Introduction
Colorectal cancer (CRC) is the third commonest type
of cancer worldwide (1). Early
clinical manifestations of CRC are nonspecific and it demonstrates
significant tumor growth and early metastatic potential (2-4).
Despite improvements in early diagnosis and systemic therapies,
China and developed countries in Europe and the United States
contribute to >50% of new CRC cases and associated mortalities
globally (5). Consequently, there
is an essential need to improve the understanding of the
pathophysiology of CRC and provide CRC patients with novel
treatment options.
Long noncoding RNAs (lncRNAs) are transcripts
>200 nucleotides that do not encode proteins (6). lncRNAs are involved in a range of
molecular mechanisms of cancer including miRNA sponging and RNA
degradation (7-10). As biological techniques have
advanced, numerous CRCrelated lncRNAs, including lncRNA GLCC1
(11), lncRNA ITGB8-AS1 (12) and H19 (13), have been linked to the progression
of CRC via a range of mechanisms. The long intergenic noncoding RNA
2038 (LINC02038), which is located on chromosome 3q29, was found to
be associated with the growth of CRC and serving as a crucial
survival-related lncRNA (14).
lncRNAs can function as competitive endogenous (ce)RNAs, which bind
to microRNAs (miRNAs/miRs) in a competitive manner (15). Therefore, investigating the
LINC02038 ceRNA pathways in CRC may uncover additional therapeutic
options for the management of CRC. The location of the family with
sequence similarity 172 member A (FAM172A) was discovered in
chromosome 5q15. According to previous studies, FAM172A expression
is highly tissue-specific and it inhibits the growth and metastasis
of hepatocellular carcinoma (16).
Overexpression of the PI3K/AKT signaling pathway has been linked to
several types of cancer, including CRC and the PI3K/AKT signaling
pathway is essential for the growth and metastasis of CRC (17).
In humans, the most prevalent form of RNA
modification is N6-methyladenosine (m6A) modification,
which regulates RNA stability (18,19).
A group of proteins regulate the cytoplasmic m6A
condition, termed 'writers' such as METTL3 and METTLI4 and
'readers' such as YTHDF2 and YTHDC2 and it has been shown that
METTL3 was an important gene for CRC cell development (20).
In the present study, a correlation between the
expression of LINC02038 and the clinical value of CRC patients was
identified. Moreover, the functions of LINC02038 in cell growth,
apoptosis and invasion from a functional analysis were ascertained.
Further investigation confirmed that LINC02038 served as a sponge
for miR-552-5p in CRC, enhancing its suppressive effect on FAM172A
through the PI3K/Akt pathway. Furthermore, METTL3 exerted its
function through weakening LINC02038 expression in an
m6A-YTHDF2-dependent manner. LINC02038 may provide novel
CRC therapeutic approaches.
Materials and methods
Patients and tissue samples
A total of 68 CRC cases in total were collected from
patients who received surgery at the First People's Hospital of
Foshan between January 2018 and January 2020. Prior to resection,
these individuals had not undergone chemotherapy or radiotherapy.
All specimens were snap-frozen in liquid nitrogen and stored at
-80°C before use or fixed and paraffin-embedded. All patients
provided informed consent and the Ethics Committee of the First
People's Hospital of Foshan approved the present study (approval
no. AF-SOP-18-1.6-2). The clinicopathological data of the patients
are presented in Table I.
 | Table IRelationship between LINC02038
expression and clinical characteristics in patients with colorectal
cancer (n=68). |
Table I
Relationship between LINC02038
expression and clinical characteristics in patients with colorectal
cancer (n=68).
