116 was
obtained from the Institute of Basic Medical Sciences, Subei
Peoples' Hospital. To generate HCT116-shRNA-MAD2 cells, a
short-hairpin RNA (shRNA) that targeted human MAD2 was cloned into
the pInducer10-MirRP lentiviral vector. The shRNA-containing
plasmid was then packaged in lentiviral particles at the Vector
Core. Lenti-PuroEMPTY-VSVG was used as the background control. A
total of 1×106 cells/10-cm plate were transduced for 48
h with cell supernatant containing a lentivirus that was diluted in
fresh serum-free medium at a ratio of 1:1. Stable cells were
selected by incubating the cells with 10 µg/ml puromycin at
37°C for 3 weeks. The cells were maintained in Dulbecco's modified
Eagle's medium (DMEM; Gibco, Carlsbad, CA, USA) supplemented with
10% FBS (Sigma-Aldrich, St. Louis, MO, USA), 100 IU/ml
penicillin-streptomycin and 2 µmM L-glutamine at 37°C in a
humidified atmosphere containing 5% CO2.
The formation of aneuploidy
We used a doxycycline-induced program to
conditionally knock down MAD2, resulting in HCT116-shRNA-MAD2
cells. The HCT116-shRNA-MAD2 cells were divided into two groups:
one group was cultured in 0.2 µg/ml doxycycline-supplemented
medium for 16 h (referred to as Dox (+) or aneuploid cells) to
conditionally knock down MAD2 (HCT116-shRNA-MAD2 cells), and the
other group was cultured in non-doxycycline medium and used as the
control group (referred to as Dox (−) or euploid cells). HCT116 is
a human colorectal cancer cell line that is chromosomal
number-stable and has a near diploid karyotype.
Metaphase spreads
At 11 days after the doxycycline washout was
performed, cells at 70~80% confluency were treated with 80 ng/ml of
the mitotic inhibitor nocodazole (Sigma-Aldrich) and 10 µM
of the protease inhibitor MG132 (Selleck Chemicals, Houston, TX,
USA) and then incubated for 6 h at 37°C. The cells were then
treated with a hypotonic solution (0.01 M sodium citrate and 0.03 M
KCl) at 37°C for 15–20 min and fixed 3 times in Carnoy's fixative
(methanol:acetic acid, 3:1) that was pre-cooled on ice. The cells
were subsequently spread on glass slides that were pre-cooled at
−20°C. Metaphase cells were stained using Giemsa (Beijing Solarbio
Science and Technology Co., Ltd., Beijing, China), and chromosomes
were counted under a fluorescence microscope for the chromosome
counting analysis. Cells with any number of chromosomes other than
45 or 46 were defined as aneuploidy [the chromosome karyotype of
the HCT116 cell line showed that the stem line chromosome number is
near diploid with a modal number of 45 (62%) and polyploid at 46
(6.8%) according to the ATCC].
Western blot analysis
Total proteins were extracted using cell lysis
buffer from cells placed on ice and then quantified using a
Bradford protein assay. Equal amounts of cellular proteins (60
µg) were separated using 10% sodium dodecyl sulfate
(SDS)-PAGE according to the manufacturer's instructions (Beyotime
Institute of Biotechnology, Shanghai, China). The resolved proteins
were then transferred onto polyvinylidene difluoride (PVDF)
membranes (Millipore, Billerica, MA, USA). The membranes were
incubated in 5% fat-free milk or 5% BSA in 1X TBS containing 0.1%
Tween-20 (TBST) at room temperature for 2 h and then probed with
the following specific primary antibodies: MAD2L1, AF4005 (1:200,
R&D Systems, Minneapolis, MN, USA), α-tubulin (1:2,000, Vazyme
Biotech Co., Ltd., Nanjing, China), Puma, ABC158 (1:1,000;
Millipore), Bax, AB2915 (1:1,500; Millipore), p21, MABE325
(1:1,000; Millipore), γ-H2AX, or 05–636, (1:400; Millipore)
overnight at 4°C. After the membranes were washed 3 times with 1X
TBST, they were incubated with horseradish peroxidase
(HRP)-conjugated secondary antibodies (1:3,000; Millipore) at room
temperature for 90 min. Reactive bands were visualized using a
chemiluminescence detection system and analyzed using Image Lab
software.
Telomerase activity
Telomerase activity of cells was quantified using a
TeloTAGGG Telomerase PCR ELISA kit (Roche Diagnostics GmbH,
Mannheim, Germany) and a process called Telomeric Repeat
Amplification Protocol (TRAP) based on the protocol described by
Kim et al (35). A total of
2.5×103 cells were harvested from each sample for the
PCR reactions. The negative controls samples were heat-treated for
5 min at 95°C to inactivate telomerase activity. The positive
control was provided by the kit. The following PCR program was run
in a Thermal cycler: one cycle of 25°C for 20 min and 94°C for 5
min, 30 cycles of 94°C for 30 sec, 50°C for 30 sec, and 72°C for 90
sec and one cycle of 72°C for 10 min. The PCR products were
subsequently analyzed using ELISA, and telomerase activity was
obtained by determining the absorbance at 450 nm (with a reference
wavelength of 690 nm). The relative telomerase activities (RTA) of
various samples were determined using the following formula: RTA =
[(AS-AS,0)/AS,IS/]/[(ATS8-ATS8,0)/ATS8,IS] × 100, where AS,
absorbance of the sample; AS,0, absorbance of the heat-treated
sample; AS,IS, absorbance of the Internal Standard sample; ATS8,
absorbance of the control template; ATS8,0, absorbance of the lysis
buffer; and ATS8,IS, absorbance of the Internal Standard control
template.
Cell viability measured by CCK-8
assay
To investigate the effect of AZT (Sigma-Aldrich) on
cell viability, both groups of cells were subjected to CCK-8
assays. Briefly, the cells were seeded in 96-well culture plates at
4×103 cells/well and treated with one of a variety of
concentrations of AZT (i.e., 0, 100, 200, 250, 500 or 1,000
µM) for one of the following incubation times: 24, 48 and 72
h. The cell suspension was then exchanged for fresh medium, and the
cells were incubated with 10 µl of CCK-8 solution
(Invitrogen, Carlsbad, CA, USA) at 37°C for 4 h. Absorbance was
measured at 450 nm using an ELISA reader. All experiments were
performed 3 times including 5 replicates for each sample.
