Introduction
The kidney is an important organ that is responsible
for maintaining homeostasis in the human body. The urine-producing
functional unit of the kidney is the nephron, which consists of a
renal corpuscle and tubules. Kidney cancer, also known as renal
cancer, originates from cells in the kidney. The two most common
types of renal cancer are renal cell carcinoma (RCC) and urothelial
cell carcinoma. RCC is a heterogeneous disease that arises from the
proximal convoluted tubular epithelium, and is the most common type
of adult renal cancer (1, 2). Clear cell (cc) RCC accounts for ~80%
of all RCC cases (3). Surgical
excision is effective for the majority of patients with ccRCC;
however, disease recurrence or distant metastasis occurs in ~30% of
patients (4,5).
The phosphatidylinositol-4,5-bisphosphate 3-kinase
(PI3K)/AKT signaling pathway is involved in numerous diverse
cellular functions, including cell proliferation (6), differentiation (7), angiogenesis (8) and autophagy (9). The PI3K/AKT signaling pathway also
affects epithelial-mesenchymal transition (EMT) via multiple
regulatory mechanisms, which influences tumor aggressiveness
(10). In addition, the PI3K/AKT
signaling pathway has been demonstrated to regulate cell
proliferation and invasion in ccRCC cell lines (11,12).
Fatty acid binding proteins (FABPs) are 14-15 kDa
proteins that are highly abundant in the cytosol of most tissues.
Nine mammalian FABPs have thus far been identified, and they
contain highly conserved structures (13,14).
FABPs display a high affinity for long-chain fatty acids (FAs) and
regulate lipid metabolism in different tissues (15), including the brain (16), intestine and liver (13). FABPs also mediate the biological
properties of tumor cells. For instance, FABP7, an FABP expressed
in the mammalian central nervous system, has been demonstrated to
regulate glioblastoma cell proliferation (17).
Recently, FABP5, which promotes tumor cell growth in
cervical cancer, was identified as a potential biomarker for lymph
node metastasis (18,19). FABP5 gene silencing
inhibited the proliferation and invasion of human SGC-7901 gastric
cancer cells in vitro (20), and FABP5 stimulated hepatocellular
carcinoma progression and metastasis via EMT (21). Considering the pivotal functions of
the PI3K/AKT signaling pathway in tumor cells, particularly ccRCC
cells, we hypothesized that FABP5 may affect ccRCC cell function
via the PI3K/AKT signaling pathway.
In the present study, the function of FABP5 in ccRCC
cell lines was investigated and the results suggest that FABP5 may
present a putative prognostic biomarker for patients with ccRCC and
provide a novel perspective for the role of FABPs in tumor
biology.
Materials and methods
Bioinformatics prediction using the The
Cancer Genome Atlas (TCGA) database
RNA sequencing data from TCGA (https://cancergenome.nih.gov/) was used to assess the
correlation between FABP5 mRNA expression levels and
clinicopathological features of patients with ccRCC. The expression
of FABP5 in all samples was sorted from low to high, and the
median expression was selected as the cutoff value to distinguish
patients with low and high expression. The median number was
75.32635. Overall survival and disease-free survival analysis were
performed according to a previously described method (22). A total of 246 patient samples with
associated clinical parameters were selected for further
analysis.
Cell culture and transfection
Caki-1 (cat. no. GCC-KI0004RT) and 786O (cat. no.
GCC-KI0003RT) ccRCC cell lines were purchased from Shanghai
GeneChem, Co., Ltd. (Shanghai, China). All cells were cultivated in
complete medium consisting of Dulbecco’s modified Eagle’s
medium/F12 (Corning Inc., Corning, NY, USA) and 10% fetal bovine
serum (Clark Bioscience, Richmond, VA, USA). The GV112 RNA
interference (RNAi) system (Shanghai GeneChem Co., Ltd.) was used
to generate lentiviruses expressing short interfering RNA sequences
targeting FABP5 (LV-FABP5-RNAi). This system contains a U6
promoter-driven multiple cloning site (MCS) and a cytomegalovirus
promoter-driven puromycin gene. The target sequence of FABP5 was
5′-TGGGAAGGAAAGCACAATA-3′ (20).
Lentiviral vectors overexpressing FABP5 (LV-FABP5) were purchased
from Shanghai GeneChem Co., Ltd. directly and were generated using
the GV492 system (Shanghai GeneChem Co., Ltd.). Briefly, expression
from an MCS combined with a 3xFLAG tag is driven by the ubiquitin
promoter, and green fluorescent protein (GFP) and puromycin
expression are driven by the cellobiohydrolase promoter. The
negative control lentiviruses, LV-NC-RNAi and LV-NC, were also
purchased from Shanghai GeneChem Co., Ltd. The scrambled sequence
used for the LV-NC-RNAi was as follows: 5′-TTCTCCGAACGTGTCACGT-3′.
An empty lentiviral vector was used to transfect cells in the LV-NC
group.
Prior to transfection, cells were seeded in six-well
plates at a density of 1×105 cells/well in complete
medium and incubated overnight. Lentiviruses (multiplicity of
infection=10) together with 5 µg/ml polybrene, was then
added to the cells. At 24 h following infection, the culture medium
was replaced with fresh medium containing 2 µg/ml puromycin
(Sangon Biotech Co., Ltd., Shanghai, China), followed by positive
selection for 3 days. The cells were then maintained in complete
medium containing 1 µg/ml puromycin. The PI3K/AKT signaling
pathway inhibitor, LY294002, was purchased from MedChem Express
(Monmouth Junction, NJ, USA) and added to the culture medium at a
concentration of 20 µM for 24 h to inactivate the PI3K/AKT
signaling pathway in vitro.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using a UNIQ-10 Spin Column
RNA Purification kit (Sangon Biotech Co., Ltd.). A total of 1
µg total RNA was prepared and first-strand cDNA was
synthesized using the RevertAid First Strand cDNA Synthesis kit
(Fermentas; Thermo Fisher Scientific, Inc., Waltham, MA, USA). cDNA
was subsequently analyzed using a StepOnePlus Real-Time PCR system
(Thermo Fisher Scientific, Inc.) using AceQ qPCR SYBR Green Master
Mix (Vazyme, Piscataway, NJ, USA). The following primers were used:
FABP5, forward, 5′-GCATTGGTTCAGCATCAG-3′, and reverse,
5′-ATCCGAGTACAGGTGACA-3′; β-actin (control reference gene),
forward, 5′-TAGTTGCGTTACACC CTTTCTTG-3′, and reverse,
5′-CACCTTCACCGTTCCAGT TTT-3′. The thermal cycling parameters were
as follows: 95°C for 5 min, followed by 40 cycles at 95°C for 10
sec and 60°C for 30 sec. The expression of FABP5 was
normalized to β-actin and the expression level was calculated using
the 2−∆∆Cq method (23).
Western blotting
Western blotting was performed according to
previously reported methods (24).
Briefly, following culture for 24 h, a Tissue or Cell Total Protein
Extraction kit (Sangon Biotech Co., Ltd.) was used to extract total
protein from cells. Protein concentrations were determined using
the Enhanced BCA Protein assay kit (Beyotime Institute of
Biotechnology, Haimen, China) and 30 µg total protein was
loaded and separated by 10% SDS-PAGE. Proteins were transferred
onto polyvinylidene fluoride (PVDF) membranes following
electrophoresis. All membranes were blocked with 5% non-fat milk
(cat. no. A600669; Sangon Biotech Co., Ltd.) for 1 h at room
temperature and then incubated with primary antibodies at 4°C
overnight. Following washing with 1X Tris-buffered saline and
Tween-20 (TBST) solution (0.1% Tween; Sangon Biotech Co., Ltd.)
three times (for 5 min each time), the membranes were then
incubated with secondary antibodies at room temperature for 4 h.
