Introduction
Ribosome-inactivating proteins (RIPs), which are
widely distributed among plants, fungi, algae and bacteria, inhibit
protein synthesis by irreversibly damaging ribosomes, which in turn
leads to cell death (1,2). RIPs are classified into three types
based on structural features. Type I RIPs, including trichosanthin
(TCS), saporin and curcin, are comprised of a single A-chain with
RNA N-glycosidase activity (3–6).
Type II RIPs are comprised of an A-chain linked to a lectin B-chain
by a disulphide bond, the most well-known of which is ricin
(7). Type III RIPs are considered
to be pro-RIPs, which require proteolytic cleavage to become active
proteins (8). In the last 10
years, RIPs, including momordica (exhibiting anti-human
immunodeficiency virus activity) (9), TCS, saporin, curcin and Abrus
agglutinin (exhibiting anti-neoplastic activity) (5,6,10,11),
have been shown to offer potential as therapeutic agents in
medicine based on their biological activities.
Vernicia fordii, which belongs to the
Euphorbiaceae family, is widely distributed in tropical and
sub-tropical regions of Asia. The leaves, roots and seeds of V.
fordii have been used as a traditional Chinese drug owing to
its anti-inflammatory and antiviral effects (12). Based on previous literature,
bioactive compounds with low molecular weights, including
α-eleostearic acid, exhibiting anti-inflammatory activity (13), and conjugated linoleic acid,
exhibiting cytotoxic (14) and
hypoglycemic activity (15), have
been extracted from V. fordii; however, the bioactive
protein from this plant remains to be fully elucidated. In our
previous study, the transcriptome of V. fordii was sequenced
to better understand the molecular basis underlying the development
of the V. fordii bioactive protein (16). Based on the transcriptome analysis,
presence of the RIP gene was confirmed in V. fordii. A
number of RIP-containing plants are reported in the pharmacopoeia
of folk medicine, mostly owing to their toxicity (17). The presence of RIP may contribute
to this activity. In the present study, fordin, a novel type I RIP,
was identified from V. fordii. The antitumor activity of
fordin was evaluated in vitro by determining its
cytotoxicity against human tumor cell lines (U-2 OS, HepG2, HeLa
and A549) and the normal MRC-5 cell line. Although the anticancer
potential of RIPs has been investigated by various groups (18–20),
the mechanism underlying RIP cytotoxicity remains to be elucidated.
Numerous studies have reported that RIPs exert anticancer
activities by inhibiting survival and inducing apoptosis in cancer
cells (4,10,21).
In the present study, it was shown that fordin also inhibited the
invasion and migration of U-2 OS and HepG2 cells.
Nuclear factor (NF)-κB consists of dimers containing
five members of the Rel protein family (p65, p50/p105, p52/p100,
Rel B and c-Rel). Inactivated NF-κB is sequestered in the cytosol
by binding with inhibitor of NF-κB (IκB)s (22). When IκBs are phosphorylated by the
IκB kinase (IKK) complex, activated NF-κB is released and
translocated into the nucleus to modulate the expression of several
genes involved in processes including cell growth and cell death
(22). It is reported that NF-κB
is key in regulation of the B-cell lymphoma-2 (Bcl-2) (23–25)
and matrix metalloproteinase (MMP) (26) families, both of which have been
confirmed to be important in cell apoptosis and invasion. It has
also been reported that RIPs induce apoptosis in cancer cells via
the downregulation of anti-apoptotic proteins, including Bcl-2
and/or Bcl-extra large (Bcl-xL) and/or the upregulation of
pro-apoptotic proteins, including Bcl-2-associated X protein (Bax)
and/or Bcl-2-associated death promoter (Bad) (4,21).
In this regard, it is important to clarify the role of NF-κB in the
multiple anticancer effects of fordin.
In the present study, the effects of fordin on cell
proliferation, apoptosis, invasion and migration were investigated.
In addition, changes in the expression of key proteins relevant to
apoptosis and metastasis in U-2 OS and HepG2 cells were determined
in an effort to better understand the molecular mechanisms
underlying the multiple anticancer effects of fordin in cancer
cells.
Materials and methods
Molecular cloning
Total RNA was extracted from the fresh leaves of
V. fordii (collected from the test field at Hefei Institutes
of Physical Science, Chinese Academy of Science, Hefei, China)
using an E. Z. N. A. Plant RNA kit (OMEGA Biotek, Inc., Norcross,
GA, USA). Subsequently, RNA samples with an optical density (OD)
260/280 wavelength ratio between 2.0 and 2.2 were used for primary
cDNA synthesis with a GoScript Reverse Transcription kit (Promega
Corporation, Madison, WI, USA). Specific primers were designed
based on the V. fordii RNA-Seq database (accession no.
GSE98631) to obtain V. fordii (vf)-RIP and its mature
peptide (fordin), as follows: vf-RIP, forward,
5′-ATTAGAAACCATCTCCTCCCTCT-3′ and reverse,
5′-CTTAGTTTGAAGCCCTTCTACTT-3′; fordin, forward, 5′-AATGGATCCAGAGTTAGCCCTTTACTA-3′ and
reverse, 5′-GGGAAGCTTCTACTTCTTTACATACAGC-3′.
The underlined bases in the primers were designated as BamHI
and HindIII restriction sites, respectively. The polymerase
chain reaction (PCR) was performed in 50-µl reaction
mixtures (Takara Biotechnology Co., Ltd., Dalian, China), each
mixture included: 0.5 µl PrimeSTAR HS DNA Polymerase, 10
µl 5X PCR Buffer, 4 µl cDNA, 4 µl dNTPs, 2
µl primers, and 29.5 µl ddH2O, according
to the following procedure: 94°C for 2 min; 30 cycles at 98°C for
10 sec, 58°C for 90 sec, and 68°C for 45 sec; and final extension
at 68°C for 10 min. The PCR products were analyzed by 1.2% agarose
electrophoresis. The fragments of expected size were purified with
an Agarose Gel Extraction kit (OMEGA Biotek, Inc.), and then cloned
into the pEASY-T1 vector (Beijing TransGen Biotech Co., Ltd.,
Beijing, China) for sequencing. Multiple sequence alignment was
performed using DNAMAN software 8.0 (LynnonBiosoft Bioinformatics
solutions, San Ramon, CA, USA).
Expression and purification of
fordin
The coding sequence of fordin was cloned into the
pET28b-SUMO vector, constructed by Professor Fan (University of
Anhui Agriculture, Anhui, China) (27,28).
The recombinant plasmid (pSUMO-Fordin) was transformed into
Escherichia coli Rosetta host strain (DE3) competent cells
(Novagen/Merck, Darmstadt, Germany). E. coli harboring the
recombinant plasmid was incubated with shaking at 200 rpm at 37°C
until the OD 600 of the culture reached 0.8. The expression of
SUMO-Fordin was induced by the addition of 0.5 mM IPTG. Following
further incubation at 16°C for 24 h, the cells were harvested and
sonicated in binding buffer containing 50 mM
NaH2PO4 (pH 8.0) and 300 mM NaCl. The
cell-free supernatant was applied to a Ni2+ resin column
(2.0×15 cm). The resin was washed with the binding and washing
buffers, containing 50 mM NaH2PO4, (pH 8.0),
300 mM NaCl, and 20 mM imidazole. The fusion protein was eluted
with elution buffer containing 50 mM NaH2PO4
(pH 8.0), 300 mM NaCl and 250 mM imidazole. To cleave the His-SUMO
tag by protease ULP, the fusion protein was then processed with
buffer through ultrafiltration columns (EMD Millipore, Billerica,
MA, USA) with membrane cartridges having molecular weight cut-offs
of 3,000 Da. The cleaved protein (fordin) was purified by
Ni2+ resin again. Finally, fordin was pooled and
measured using a BCA protein assay kit (Sangon Biotech Co., Ltd.,
Shanghai, China). The purity of fordin was analyzed by 12% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
Western blot analysis using rabbit polyclonal antibodies specific
for 6X His-tag (1:1000, cat. no. D110002; Sangon Biotech Co., Ltd.)
was performed to confirm the presence of the fusion protein. The
identification of fordin was assessed by liquid chromatography-mass
spectrometry with a MALDI TOF/TOF analyzer (ABI; Thermo Fisher
Scientific, Inc., Waltham, MA, USA).
RNA N-glycosidase activity assay
The assay was performed according to the method
described by Wu et al (29)
with several revisions. Total RNA was extracted from Sapium
sebiferum leaves (collected from the test field at Hefei
Institutes of Physical Science) by the method mentioned above.
Subsequently, 3 µg total RNA was treated with 0.4 µg
of fordin. The mixture was incubated at 30°C for 30 min, and then
treated with 2 M fresh aniline/acetate (pH 4.5). RNA was recovered
by ethanol precipitation and examined by 2.0% agarose
electrophoresis.
Cell culture
Human osteosarcoma (U-2 OS), hepatoblastoma (HepG2)
(30), cervical carcinoma (HeLa),
lung carcinoma (A549) and fetal lung fibroblast (MRC-5) cell lines
were purchased from the American Type Culture Collection (Manassas,
VA, USA) and cultured in DME/F-12 medium (GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS; BioWest, Miami, FL, USA). The cells were incubated in a
humidified atmosphere with 5% CO2 at 37°C.
Cell viability assay
The cell viability was assessed using a Cell
Counting Kit-8 (CCK-8) assay (Sangon Biotech, Co., Ltd.). The cell
suspension (100 µl) was dispensed (7×103
cells/well) into 96-well plates and exposed to various
concentrations of fordin (0.2–10 µM) for 24, 48 and 72 h.