Reverse transcription (RT)-PCR
Total RNA was extracted from cultured cells using
TRIzol (Invitrogen). cDNA was obtained using an ABI7300 real-time
PCR system with SYBR-Green PCR Master Mix (Vazyme Biotech). The
primers used in these experiments are listed in Table I. The following cycling program was
used: one cycle at 95°C for 5 min, 40 cycles at 95°C for 10 sec,
60°C for 30 sec, 95°C for 15 sec, 60°C for 60 sec and 95°C for 15
sec. The samples were then held at 4°C until retrieved. The
relative level of RNA was normalized to that of GAPDH using the
2−ΔΔCt method.
 | Table IPrimers used for PCR
amplification. |
Table I
Primers used for PCR
amplification.
| Genes | Forward primer | Reverse primer |
|---|
| GAPDH |
5′-CGTCTTCACCACCATGGAGA-3′ |
5′-CGGCCATCGCCACAGTTT-3′ |
| hTERT |
5′-AAGTTCCTGCACTGGCTGATG-3′ |
5′-GCTTTGCAACTTGCTCCAGAC-3′ |
| Bax |
5′-GGCAGCTGACATGTTTTCTGAC-3′ |
5′-CACCCAACCACCCTGGTCTT-3′ |
| Puma |
5′-GACCTCAACGCACAGTACGAG-3′ |
5′-AGGAGTCCCATGATGAGATTGT-3′ |
| Noxa |
5′-GTGTTCCTGTTGGGCGTTAC-3′ |
5′-GGAGCATTTTCCGAACCTT-3′ |
| p21 |
5′-CCTGTCACTGTCTTGTACCCT-3′ |
5′-GCGTTTGGAGTGGTAGAAATCT-3′ |
Data analysis
All statistical analyses were performed using SPSS
18.0 for Windows 10 (SPSS, Inc., Chicago, IL, USA). Two-way ANOVA
with a Bonferroni post hoc test was used for multiple comparisons
of two groups, as indicated. The expression of hTERT and telomerase
activity were evaluated in the groups using two-tailed Student's
t-tests. The data are represented as the mean ± standard deviation
of the mean. A P<0.05 was defined as statistically
significant.
Results
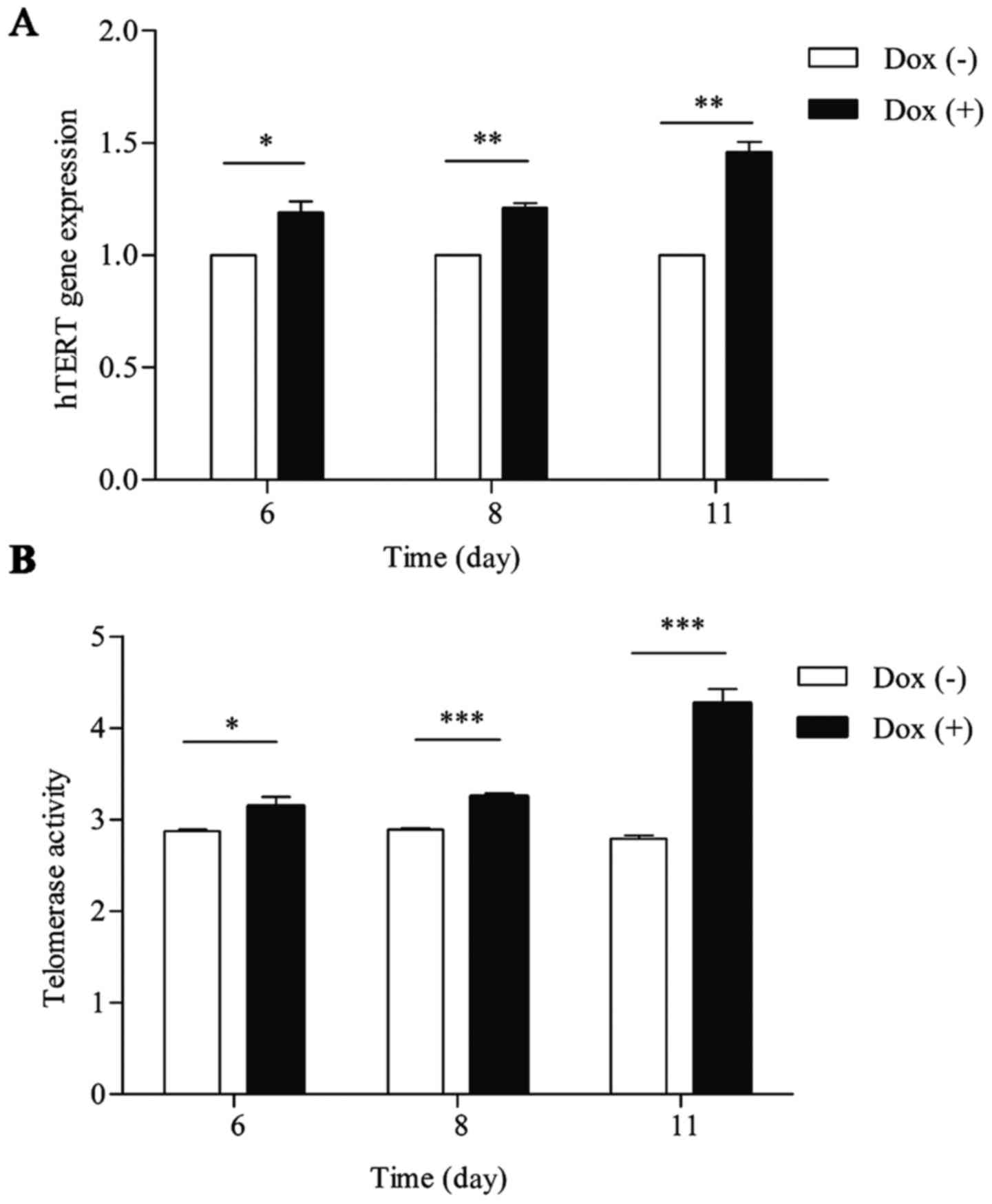
In aneuploid cells, hTERT gene expression
and telomerase activity gradually increased after doxycycline
washout
Telomerase regulation occurs mainly at the level of
hTERT transcription. We sought to investigate the alterations in
hTERT gene expression and telomerase activity that occurred in
aneuploid cells in response to doxycycline washout. At 6, 8 and 11
days after doxycycline washout, there was a gradual increase in
hTERT gene expression and telomerase activity in aneuploid cells,
and this increase peaked at 11 days after doxycycline washout
(1.46-fold vs. euploid cells; P<0.05), as shown in Fig. 1A. Telomerase activity peaked at 11
days after doxycycline washout (2.79 vs. 4.26 in euploid and
aneuploid cells, respectively; P<0.05), as shown in Fig. 1B. Taken together, we found that
hTERT gene expression and telomerase activity were higher in
aneuploid cells than in euploid cells. Because we found that the
differences in hTERT gene expression and telomerase activity
between aneuploid and euploid cells were most obvious at 11 days,
this time-point was used in the following tests.