The PVDF membranes were incubated with the following primary
antibodies: Mouse anti-β-actin (dilution, 1:1,000; cat. no. 3700;
Cell Signaling Technology, Inc., Danvers, MA, USA); rabbit
anti-FABP5 (dilution, 1:1,000; cat. no. 33191; GeneTex, Inc.,
Irvine, CA, USA), mouse anti-FLAG (dilution, 1:1,000; cat. no.
AF519; Beyotime Institute of Biotechnology), rabbit anti-AKT
(dilution, 1:1,000; cat. no. 4685; Cell Signaling Technology,
Inc.), rabbit anti-phosphorylated (p)-AKT (Ser 473; dilution,
1:1,000; cat. no. 4060; Cell Signaling Technology, Inc.) and rabbit
anti-p-AKT (Thr 308; dilution, 1:1,000; cat. no. 13038; Cell
Signaling Technology, Inc.). The secondary antibodies used were as
follows: Horseradish peroxidase (HRP)-conjugated goat anti-mouse
(dilution, 1:3,000; cat. no. D110103; Sangon Biotech Co., Ltd.) and
HRP-conjugated goat anti-rabbit (dilution, 1:3,000; cat. no.
D110058; Sangon Biotech Co., Ltd.). The membranes were washed with
TBST 3 times (for 5 min each time) and detected using enhanced
chemiluminescence reagent (cat. no. E411; Vazyme). The
immunoreactive membranes were scanned using the Chemidoc XRS system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Relative protein
expression levels were calculated using Image Lab software (Bio-Rad
Laboratories, Inc.).
Cell viability assay
Cells were first seeded at a density of
2×103 cells/well in 96-well plates for 24, 48 and 72 h.
A total of 10 µl Cell Counting kit-8 (CCK-8) reagent
(Beyotime Institute of Biotechnology) was then added to each well
and cells were incubated for 1 h. The optical density (OD) at 450
nm was then measured using a Synergy 2 Enzyme Mark instrument
(BioTek Instruments, Inc., Winooski, VT, USA). Cell viability was
expressed as a percentage and was calculated using the following
formula: (ODdrug-treated group / ODcontrol
group) × 100 (25).
5-ethynyl-2′-deoxyuridine (EdU)
assay
Cells were seeded at a density of 2×105
cells/well in 6-well plates and incubated for 24 h. The cells were
then treated with 50 µM EdU reagent (Guangzhou RiboBio Co.,
Ltd., Guangzhou, China) followed by Apollo-567 reaction cocktail
(Guangzhou RiboBio Co., Ltd.) for 30 min. Following three washes
with phosphate-buffered saline, cells were counterstained with
Hoechst (dilution 1:1,000; Guangzhou RiboBio Co., Ltd.) for 10 min
at room temperature for nuclear staining. All cells were observed
using an EVOS FL Imaging System (Thermo Fisher Scientific,
Inc.).
Cell migration assay
A wound-healing assay was performed using a 35 mm
μ-Dish with a culture insert (ibidi GmbH, Planegg, Germany).
Briefly, 2×104 cells/well were seeded onto the inserts
and cultured in 70 µl complete medium. To estimate cell
migration ability, the culture inserts were gently removed to
create a space ~500 µm in diameter. Each well was then
filled with 1 ml fresh medium. Photographs of the wound areas were
taken using the EVOS FL Imaging System (Thermo Fisher Scientific,
Inc.) immediately (0-h time point), and at 6, 12 and 24 h following
generation of the wound. The migration index was subsequently
calculated, which was considered as the distance migrated by the
treated group relative to the distance migrated in the control
group (26).
Cell invasion assay
Transwell chambers with 8-µm pores (EMD
Millipore, Billerica, MA, USA) were used to measure the invasion
ability of cells. Briefly, the upper chambers were pre-coated with
Matrigel (BD Biosciences; Becton, Dickinson and Company, Franklin,
Lakes, NJ, USA) for 1 h, and 2×104 cells in 300
µl serum-free medium were seeded into the upper chamber. A
total of 1 ml complete medium was added into the lower chamber.
Following incubation at 37°C for 24 h, the cells on the upper
surface of the chamber were removed, fixed in 4% paraformaldehyde
for 10 min at room temperature, and stained with 0.1% crystal
violet (Sangon Biotech, Co., Ltd.) for 1 h at room temperature.
Stained cells were counted using the EVOS FL Imaging System (Thermo
Fisher Scientific, Inc.).
Xenograft model and immunohistochemistry
analysis
A total of 24 BALB/c male mice (age, 4-6 weeks) were
purchased from the Animal Experimental Center of Nantong University
(Nantong, China). All mice were treated under pathogen-free
conditions in cages according to protocols approved by Nantong
University, and the present study was approved by the Ethics
Committee of Nantong University. Briefly, 5×106 Caki-1
cells were diluted in 100 µl serum-free medium and
inoculated subcutaneously into the left flank of each mouse. At 60
days following inoculation, all mice were euthanized by
CO2, where the flow rate displace no more than 30% of
the chamber volume/minute according to the American Veterinary
Medical Association guidelines (27). The tumor tissues were then excised,
and the tumor volumes (V) were calculated using the following
formula: V = ½ (length × width2). Tumor tissues
were fixed in 4% paraformaldehyde for 2 h at room temperature, and
subsequently placed in a 20% sucrose solution for 24 h. All tissues
were then frozen at -20°C and cut into 10-µm sections for
immunohistochemistry analysis. Tissue sections were blocked using
5% bovine serum albumin (cat. no. A500023; Sangon Biotech, Co.,
Ltd.) for 2 h at room temperature. Then, tissue sections were
incubated with rabbit anti-Ki67 antibodies (dilution, 1:100; cat.
no. PA5-19462; Thermo Fisher Scientific, Inc.) overnight at 4°C,
followed by incubation with Alexa Fluor 594-labeled anti-rabbit IgG
(dilution, 1:1,000; cat. no. 8889S; Cell Signaling Technology,
Inc.) at room temperature for 2 h. Cells were then stained with
Hoechst solution (dilution 1:1,000; Guangzhou RiboBio Co., Ltd.)
for 10 min at room temperature. Cells were examined using an
Olympus laser confocal microscope (Olympus Corporation, Tokyo,
Japan).
Statistical analysis
Data from at least three independent experiments
were collected and expressed as the mean ± standard error of the
mean. A Student’s t-test or one-way analysis of variance followed
by Student-Newman-Keuls or least significant difference post hoc
tests was used to analyze the results. The SPSS 23.0 (IBM Corp.,
Armonk, NY, USA) software package was employed for statistical
analyses. Survival analysis was performed using the Kaplan-Meier
method and groups were compared using the log-rank test, according
to a previous report (22). A
χ2 test was performed on selected clinical samples to
determine the association between FABP5 expression and
clinicopathological features of patients with ccRCC. Univariate and
multivariate Cox regression analysis was used to evaluate the
prognostic significance of specific features. P<0.05 was
considered to indicate a statistically significant difference.