| Parameter | Number of
patients | LINC02038
expression
| P-value |
|---|
| Low | High |
|---|
| Patients (n) | 68 | 35 | 33 | |
| Age (years) | | | | 0.902 |
| ≤60 | 32 | 17 | 15 | |
| >60 | 36 | 18 | 18 | |
| Sex | | | | 0.736 |
| Female | 26 | 13 | 13 | |
| Male | 42 | 22 | 20 | |
| TNM stage | | | | |
| I-II | 29 | 10 | 19 | 0.011 |
| III-IV | 39 | 25 | 14 | |
| Distant
metastasis | | | | 0.024 |
| Absent | 57 | 27 | 30 | |
| Present | 11 | 8 | 3 | |
|
Differentiation | | | | |
| Poorly or
moderately | 38 | 21 | 17 | 0.411 |
| Well | 30 | 14 | 16 | |
| Tumor depth | | | | |
| T1T2 | 32 | 12 | 20 | 0.022 |
| T3T4 | 36 | 23 | 13 | |
Cell culture and transfection
The human colorectal carcinoma cell lines used were
LoVo, RKO, SW480, HCT116, DLD-1 and HT-29 cells and NCM460 normal
colonic epithelial cell line were used as the control cells. All
cells were obtained from the Cell Bank of Type Culture Collection
of the Chinese Academy of Sciences. STR profiling was performed to
verify the identity of the HT-29 cells. Cells were cultured in
RPMI-1640 media supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) in a humidified incubator (95% humidity) supplied
with 5% CO2. Cells were routinely checked for Mycoplasma
contamination. The overexpression vectors pcDNA3.1/Control (Vector)
and pcDNA3.1/LINC02038 (LINC02038) were purchased from Shanghai
GenePharma Co., Ltd.. Empty vector was used as the negative control
(NC) for LINC02038 overexpression. A total of 3×105
cells/well were transfected with empty vector or pcDNA3.1-LINC02038
plasmids (1.5 µg/ml), a total of 30 pmol/l miR-552-5p mimics
or mimic NC at room temperature for 48 h. The cells were collected
for subsequent experiments at 48 h following transfection.
miR-552-5p mimic and its negative control mimic for miR-552-5p,
miR-552-5p inhibitor, were purchased from Shanghai GenePharma Co.,
Ltd.. The sequences were as follows: miR-552-5p mimics 5′-GUU UAA
CCU UUU GCC UGU UGG-3′ and mimic NC 5′-UCA CAA CCU CCU AGA AAG AGU
AGA-3′. Small interfering (si)RNAs against FAM172A (siFAM172A) and
its scramble control siRNA were purchased from Shanghai GenePharma
Co., Ltd. Vectors containing short hairpin (sh)RNAs targeting
METTL3 (shMETTL3, i.e., shMETTL3-1 and shMETTL3-2) or its
non-targeting shRNA sequence negative control (shNC METTL3),
vectors containing short hairpin RNAs (shRNAs) targeting YTHDF2
(shYTHDF2, i.e., shYTHDF2-1 and shYTHDF2-2) or its non-targeting
shRNA sequence negative control (shNC YTHDF2) were subcloned into
Lv5 lentiviruses (Shanghai GenePharma Co., Ltd.) and were then
transduced into the target cells. Transfection was performed using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The efficacy of transfection was confirmed by
reverse transcription-quantitative (RT-q) PCR.
Bioinformatics prediction
LINC02038, miR-552-5p and FAM172A levels were
determined using The Cancer Genome Atlas (TCGA; https://cancergenome.nih.gov/) and the GEPIA
(http://gepia.cancer-pku.cn) databases in
CRC. The biological roles of LINC02038 were identified using the
Kyoto Encyclopedia of Genes and Genomes (KEGG; https://www.genome.jp/kegg/). The miRcode (https://www.mircode.org/) and DIANA-LncBase Predicted
v.2 (https://diana.e-ce.uth.gr/lncbasev2/home) online set
of tools was used to predict the miRNAs targeted by LINC02038.
TargetScan (http://www.targetscan.org/vert72/) and PicTar
(https://pictar.mdcberlin.de) were used
to predict mRNA target sites of miR-552-5p. SRAMP (http://www.cuilab.cn/sramp) was used to predict the
possible m6A modification locations of LINC02038.
RTqPCR
TRIzol® (Thermo Fisher Scientific, Inc.)
was used to isolate total RNA from tissues and cells at a density
of 3×106. A total of 500 ng RNA was used to synthesize
cDNA by using a reverse transcriptase cDNA synthesis kit according
to the manufacturer's protocol (Takara Bio, Inc.). The Hairpin-it
MicroRNA and U6 snRNA Normalization RT-PCR Quantification kits were
used to perform RT for miRNAs (Shanghai GenePharma, Co., Ltd.)
according to the manufacturer's protocol. mRNA expression was
determined using SYBR Premix ExTaq II (Takara Bio, Inc.) according
to the manufacturer's protocol. The internal controls were GAPDH
and U6. The thermocycling conditions were: Initial denaturation at
95°C for 40 sec followed by 40 cycles of 95°C for 10 sec, 20 sec at
60°C and dissociation at 72°C for 30 sec with a final extension
step of 72°C for 6 min. The 2−∆∆Cq method was used to
calculate the fold change (21).