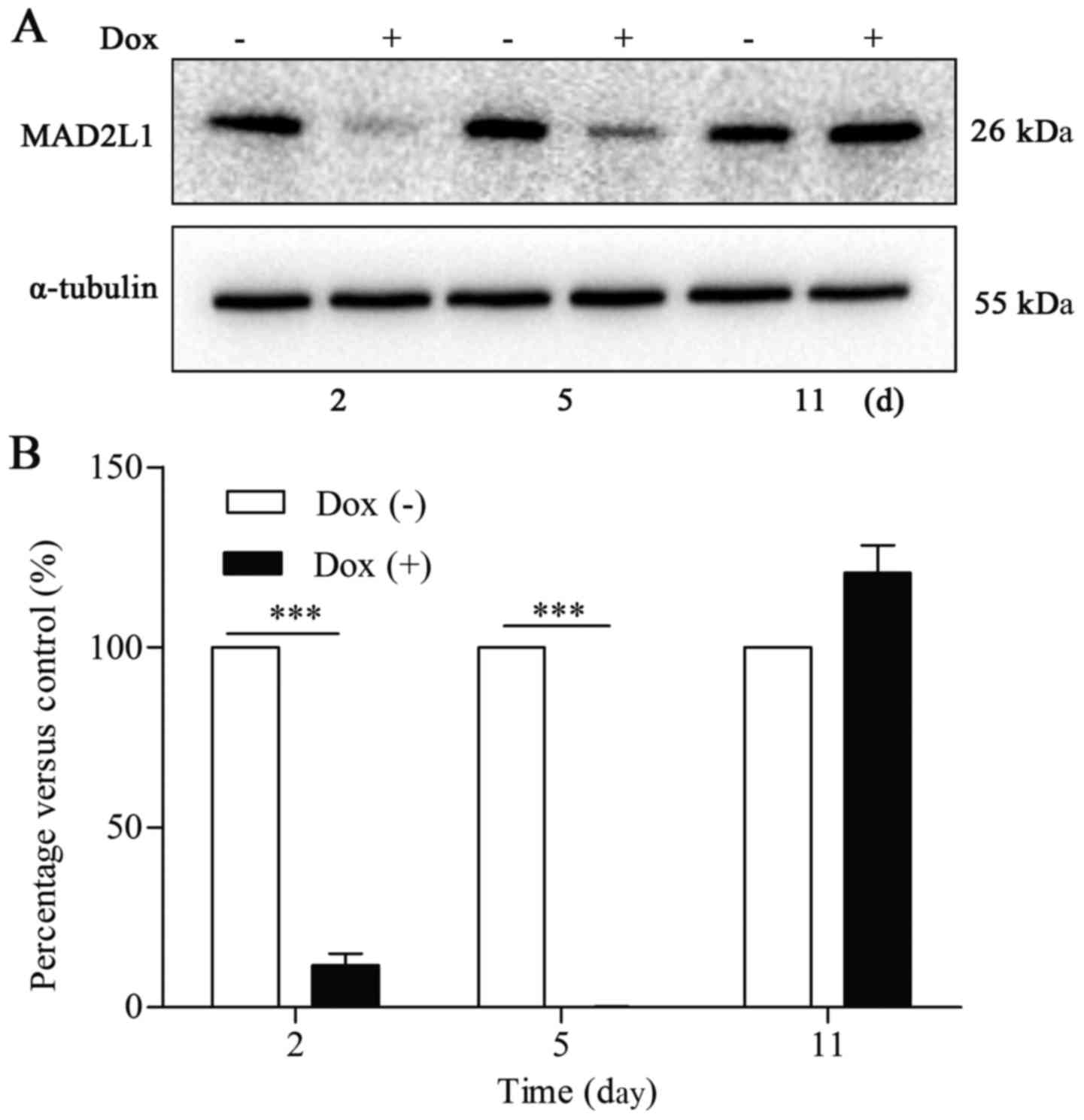
The rate of aneuploidy following MAD2
knockdown
At 11 days after doxycycline washout, a karyotyping
analysis of Dox (+) and Dox (−) groups was performed using
metaphase spreads. As shown in Table
II, the rate of aneuploidy was 112 out of 150 (74.67%) in the
Dox (+) group and 10 out of 150 (6.67%) in the Dox (−) group. We
next evaluated MAD2L1 protein expression in the Dox (+) and Dox (−)
groups at 2, 5, and 11 days after doxycycline washout. Western blot
analysis demonstrated that the protein level of MAD2L1 in aneuploid
cells had decreased at day 2, had gradually increased at day 5, and
had recovered to normal at day 11 after doxycycline washout. The
fact that MAD2L1 protein levels had recovered to normal at day 11
after washout demonstrated that Dox (+) cells are capable of
proliferating at 11 days after doxycycline washout, as shown in
Fig. 2.
| Table IIThe karyotyping analysis of euploid
and aneuploid group cells. |
Table II
The karyotyping analysis of euploid
and aneuploid group cells.
| Groups | Amounts | Percentage of
aneuploidy |
Chromosome
numbers of indicated karyotype
|
|---|
| 37 | 38 | 39 | 40 | 41 | 42 | 43 | 44 | 45 | 46 | 47 | 48 | 49 | 50 | 51 | 52 | 53 | 54 | 66 |
|---|
| Dox (−) | 150 | 6.67 | | | | | | 2 | 1 | 1 | 70 | 70 | 1 | 1 | 1 | 1 | | 2 | | | |
| Dox (+) | 150 | 74.67 | 2 | 3 | 3 | 6 | 8 | 12 | 11 | 19 | 12 | 26 | 17 | 8 | 7 | 5 | 4 | 2 | 1 | 2 | 2 |
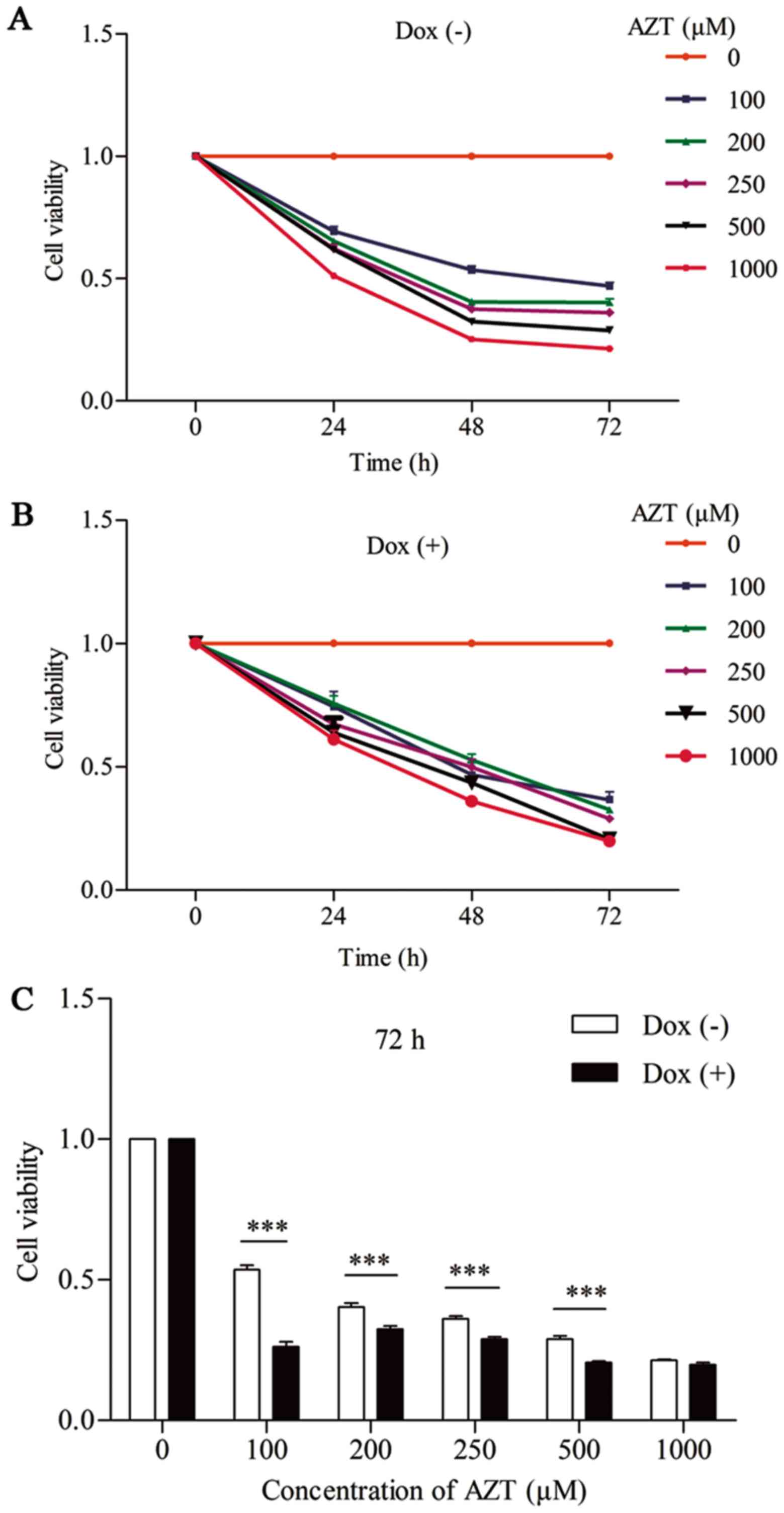
AZT inhibits cell proliferation in Dox
(+) and Dox (−) cells
The effect of different concentrations of AZT on
cell viability in Dox (+) and Dox (−) cells was determined using
CCK-8 assays. As shown in Fig. 3A and
B, viability was substantially decreased in a dose- and
time-dependent manner in Dox (+) and Dox (−) cells following
exposure to AZT. As shown in Fig.