CCK-8 solution was added (10 µl/well) and incubated for 30
min. The absorbance was measured at 450 nm using a microplate
reader (Molecular Devices LLC, Sunnyvale, CA, USA). Cell viability
was calculated as the percentage of untreated controls. Inhibitory
concentrations (ICs) of fordin were calculated using SPSS 16.0
software (SPSS, Inc., Chicago, IL, USA). In a subsequent
experiment, to avoid antiproliferative effects, relatively low
concentrations of fordin (1 µM and IC20) were
used for investigating the scratch-healing and invasive behavior of
U-2 OS and HepG2 cells. Low (IC20), medium
(IC50), and high (IC70) fordin concentrations
were used to evaluate concentration-dependent effects on U-2 OS and
HepG2 cells via nuclear staining, flow cytometry, and reverse
transcription-quantitative PCR (RT-qPCR) and western blot
analyses.
Scratch assay
The U-2 OS and HepG2 cells were seeded into 6-well
plates. The confluent monolayers of cells were scratched using
200-µl sterile pipette tips. Following removal of the
floating cells, the cells were cultured in medium without serum.
The effect of fordin (1 µM and IC20) on the
scratch-healing response was determined under an inverted
microscope (Olympus, Tokyo, Japan). A total of six randomly
selected images were selected per sample at 0 and 24 h.
Cell invasion assay
The effect of fordin on cell invasion ability was
investigated using a Transwell assay. Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA) diluted 1:8 in cold serum-free culture
media was added to each 24-well Transwell chamber with an
8-µm pore polycarbonate membrane (Corning Incorporated,
Corning, NY, USA) and incubated under standard conditions for at
least 12 h to gel. The cells treated with fordin (1 µM and
IC20) for 24 and 48 h were seeded onto the Matrigel.
Each lower chamber contained 600 µl of culture media
containing 10% FBS. Following incubation for 24 h, transmembrane
cells were stained by 0.1% crystal violet. Six fields were randomly
selected under the inverted fluorescence microscope, and the number
of transmembrane cells was counted.
Nuclear staining
The selected cells were seeded into 12-well plates
containing sterilized coverslips. Following treatment with fordin
(IC20, IC50, or IC70) for 24 h,
the cells were fixed with 4% paraformaldehyde, and stained with 4
µg/ml of Hoechst 33342 solution (Invitrogen; Thermo Fisher
Scientific, Inc.). The coverslips with cells were inverted onto
slides. Six random images were captured with the inverted
fluorescent microscope.
Annexin V/propidium iodide (PI)
analysis
Following treatment with fordin (IC20,
IC50, or IC70) for 24 h, the U-2 OS and HepG2
cells were harvested, and the extent of apoptosis was determined
through flow cytometry using an Annexin V-FITC/Dead Cell Apoptosis
kit (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Cell apoptosis was analyzed in triplicate
for each treatment using a FACSCalibur platform (BD Biosciences)
and FlowJo software 7.6 (Tree Star, Inc., Ashland, OR, USA).
Small interfering (si)RNA
transfection
NF-κB/p65 siRNA was purchased from GenePharma
(Shanghai, China). Non-silencing siRNA was used as the negative
control. U-2 OS and HepG2 cells were transfected with p65 siRNA or
control siRNA using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for 48 h according to the manufacturer's
protocol. Following transfection, the cells were treated with or
without fordin (IC50) for 24 h. Subsequently, certain
proteins involved in the NF-κB signaling pathway were analyzed by
western blot analysis.
RT-qPCR analysis
Total RNA was extracted using the E. Z. N. A RNA kit
from cells treated with fordin (IC20, IC50,
or IC70) for 24 h. cDNA corresponding to 1 µg of
total RNA was used to examine the mRNA levels of the selected genes
by a Light Cycler 96 Real-Time PCR system (Roche Diagnostics GmbH,
Mannheim, Germany). FastStart Essential DNA Green Master mix (Roche
Diagnostics GmbH) was applied for the RT-qPCR analysis. Each
reaction mixture included 10 µl 2x Master Mix, 1 µl
cDNA, 0.8 µl primers, and 8.2 µl ddH2O.
The reactions were performed according to the following conditions:
94°C for 30 sec, 40 cycles at 94°C for 5 sec, the optimal
temperature for primers of the selected genes (Table I) for 15 sec and 72°C for 10 sec.
After 40 cycles, the dissociation curve was determined. All samples
were performed as triplicate. The 2−ΔΔCq method
(31) was applied to calculate the
relative mRNA expression of the selected genes using GAPDH to
normalize the data.
 | Table INucleotide sequences of the primers
for reverse transcription-quantitative polymerase chain reaction
analysis. |
Table I
Nucleotide sequences of the primers
for reverse transcription-quantitative polymerase chain reaction
analysis.
| Gene | Forward primer
(5′–3′) | Reverse primer
(5′–3′) | Amplification
length (bp) |
|---|
| Bcl-2 |
CGGTGGGGTCATGTGTGTG |
CGGTTCAGGTACTCAGTCATCC | 90 |
| Bax |
TCATGGGCTGGACATTGGAC |
GAGACAGGGACATCAGTCGC | 114 |
| Bfl-1 |
TACAGGCTGGCTCAGGACTAT |
CGCAACATTTTGTAGCACTCTG | 96 |
| Bcl-xL |
GAGCTGGTGGTTGACTTTCTC |
TCCATCTCCGATTCAGTCCCT | 138 |
| MMP-2 |
AGGGAATGAATACTGGATCTACT |
GCTCCAGTTAAAGGCGGCATCCAC | 119 |
| MMP-9 |
GGGACGCAGACATCGTCATC |
TCGTCATCGTCGAAATGGGC | 139 |
| Timp-3 |
ACCGAGGCTTCACCAAGATG |
GGGGTCTGTGGCATTGATGA | 125 |
| uPA |
TCAAAAACCTGCTATGAGGGGA |
GGGCATGGTACGTTTGCTG | 121 |
| uPAR |
TATTCCCGAAGCCGTTACCTC |
TCGTTGCATTTGGTGGTGTTG | 275 |
| Cathepsin B |
CTGTCGGATGAGCTGGTCAAC |
TCGGTAAACATAACTCTCTGGGG | 152 |
| IKKα |
AAGTTGAACCATGCCAATGTTGT |
TCTCCTCCAGAACAGTATTCCAT | 107 |
| IKKβ |
CACCATCCACACCTACCCTG |
CTTATCGGGGATCAACGCCAG | 136 |
| NEMO |
CTTCCAAGAATACGACAACCACA |
CGGAACGGTCTCCATCACAAT | 187 |
| GAPDH |
GCACCGTCAAGGCTGAGAAC |
TGGTGAAGACGCCAGTGGA | 138 |
Western blot analysis
Following the various treatments, the cells were
maintained in lysis buffer for 10 min on ice for preparing the
whole cell lysates. Nuclear and cytoplasmic extracts were prepared
using NE-PER™ nuclear and cytoplasmic extraction reagents (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
The protein concentrations were measured using a BCA Protein Assay
kit. Equal quantities (50 µg) of protein were separated on a
12% SDS-PAGE gel and then transferred onto a PVDF membrane (EMD
Millipore), which was then blocked in 5% skim milk for 1 h at room
temperature. The membrane was then incubated overnight with primary
antibodies at 4°C. The primary antibodies were as follows:
Anti-Bcl-2-associated X protein (Bax; 1:2,000, cat. no. sc-70408)
and anti-Bcl-2 (1:1,000, cat. no. sc-130308) from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). The other primary antibodies
were anti-Bcl-2-related protein A1 (Bfl-1; 1:200, cat. no.
ab45413), anti-MMP-2 (1:500, cat. no. ab92536), anti-MMP-9 (1:500,
cat. no. ab76003), anti-p65 [(phosphorylated S536; 1:1,000, cat.
no. ab76302) and total (1:1,000, cat. no. ab76311)], anti-IKKα
(1:1,000, cat. no. ab32518), anti-IKKβ (1:1,000, cat. no.
ab124957), and anti-IκB [(phosphorylated S36; 1:1,000, cat. no.
ab133462) and total (1:1,000, cat. no. ab32518)], all of which were
purchased from Abcam (Cambridge, MA, USA). The membrane was
incubated with the secondary antibody (goat anti-mouse IgG-HRP,
1:1,000, cat. no. sc-2005; goat anti-rabbit IgG-HRP, 1:1,000, cat.
no. sc-2004; Santa Cruz Biotechnology, Inc.) for 2 h at room
temperature and visual-ized by chemiluminescence. β-actin and
Histone H3 were used as internal controls for the whole cell
lysates, cytosolic extracts and nuclear extracts, respectively.
Statistical analysis
The statistical analysis was performed using
GraphPad Prism 6 software (GraphPad Software, Inc., La Jola CA,
USA). All data are expressed as the mean ± standard deviation.