Results
FABP5 is upregulated and correlates with
poor survival in patients with ccRCC
Using the profiles published in TCGA, FABP5
expression was observed to be upregulated in ccRCC samples when
compared with adjacent normal samples (P<0.001; Fig. 1A). The overall survival
(P<0.001; Fig. 1B) and
disease-free survival curves demonstrated that patients with higher
FABP5 expression exhibited significantly shorter survival
rates when compared with patients exhibiting lower FABP5
expression levels (P<0.01; Fig.
1C). These results indicate that higher FABP5 expression
in patients with ccRCC may be associated with poor survival.
The TNM classification system is a recognized
standard for grading the extent of disease in patients with cancer
(28). ‘T’ represents the size of
the primary tumor, ‘N’ reflects the extent of metastasis to
regional lymph nodes, and ‘M’ refers to the distant metastasis of
tumor cells. In addition, the grade (G) classification is used to
describe the degree of differentiation of cancer cells. As shown in
Table I, further analysis revealed
that FABP5 was expressed at lower levels in early-stage
tumors (Stages I + II) and at higher levels in more advanced stages
(III + IV). In addition, an association between FABP5
expression and T and M classification was observed (P<0.05);
however, FABP5 expression did not correlate with N or G
classification, age or gender (P>0.05; Table I). Furthermore, univariate Cox
regression analysis demonstrated that high FABP5 expression
was an unfavorable prognostic parameter in patients with ccRCC
(P<0.01; Table II).
Multivariate Cox regression analysis revealed that age (P<0.05)
and M stage (P<0.01) were also unfavorable prognostic
indicators.
 | Table IAssociation between FABP5
expression and the clini-copathological characteristics of patients
with clear cell renal cell carcinoma. |
Table I
Association between FABP5
expression and the clini-copathological characteristics of patients
with clear cell renal cell carcinoma.
| Characteristic | FABP5 gene
expression (patients, no.)
| Total | P-value | χ2 |
|---|
| Low | High |
|---|
| Age (years) | | | | 0.369 | 1.052 |
| ≤60 | 59 | 51 | 110 | | |
| >60 | 64 | 72 | 136 | | |
| Sex | | | | 0.794 | 0.153 |
| Female | 47 | 50 | 97 | | |
| Male | 76 | 73 | 149 | | |
| T
classification | | | | 0.002 | 10.498 |
| T1+T2 | 85 | 60 | 145 | | |
| T3+T4 | 38 | 63 | 101 | | |
| N
classification | | | | 0.784 | 0.303 |
| N0 | 117 | 115 | 232 | | |
| N1+N2 | 6 | 8 | 14 | | |
| M
classification | | | | 0.039 | 4.946 |
| M0 | 109 | 96 | 205 | | |
| M1 | 14 | 27 | 41 | | |
| G
classification | | | | 0.072 | 3.707 |
| G1+G2 | 62 | 47 | 109 | | |
| G3+G4 | 61 | 76 | 137 | | |
| Stage
classification | | | | 0.005 | 8.659 |
| Stage I+II | 78 | 55 | 133 | | |
| Stage III+IV | 45 | 68 | 113 | | |
| Survival
status | | | | 0.002 | 10.805 |
| Live | 89 | 64 | 153 | | |
| Dead | 34 | 59 | 93 | | |
| Table IIUnivariate and multivariate Cox
regression analysis of FABP5 expression with the
clinicopathological characteristics of patients with clear cell
renal cell carcinoma. |
Table II
Univariate and multivariate Cox
regression analysis of FABP5 expression with the
clinicopathological characteristics of patients with clear cell
renal cell carcinoma.
|
Characteristics | Univariate Cox
| Multivariate Cox
|
|---|
| P-Value | HR | 95% CI
| P-Value | HR | 95% CI
|
|---|
| Lower | Upper | Lower | Upper |
|---|
| Sex | 0.789 | 1.059 | 0.697 | 1.608 | | | | |
| Age | 0.042 | 1.548 | 1.015 | 2.362 | 0.019 | 1.670 | 1.090 | 2.561 |
| T stage |
<0.001 | 2.947 | 1.945 | 4.466 | 0.489 | 1.349 | 0.577 | 3.151 |
| N stage | 0.001 | 3.114 | 1.609 | 6.029 | 0.136 | 1.695 | 0.847 | 3.394 |
| M stage |
<0.001 | 4.042 | 2.618 | 6.242 | 0.001 | 2.540 | 1.489 | 4.335 |
| G stage |
<0.001 | 2.487 | 1.580 | 3.915 | 0.058 | 1.609 | 0.983 | 2.633 |
| Stage |
<0.001 | 3.329 | 2.159 | 5.132 | 0.534 | 1.348 | 0.526 | 3.458 |
| FABP5 gene
expression | 0.001 | 2.032 | 1.332 | 3.1 | 0.107 | 1.45 | 0.923 | 2.276 |
FABP5 knockdown inhibits ccRCC cell
growth and decreases p-AKT expression
To investigate the function of FABP5 in ccRCC cells,
Caki-1 and 786O cell lines transfected with FABP5-RNAi were
generated and FABP5 expression was first determined. These
cells were observed to be stably transfected with FABP5-RNAi, as
indicated by the significantly lower levels of FABP5 mRNA in
the FABP5-RNAi group compared with their respective negative
controls (P<0.01; Fig. 2A). As
demonstrated in Fig. 2B and C, the
viability of these cell lines was significantly reduced in the
FABP5-RNAi group when compared with the NC-RNAi group at 24, 48 and
72 h following transfection (all P<0.001 vs. NC-RNAi group apart
from 786O cells at 24 h, P<0.01). An EdU assay was then used to
measure the proliferative ability of the FABP5-RNAi cell lines. The
results demonstrated that the proportion of EdU-positive Caki-1 and
786O cells was decreased in the FABP5-RNAi group compared with the
respective control cells (P<0.05; Fig. 2D and E). Considering the pivotal
functions of the PI3K/AKT signaling pathway in tumor cells,
particularly ccRCC cells (11,12),
the level of p-AKT in FABP5-RNAi cells was examined. As shown in
Fig. 3A-D, FABP5 and p-AKT levels
were decreased in Caki-1 and 786O cells [all P<0.001 vs. NC-RNAi
group apart from p-AKT (Ser 473), P<0.01] suggesting that
reduced FABP5 expression may decrease cell proliferation by
inhibiting the PI3K/AKT signaling pathway.
Overexpression of FABP5 promotes ccRCC
cell growth
In order to investigate the role of FABP5 in ccRCC
further, the effect of exogenous FABP5 expression on ccRCC cells
was examined. Transfected Caki-1 and 786O cells expressed GFP
(Fig. 4A) and cells in the
LV-FABP5 group exhibited significantly higher FABP5 mRNA
levels when compared with controls (P<0.01; Fig. 4B). CCK-8 assay analysis indicated
that the proliferative ability of FABP5-overexpressing cells was
significantly increased in Caki-1 and 786O cells (all P<0.001
vs. LV-NC group apart from Caki-1 cells at 24 h, P<0.01;
Fig. 4C and D). Consistent with
these results, the EdU assay also demonstrated an increased
proportion of proliferating Caki-1 and 786O cells in the
FABP5-overexpression group when compared with the negative controls
(P<0.05; Fig. 4E and F).