3C, the cytotoxic impact of AZT was concentration-dependent.
Cell viability was decreased by ~53, 60, 64, 71 and 79% in Dox (−)
cells and by ~64, 68, 71, 79 and 80% in Dox (+) cells after 72 h of
exposure to 100, 200, 250, 500 or 1,000 µM of AZT,
respectively. The rate of reduction in cell viability was therefore
higher in Dox (+) cells than in Dox (−) cells. These data indicate
that the Dox (+) cells were more sensitive to the cytotoxic impact
of AZT than the Dox (−) cells.
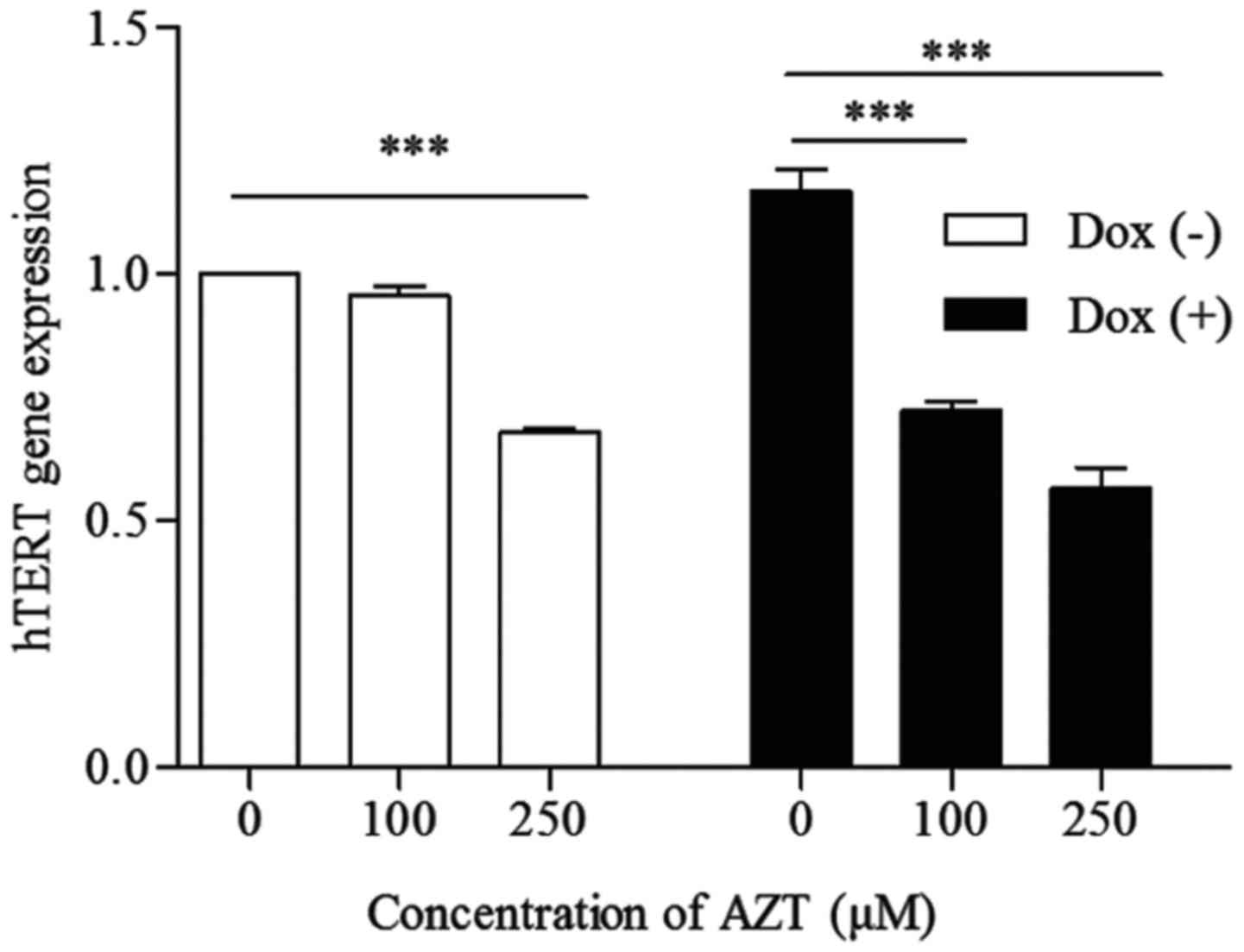
AZT downregulates the expression of hTERT
in Dox (+) and Dox (−) cells
Telomerase is regulated mainly by the level of hTERT
transcription. As shown in Fig. 4,
there was a significant decrease in hTERT gene expression following
72 h of treatment with 100 or 250 µM AZT in both the Dox (+)
and Dox (−) cells. This change in hTERT gene expression was
concentration-dependent and decreased by ~5 and 32% in the Dox (−)
cells and ~38.46 and 51.28% in the Dox (+) cells, respectively. The
rate of reduction in hTERT gene expression was higher in the Dox
(+) cells than in the Dox (−) cells (P<0.05), indicating that
the Dox (+) cells were more sensitive to AZT than the Dox (−)
cells.