Statistical analysis between groups was performed by one-way
analysis of variance followed by the LSD post hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Molecular cloning and expression of
fordin
The coding sequence of the vf-RIP gene
(GenBank accession no. MF521485) is 840 bp in length with
translation initiation (ATG) and stop (TAG) codons. The open
reading frame encodes a 279-amino acid polypeptide, of which the
putative molecular mass is 30.8 kDa. An endoplasmic reticulum
signal peptide (residues 1–21) was identified, followed by the
mature peptide (fordin; residues 22–279) with a molecular mass of
28.3 kDa (Fig. 1). The results of
sequence alignment showed that fordin showed similarity with other
type I RIPs, including TCS (from Trichosanthes kirilowi),
curcin (from Jatropha curcas), momorcharin (from
Momordica charantia) and ricin A-chain (from Ricinus
communis) with respect to the conserved region. The fordin
amino acid residues, Tyr-109 (Y), Tyr-148 (Y), Glu-197 (E), Arg-200
(R), and Trp-229 (W), aligned perfectly with the consensus active
site sequences of RIPs (Fig. 1)
(32).
E. coli strains harboring the recombinant
plasmid (pSUMO-Fordin) and non-recombinant plasmid (pSUMO) were
incubated for expression analysis. The recombinant protein with an
expected band of ~45 kDa was detected in the soluble fraction
(Fig. 2A), which consisted of the
SUMO protein with a molecular weight of 20 kDa fused to fordin. The
His-SUMO tag allowed the fused protein to be purified by Ni-NTA
affinity chromatography and be confirmed by western blot analysis
using the rabbit polyclonal anti-His antibody (Fig. 2B). The His-SUMO tag was cleaved
with protease ULP, and fordin was measured. The purity of the fused
protein and fordin was identified via the presence of a single band
at ~45 and 28 kDa on the gel, respectively (Fig. 2A). The yields of the fused protein
and fordin were ~25 and 13 mg of protein/l of bacterial culture,
respectively. The expressed protein was characterized by mass
spectrometry (MS) analysis, and the theoretical molecular mass
obtained by MS methods was 28.3 kDa. The sequence from MS analysis
had 54% sequence identity with the putative polypeptide based on
the vf-RIP gene, with 21 unique matching peptides (partially
in Table II).
 | Figure 2Expression analysis and N-glycosidase
activity analysis of fordin. (A) Sodium dodecyl
sulfate-polyacrylamide gel electrophoresis analysis of fordin
expressed in E. coli. Lane M, protein molecular weight
marker (kDa); lane 1, supernatant of the cell lysate of Rosetta
transformed with the empty vector pSUMO; lane 2, supernatant of the
cell lysate of Rosetta transformed with recombinant vector
pSUMO-Fordin; lane 3, inclusion bodies of Rosetta transformed with
the empty vector pSUMO; lane 4, inclusion bodies of Rosetta
transformed with recombinant vector pSUMO-Fordin; lane 5, purified
fusion protein; lane 6, fusion protein following treatment with
ULP; lane 7 purified fordin. (B) Western blot analysis using
polyclonal anti-His antibodies. Lane 1, purified fusion protein;
lane 2, fusion protein following treatment with ULP; lane 3,
purified fordin. (C) RNA N-glycosidase activity of fordin. Lane 1,
control rRNA without fordin treatment; lane 2, rRNA treated with
0.4 µg fordin. |
| Table IIIdentification of the expressed
protein by mass spectrometry analysis. |
Table II
Identification of the expressed
protein by mass spectrometry analysis.
| Identified
protein | NCBI accession | MW (kDa) | Coverage (%) | Score sequest
HT | Matched
peptides |
|---|
| vf-RIP | MF521485 | 28.3 | 54 | 518.05 |
LQTQLGVLALSDGIYTLNK; LSSGIQTADANGK; SGNSN
GIPVLSATVAEAK; VITNFG QR; ATTYLFKGTK; FKPLP DVTSRENNWGK; KGADRL
QTQLGVLALSDGIYTLNK; FSGSYDDLKKK; GADRLQ TQLGVLALSDGIYTLNK;
GTKQTMLK |
The N-glycosidase activity of fordin was examined
against Sapium sebiferum rRNA. A specific endo-fragment was
generated following fordin treatment due to the N-glycosidase
activity of fordin (Fig. 2C). The
negative control (no fordin treatment), did not yield the same
fragment.
Fordin inhibits viability of cancer
cells
The selected tumor and normal cell lines were
treated with increasing concentrations of Fordin (0.2–10 µM)
for 24, 48 and 72 h. Following fordin exposure, effective cytotoxic
activity was observed as a decline in survival rates in a dose- and
time-dependent manner in the four cancer cell lines (Fig. 3A–D). The ICs of fordin were
calculated based on the viability assay (Table III). Considering the
IC50 values, fordin was more toxic against the four
tumor cell lines, particularly the U-2 OS and HepG2 cells, than the
normal cell line (Fig. 3E).
Specifically, a sharp decline in survival was observed in the U-2
OS and HepG2 cells at a fordin concentration >1 µM at the
24- and 48-h time intervals, as shown in Fig. 3A and B.
| Table IIIICs of fordin in the selected cell
lines. |
Table III
ICs of fordin in the selected cell
lines.
| Cell line | IC20
(µM)
| IC50
(µM)
| IC70
(µM)
|
|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h |
|---|
| U-2 OS | 3.66 | 2.22 | 0.18 | 6.85 | 5.43 | 2.22 | 9.25 | 6.75 | 6.12 |
| Hep G2 | 4.19 | 2.38 | 1.48 | 7.48 | 6.28 | 5.88 | 9.94 | 8.72 | 7.58 |
| HeLa | 7.64 | 5.05 | | 12.97 | 10.25 | | 14.12 | 13.22 | |
| A549 | 8.61 | 6.38 | | 15.55 | 10.50 | | 15.26 | 13.21 | |
| MRC-5 | >200 | 28.91 | | >400 | 34.64 | | >400 | 38.02 | |
Fordin exposure induces apoptosis
To confirm fordin induces apoptosis, the nuclear
morphology of U-2 OS, Hep G2 and MRC-5 cells was determined by
Hoechst 33342 staining using fluorescence microscopy. Exposure of
the cells to fordin (IC20, IC50, or
IC70) for 24 h resulted in clear destructive changes
(chromatin condensation and nuclear fragmentation) in the cancer
cell nuclei. At the fordin IC20, nuclear shrinkage and
chromatin condensation were noted, whereas at IC50 or
IC70, nuclear fragmentation was also observed in the
cells (Fig. 4A). The apoptotic
effects in response to fordin exposure were more profound in U-2 OS
and HepG2 cells compared with MRC-5 cells (Fig. 4A and B).
To further verify fordin as an inducer of apoptosis,
an Annexin V-based assay was performed. The cells treated with
fordin (IC20, IC50, or IC70) for
24 h were examined by FACS analysis. Significant increases, from
2.03 to 35.65% and 1.30 to 13.95%, occurred in Annexin
V-FITC-positive cells with increasing concentrations of fordin in
U-2 OS and HepG2 cells, respectively (Fig. 4C). However, this increase was lower
(from 0.84 to 2.90%) in the MRC-5 cells.
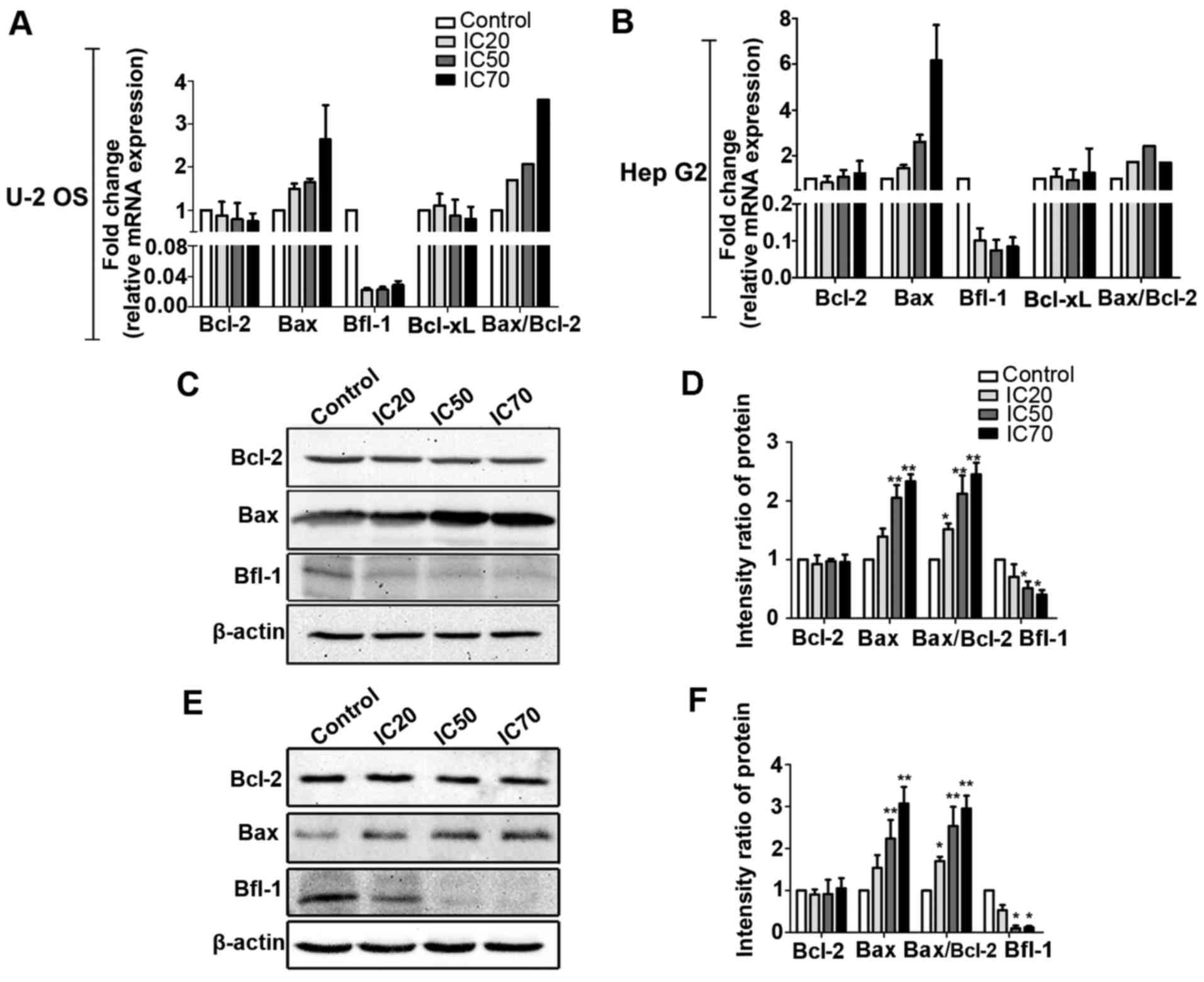
Following fordin treatment for 24 h,
apoptosis-related gene expression of the U-2 OS and HepG2 cells was
examined by RT-qPCR and western blot analyses. The results showed
the potential of fordin to influence the expression of Bcl-2 family
genes (Fig. 5). Fordin exposure
significantly increased the mRNA level of Bax (maximum, 2.6-fold),
and inhibited the mRNA level of Bfl-1 (maximum, 46.0-fold) in U-2
OS cells. The mRNA expression levels of Bcl-2 and Bcl-xL in the
treated cells were essentially unchanged compared with those in the
untreated cells. Similar effects were observed in HepG2 cells, in
which maximum induction was 2.7-fold for the Bax gene and maximum
inhibition was 47.9-fold for the Bfl-1 gene. These results are
consistent with the western blot data. The expression of Bax was
increased in a concentration-dependent manner in the U-2 OS
(Fig. 5C and D) and HepG2 cells
(Fig. 5E and F), with maximum
increases of 2.3- and 3.1-fold, respectively (P<0.001). The
expression of Bfl-1 was decreased in the U-2 OS (maximum, 1.5-fold,
P<0.05) and HepG2 (maximum, 7.6-fold, P<0.05) cells. The
expression of Bcl-2 did not differ between the treated cells and
untreated control cells.