Considering that FABP5 knockdown inhibited ccRCC cell growth and
decreased p-AKT expression, the authors of the current study
hypothesized that exogenous FABP5 may promote the proliferation of
ccRCC cells via activating the PI3K/AKT signaling pathway.
Inhibition of PI3K/AKT signaling
alleviates the pro-proliferative effects of exogenous FABP5
expression
To investigate the role of FABP5 in regulating the
PI3K/AKT signaling pathway in ccRCC cells further, 20 µM
LY294002 was used to inhibit the PI3K/AKT signaling pathway in
Caki-1 and 786O cells in vitro. As shown in Fig. 5A and B, LY294002 treatment
significantly reduced the viability of FABP5-overexpressing cells,
as demonstrated by the CCK-8 assay results in Caki-1 (all
P<0.001 vs. LV-FABP5 group; LV-NC group vs. LV-NC+LY294002
group, P<0.01; LV-FABP5+LY294002 group vs. LV-NC+LY294002 group,
P<0.05; Fig. 5A) and in 786O
(all P<0.001 apart from LV-NC group vs. LV-FABP5+LY294002 group,
P<0.05; Fig. 5B) cells.
Consistent with these observations, the results of the EdU assay
(Fig. 5C-E) also indicated that
the number of EdU-positive FABP5-overexpressing Caki-1 (all
P<0.01 vs. LV-FABP5 group apart from LV-NC group, P<0.05;
LV-NC vs. LV-NC+LY294002, P<0.05; Fig. 5D) and 786O (all P<0.001 vs.
LV-FABP5 group apart from LV-NC group, P<0.01; LV-NC group vs.
LV-NC+LY294002 group, P<0.05; Fig.
5E) cells were significantly decreased following treatment with
LY294002.
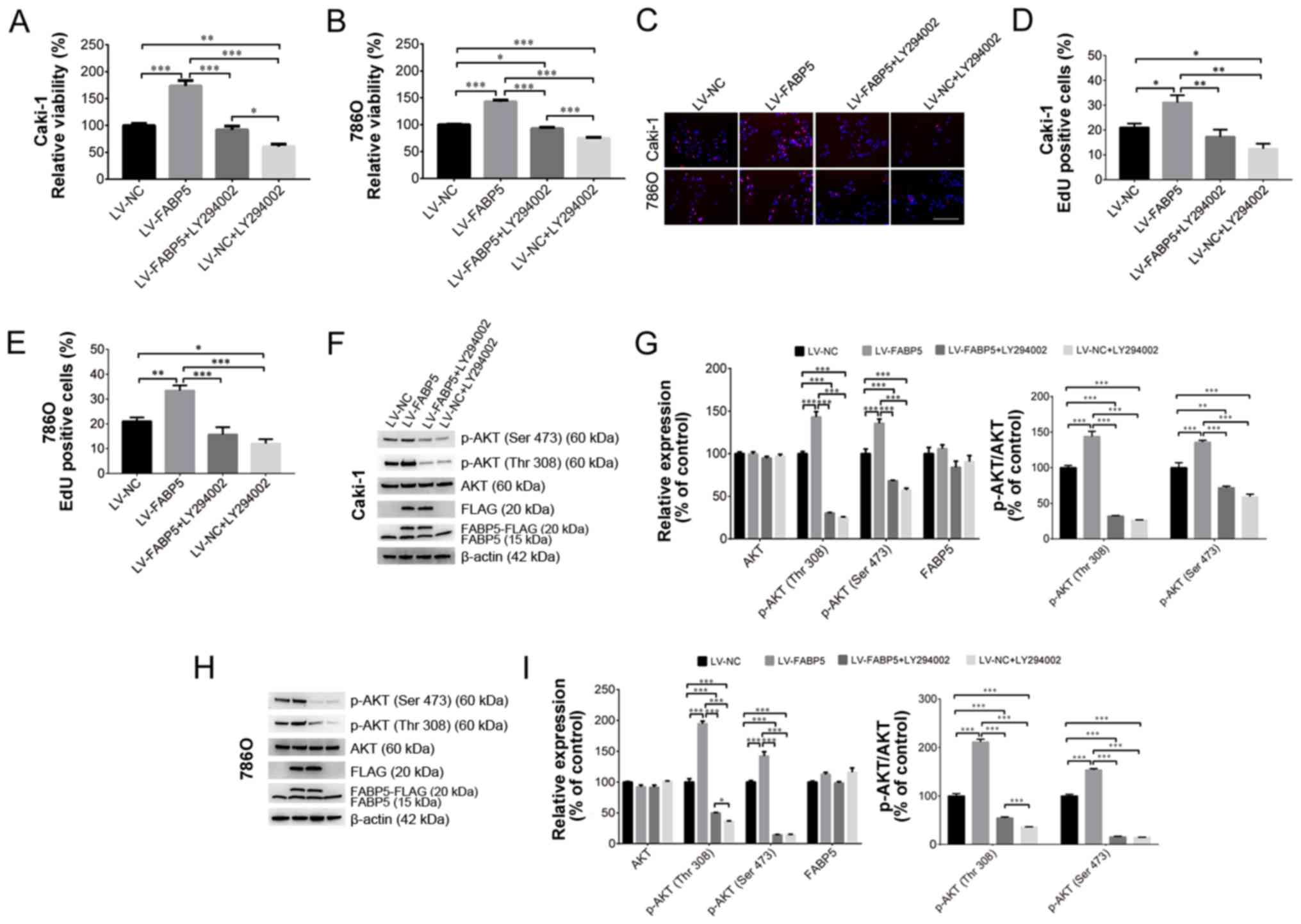 | Figure 5Exogenous FABP5 expression
increased the viability of (A) Caki-1 and (B) 786O cells, whereas
LY294002 treatment decreased the viability of FABP5-overexpressing
cells as determined using the CCK-8 assay. (C) An EdU assay
demonstrated that the proportion of EdU-positive Caki-1 and 786O
cells in the LV-FABP5 group were decreased following LY294002
treatment (scale bar, 200 µm). Quantification of the EdU
staining results in (D) Caki-1 and (E) 786O cells. (F) Western
blotting results demonstrating exogenous FABP5 expression in the
LV-FABP5 group (indicated as FABP5-FLAG and FLAG) and the
upregulation of p-AKT in Caki-1 cells from the LV-FABP5 group.
LY294002 treatment decreased the level of p-AKT in
FABP5-overexpressing Caki-1 cells. (G) Quantification of the
western blotting results in Caki-1 cells. (H) Western blotting
results demonstrating exogenous FABP5 expression in the LV-FABP5
group (indicated as FABP5-FLAG and FLAG) and the upregulation of
p-AKT in 786O cells from the LV-FABP5 group. LY294002 treatment
decreased the level of p-AKT in FABP5-overexpressing 786O cells.
(I) Quantification of the western blotting results in 786O cells.