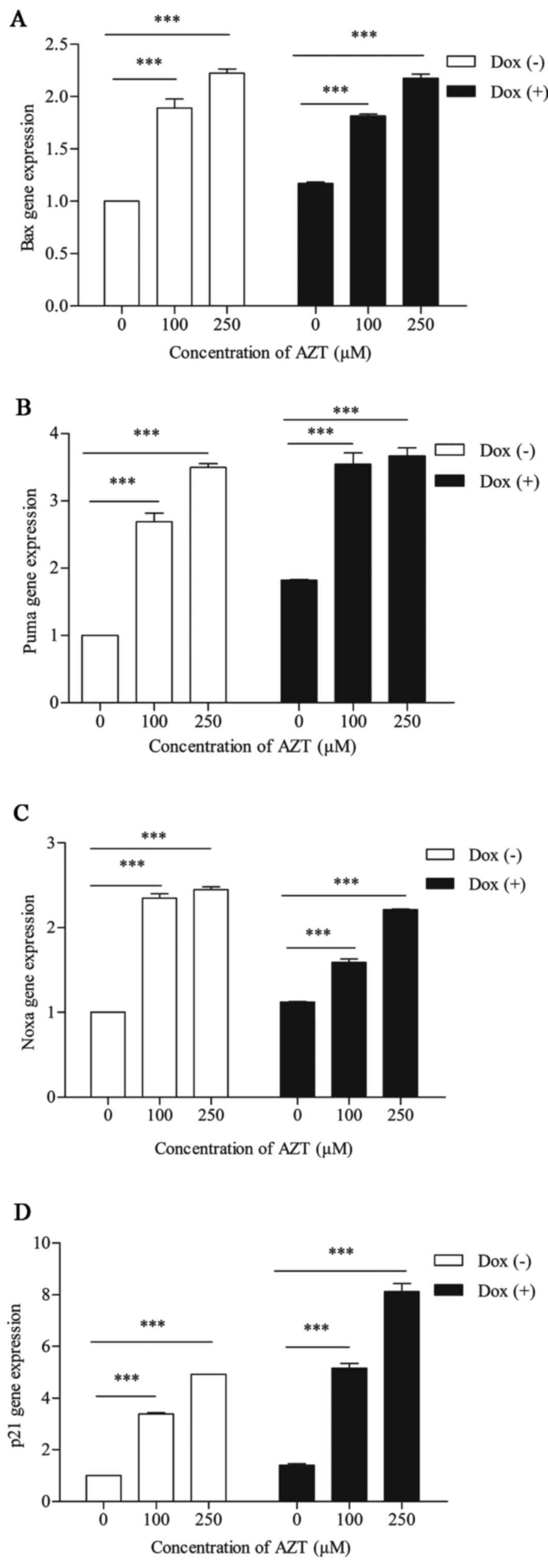
Effects of AZT on the roles of Puma, Bax,
Noxa and p21 in Dox (+) and Dox (−) cells
To determine the effect of AZT on apoptosis in the
two groups of cells, we first used RT-PCR to detect the mRNA
expression levels of p53-targeted genes (Puma, Bax, Noxa and p21).
We found that these genes were positively regulated by p53 and
played pro-apoptotic roles. The mRNA levels of these genes were
upregulated in a dose-dependent manner following exposure to 100 or
250 M AZT for 72 h, as shown in Fig.
5. The levels of Puma, Bax and Noxa gene expression following
exposure to 0, 100 and 250 µM AZT were 1:2.68:3.49,
1:1.89:2.22 and 1:2.34:2.44, respectively in the Dox (−) cells and
1:1.94:2.01, 1:1.54:1.85 and 1:1.42:1.97, respectively, in the Dox
(+) cells, and the differences between the responses of the two
groups of cells were significantly different (P<0.05). The level
of p21 gene expression following exposure to 0, 100 and 250
µM AZT for 72 h was 1:3.38:4.91 in the Dox (−) cells and
1:3.67:5.78 in the Dox (+) cells, and the differences in the
responses of the two lines of cells were significantly different
(P<0.05). These data indicate that the euploid cells were more
sensitive than the aneuploid cells to the induction of the
p53-Puma/Bax/Noxa pathways during AZT-induced apoptosis and that
the aneuploid cells were more sensitive than the euploid cells to
the induction of the p53-p21 pathways during AZT-induced cell cycle
arrest.
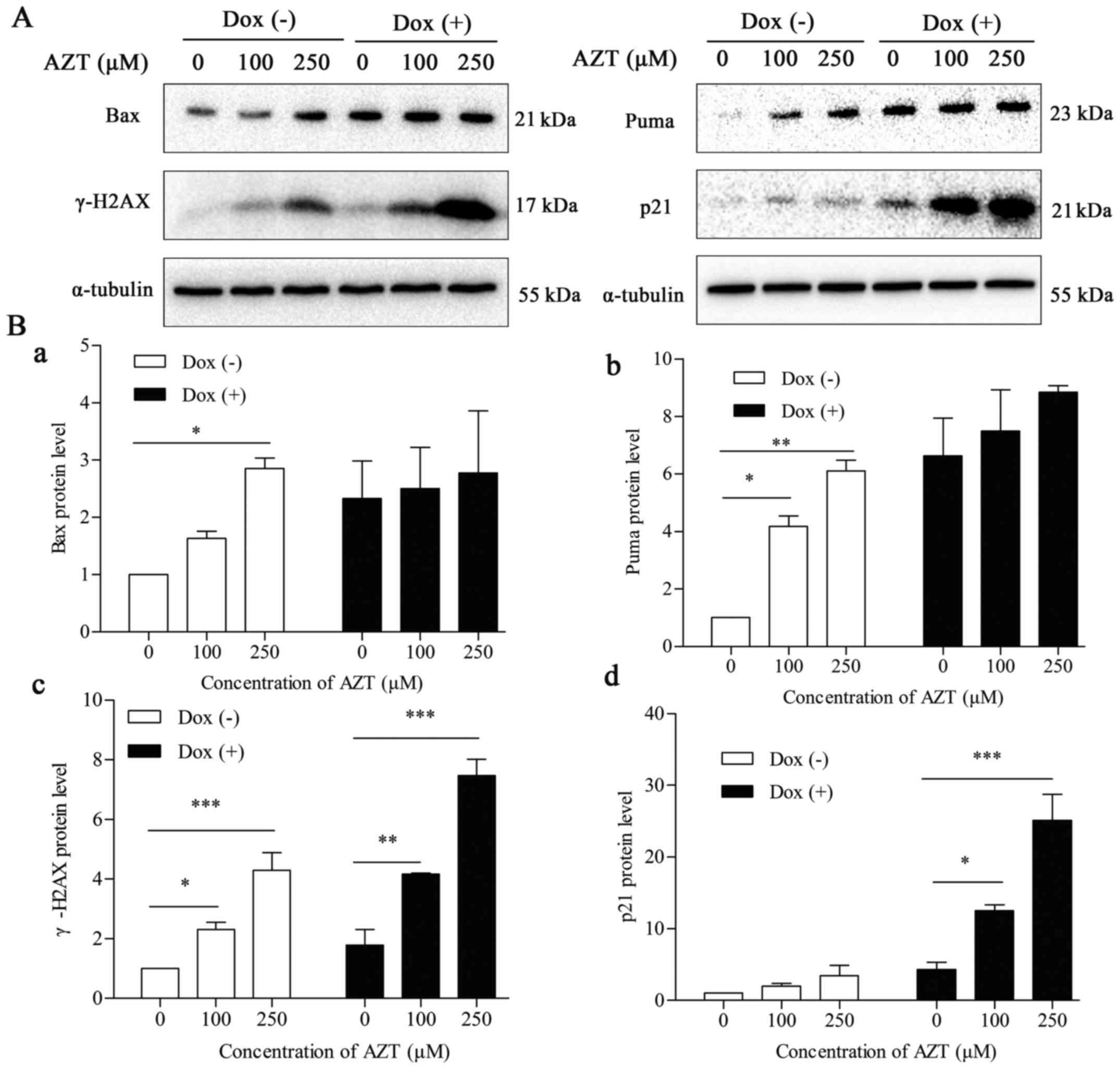
Effects of AZT on the roles of the Puma,
Bax, p21 and γ-H2AX proteins in aneuploid and euploid cells
Puma proteins are pro-apoptotic participants in the
Bcl-2 family that could impact the levels of apoptosis-related
proteins (36). Cell cycle arrest
occurs in G1 phase when the p53-p21-cip1 pathway is activated
(37). The phosphorylation of the
H2AX histone (γ-H2AX, phosphorylated histone H2AX on serine 139) is
an early indicator of DNA double-strand breaks (DSBs). We therefore
determined the protein level of the pro-apoptosis proteins Puma and
Bax, the cell cycle-arrest protein p21, and the DNA double-strand
break protein γ-H2AX using western blot analysis. As shown in
Fig. 6, the Puma, Bax, p21 and
γ-H2AX proteins were upregulated in a dose-dependent manner
following exposure to 100 and 250 µM AZT for 72 h in the Dox
(−) cells. In the Dox (+) cells, p21 and γ-H2AX were upregulated in
a dose-dependent manner following exposure to 100 and 250 µM
AZT for 72 h. However, Puma and Bax levels were not altered. The
relative protein levels of p21 and γ-H2AX in cells exposed to 0,
100 and 250 µM AZT were 1:2.23:3.94 and 1:1.97:3.43 in the
Dox (−) cells and 1:2.93:5.32 and 1:3.02:5.91 in the Dox (+) cells,
respectively. There was a significant difference in the levels of
the p21 and γ-H2AX proteins between the two groups of cells
following exposure to 0, 100 and 250 µM AZT (P<0.05). AZT
upregulated the level of Puma, Bax, p21 and γ-H2AX in the Dox (−)