The primer sequences were: LINC02038 forward, 5′-ACC GTT TTT GAT
GGT GCT GC-3′ and reverse, 5′-AAA ACG CCT CTG TCG CAA AC-3′; METTL3
forward, 5′-AAG CTG CAC TTC AGA CGA AT-3′ and reverse,5′-GGA ATC
ACC TCC GAC ACT C-3′; YTHDF2 forward, 5′-AGC CCC ACT TCC TAC CAG
ATG-3′ and reverse, 5′-TGA GAA CTG TTA TTT CCC CAT GC-3′; GAPDH
forward, 5′-AAT GGG CAG CCG TTA GGA AA-3′ and reverse, 5′-GCC CAA
TAC GAC CAA ATC AGA G-3′; miR-552-5p forward, 5′-GTT TAA CCT TTT
GCC TGT TGG-3′ and reverse, 5′-CGA ACG CTT CAC GAA TTT G-3′; U6
forward, 5′-CTC GCT TCG GCA GCA CA-3′ and reverse, 5′-AAC GCT TCA
CGA ATT TGC GT-3′; and FAM172A forward, 5′-CAA CGA GAA GCC GAT
GTA-3′ and reverse, 5′-GAT GTG TCT AAT GGT TCT GAG-3′. All assays
were repeated at least three times.
Cell Counting Kit8 (CCK8) assay
Cell vitality was determined using a CCK-8 assay
(Dojindo Molecular Technologies, Inc.). LoVo and SW620 cells
(1×103 cells/well) were plated into 96-well plates.
After 24, 48, 72 or 96 h of transfection, each well was filled with
RPMI-1640 containing 10% CCK-8 solution and cells were further
incubated at 37°C for 1 h. To assess the effectiveness of cell
growth, a microplate reader was used to measure the absorption at
450 nm (Thermo Fisher Scientific, Inc.).
Flow cytometry
To fix cells, cells (2×102) were placed
in 70% precooled ethanol overnight at 4°C. To analyze cell cycle
distribution, 400 µl propidium iodide (PI; Beyotime
Institute of Biotechnology) was added to stain the cells, followed
by incubation in the dark for 25 min at room temperature at 37°C. A
flow cytometer (FACScan; BD Biosciences) was used to determine the
proportion of cells in each phase of the cell cycle. The data were
analyzed using FlowJo 10 software (FlowJo LLC).
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) kits (Beyotime Institute of
Biotechnology) were used to examine cell apoptosis. The cell pellet
was washed with 4°C precooled PBS followed by 1X binding buffer
(centrifugation 300 × g, 5 min at 4°C). A total of 250 µl 1X
binding buffer was used to resuspend the cells and 5 µl
Annexin V-FITC/PI staining was applied to the cells for 20 min at
room temperature in the dark. Subsequently, 100 µl 1X
binding buffer was added and flow cytometry was performed. The data
(the percentage of early + late apoptotic cells) was examined using
FlowJo 10 (FlowJo LLC).
Cell colony formation assay
A total of 1×103 cells/well were
incubated in 6-well plates using RPMI-1640 media containing 10% FBS
at 37°C and 5% CO2 for 14 days. Subsequently, cells were
fixed using methanol for 15 min and stained with 0.5% crystal
violet at room temperature for 8 min. The colonies were counted
using ImageJ software (version 1.5; National Institutes of
Health).
Transwell assays
Cell migration and invasion were assessed using
Transwell inserts pre-coated with Matrigel at 37°C for 1 h or
without Matrigel (Corning Inc.). To the upper chamber,
1×104 cells suspended in 200 µl serum-free RPMI
1640 was added. To the lower chamber, 600 µl supplemented
media was added. After 48 h of incubation, the cells that had
migrated to the lower chamber were fixed using 4% paraformaldehyde
for 30 min at room temperature and then stained with Giemsa for 15
min at room temperature. The number of cells in five randomly
selected fields of view were imaged using a light microscope
(Olympus Corporation). The number of cells that had migrated or
invaded per field was determined using ImageJ (version 1.5;
National Institutes of Health).