*P<0.05, **P<0.01 and
***P<0.001, as indicated. FABP5, fatty acid binding
protein 5; CCK-8, Cell Counting kit-8; EdU,
5-ethynyl-2′-deoxyuridine; LV, lentivirus; p-, phosphorylated; NC,
negative control. |
Western blotting analysis verified that exogenous
FABP5 expression (indicated as FABP5-FLAG or FLAG; Fig. 5F and H) could be detected in
FABP5-overexpressing cells. FABP5-FLAG or FLAG expression was
detected in the LV-FABP5 group, indicating the exogenous FABP5 was
successfully expressed in these cells. By contrast, FABP5-FLAG or
FLAG was not detected in the LV-NC group, which confirmed that
there was no exogenous FABP5 expression in the LV-NC group. These
results demonstrated that exogenous FABP5 was successfully
expressed in the LV-FABP5 group of cells. As shown in Fig. 5F-I, the level of p-AKT in Caki-1
and 786O cells from the LV-FABP5 group was significantly increased
when normalized to β-actin and compared with controls. Accordingly,
treatment with LY294002 significantly decreased p-AKT levels in
FABP5-overexpressing Caki-1 (P<0.001; Fig. 5G) and 786O cells (all P<0.001
apart from p-AKT (Thr308) in LV-FABP5+LY294002 group vs.
LV-NC+LY294002 group, P<0.05; Fig.
5I). However, LY294002 treatment did not affect the expression
of endogenous FABP5 (indicated as FABP5 only; Fig. 5F-H). Taken together, these results
suggest that the PI3K/AKT signaling pathway may participate in
FABP5-induced proliferation of ccRCC cells, and that inhibiting
PI3K/AKT signaling may suppress the pro-proliferative effects of
FABP5 in ccRCC cells.
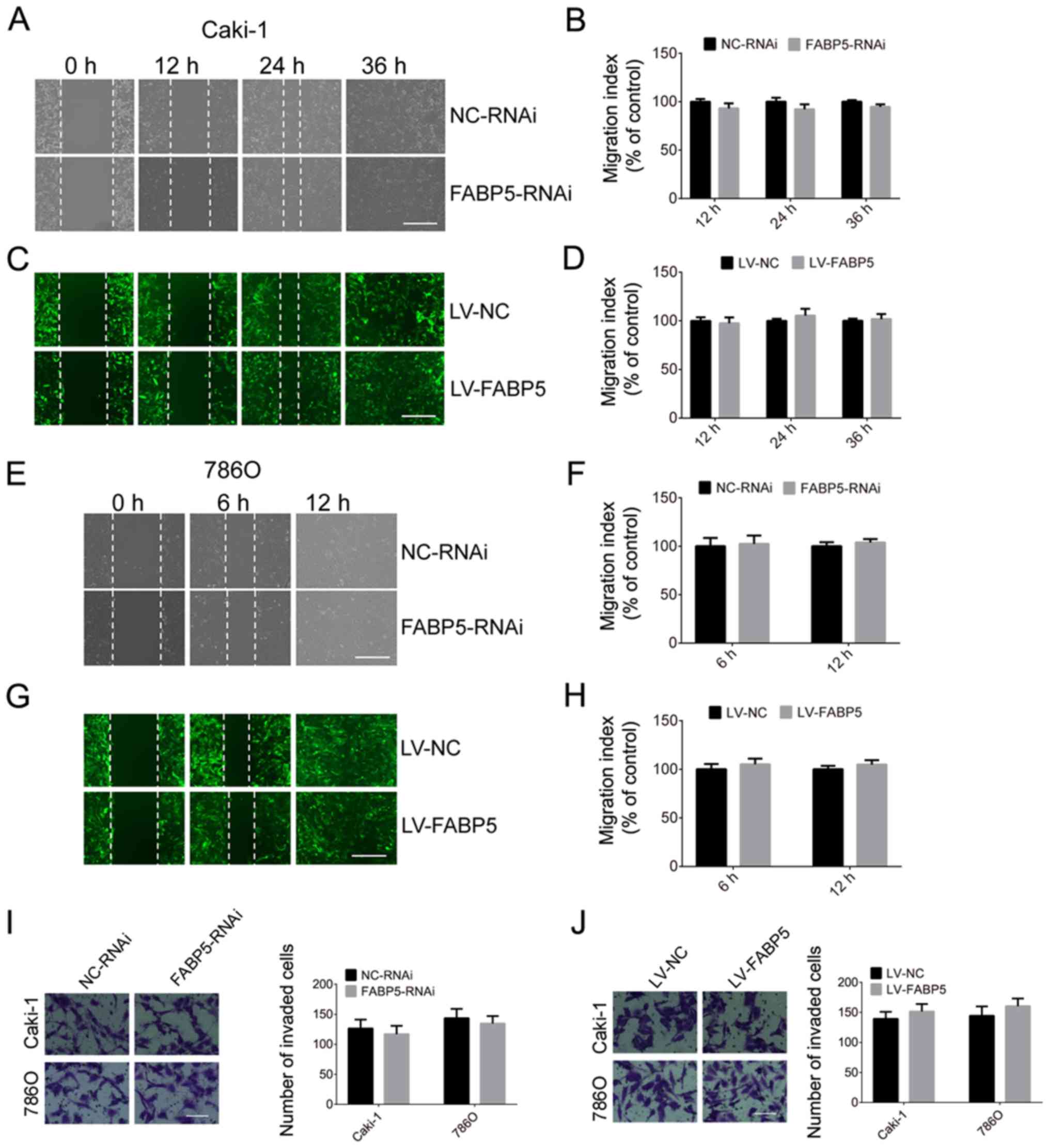
The migration and invasion abilities of Caki-1 and
786O cells in the FABP5-RNAi and NC-RNAi groups were then
investigated in the present study. As indicated in Fig. 6, silencing of FABP5 did not affect
the migration and invasion abilities of ccRCC cells at all time
points. Similarly, overexpression of FABP5 was not associated with
a significant effect on the migration or invasion of Caki-1 and
786O cells when compared with controls (Fig. 6).
FABP5 affects tumorigenesis in nude
mice
To evaluate the effect of FABP5 on tumorigenesis,
Caki-1 cells were injected into nude mice. The tumor volumes in the
FABP5-RNAi group of mice were significantly smaller than those in
the NC-RNAi groups (P<0.01; Fig. 7A
and B), and the maximum tumor diameter was 1.01 cm. The
proportion of Ki67-positive cells in the FABP5-RNAi group was also
significantly lower than that in the control group (P<0.01;
Fig. 7C and D). Furthermore, the
protein expression were normalized to β-actin, the FABP5 and p-AKT
were decreased in the FABP5-RNAi group (all P<0.001 vs. NC-RNAi
group apart from p-AKT (Thr308), P<0.01; Fig. 7E and F). However, following
inoculation of mice with FABP5-overexpressing Caki-1 cells, the
average volume of tumors in these mice (LV-FABP5 group) was
significantly larger than those in the LV-NC group (P<0.05;
Fig. 8A and B), and the maximum
tumor diameter was 1.41 cm. In addition, the proportion of
Ki67-positive cells was increased in LV-FABP5 group (P<0.01;
Fig. 8C and D), and the expression
of p-AKT in the LV-FABP5 group were significantly higher than that
in the LV-NC group when normalized to β-actin (P<0.01; Fig. 8E and F). The primary FABP5 antibody
is able to detect both endogenous FABP5 and exogenous FABP5-FLAG
expression. Exogenous expression of FABP5 (indicated as FABP5-FLAG
or FLAG) was also detected in FABP5-overexpressing cells,
indicating that exogenous FABP5 was successfully expressed in cells
of LV-FABP5 group Using the GV492 vector, the FABP5 sequence was
directly combined with the FLAG tag. Following transfection of
cells with LV-FABP5, exogenous FABP5-FLAG could be detected,
however this did not affect endogenous FABP5 expression (Fig. 8E and F). In addition, the FLAG tag
is ~3 kDa and FABP5 is ~15 kDa; therefore, exogenous FABP5-FLAG is
~18 kDa in size. As a result, the use of the FABP5 or FLAG
antibodies to detect exogenous FABP5-FLAG expression identified the
~18 kDa protein band in cells transfected with LV-FABP5, but not in
the LV-NC group. This confirmed that there was no exogenous FABP5
expression in the LV-NC group. These results support the notion
that FABP5 exerts a pro-proliferative role and contributes to
tumorigenesis in ccRCC in vivo.