Fordin treatment interferes with cell
invasion and migration
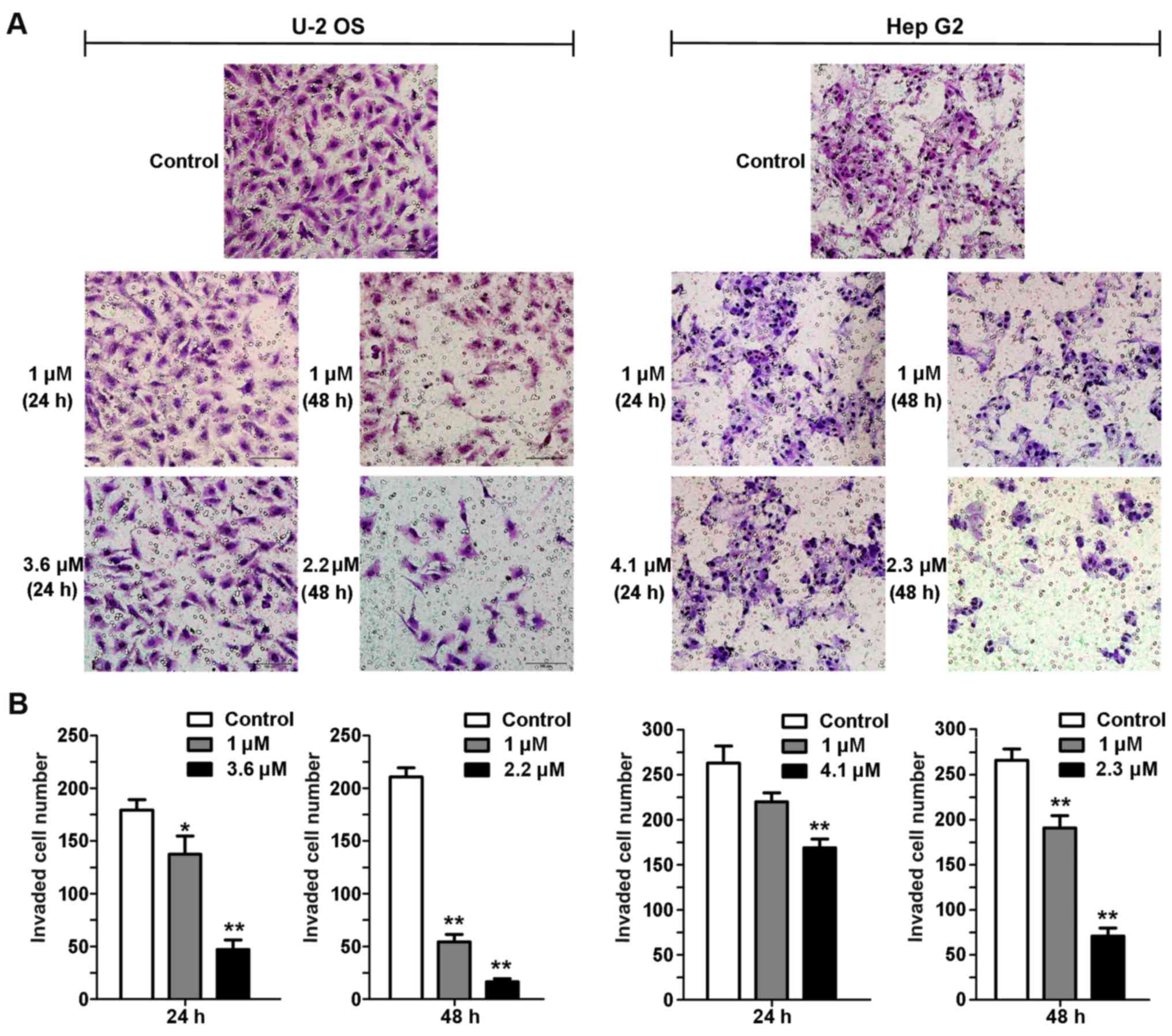
The Transwell assay was used to evaluate the effects
of fordin on the invasive activity of U-2 OS and HepG2 cells. The
invasive behavior of the cells was examined following 24 and 48 h
treatment with fordin (1 µM and IC20). The two
cell lines exhibited a reduction in the number of trans-membrane
cells (Fig. 6). The number of U-2
OS cells was reduced by 23% (P<0.05) and 74% (P<0.001)
following 24 h treatment with fordin (1 µM and
IC20), respectively. The reduction was more pronounced
for the cells treated with fordin (1 µM and IC20)
for 48 h (74 and 92%, respectively; P<0.001). The number of
HepG2 cells was decreased only by 16% (P=0.0548) and 35%
(P<0.001) following 24 and 48 h treatment with fordin (1
µM), respectively. Following 24 and 48 h treatment with
fordin (IC20), the reduction in invasive cell number
increased to 28% (P<0.001) and 73% (P<0.001),
respectively.
A scratch assay was used to monitor the ability of
U-2 OS and HepG2 cells to cover the vacant scuffed area introduced
by scratching for 24 h. As shown in Fig. 7, the ability of cells treated with
fordin to cover the scratched area was reduced. No apparent
morphologic signs of treatment-related toxicity were shown at a low
concentration of fordin (1 µM and IC20). U-2 OS
and the HepG2 control cells covered 1,427±201 and 629±36 µm
distances between the scratched edges following a 24-h incubation
period, respectively. Fordin (1 µM) exposure inhibited the
migration potential by 20% (1,143±77 µm) and 51% (310±51
µm) in the U-2 OS and HepG2 cells, respectively. The effects
were more marked at a higher concentration of fordin
(IC20), with a 49% (730±79 µm) and 77% (143±43
µm) decrease of the migration potential of U-2 OS and HepG2
cells, respectively.