Fluorescence in situ hybridization (FISH)
assays
GFP-labeled LINC02038 probes were supplied by
Shanghai GenePharma Co., Ltd. A FISH kit (Shanghai GenePharma Co.,
Ltd.) was used to perform the hybridization. After being preserved
with 4% paraformaldehyde at room temperature for 10 min,
2×104 cells/well were permeabilized with 0.5% Triton
X-100. and then washed three times with PBS. The cells were then
supplemented with a prehybridization solution and blocked for 25
min at 37°C. Cy3-labeled lncRNA FISH probes (Shanghai GenePharma
Co., Ltd.) were used to detect LINC02038. The 2 µl and 15
µM probe mixture were added to the 100 µl
hybridization solution and this solution was added to cells for
hybridization overnight at 37°C. The nuclei were stained for
fluorescence detection with DAPI at room temperature for 10 min and
then mounted on glass slides in the dark setting. The fluorescence
images were captured with an IX81 inverted fluorescence microscope
with 5 fields of view being assessed per slide (Olympus
Corporation).
Dual-luciferase reporter assay
In LoVo and SW620 cells, miR-552-5p interactions
with LINC02038 or FAM172A were identified. Cells were
co-transfected with vector, pmirGLO LINC02038 wild type (WT) or
LINC02038 mutant (MUT), FAM172A WT or FAM172A MUT (Promega
Corporation) as well as miR-552-5p mimic using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Luciferase activity was determined after 48 h of
transfection using the Dual-Luciferase Reporter assay system
(Promega Corporation) according to the manufacturer's protocol.
RNA immunoprecipitation (RIP)
To examine the relationship between miR-552-5p with
LINC02038 or FAM172A in CRC cells, a Magna RIP RNA-Binding Protein
Immunoprecipitation kit (cat: no. 17-700; Sigma, Millipore) and
anti-argonaute 2 (Ago2) antibody (5 µg; cat. no. ab32381;
Abcam) was used according to the manufacturer's protocol. Cells
were centrifuged at 1,000 × g for 8 min at 4°C and lysed in 600
µl RIP buffer (Millipore, Sigma). A total of 100 µl
lysate was then treated with 5 µl normal IgG or 5 µl
Ago2 antibody. The levels of LINC02038 or FAM172A and miR-552-5p
enriched on beads were evaluated by RT-qPCR.
Western blotting
RIPA lysate solution (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to lyse the cells. A BCA assay (Beyotime
Institute of Biotechnology) was used to determine the protein
concentration. Total protein extracts (20 µg) was loaded on
a 10% SDS-gel, resolved using SDS-PAGE, transferred to PVDF
membranes (MilliporeSigma) and blocked for 1 h with 5% non-fat milk
at room temperature. After blocking, membranes were incubated with
one of the primary antibodies overnight at 4°C. The antibodies used
were: anti-FAM172A (cat. no. ab121364; 1:2,000; Abcam), anti-METTL3
(cat. no. ab195352; 1:1,000; Abcam), anti-YTHDF2 (cat. no.
ab220163; 1:1,000; Abcam), anti-AKT (T308) (cat. no. ab38449;
1:1,000; Abcam), anti-AKT (cat. no. ab8805; 1:500; Abcam),
anti-PI3K (cat. no. ab302958; 1:1,000; Abcam) and anti-GAPDH (cat.
no. ab8245; 1:4,000; Abcam). After washing with PBS, the membranes
were probed for 1 h at room temperature and incubated with goat
anti-rabbit IgG secondary antibody (cat. no. ab97080; 1:5,000;
Abcam) and goat anti-mouse IgG secondary antibody (cat. no.
ab97040; 1:5,000; Abcam). Pierce ECL reagent (Pierce; Thermo Fisher
Scientific, Inc.) was used to observe the protein bands and
ImagePro Plus software version 6.0 (Media Cybernetics, Inc.) was
used for densitometry analysis.
In vivo experiments
A total of 18 male BALB/c nude mice (age, 4 weeks;
weight, 14-18 g) were purchased from Southern Medical University
Experimental Animal Centre and were kept in the pathogen-free-grade
research center at 28°C with 50% humidity, with a 12 h light/dark
cycle and ad libitum access to food and water. The
Institutional Animal Care and Use Committee of the Southern Medical
University approved the animal experiments (approval no. 2032101).