cells and upregulated the levels of p21 and γ-H2AX in the Dox (+)
cells. Collectively, these findings demonstrate that the aneuploid
cells were more sensitive than the euploid cells to AZT-induced
cell cycle arrest (indicated by p53-p21 levels) and DNA
double-strand breaks (indicated by γ-H2AX levels), whereas euploid
cells were more sensitive than aneuploid cells to AZT-induced
apoptosis (indicated by p53-Puma/Bax/Noxa levels).
Discussion
The relationship between aneuploidy and
tumorigenesis has long been recognized (38). Some authors have suggested that
aneuploidy simply represents a side-effect of the course of tumor
formation, while others have argued that aneuploidy is associated
with oncogenic transformation. Studies of certain aneuploidy
karyotypes performed in mouse models of chromosomal instability
have suggested that aneuploidy can either promote or inhibit
tumorigenesis. Mouse trisomy for chromosome 16 is resistant to
adenomatous polyposis coli (APC)-induced colon cancer (39). Humans with chromosome 21 trisomy
are less likely to develop tumors than euploid individuals
(40). In contrast, trisomy 8
appears to increase the risk of hematopoietic malignancies. In
total, 5% of all cases of acute lymphoblastic leukemia (ALL),
10–15% of all cases of acute myeloid leukemia (AML) and 25% of all
cases of chronic myeloid leukemia (CML) have an extra copy of
chromosome 8 (39,41). It has been suggested that a low
frequency of chromosome mis-segregation accelerates tumorigenesis
by increasing the likelihood of producing a tumorigenesis-promoting
karyotype. When the rate of chromosome mis-segregation is too high,
tumor cells cannot survive with these types of tumor-promoting
karyotypes. Instead, cells with inviable karyotypes are constantly
produced, resulting in cell death and, consequentially, tumor
repression (42).
Chromosome mis-segregation can lead to aneuploidy.
The currently available data suggest that a series of biochemical
pathways enable the faithful segregation of sister chromosomes, and
malfunctions in these pathways can cause chromosomal
mis-segregation, for example, by disrupting the spindle assembly
checkpoint, inducing centrosome abnormalities, altering
microtubule-kinetochore dynamics, or causing defects in chromosome
cohesion (17). Short hairpin RNAs
(shRNAs) against GJB3, RXFP1, OSBPL3 and STARD9 were introduced
into BJ and IMR90 fibroblast cell lines as per Meena et al
to induce aneuploidy (27). These
4 candidate genes are directly associated with ploidy-affecting
pathways (27). For example, Li
et al (26) knocked down
MAD2L1 using siRNA to produce aneuploidy. In the present study, we
used a MAD21-targeting shRNA to generate an inducible expression
system that was triggered by doxycycline. MAD2L1 is a component of
the SAC that is necessary for proper sister chromatid segregation.
Defects in MAD2L1 can cause chromosome mis-segregation, leading to
the formation of aneuploidy (17).