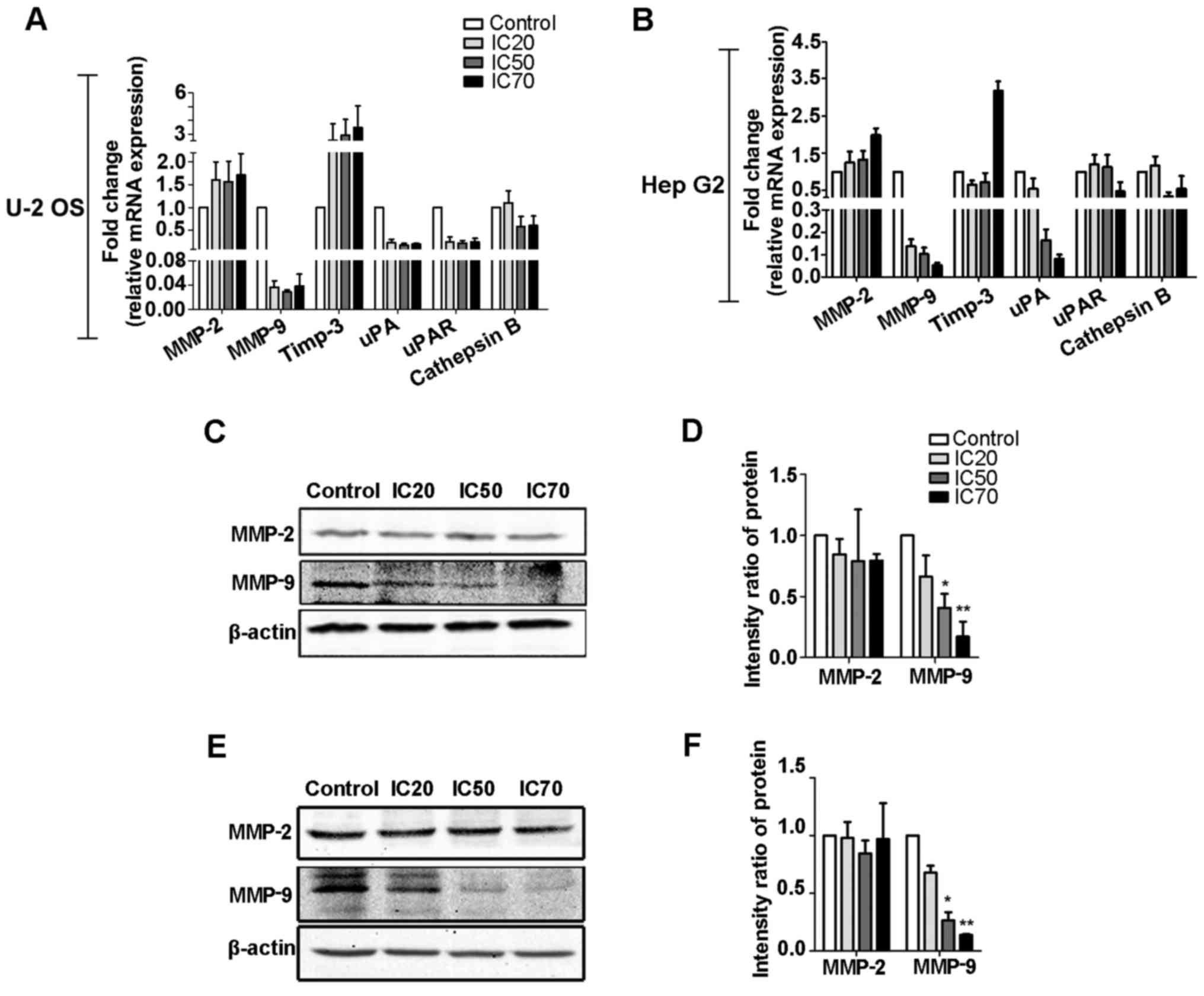
RT-qPCR and western blot analysis were used to
examine the effects of fordin on several invasion-relevant genes
(Fig. 8). Following fordin
exposure of the U-2 OS cells (Fig. 8A,
C and D), the maximum inhibition of the mRNA and protein levels
of the MMP-9 gene with migratory potential was 35- and 4.8-fold
(P<0.001), respectively. A maximum induction of 3.5-fold for the
mRNA level of tissue inhibitor of metalloproteinase (Timp)-3, an
inhibitor of the MMP family, was also observed. In the HepG2 cells
(Fig. 8B, E and F), the mRNA and
protein levels of MMP-9 were decreased in a dose-dependent manner
with a maximum reduction of 19- and 6.2-fold (P<0.001),
respectively. The mRNA level of Timp-3 was not affected by fordin
treatment. The mRNA and protein levels of MMP-2 gene did not differ
significantly between control and treated groups in either cell
line. In addition, in the two cell lines, fordin had inhibitory
effects on the mRNA levels of cathepsin B (maximum, 1.0- and
2.0-fold), urokinase plasminogen activator (uPA; maximum, 5.1- and
11.7-fold), and uPA receptor (maximum, 3.8- and 1.6-fold) (Fig. 8A and B), which have significant
roles in tumor invasion. Downregulation of the aforementioned
metastasis-related genes may be important in the suppression of U-2
OS and HepG2 cell invasion and migration caused by fordin.
Effects of fordin on the regulation of
NF-κB signaling pathways
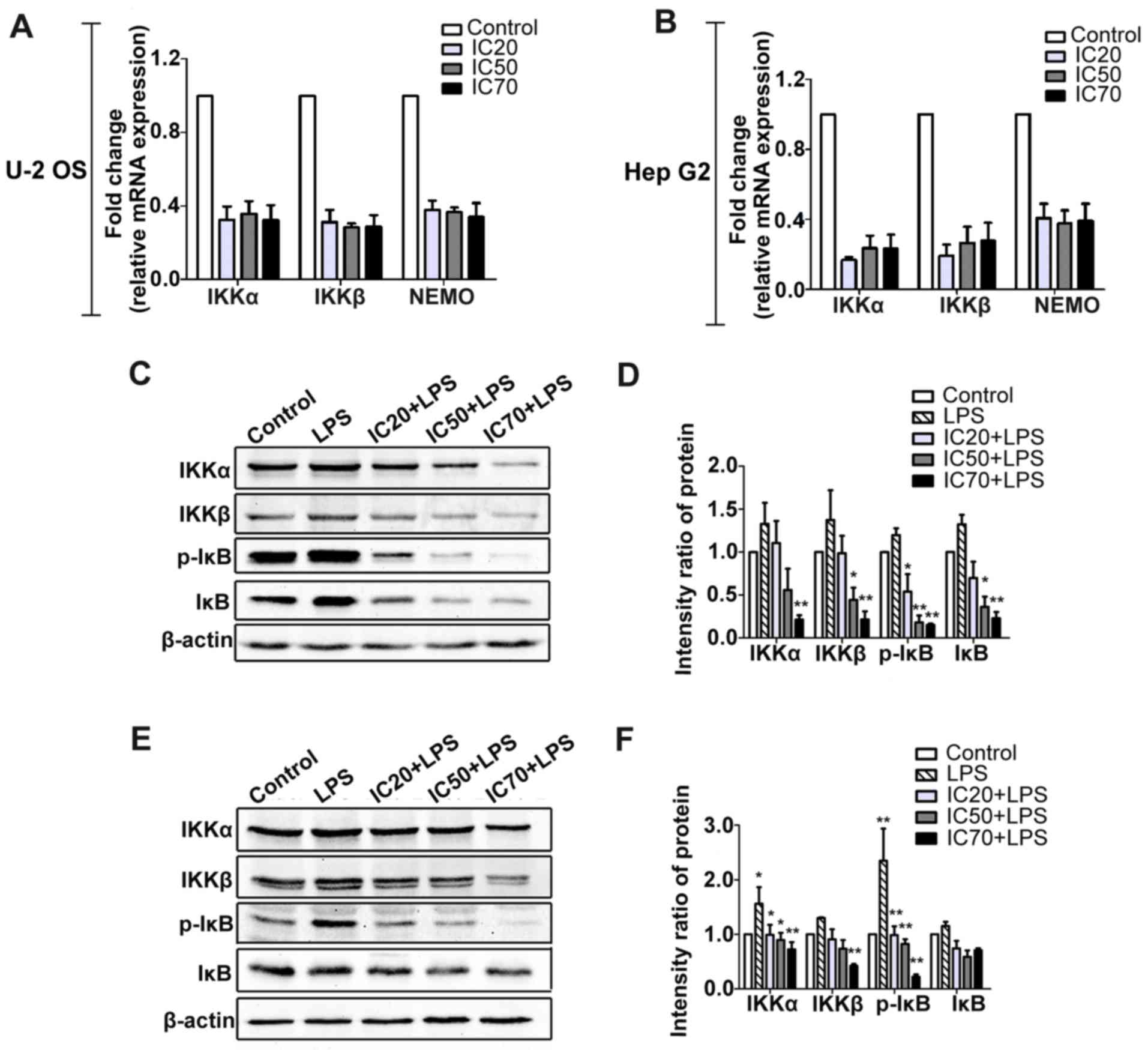
To investigate the effects of fordin on NF-κB
activation, the protein levels of p65 (total and phosphorylated)
were determined in U-2 OS and HepG2 cells. As shown in Fig. 9A and B, the expression of
phosphorylated p65 was significantly decreased in the treated
cells; however, the total p65 expression was not changed. To
further verify the effects of fordin on the inhibition of p65
activation, lipopolysac-charide (10 µg/ml) was used to
induce NF-κB activation. The protein levels of intra-nuclear p65
(phosphorylated and total) were markedly suppressed (maximum, 10.3-
and 9.6-fold, respectively; P<0.001) in U-2 OS cells treated
with LPS and fordin (IC20, IC50, or
IC70) compared with the cells treated with LPS alone
(Fig. 9C). Similar findings were
observed in the HepG2 cells, in which the maximum reductions in
intra-nuclear p65 (phosphorylated and total) expression were 8.5-
and 2.6-fold (P<0.001) (Fig.
9D), respectively. In the two cell lines, the protein levels of
total p65 in the cytoplasm showed negligible change; however, the
expression of phosphorylated p65 in the cytoplasm was downregulated
in Hep G2 cells (maximum, 3.2-fold, P<0.05). These results
demonstrated that fordin inhibited the nuclear accumulation and
phosphorylation of NF-κB in cancer cells.