Discussion
FABPs are a group of small, highly conserved
proteins that bind long-chain FAs (29). As FA transportation-associated
proteins, FABPs are divided into two groups according to their
cellular location; plasma membrane-associated FABPs and cytoplasmic
FABPs (30,31). Cytoplasmic FABPs are a group of
lipid-binding proteins with low molecular masses of ~14-15 kDa. The
primary function of cytoplasmic FABPs is their involvement in lipid
metabolism. Various FABPs exhibit unique patterns of expression in
tissues; however, FABPs are not exclusively expressed in certain
cell types (32). For instance,
FABP5 and FABP7 were identified in adult rat hippocampal neural
progenitor cells and in newborn rat neurons (33).
FAs function as signaling compounds to regulate
metabolic networks, survival pathways and inflammatory responses in
various cancers. As transporters of FAs, FABPs also modulate tumor
cell growth, metabolism, migration, differentiation, and
development (34). FABPs have been
identified as target genes of peroxisome proliferator-activated
receptors (PPARs) (35,36), and PPARs may also be stimulated by
FABPs to enhance their transcriptional activities in cancer cells
(37,38). In addition, numerous signaling
pathways and molecules may facilitate FABP-induced tumor
progression, such as the mitogen-activated protein kinase (MAPK)
(39), the Src/focal adhesion
kinase/cell division cycle 42 (40), reactive oxygen species (41) and epidermal growth factor receptor
(EGFR) (42) signaling
pathways.
FABP5, also known as epidermal FABP, is upregulated
in many types of tumors, and previous studies have also
demonstrated that FABP5 is overexpressed in several types of human
cancer cells (13,18,20,21,34).
In prostate cancer cells, FABP5 contains a typical CpG
island around its promoter region, and the overexpression of FABP5
is attributed to hypomethylation of this CpG island (43). Consequently, silencing of
FABP5 led to a significant decrease in cell proliferation.
FABP5 facilitates the malignant progression and tumorigenicity of
prostate cancer cells via the FABP5-PPARγ-vascular endothelial
growth factor signal transduction axis (44). FABP5 is also expressed at higher
levels in breast cancer tissues; the FABP5/PPARδ axis induces
breast cancer cell proliferation, migration and invasion by
activating EGFR (45,46), and suppression of FABP5 may present
a strategy to overcome retinoic acid (RA)-resistant breast cancer
(36). In addition, FABP5 promotes
tumorigenesis in numerous additional types of cancer, such
colorectal cancer (47), cervical
cancer (18), hepatocellular
carcinoma (21), and gastric
cancer (20). In the present
study, FABP5 expression was observed to be significantly higher in
samples from patients with ccRCC when compared with normal control
specimens, and FABP5 expression was significantly correlated with
tumor stage. These results suggest that FABP5 may be associated
with the malignant progression of ccRCC cells and may serve an
important role in tumorigenesis. In addition, the overall survival
and disease-free status curves indicated that survival rates were
significantly decreased in patients with higher FABP5
expression. Univariate Cox regression analysis further revealed
that high FABP5 expression was an unfavorable prognostic
parameter. Taken together, these results suggest that FABP5
expression significantly correlates with the clinical
characteristics and survival of patients with ccRCC.
The PI3K/AKT signaling pathway is normally activated
by extracellular signals in cells and functions as a crucial
intracellular signaling pathway in tumorigenesis. This pathway
regulates a number of functions in cancer cells, such as cellular
metabolism, tumor development, growth, proliferation and metastasis
(48,49). Progress in uncovering PI3K/AKT
alterations and their roles in tumorigenesis has enabled the
development of novel targeted molecules for anticancer treatment
(48-51). ccRCC is the most aggressive subtype
of RCC and accounts for the vast majority of kidney
cancer-associated deaths. This subtype is notoriously resistant to
traditional chemotherapy and radiotherapy (52). The primary signaling pathways
underlying ccRCC pathogenesis include the PI3K/AKT and MAPK
signaling pathways (53). Elevated
Notch1 signaling exerts its tumor growth-promoting effects via the
PI3K/AKT signaling pathway (54,55)
and the oncogenic effects of specific microRNAs and the long
non-coding RNA, promoter of CDKN1A antisense DNA damage activated
RNA, also involve the activation of this pathway (56,57).
In the present study, FABP5 was observed to promote
the proliferation of Caki-1 and 786O ccRCC cells. Silencing of
FABP5 significantly inhibited ccRCC cell proliferation,
while overexpressing FABP5 promoted cell proliferation. The
results also revealed that silencing of FABP5 significantly
decreased the p-AKT expression levels, while overexpression of
FABP5 upregulated p-AKT levels. The authors of the current
study hypothesized that FABP5 may regulate ccRCC cell proliferation
via the PI3K/AKT signaling pathway. To test this hypothesis, the
AKT inhibitor, LY294002, was used to inactivate the PI3K/AKT
signaling pathway. The results demonstrated that LY294002 treatment
attenuated the pro-proliferative effects of FABP5 in Caki-1 and
786O cells, suggesting that the PI3K/AKT signaling pathway, at
least in part, may be involved in FABP5-induced cell
proliferation.
It was previously reported that FABP5
silencing inhibited the invasion and migration of gastric cancer
cells and breast cancer cells (18-20).
By contrast, the present study observed that FABP5 silencing
did not affect the migration or invasion of ccRCC cells, indicating
that the intrinsic properties of ccRCC cells may differ from
gastric and breast cancer cells, and FABP5 might exert different
functions in different cancer cells, which required further
investigation. To better understand the role of FABP5 in
ccRCC, its effects on cell cycle progression, cell proliferation
and colony formation require further investigation in future
studies. In addition, future experiments investigating the
association between FABP5, AKT phosphorylation and cancer
metastasis will be performed.
In conclusion, the present study revealed that
FABP5 expression was upregulated in ccRCC samples when
compared with normal tissues. High FABP5 expression was
significantly correlated with tumor stage and predicted poor
patient survival. In addition, the results indicated that FABP5
exerted a pro-proliferative role in Caki-1 and 786O ccRCC cells
in vitro. Furthermore, the results indicated that the
PI3K/AKT signaling pathway may be involved in mediating
FABP5-induced ccRCC cell proliferation.
Funding
The present study was supported by the Health and
Family Planning Commission of Shanghai (grant no. 201740114), and
the Cultivation Project of Tongji Hospital in the National Natural
Science Foundation.
Availability of data and materials
All data used or analyzed in this study are included
in this published article or are available from the corresponding
author on reasonable request.
Authors’ contributions
QL, GW, YZ performed the majority of this
experiments. QL, GW and HL wrote the manuscript. XH, HL performed
the animal studies. WL, MZ acquired experimental materials. QD, CM
and PW contributed to the conception and design of this study. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Nantong University (Nantong, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Acknowledgments
Not applicable.