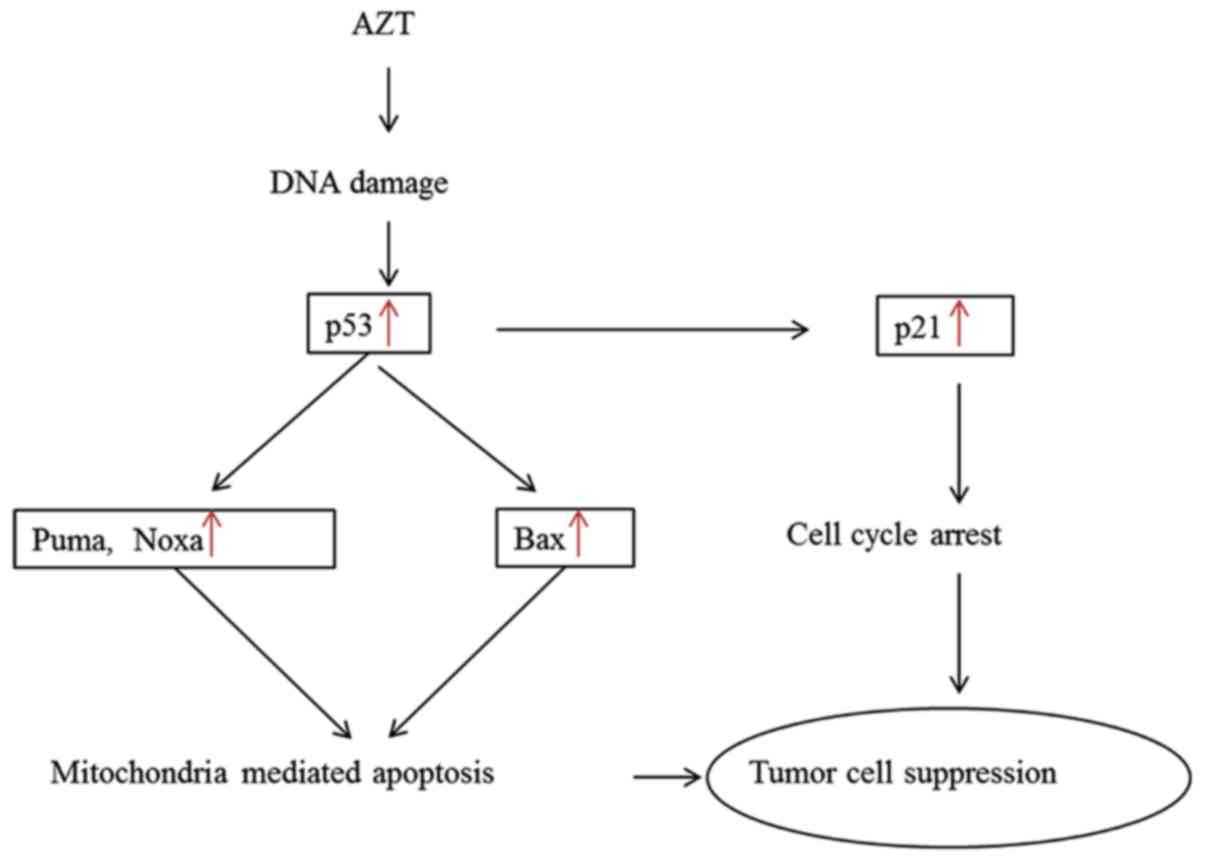
p53 is well-known for its role as the guardian of
the genome. It also plays a pivotal role in suppressing
tumorigenesis (43). Our
experimental data demonstrate that two p53-dependent pathways play
pivotal roles in this process, as shown in Fig. 7. We first showed that the DNA
damage caused by AZT in aneuploid and euploid cells is capable of
inducing the expression of the p53-Puma/Bax/Noxa pathways,
ultimately leading to cellular apoptosis. Second, p21 is widely
accepted as a transcriptional target of p53. Our data clearly
indicate that the p53-p21 pathway is linked to AZT-induced cell
cycle arrest. We found that aneuploid cells were more sensitive to
AZT-induced cell cycle arrest (p53-p21) and DNA double-strand
breaks (γ-H2AX), while euploid cells were more sensitive to
AZT-induced apoptosis (p53-Puma/Bax/Noxa). It has previously been
shown that aneuploidy plays dual roles in promoting or inhibiting
the process of tumor formation and that specific aneuploidy
karyotypes promote tumorigenesis. A doxycycline strategy was
utilized to disrupt SAC function and therefore trigger aneuploidy
in cells. Aneuploid cells exhibited a high rate of early cell death
but recovered after 11 days, before exposure to AZT. During this
early period, the aneuploid cells that were unfit for survival were
induced to undergo death, while those that survived aneuploidy were
likely to undergo tumorigenesis as a result of gaining a specific
chromosome karyotype that made them resistant to AZT-induced
apoptosis. Hence, the euploid cells were more sensitive to
AZT-induced apoptosis (p53-Puma/Bax/Noxa). Our data show that Puma
and Bax gene expression are correlated with relevant protein
expression levels in euploid cells but not in aneuploid cells.
Whether the process of translation is disrupted or the apoptotic
pathway differs between aneuploid and euploid cells remains
unknown.
Taken together, our data indicate that hTERT gene
expression and telomerase activity are higher in aneuploid cells
than in euploid cells. Both apoptosis (indicated by the
p53-Puma/Bax/Noxa pathway) and cell cycle arrest (indicated by the
p53-p21 pathway) were involved in AZT-induced cell death in both
aneuploid and euploid cells. The apoptotic pathway in aneuploid
cells requires additional study.
Acknowledgments
The present study was supported by the National
Natural Science Funds (81302016), the Clinical Medicine Science and
Technology Projects of Jiangsu Province (BL2012054) in China, the
National Natural Science Foundation of China (81402484), Jiangsu
Provincial Natural Science Foundation (BK20140497), the Natural
Science Fund for Colleges and Universities in Jiangsu Province
(14KJB310024), Jiangsu Provincial Medical Youth Talent Program
(QNRC2016322), and Yangzhou Municipal Natural Science Foundation
(YZ2014059). We thank Dr Pumin Zhang and Dr Guojun Lu at Baylor
College of Medicine (Houston, TX, USA) and Beijing Proteome
Research Center (Beijing, China) for providing us the plasmids.
References
|
1
|
Blackburn EH: Switching and signaling at
the telomere. Cell. 106:661–673. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chiodi I and Mondello C: Telomere and
telomerase stability in human diseases and cancer. Front Biosci
(Landmark Ed). 21:203–224. 2016. View
Article : Google Scholar
|
|
3
|
Shay JW and Wright WE: Senescence and
immortalization: Role of telomeres and telomerase. Carcinogenesis.
26:867–874. 2005. View Article : Google Scholar
|
|
4
|
Verdun RE and Karlseder J: Replication and
protection of telomeres. Nature. 447:924–931. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang N, Li J, Zhong X, An X and Hou J:
Reversible immortalization of sheep fetal fibroblast cells by
tetracycline-inducible expression of human telomerase reverse
transcriptase. Biotechnol Lett. 38:1261–1268. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wieser M, Stadler G, Jennings P, Streubel
B, Pfaller W, Ambros P, Riedl C, Katinger H, Grillari J and
Grillari-Voglauer R: hTERT alone immortalizes epithelial cells of
renal proximal tubules without changing their functional
characteristics. Am J Physiol Renal Physiol. 295:F1365–F1375. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kapanadze B, Morris E, Smith E and
Trojanowska M: Establishment and characterization of scleroderma
fibroblast clonal cell lines by introduction of the hTERT gene. J
Cell Mol Med. 14:1156–1165. 2010.
|
|
8
|
Gu T, Wang X, Wang X, Wang W, Liu Y, Zhang
B, Shi Y, Zhang Z, Sun Q, Xue T, et al: The value of detecting
telomerase activity on early diagnosis of lung cancer. Zhongguo Fei
Ai Za Zhi. 4:37–40. 2001.In Chinese. PubMed/NCBI
|
|
9
|
Cavallo D, Casadio V, Bravaccini S,
Iavicoli S, Pira E, Romano C, Fresegna AM, Maiello R, Ciervo A,
Buresti G, et al: Assessment of DNA damage and telomerase activity
in exfoliated urinary cells as sensitive and noninvasive biomarkers
for early diagnosis of bladder cancer in ex-workers of a rubber
tyres industry. Biomed Res Int. 2014:3709072014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Glybochko PV, Zezerov EG, Glukhov AI,
Alyaev YG, Severin SE, Polyakovsky KA, Varshavsky VA, Severin ES
and Vinarov AZ: Telomerase as a tumor marker in diagnosis of
prostatic intraepithelial neoplasia and prostate cancer. Prostate.
74:1043–1051. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harley CB: Telomerase and cancer
therapeutics. Nat Rev Cancer. 8:167–179. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Low KC and Tergaonkar V: Telomerase:
Central regulator of all of the hallmarks of cancer. Trends Biochem
Sci. 38:426–434. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gordon DJ, Resio B and Pellman D: Causes
and consequences of aneuploidy in cancer. Nat Rev Genet.