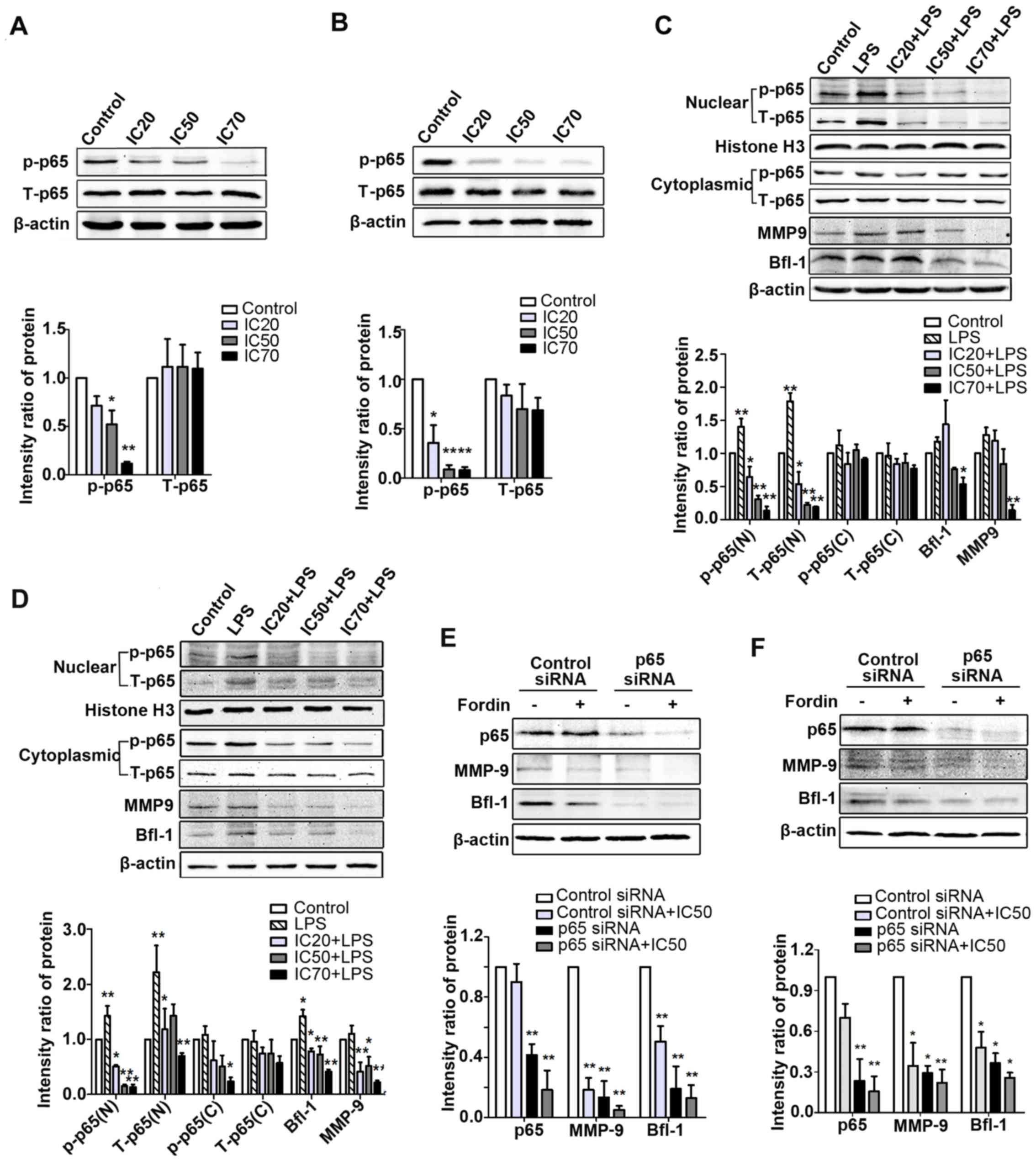 | Figure 9NF-κB/p65 is responsible for the
downregulation of MMP-9 and Bfl-1 mediated by fordin in U-2 OS and
HepG2 cells. The expression of p65 (phosphorylated and total),
MMP-9 and Bfl-1 were determined by western blot analysis in the
various treated cells and normalized to β-actin or Histone H3. (A)
U-2 OS and (B) HepG2 cells were incubated with various
concentrations of fordin for 24 h; *P<0.05 and
**P<0.001 vs. untreated control group. (C) U-2 OS and
(D) HepG2 cells were treated with fordin and/or LPS for 24 h;
*P<0.05 and **P<0.001 vs. control
group. (E) U-2 OS and (F) HepG2 cells transfected with p65 siRNA or
control siRNA were treated with or without fordin
(IC50); *P<0.05 and
**P<0.001 vs. control siRNA group (n=3). siRNA, small
interfering RNA; LPS, lipopolysaccharide; p-, phosphorylated; T,
total; N, nuclear; C, cytoplasmic; MMP, matrix metalloproteinase;
Bfl-1, B-cell lymphoma-related protein A1; IC, inhibitory
concentration. |
In addition, understanding whether NF-κB is involved
in the regulation of the expression of Bfl-1 and MMP-9 mediated by
fordin, LPS and p65 siRNA were used to regulate the activation of
NF-κB in U-2 OS and HepG2 cells. As shown in Fig. 9C and D, upregulation of the
intra-nuclear expression of p65, Bfl-1 and MMP-9 due to LPS was
reduced by fordin treatment in the U-2 OS (maximum, 8.6-, 1.2- and
8.2-fold; P<0.05) and HepG2 (maximum, 2.2-, 2.4- and 4.0-fold,
respectively; P<0.001) cells. In Fig. 9E and F, the knockdown of p65
reduced the protein levels of MMP-9 and Bfl-1 in U-2 OS (6.5- and
4.3-fold, respectively; P<0.001 vs. control siRNA transfected
cells) and HepG2 cells (2.4- and 1.7-fold, respectively; P<0.05
vs. control siRNA transfected cells). The effects of fordin on the
expression of MMP-9 and Bfl-1 were weaker in the p65
siRNA-transfected cells (U-2 OS: 1.7- and 0.5-fold; HepG2: 0.3- and
0.4-fold, respectively; P>0.05) than in control
siRNA-transfected cells (U-2 OS: 4.5- and 1.0-fold; Hep G2: 1.9-
and 1.1-fold, respectively; P<0.05). These findings revealed
that NF-κB regulated the expression of Bfl-1 and MMP-9 in U-2 OS
and HepG2 cells, and fordin downregulated the expression of Bfl-1
and MMP-9, at least in part, associated with the inactivation of
NF-κB.
The finding that fordin inhibited the nuclear
accumulation and phosphorylation of NF-κB in cancer cells was
verified by a corollary study (Fig.
10A–F). Fordin treatment decreased the mRNA levels of IKKα
(maximum, 2.2- and 5.0-fold) and IKKβ (maximum, 2.6- and 4.7-fold)
in the U-2 OS and HepG2 cells, respectively. The protein levels of
IKKα and IKKβ were decreased by a maximum of 4.5- and 4.3-fold
(P<0.001) in U-2 OS cells, respectively. The maximum reduction
in IKKα and IKKβ was 0.9- and 2.1-fold (P<0.001) in the HepG2
cells, respectively. The protein levels of phosphorylated IκB were
also reduced in U-2 OS and HepG2 cells (maximum, 6.6- and
11.6-fold, respectively; P<0.001).
Discussion
V. fordii, belonging to Euphorbiaceae, is a
woody oil plant and prevalent in southern China. The seeds of V.
fordii are well-known for their potential as a raw material to
produce biodiesel (16,33). The roots and leaves of V.
fordii have been widely used in folk medicine. Based on our
previous transcriptome analysis, information on a novel RIP
(fordin) gene from V. fordii was retrieved. Fordin is a
279-amino-acid polypeptide with a molecular weight of 30.781 kDa.
The alignment results with other RIP sequences have demonstrated
that fordin is closely associated with type I RIPs with respect to
the conserved region, which is also the ribosome-inactivating
region. In the present study, the open reading frame of the fordin
gene was successfully obtained from V. fordii leaves. In
consideration of the difficulty in purifying this protein from the
plant, a novel method was established to obtain soluble and active
fordin from the E. coli expression system. This technique
allows mass quantities of fordin to be obtained for further
biological activity investigations and application. The results of
the enzyme activity assay showed that fordin had N-glycosidase
activity which may lead to ribosome inactivation, as with the
majority of the reported RIPs from different plant species
(32).
To determine the potential of fordin in cancer
therapy, the present study examined its specific action on the
viability of tumor and normal cells. Considering the
IC50 values in the viability assay, fordin exhibited
more toxicity towards four tumor cell lines than the normal cell
line. This finding may be due to fordin being a type I RIP without
a B-chain to bind the cell surface glycoconjugate to increase the
internalization of activity A-chain into normal cells. Compared
with fordin, ricin exhibits marked toxicity towards normal cells,
as the LD50 of mice can be as low as 1–10 µg/kg (34). The inhibitory effect of fordin on
four tumor cell lines was exerted in a time- and dose-dependent
manner. The U-2 OS and HepG2 cell lines were more susceptible to
the fordin cytotoxicity. It is reported that type I RIPs show
diverse toxic effects against different tumor cells due to the
differential expression of receptors for binding, the presence of
sialyl caps, and specific molecular routes developed by cancer
cells over time (35,36). The mechanism of the internalization
of fordin requires further investigation. Furthermore, in U-2 OS
and HepG2 cells, a sharp decline in survival rates was observed
following fordin (>1 µM) exposure. However, the effects
were not in proportion to further increases in fordin
concentrations (>7.5 µM). This observation indicates the
presence of possible feedback mechanisms and successful resistance
to high fordin concentrations in cancer cells.
Fordin also led to the inhibition of invasion and
migration of U-2 OS and HepG2 cells, as observed by scratch and
Transwell assays. The migratory and invasive abilities of cancer
cells from the primary site to distant organs are essential for
metastasis. At the molecular level, fordin exposure suppressed the
mRNA levels of uPA, uPAR, and cathepsin B, which are considered to
be crucial in the invasion of tumor cells by degradation of the
extracellular matrix (ECM) (37).