References
|
1
|
Volpe A and Patard JJ: Prognostic factors
in renal cell carcinoma. World J Urol. 28:319–327. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wood LS: Renal cell carcinoma: Screening,
diagnosis, and prognosis. Clin J Oncol Nurs. 13:3–7. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheville JC, Lohse CM, Zincke H, Weaver AL
and Blute ML: Comparisons of outcome and prognostic features among
histologic subtypes of renal cell carcinoma. Am J Surg Pathol.
27:612–624. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levy DA, Slaton JW, Swanson DA and Dinney
CP: Stage specific guidelines for surveillance after radical
nephrectomy for local renal cell carcinoma. J Urol. 159:1163–1167.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Figlin RA: Renal cell carcinoma:
Management of advanced disease. J Urol. 161:381–386; discussion
386–387 1999. PubMed/NCBI
|
|
6
|
Zhong C, Chen Y, Tao B, Peng L, Peng T,
Yang X, Xia X and Chen L: LIM and SH3 protein 1 regulates cell
growth and chemosensitivity of human glioblastoma via the PI3K/AKT
pathway. BMC Cancer. 18:7222018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shen GY, Ren H, Huang JJ, Zhang ZD, Zhao
WH, Yu X, Shang Q, Qiu T, Zhang YZ, Tang JJ, et al: Plastrum
Testudinis Extracts Promote BMSC Proliferation and Osteogenic
Differentiation by Regulating Let-7f–5p and the TNFR2/PI3K/AKT
Signaling Pathway. Cell Physiol Biochem. 47:2307–2318. 2018.
View Article : Google Scholar
|
|
8
|
Hou T, Zhou L, Wang L, Kazobinka G, Chen
Y, Zhang X and Chen Z: Leupaxin Promotes Bladder Cancer
Proliferation, Metastasis, and Angiogenesis Through the PI3K/AKT
Pathway. Cell Physiol Biochem. 47:2250–2260. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Y, Wang W, Li D, Li M, Wang P, Wen J,
Liang M, Su B and Yin Y: IGF-1 alleviates NMDA-induced
excitotoxicity in cultured hippocampal neurons against autophagy
via the NR2B/PI3K-AKT-mTOR pathway. J Cell Physiol. 229:1618–1629.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu W, Yang Z and Lu N: A new role for the
PI3K/Akt signaling pathway in the epithelial-mesenchymal
transition. Cell Adhes Migr. 9:317–324. 2015. View Article : Google Scholar
|
|
11
|
Sun P, Wang L, Lu Y, Liu Y, Li L, Yin L,
Zhang C, Zhao W, Shen B and Xu W: MicroRNA-195 targets VEGFR2 and
has a tumor suppressive role in ACHN cells via PI3K/Akt and
Raf/MEK/ERK signaling pathways. Int J Oncol. 49:1155–1163. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou J, Zhu G, Huang J, Li L, Du Y, Gao Y,
Wu D, Wang X, Hsieh JT, He D, et al: Non-canonical GLI1/2
activation by PI3K/AKT signaling in renal cell carcinoma: A novel
potential therapeutic target. Cancer Lett. 370:313–323. 2016.
View Article : Google Scholar
|
|
13
|
Gajda AM and Storch J: Enterocyte fatty
acid-binding proteins (FABPs): Different functions of liver and
intestinal FABPs in the intestine. Prostaglandins Leukot Essent
Fatty Acids. 93:9–16. 2015. View Article : Google Scholar
|
|
14
|
Storch J and Corsico B: The emerging
functions and mechanisms of mammalian fatty acid-binding proteins.
Annu Rev Nutr. 28:73–95. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Richieri GV, Ogata RT, Zimmerman AW,
Veerkamp JH and Kleinfeld AM: Fatty acid binding proteins from
different tissues show distinct patterns of fatty acid
interactions. Biochemistry. 39:7197–7204. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maximin E, Langelier B, Aïoun J, Al-Gubory
KH, Bordat C, Lavialle M and Heberden C: Fatty acid binding protein
7 and n-3 p oly unsaturated fatty acid supply in early rat brain
development. Dev Neurobiol. 76:287–297. 2016. View Article : Google Scholar
|
|
17
|
Lin H, Patel S, Affleck VS, Wilson I,
Turnbull DM, Joshi AR, Maxwell R and Stoll EA: Fatty acid oxidation
is required for the respiration and proliferation of malignant
glioma cells. Neuro-oncol. 19:43–54. 2017. View Article : Google Scholar :
|
|
18
|
Wang W, Chu HJ, Liang YC, Huang JM, Shang
CL, Tan H, Liu D, Zhao YH, Liu TY and Yao SZ: FABP5 correlates with
poor prognosis and promotes tumor cell growth and metastasis in
cervical cancer. Tumour Biol. 37:14873–14883. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang W, Jia HL, Huang JM, Liang YC, Tan H,
Geng HZ, Guo LY and Yao SZ: Identification of biomarkers for lymph
node metastasis in early-stage cervical cancer by tissue-based
proteomics. Br J Cancer. 110:1748–1758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao G, Wu M, Wang X, Du Z and Zhang G:
Effect of FABP5 gene silencing on the proliferation, apoptosis and
invasion of human gastric SGC-7901 cancer cells. Oncol Lett.
14:4772–4778. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ohata T, Yokoo H, Kamiyama T, Fukai M,
Aiyama T, Hatanaka Y, Hatanaka K, Wakayama K, Orimo T, Kakisaka T,
et al: Fatty acid-binding protein 5 function in hepatocellular
carcinoma through induction of epithelial-mesenchymal transition.
Cancer Med. 6:1049–1061. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang W, Zhang M, Peng Y and He J:
Ubiquitin Associated Protein 2-Like (UBAP2L) Overexpression in
Patients with Hepatocellular Carcinoma and its Clinical
Significance. Med Sci Monit. 23:4779–4788. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) μethod. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Han X, Li H, Zhang Y, Qin J, Yang Q, Wang
L, Yuan M and Xia C: Brain lipid-binding protein promotes
proliferation and modulates cell cycle in C6 rat glioma cells. Int
J Oncol. 51:1439–1448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang N, Meng C, Han X, Guo J, Li H and Yu
Z: Low-dose cisplatin causes growth inhibition and loss of
autophagy of rat astrocytes in vitro. Neurosci Lett. 682:112–117.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shi H, Xu J, Zhao R, Wu H, Gu L and Chen
Y: FGF2 regulates proliferation, migration, and invasion of ECA109
cells through PI3K/Akt signalling pathway in vitro. Cell Biol Int.
40:524–533. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leary S, Underwood W, Anthony R, Cartner
S, Corey D, Grandin T, Greenacre CB, Gwaltney-Bran S, McCrackin MA
and Meyer R: AVMA Guidelines for the Euthanasia of Animals. 2013
Edition. University of Alaska Anchorage; Schaumburg: 2013
|
|
28
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang W, Chen R, Yang T, Xu N, Chen J, Gao
Y and Stetler RA: Fatty acid transporting proteins: Roles in brain
development, aging, and stroke. Prostaglandins Leukot Essent Fatty
Acids. 136:35–45. 2018. View Article : Google Scholar
|
|
30
|
Berk PD, Wada H, Horio Y, Potter BJ,
Sorrentino D, Zhou SL, Isola LM, Stump D, Kiang CL and Thung S:
Plasma membrane fatty acid-binding protein and mitochondrial
glutamic-oxalo-acetic transaminase of rat liver are related. Proc
Natl Acad Sci USA. 87:3484–3488. 1990. View Article : Google Scholar
|
|
31
|
Stremmel W, Kochwa S and Berk PD: Studies
of oleate binding to rat liver plasma membranes. Biochem Biophys
Res Commun. 112:88–95. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Storch J and Thumser AE: The fatty acid
transport function of fatty acid-binding proteins. Biochim Biophys
Acta. 1486:28–44. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Matsumata M, Sakayori N, Maekawa M, Owada
Y, Yoshikawa T and Osumi N: The effects of Fabp7 and Fabp5 on
postnatal hippocampal neurogenesis in the mouse. Stem Cells.