13:189–203. 2012.PubMed/NCBI
|
|
14
|
Pfau SJ and Amon A: Chromosomal
instability and aneuploidy in cancer: From yeast to man. EMBO Rep.
13:515–527. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Holland AJ and Cleveland DW: Losing
balance: The origin and impact of aneuploidy in cancer. EMBO Rep.
13:501–514. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Musacchio A and Salmon ED: The
spindle-assembly checkpoint in space and time. Nat Rev Mol Cell
Biol. 8:379–393. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fang X and Zhang P: Aneuploidy and
tumorigenesis. Semin Cell Dev Biol. 22:595–601. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sheltzer JM: A transcriptional and
metabolic signature of primary aneuploidy is present in
chromosomally unstable cancer cells and informs clinical prognosis.
Cancer Res. 73:6401–6412. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dürrbaum M, Kuznetsova AY, Passerini V,
Stingele S, Stoehr G and Storchová Z: Unique features of the
transcriptional response to model aneuploidy in human cells. BMC
Genomics. 15:1392014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Beroukhim R, Mermel CH, Porter D, Wei G,
Raychaudhuri S, Donovan J, Barretina J, Boehm JS, Dobson J,
Urashima M, et al: The landscape of somatic copy-number alteration
across human cancers. Nature. 463:899–905. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Munné S, Bahçe M, Sandalinas M, Escudero
T, Márquez C, Velilla E, Colls P, Oter M, Alikani M and Cohen J:
Differences in chromosome susceptibility to aneuploidy and survival
to first trimester. Reprod Biomed Online. 8:81–90. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Williams BR, Prabhu VR, Hunter KE, Glazier
CM, Whittaker CA, Housman DE and Amon A: Aneuploidy affects
proliferation and spontaneous immortalization in mammalian cells.
Science. 322:703–709. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Davoli T, Xu AW, Mengwasser KE, Sack LM,
Yoon JC, Park PJ and Elledge SJ: Cumulative haploinsufficiency and
triplosensitivity drive aneuploidy patterns and shape the cancer
genome. Cell. 155:948–962. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Baker DJ and van Deursen JM: Chromosome
missegregation causes colon cancer by APC loss of heterozygosity.
Cell Cycle. 9:1711–1716. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schvartzman JM, Sotillo R and Benezra R:
Mitotic chromosomal instability and cancer: Mouse modelling of the
human disease. Nat Rev Cancer. 10:102–115. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li M, Fang X, Baker DJ, Guo L, Gao X, Wei
Z, Han S, van Deursen JM and Zhang P: The ATM-p53 pathway
suppresses aneuploidy-induced tumorigenesis. Proc Natl Acad Sci
USA. 107:14188–14193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Meena JK, Cerutti A, Beichler C, Morita Y,
Bruhn C, Kumar M, Kraus JM, Speicher MR, Wang ZQ, Kestler HA, et
al: Telomerase abrogates aneuploidy-induced telomere replication
stress, senescence and cell depletion. EMBO J. 34:1371–1384. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gomez DE, Armando RG and Alonso DF: AZT as
a telomerase inhibitor. Front Oncol. 2:1132012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhihua Z, Miaomiao S, Jianping Y and
Kuisheng C: Research for the influence of telomerase inhibitors on
myeloma cell and therapy. Pak J Pharm Sci. 27(Suppl): 1641–1644.
2014.PubMed/NCBI
|
|
30
|
He M, Jiang YY, Zhu M, Wei X, Qin J, Zhang
ZY and Li L: Effects of 3′-azido-deoxythymidine on telomerase
activity and protein expression of hepatocarcinoma cell line
SMMC-7721. Ai Zheng. 25:543–548. 2006.In Chinese. PubMed/NCBI
|
|
31
|
Souza Sobrinho CP, Gragnani A, Santos ID,
Oliveira AF, Lipay MV and Ferreira LM: AZT on telomerase activity
and cell proliferation in HS 839. T melanoma cells Acta Cir Bras.
27:855–860. 2012. View Article : Google Scholar
|
|
32
|
Brown T, Sigurdson E, Rogatko A and
Broccoli D: Telomerase inhibition using azidothymidine in the HT-29
colon cancer cell line. Ann Surg Oncol. 10:910–915. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stedt H, Samaranayake H, Kurkipuro J,
Wirth G, Christiansen LS, Vuorio T, Määttä AM, Piškur J and
Ylä-Herttuala S: Tomato thymidine kinase-based suicide gene therapy
for malignant glioma - an alternative for Herpes Simplex virus-1
thymidine kinase. Cancer Gene Ther. 22:130–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sabokrouh A, Goodarzi MT, Vaisi-Raygani A,
Khatami S and Taghizadeh-Jahed M: Effects of treatment with
platinum azidothymidine and azidothymidine on telomerase activity
and bcl-2 concentration in hepatocellular carcinoma-induced rats.
Avicenna J Med Biotechnol. 6:200–209. 2014.PubMed/NCBI
|
|
35
|
Kim NW and Wu F: Advances in
quantification and characterization of telomerase activity by the
telomeric repeat amplification protocol (TRAP). Nucleic Acids Res.
25:2595–2597. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Junttila MR and Evan GI: p53 - a Jack of
all trades but master of none. Nat Rev Cancer. 9:821–829. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yu Z, Wang H, Zhang L, Tang A, Zhai Q, Wen
J, Yao L and Li P: Both p53-PUMA/NOXA-Bax-mitochondrion and
p53-p21cip1 pathways are involved in the CDglyTK-mediated tumor
cell suppression. Biochem Biophys Res Commun. 386:607–611. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Boveri T: Concerning the origin of
malignant tumours by Theodor Boveri. Translated and annotated by
Henry Harris. J Cell Sci. 121(Suppl 1): 1–84. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Paulsson K and Johansson B: Trisomy 8 as
the sole chromosomal aberration in acute myeloid leukemia and
myelodysplastic syndromes. Pathol Biol (Paris). 55:37–48. 2007.
View Article : Google Scholar
|
|
40
|
Baek KH, Zaslavsky A, Lynch RC, Britt C,
Okada Y, Siarey RJ, Lensch MW, Park IH, Yoon SS, Minami T, et al:
Down's syndrome suppression of tumour growth and the role of the
calcineurin inhibitor DSCR1. Nature. 459:1126–1130. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jones L, Wei G, Sevcikova S, Phan V, Jain
S, Shieh A, Wong JC, Li M, Dubansky J, Maunakea ML, et al: Gain of
MYC underlies recurrent trisomy of the MYC chromosome in acute
promyelocytic leukemia. J Exp Med. 207:2581–2594. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Santaguida S and Amon A: Short- and
long-term effects of chromosome mis-segregation and aneuploidy. Nat
Rev Mol Cell Biol. 16:473–485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xie C, Subhash VV, Datta A, Liem N, Tan
SH, Yeo MS, Tan WL, Koh V, Yan FL, Wong FY, et al: Melanoma
associated antigen (MAGE)-A3 promotes cell proliferation and
chemotherapeutic drug resistance in gastric cancer. Cell Oncol
(Dordr). 39:175–186. 2016. View Article : Google Scholar
|