Reports have also claimed that cathepsin-dependent and uPA-mediated
tumor invasion are achieved by inducing the expression of MMP-2 and
MMP-9 (22). Overexpression of the
MMP family genes has been reported to be correlated with cancer
metastasis (38–41). Therefore, the present study
evaluated the effects of fordin on the expression of MMP-2 and
MMP-9. The results revealed that fordin exposure downregulated the
mRNA and protein levels of MMP-9, which demonstrated the importance
in tumorigenesis and metastasis within the MMP family (42). Therefore, the observed
anti-metastatic effects of fordin in cancer cells are associated
with the downregulation of metastasis-dependent gene (MMP-9)
expression.
To clarify the reasoning behind the fordin-mediated
anti-proliferative and cytotoxic effects, the present study
investigated the possible effects on the induction of apoptosis in
cancer (U-2 OS and HepG2) cells and normal (MRC-5) cells. Fordin
induced more considerable signs of apoptosis in the two cancer
cells than in the normal cells. These apoptotic effects were
further confirmed by nuclear staining, which showed chromatin
condensation and nuclear fragmentation, and the Annexin V-FITC
assay with an increased percentage of Annexin V-FITC-positive
cells. Searching for the molecular logic of these apoptotic
effects, it was found that fordin exposure led to the induction of
pro-apoptotic (Bax) genes, as shown by RT-qPCR and western blot
analyses. Several studies have indicated the suppression of
anti-apoptotic (Bcl-2 and Bcl-xL) gene expression following RIP
treatment (18,21,43).
However, in the present study, fordin had no significant effect on
the levels of Bcl-2 and Bcl-xL. The expression ratio of Bax/Bcl-2
was increased in a concentration-dependent manner. The mRNA and
protein levels of Bfl-1 were also markedly downregulated following
fordin exposure. Bfl-1, known as A1, is a Bcl-2 homolog and a
direct target of NF-κB (44,45).
It is required to prevent apoptosis in various cells (46). However, there are few reports on
its role in the apoptosis induced by RIPs. In the present study,
the alterations in the expression of Bfl-1 and Bax were crucial in
the apoptosis induced by fordin in U-2 OS and HepG2 cells.
Following fordin treatment, suppression of the
nuclear accumulation of NF-κB was observed, which resulted from a
decrease in the phosphorylation level of IκB caused by quelling the
expression of IKKα and IKKβ. Suppression of the nuclear
accumulation and phosphorylation of NF-κB by fordin treatment
effectively inhibited the activation of NF-κB in the cells. A
number of studies have shown the pivotal position of NF-κB in tumor
initiation, progression, and invasion in various types of human
cancer (47–50). Several RIPs, including TCS, Viscum
album agglutinin and Abrus agglutinin, have been shown to
induce apoptosis through quelling activation of the NF-κB signaling
pathway (51–54). In consideration of these findings,
the ability of fordin to suppress NF-κB in cancer cells is of
paramount importance. Furthermore, it is widely reported that the
overexpression of MMP and Bcl-2 family are closely associated with
the activation of NF-κB (22,55).
The present focused on the effects of fordin on the expression of
MMP-9 and Bfl-1 in cells treated with LPS, which is known to
activate the NF-κB pathway. The results revealed that the
expression of intranuclear p65 (phosphorylated and total), Bfl-1
and MMP-9 were increased in the LPS-treated cells, but decreased
following fordin treatment. In addition, the protein levels of
MMP-9 and Bfl-1 were decreased in the cells with siRNA-induced p65
knockdown. The effects of fordin on the expression of MMP-9 and
Bfl-1 were partly inhibited in the p65 siRNA-transfected cells.
These findings suggested that NF-κB is responsible for the
downregulated expression of MMP-9 and Bfl-1 mediated by fordin.
This suggests that an NF-κB-mediated pathway is involved in the
inhibition of invasion and induction of apoptosis by fordin
exposure in U-2 OS and HepG2 cells (Fig. 11).
In conclusion, the present study revealed for the
first time, to the best of our knowledge, the presence of fordin
and the antitumor activity of fordin in vitro. The results
demonstrated that fordin induced multi-faceted antineoplastic
effects in U-2 OS and HepG2 cells, including antiproliferation and
anti-invasion effects, and the induction of apoptosis. It appears
that NF-κB is one of the main driving factors for fordin-mediated
responses. The results suggested that fordin inhibited the invasion
of cancer cells, possibly via suppression of the NF-κB-dependent
activation of MMPs, and induced apoptosis partly through
NF-κB-mediated pro-apoptotic mechanisms. Taken together, the
diverse anticancer effects of fordin suggest its therapeutic
potential on restricting tumor growth and reducing the risks of
cancer metastasis as a naturally therapeutic agent. However, fordin
also may possess adverse effects, including severe systemic
anaphylaxis, immunogenicity and toxicity, as with other reported
RIPs. Further investigations of fordin, in terms of its antitumor
activity in vivo, the precise mechanisms underlying its
effects, and targeted toxins, are warranted.
Acknowledgments
The authors wish to acknowledge Professor Fanfor
for his provision of the pET28b-SUMO and p28ULP expression
vector.
References
|
1
|
Girbés T, Ferreras JM, Arias FJ and Stirpe
F: Description, distribution, activity and phylogenetic
relationship of ribosome-inactivating proteins in plants, fungi and
bacteria. Mini Rev Med Chem. 4:461–476. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stirpe F: Ribosome-inactivating proteins.
Toxicon. 44:371–383. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stirpe F and Battelli MG:
Ribosome-inactivating proteins: Progress and problems. Cell Mol
Life Sci. 63:1850–1866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li J, Xia X, Ke Y, Nie H, Smith MA and Zhu
X: Trichosanthin induced apoptosis in HL-60 cells via mitochondrial
and endoplasmic reticulum stress signaling pathways. Biochim
Biophys Acta. 1770:1169–1180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Polito L, Bortolotti M, Mercatelli D,
Battelli MG and Bolognesi A: Saporin-S6: A useful tool in cancer
therapy. Toxins (Basel). 5:1698–1722. 2013. View Article : Google Scholar
|
|
6
|
Mohamed MS, Veeranarayanan S, Poulose AC,
Nagaoka Y, Minegishi H, Yoshida Y, Maekawa T and Kumar DS: Type 1
ribotoxin-curcin conjugated biogenic gold nanoparticles for a
multimodal therapeutic approach towards brain cancer. Biochim
Biophys Acta. 1840:1657–1669. 2014. View Article : Google Scholar
|
|
7
|
Smith ME and Hayoun MA: Toxicity, Ricin.
StatPearls (Internet); Jun 10–2017, Epub ahead of print.
|
|
8
|
Puri M, Kaur I, Perugini MA and Gupta RC:
Ribosome-inactivating proteins: Current status and biomedical
applications. Drug Discov Today. 17:774–783. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Puri M: 'Momordica balsamina: a medicinal
and neutraceutical plant for health care management'. Comments:
biotechnological potential of M. balsamina revealed. Curr Pharm
Biotechnol. 11:229. 2010. View Article : Google Scholar
|
|
10
|
Polito L, Bortolotti M, Pedrazzi M,
Mercatelli D, Battelli MG and Bolognesi A: Apoptosis and
necroptosis induced by stenodactylin in neuroblastoma cells can be
completely prevented through caspase inhibition plus catalase or
necrostatin-1. Phytomedicine. 23:32–41. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Panda PK, Behera B, Meher BR, Das DN,
Mukhopadhyay S, Sinha N, Naik PP, Roy B, Das J, Paul S, et al:
Abrus Agglutinin, a type II ribosome inactivating protein inhibits
Akt/PH domain to induce endoplasmic reticulum stress mediated
autophagy-dependent cell death. Mol Carcinog. 56:389–401. 2017.
View Article : Google Scholar
|
|
12
|
College JN: Dictionary of Chinese Materia
Medica. Journal. 1977.
|
|
13
|
Piccirilli A, Piccardi N and Miska P: Use
of at least one conjugated triene-containing fatty acid for
preparing a medicine for treating inflammation. US Patent
2008/0045594 A1. Filed July 26, 2004; issued February 2, 2008.
|
|
14
|
Igarashi M and Miyazawa T: Newly
recognized cytotoxic effect of conjugated trienoic fatty acids on
cultured human tumor cells. Cancer Lett. 148:173–179. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Belury MA: Dietary conjugated linoleic
acid in health: Physiological effects and mechanisms of action.
Annu Rev Nutr. 22:505–531. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mao Y, Liu W, Chen X, Xu Y, Lu W, Hou J,
Ni J, Wang Y and Wu L: Flower development and sex determination
between male and female flowers in Vernicia fordii. Front Plant
Sci. 8:12912017. View Article : Google Scholar :
|
|
17
|
Polito L, Bortolotti M, Maiello S,
Battelli MG and Bolognesi A: Plants producing ribosome-inactivating
proteins in traditional medicine. Molecules. 21:212016. View Article : Google Scholar
|
|
18
|
Fang EF, Zhang CZ, Wong JH, Shen JY, Li CH
and Ng TB: The MAP30 protein from bitter gourd (Momordica
charantia) seeds promotes apoptosis in liver cancer cells in vitro
and in vivo. Cancer Lett. 324:66–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pan WL, Wong JH, Fang EF, Chan YS, Ng TB
and Cheung RC: Preferential cytotoxicity of the type I ribosome
inactivating protein alpha-momorcharin on human nasopharyngeal
carcinoma cells under normoxia and hypoxia. Biochem Pharmacol.