A total of 2×104 LoVo cells stably overexpressing
LINC02038 or LINC02038 or pcDNA3.1/Control (Vector) were
subcutaneously injected in the dorsal flank of mice. The weight and
growth of the tumor were assessed every 5 days. The volume of the
tumor was calculated as follows: Volume
(mm3)=(width2 × length)/2. At 3 weeks
following inoculation, cervical dislocation was used to euthanize
the mice after isoflurane inhalation (5% for 5 min). Finally, tumor
tissues were used for immunohistochemistry (IHC) analysis. The
National Institutes of Health Guide for the Care and Use of
Laboratory Animals was followed when conducting the study. The
maximum tumor volume observed was 534 mm3 and the
maximum tumor diameter was 17 mm.
Immunohistochemistry (IHC)
After being fixed with 4% paraformaldehyde at 4°C
for 24 h and then dehydrated in ethanol from low to high
concentration, permeabilized using xylene and embedded in paraffin,
tumor tissues were embedded in paraffin and sectioned at 4
µm. For inhibiting endogenous peroxidases, dewaxed portions
were cleared with xylene and were then rehydrated in an descending
ethanol gradient at room temperature for 15 min prior to incubation
with 3% H2O2 for 10 min. For antigen
retrieval, samples were placed in citrate solution (pH 6.0) at 95°C
for 15 min. The plates were treated with primary antibodies against
Ki-67 (cat. no. ab15580; 1:400; Abcam) and FAM172A (cat. no.
ab121364; 1:300; Abcam) overnight at 4°C. Subsequently, the
sections were treated with an HRP-conjugated secondary antibody for
60 min at 37°C prior to incubation with diaminobenzidine (DAB). The
slices were counterstained with hematoxylin for 3 min at room
temperature. Images were captured using a light microscope (Olympus
Corporation).
m6A quantification
Total RNA was quantified and purified using a
Dynabeads mRNA Purification kit (Thermo Fisher Scientific, Inc.).
The m6A quantification of mRNA was measured using the
EpiQuik m6A Methylation Quantification kit according to
the manufacturer's protocol (Colorimetric; EpigenTek).
m6A-RNA
immunoprecipitation-qPCR (MeRIP-qPCR)
MeRIP-qPCR was performed as described previously
(22). Briefly, mRNA was isolated,
purified and fragmented chemically using Ambion fragmentation
reagent (Thermo Fisher Scientific, Inc.) into 100-nt pieces prior
to being incubated with 15 µl ChIP Protein Magnetic Beads
(MilliporeSigma) conjugated to 2 µg of anti-m6A
polyclonal antibody (cat. no. ab208577; Abcam) or mouse control IgG
in 1X IPP buffer, 250 ng mRNA fragments were denatured at 65°C for
5 min. Subsequently, incubation of mRNA with m6A-bound
beads in IPP buffer at 4°C with constant rotation for 2 h was
washed with different types of buffers in turn. As the final step,
the RNA was eluted and purified using Qiagen RNeasy columns (Qiagen
GmbH) in 200 µl RNase-free water. The abundance of RNA was
assessed using qPCR as described above according to the
manufacturer's protocol. The experiments were repeated three
times.
RNA stability
A total of 1×103 cells/well were
incubated with 10 g/ml actinomycin D overnight in a
CO2-free humidified incubator. Total RNA was isolated
and quantified after 0, 1, 2, 4, or 8 h of actinomycin D treatment,
using RT-qPCR as described above according to the manufacturer's
protocol. The experiments were repeated three times.
Statistical analysis
Analysis was performed using GraphPad Prism version
9.0 (Dotmatics). Data are presented as the mean ± standard
deviation. A Student's t-test or a one-way ANOVA followed by
Tukey's test was used to evaluate differences between groups. The
association between LINC02038 transcript and clinicopathological
characteristics were investigated using a Fisher's exact test or a
χ2 test. Using linear regression analysis, the
association between two genes was analyzed. Kaplan-Meier analysis
with log-rank test was used to assess the overall survival rate.
P<0.05 was considered to indicate a statistically significant
difference.
Results
LINC02038 is downregulated in CRC
The expression patterns of lncRNAs in CRC specimens
were obtained from the TCGA and GTEx databases. LINC02038 was one
of the most downregulated lncRNAs with a fold change (FC) of ≥5 and
a P-value of <0.05 (Fig. 1A),
indicating that it could be a tumor suppressor lncRNA as well as a
therapeutic target and prognostic biomarker in CRC. A total of
1,263 differentially expressed lncRNAs were identified (Fig. 1B). The expression of LINC02038 in
various tumor types was assessed and found to be lower than in
normal samples when analyzed using the TCGA and GTEx datasets
(Fig. 1C). Additionally, GEO
databases were examined using the R tool to determine LINC02038
expression. The results showed that LINC02038 expression was
typically lower in tumor specimens compared to normal tissues
(Fig. 1D and E). Furthermore,
LINC02038 expression was positively associated with higher overall
survival (OS) according to the GEPIA database (Fig. 1F). KEGG analysis showed that
LINC02038 was closely related to the PI3K/Akt signaling pathway
(Fig. 1G). Subsequently, the
potential clinical relevance of LINC02038 was investigated. The
results showed positive associations between low LINC02038
expression and high invasion, distant metastasis and higher TNM
stage (Table I).
 | Figure 1LINC02038 is downregulated in CRC.