30:1532–1543. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Amiri M, Yousefnia S, Seyed Forootan F,
Peymani M, Ghaedi K and Nasr Esfahani MH: Diverse roles of fatty
acid binding proteins (FABPs) in development and pathogenesis of
cancers. Gene. 676:171–183. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Boiteux G, Lascombe I, Roche E,
Plissonnier ML, Clairotte A, Bittard H and Fauconnet S: A-FABP, a
candidate progression marker of human transitional cell carcinoma
of the bladder, is differentially regulated by PPAR in urothelial
cancer cells. Int J Cancer. 124:1820–1828. 2009. View Article : Google Scholar
|
|
36
|
Schug TT, Berry DC, Toshkov IA, Cheng L,
Nikitin AY and Noy N: Overcoming retinoic acid-resistance of
mammary carcinomas by diverting retinoic acid from PPARbeta/delta
to RAR. Proc Natl Acad Sci USA. 105:7546–7551. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Morgan E, Kannan-Thulasiraman P and Noy N:
Involvement of Fatty Acid Binding Protein 5 and PPARβ/δ in Prostate
Cancer Cell Growth. PPAR Res. 2010.pii: 234629. 2010. View Article : Google Scholar
|
|
38
|
De Rosa A, Pellegatta S, Rossi M, Tunici
P, Magnoni L, Speranza MC, Malusa F, Miragliotta V, Mori E,
Finocchiaro G, et al: A radial glia gene marker, fatty acid binding
protein 7 (FABP7), is involved in proliferation and invasion of
glioblastoma cells. PLoS One. 7:e521132012. View Article : Google Scholar
|
|
39
|
Lee D, Wada K, Taniguchi Y, Al-Shareef H,
Masuda T, Usami Y, Aikawa T, Okura M, Kamisaki Y and Kogo M:
Expression of fatty acid binding protein 4 is involved in the cell
growth of oral squamous cell carcinoma. Oncol Rep. 31:1116–1120.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ku CY, Liu YH, Lin HY, Lu SC and Lin JY:
Liver fatty acid-binding protein (L-FABP) promotes cellular
angiogenesis and migration in hepatocellular carcinoma. Oncotarget.
7:18229–18246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Song GX, Shen YH, Liu YQ, Sun W, Miao LP,
Zhou LJ, Liu HL, Yang R, Kong XQ, Cao KJ, et al: Overexpression of
FABP3 promotes apoptosis through inducing mitochondrial impairment
in embryonic cancer cells. J Cell Biochem. 113:3701–3708. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liang Y, Bollen AW, Aldape KD and Gupta N:
Nuclear FABP7 immunoreactivity is preferentially expressed in
infiltrative glioma and is associated with poor prognosis in
EGFR-overexpressing glioblastoma. BMC Cancer. 6:972006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kawaguchi K, Kinameri A, Suzuki S, Senga
S, Ke Y and Fujii H: The cancer-promoting gene fatty acid-binding
protein 5 (FABP5) is epigenetically regulated during human prostate
carcinogenesis. Biochem J. 473:449–461. 2016. View Article : Google Scholar
|
|
44
|
Forootan FS, Forootan SS, Gou X, Yang J,
Liu B, Chen D, Al Fayi MS, Al-Jameel W, Rudland PS, Hussain SA, et
al: Fatty acid activated PPARγ promotes tumorigenicity of prostate
cancer cells by up regulating VEGF via PPAR responsive elements of
the promoter. Oncotarget. 7:9322–9339. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Levi L, Lobo G, Doud MK, von Lintig J,
Seachrist D, Tochtrop GP and Noy N: Genetic ablation of the fatty
acid-binding protein FABP5 suppresses HER2-induced mammary
tumorigenesis. Cancer Res. 73:4770–4780. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Powell CA, Nasser MW, Zhao H, Wochna JC,
Zhang X, Shapiro C, Shilo K and Ganju RK: Fatty acid binding
protein 5 promotes metastatic potential of triple negative breast
cancer cells through enhancing epidermal growth factor receptor
stability. Oncotarget. 6:6373–6385. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kawaguchi K, Senga S, Kubota C, Kawamura
Y, Ke Y and Fujii H: High expression of Fatty Acid-Binding Protein
5 promotes cell growth and metastatic potential of colorectal
cancer cells. FEBS Open Bio. 6:190–199. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ciruelos Gil EM: Targeting the
PI3K/AKT/mTOR pathway in estrogen receptor-positive breast cancer.
Cancer Treat Rev. 40:862–871. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Singh SS, Yap WN, Arfuso F, Kar S, Wang C,
Cai W, Dharmarajan AM, Sethi G and Kumar AP: Targeting the PI3K/Akt
signaling pathway in gastric carcinoma: A reality for personalized
medicine? World J Gastroenterol. 21:12261–12273. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Polivka J Jr and Janku F: Molecular
targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol
Ther. 142:164–175. 2014. View Article : Google Scholar
|
|
51
|
Fumarola C, Bonelli MA, Petronini PG and
Alfieri RR: Targeting PI3K/AKT/mTOR pathway in non small cell lung
cancer. Biochem Pharmacol. 90:197–207. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Guo H, German P, Bai S, Barnes S, Guo W,
Qi X, Lou H, Liang J, Jonasch E, Mills GB, et al: The PI3K/AKT
Pathway and Renal Cell Carcinoma. J Genet Genomics. 42:343–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Apanovich NV, Peters MV, Apanovich PV,
Kamolov BS, Matveev VB, Ginter EK and Karpukhin AV: Expression
Profiles of Genes-Potential Therapy Targets-and Their Relationship
to Survival in Renal Cell Carcinoma. Dokl Biochem Biophys.
478:14–17. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xu L, Zhu Y, Xu J, Wu K, Li J, Xu W, Liu
H, Wang S, Yin H, Chen L, et al: Notch1 activation promotes renal
cell carcinoma growth via PI3K/Akt signaling. Cancer Sci.
103:1253–1258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Liu S, Ma X, Ai Q, Huang Q, Shi T, Zhu M,
Wang B and Zhang X: NOTCH1 functions as an oncogene by regulating
the PTEN/PI3K/AKT pathway in clear cell renal cell carcinoma. Urol
Oncol. 31:938–948. 2013. View Article : Google Scholar
|
|
56
|
Nogueira I, Dias F, Teixeira AL and
Medeiros R: miRNAs as potential regulators of mTOR pathway in renal
cell carcinoma. Pharmacogenomics. 19:249–261. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xu Y, Tong Y, Zhu J, Lei Z, Wan L, Zhu X,
Ye F and Xie L: An increase in long non-coding RNA PANDAR is
associated with poor prognosis in clear cell renal cell carcinoma.
BMC Cancer. 17:3732017. View Article : Google Scholar : PubMed/NCBI
|