89:329–339. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roy S, Axup JY, Forsyth JS, Goswami RK,
Hutchins BM, Bajuri KM, Kazane SA, Smider VV, Felding BH and Sinha
SC: SMI-Ribosome inactivating protein conjugates selectively
inhibit tumor cell growth. Chem Commun (Camb). 53:4234–4237. 2017.
View Article : Google Scholar
|
|
21
|
Wang P, Xu J and Zhang C: CREB, a possible
upstream regulator of Bcl-2 in trichosanthin-induced HeLa cell
apoptosis. Mol Biol Rep. 37:1891–1896. 2010. View Article : Google Scholar
|
|
22
|
Baud V and Karin M: Is NF-kappaB a good
target for cancer therapy? Hopes and pitfalls. Nat Rev Drug Discov.
8:33–40. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sweeney CJ, Mehrotra S, Sadaria MR, Kumar
S, Shortle NH, Roman Y, Sheridan C, Campbell RA, Murry DJ, Badve S,
et al: The sesquiterpene lactone parthenolide in combination with
docetaxel reduces metastasis and improves survival in a xenograft
model of breast cancer. Mol Cancer Ther. 4:1004–1012. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lwin T, Hazlehurst LA, Li Z, Dessureault
S, Sotomayor E, Moscinski LC, Dalton WS and Tao J: Bone marrow
stromal cells prevent apoptosis of lymphoma cells by upregulation
of anti-apoptotic proteins associated with activation of NF-kappaB
(RelB/p52) in non-Hodgkin's lymphoma cells. Leukemia. 21:1521–1531.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao JP, Niu HY, Wang HJ, Huang XG and Gao
DS: NF-κB p65/p52 plays a role in GDNF up-regulating Bcl-2 and
Bcl-w expression in 6-OHDA-induced apoptosis of MN9D cell. Int J
Neurosci. 123:705–710. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hecht M, von Metzler I, Sack K, Kaiser M
and Sezer O: Interactions of myeloma cells with osteoclasts promote
tumour expansion and bone degradation through activation of a
complex signalling network and upregulation of cathepsin K, matrix
metalloproteinases (MMPs) and urokinase plasminogen activator
(uPA). Exp Cell Res. 314:1082–1093. 2008. View Article : Google Scholar
|
|
27
|
Feng AH, Zhang GM, Qi ZG and Fan J:
Functional determination of recombinant maize f and m type
rhioredoxin and entrapment of target proteins. Zhongguo Sheng Wu
Gong Cheng Za Zhi. 30:63–68. 2010.In Chinese.
|
|
28
|
Malakhov MP, Mattern MR, Malakhova OA,
Drinker M, Weeks SD and Butt TR: SUMO fusions and SUMO-specific
protease for efficient expression and purification of proteins. J
Struct Funct Genomics. 5:75–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu Y, Mao Y, Jin S, Hou J, Du H, Yang M
and Wu L: Identification, characterization and structure analysis
of a type I ribosome-inactivating protein from Sapium sebiferum
(Euphorbiaceae). Biochem Biophys Res Commun. 463:557–562. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
32
|
Severino V, Paiardini A, Pascarella S,
Parente A and Chambery A: Structural analysis of toxic volkensin, a
type 2 ribosome inactivating protein from Adenia volkensii Harm
(kilyambiti plant): Molecular modeling and surface analysis by
computational methods and limited proteolysis. Int J Biol Macromol.
45:407–413. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chang CH, Lin HC, Chang CH, Chang CC and
Hsu SH: Potential of domestically produced and imported tung
(Vernicia fordii) seeds for biofuels. J Biobased Mater Bioenergy.
7:512–515. 2013. View Article : Google Scholar
|
|
34
|
Schep LJ, Temple WA, Butt GA and Beasley
MD: Ricin as a weapon of mass terror - separating fact from
fiction. Environ Int. 35:1267–1271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bayer H, Essig K, Stanzel S, Frank M,
Gildersleeve JC, Berger MR and Voss C: Evaluation of riproximin
binding properties reveals a novel mechanism for cellular
targeting. J Biol Chem. 287:35873–35886. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pervaiz A, Adwan H and Berger MR:
Riproximin: A type II ribosome inactivating protein with
anti-neoplastic potential induces IL24/MDA-7 and GADD genes in
colorectal cancer cell lines. Int J Oncol. 47:981–990. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo M, Mathieu PA, Linebaugh B, Sloane BF
and Reiners JJ Jr: Phorbol ester activation of a proteolytic
cascade capable of activating latent transforming growth
factor-betaL a process initiated by the exocytosis of cathepsin B.
J Biol Chem. 277:14829–14837. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Egeblad M and Werb Z: New functions for
the matrix metal-loproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ranuncolo SM, Armanasco E, Cresta C, Bal
De Kier Joffe E and Puricelli L: Plasma MMP-9 (92 kDa-MMP) activity
is useful in the follow-up and in the assessment of prognosis in
breast cancer patients. Int J Cancer. 106:745–751. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Talvensaari-Mattila A, Pääkkö P and
Turpeenniemi-Hujanen T: Matrix metalloproteinase-2 (MMP-2) is
associated with survival in breast carcinoma. Br J Cancer.
89:1270–1275. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Grünwald B, Vandooren J, Gerg M, Ahomaa K,
Hunger A, Berchtold S, Akbareian S, Schaten S, Knolle P, Edwards
DR, et al: Systemic ablation of MMP-9 triggers invasive growth and
metastasis of pancreatic cancer via deregulation of IL6 expression
in the bone marrow. Mol Cancer Res. 14:1147–1158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Roy R, Yang J and Moses MA: Matrix
metalloproteinases as novel biomarkers and potential therapeutic
targets in human cancer. J Clin Oncol. 27:5287–5297. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hu R, Zhai Q, Liu W and Liu X: An insight
into the mechanism of cytotoxicity of ricin to hepatoma cell: Roles
of Bcl-2 family proteins, caspases, Ca(2+)-dependent proteases and
protein kinase. C J Cell Biochem. 81:583–593. 2001. View Article : Google Scholar
|
|
44
|
Sandur SK, Ichikawa H, Sethi G, Ahn KS and
Aggarwal BB: Plumbagin (5-hydroxy-2-methyl-1,4-naphthoquinone)
suppresses NF-kappaB activation and NF-kappaB-regulated gene
products through modulation of p65 and IkappaBalpha kinase
activation, leading to potentiation of apoptosis induced by
cytokine and chemotherapeutic agents. J Biol Chem. 281:17023–17033.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tarte K, Jourdan M, Veyrune JL, Berberich
I, Fiol G, Redal N, Shaughnessy J Jr and Klein B: The Bcl-2 family
member Bfl-1/A1 is strongly repressed in normal and malignant
plasma cells but is a potent anti-apoptotic factor for myeloma
cells. Br J Haematol. 125:373–382. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Grumont RJ, Rourke IJ and Gerondakis S:
Rel-dependent induction of A1 transcription is required to protect
B cells from antigen receptor ligation-induced apoptosis. Genes
Dev. 13:400–411. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mödder UI, Oursler MJ, Khosla S and Monroe
DG: Wnt10b activates the Wnt, notch, and NFκB pathways in U2OS
osteo-sarcoma cells. J Cell Biochem. 112:1392–1402. 2011.
View Article : Google Scholar
|
|
48
|
Westhoff MA, Zhou S, Nonnenmacher L,
Karpel-Massler G, Jennewein C, Schneider M, Halatsch ME, Carragher
NO, Baumann B, Krause A, et al: Inhibition of NF-κB signaling
ablates the invasive phenotype of glioblastoma. Mol Cancer Res.
11:1611–1623. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Iyer SV, Ranjan A, Elias HK, Parrales A,
Sasaki H, Roy BC, Umar S, Tawfik OW and Iwakuma T: Genome-wide RNAi
screening identifies TMIGD3 isoform1 as a suppressor of NF-κB and
osteosarcoma progression. Nat Commun. 7:135612016. View Article : Google Scholar
|
|
50
|
Jiang C, Fang X, Zhang H, Wang X, Li M,
Jiang W, Tian F, Zhu L and Bian Z: AMD3100 combined with triptolide
inhibit proliferation, invasion and metastasis and induce apoptosis
of human U2OS osteosarcoma cells. Biomed Pharmacother. 86:677–685.
2017. View Article : Google Scholar
|
|
51
|
Li CT, Lin CH, Kao TY, Wu MF, Yeh CS, Yeh
KT and Ko JL: The mechanisms of action of Tianhua(™) on antitumor
activity in lung cancer cells. Pharm Biol. 48:1302–1309. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li M, Li X and Li JC: Possible mechanisms
of trichosanthin-induced apoptosis of tumor cells. Anat Rec
(Hoboken). 293:986–992. 2010. View Article : Google Scholar
|
|
53
|
Mukhopadhyay S, Panda PK, Das DN, Sinha N,
Behera B, Maiti TK and Bhutia SK: Abrus agglutinin suppresses human
hepatocellular carcinoma in vitro and in vivo by inducing
caspase-mediated cell death. Acta Pharmacol Sin. 35:814–824. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Khil LY, Kim W, Lyu S, Park WB, Yoon JW
and Jun HS: Mechanisms involved in Korean mistletoe lectin-induced
apoptosis of cancer cells. World J Gastroenterol. 13:2811–2818.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|