(A) Heatmap of lncRNAs. lncRNAs with an FC ≥5 and P<0.05 were
considered down- or upregulated. Downregulation is represented by a
blue scale, while upregulation is represented by a red scale. (B)
lncRNA volcano diagram of CRC vs. non-tumorous tissues. Upregulated
genes are indicated in red and downregulated genes are indicated in
green (defined by FC ≥1 and P<0.05). Genes in black were
differentially expressed by FC <1. (C) Expression of LINC02038
based on pan-cancer analysis utilizing the TCGA and GTEx databases.
LINC02038 expression was lower in COAD tissues than in normal
colorectal tissues. (D and E) The expression of LINC02038 in CRC
and normal colorectal tissues based on data obtained from GTEx and
TCGA datasets. (F) The relationship between the LINC02038 and OS in
the obtained datasets. (G) Bubble plots showing the top 21
dysregulated KEGG pathways in terms of genes associated with
LINC02038 in CRC. *P<0.05, ***P<0.001.
CRC, colorectal cancer; lncRNA, long non-coding RNA; TCGA, The
Cancer Genome Atlas; GTEx, Genotype-Tissue Expression project;
KEGG, Kyoto Encyclopedia of Genes and Genomes; COAD, colon
adenocarcinoma; OS, overall survival; ns, no significance. |
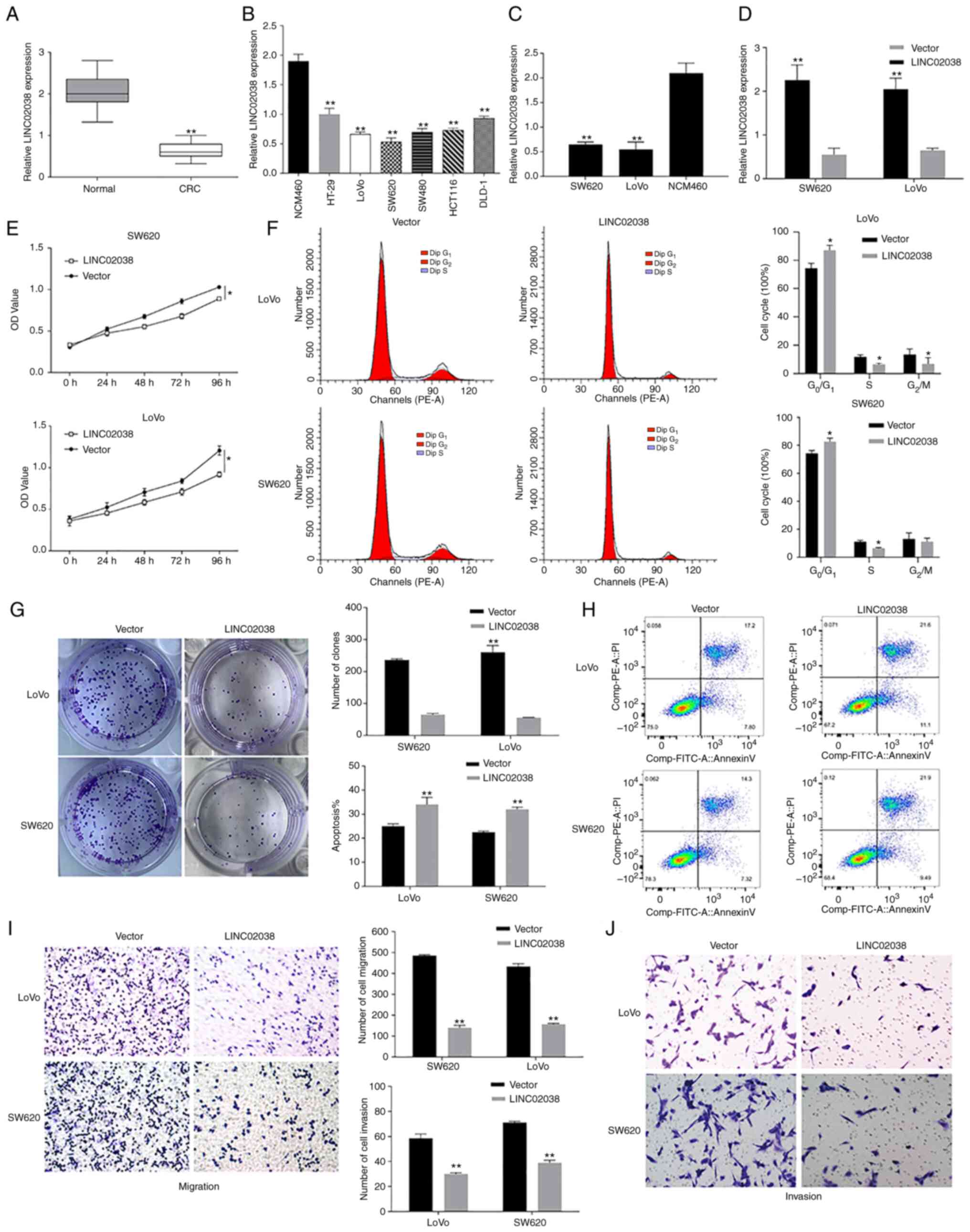
LINC02038 regulates apoptosis and
proliferation, migration and invasion of CRC cells
To further verify the potential of LINC02038, its
expression was analyzed in colorectal cancer (CRC) specimens and
cell lines (HT-29, LoVo, SW620, SW480, HCT116 and DLD-1) using
RT-qPCR. RT-qPCR analysis showed that LINC02038 was considerably
downregulated in CRC tissues relative to adjacent normal colorectal
tissues (n=68 pairs; Fig. 2A). In
comparison with normal colorectal epithelial cells (NCM460), the
expression of LINC02038 was greatly decreased in various CRC cell
lines (Fig. 2B,C). To determine
the role of LINC02038 in CRC progression, LoVo and SW620 cells were
transfected with pcDNA3.1-LINC02038. RT-qPCR results showed that
transfection with the LINC02038 vector dramatically increased
LINC02038 expression (Fig. 2D).
CCK-8 assay indicated that the LINC02038-overexpressed LoVo and
SW620 cells proliferated a reduced amount than the control cells
(Fig. 2E). Colony formation assay
results demonstrated that the upregulation of LINC02038 inhibited
the growth of LoVo and SW620 cells (Fig. 2G). Notably, an increasing number of
CRC cells in the G1/G0 phase and a decrease
in the number in the S phase were detected in response to LINC02038
overexpression compared to control groups using flow cytometry
(Fig. 2F). Furthermore, flow
cytometry results showed that overexpression of LINC02038 increased
the percentage of apoptotic cells compared to the empty vector
group (Fig. 2H). Cell motility was
measured using Transwell assays and the results showed that
overexpression of LINC02038 hindered migration and invasion
capacities of LoVo and SW620 cells (Fig. 2I and J).
LINC02038 acts as a sponge for miR-552-5p
in CRC
As the role of lncRNAs can be determined by their
subcellular location, the subcellular localization of LINC02038 was
observed using FISH. The results showed that LINC02038 was
predominantly localized in the cytoplasm (Fig. 3A). Additionally, the binding sites
between LINC02038 and miR-552-5p were predicted using miRcode and
LncBase Predicted v.2 (Fig. 3B).
Furthermore, the relationship between LINC02038 and miR-552-5p was
investigated in data on patients with primary colon adenocarcinoma
(COAD) obtained from TCGA. The results showed that LINC02038
expression was found to be inversely associated with miR-552-5p
expression in CRC (Fig. 3C). Using
the TCGA-COAD dataset, it was demonstrated that miR-552-5p
expression was significantly increased in CRC tissues when compared
with the adjacent normal tissues (Fig.
3D). This result was confirmed using RT-qPCR analysis which
showed that miR-552-5p expression was higher compared with adjacent
tissues (Fig. 3E) and cells
(Fig. 3F) than that of the
adjacent non-tumor tissues and NCM460 cells. Additionally,
LINC02038 overexpression substantially reduced miR-552-5p levels of
CRC cells based on the results of RT-qPCR (Fig. 3G).
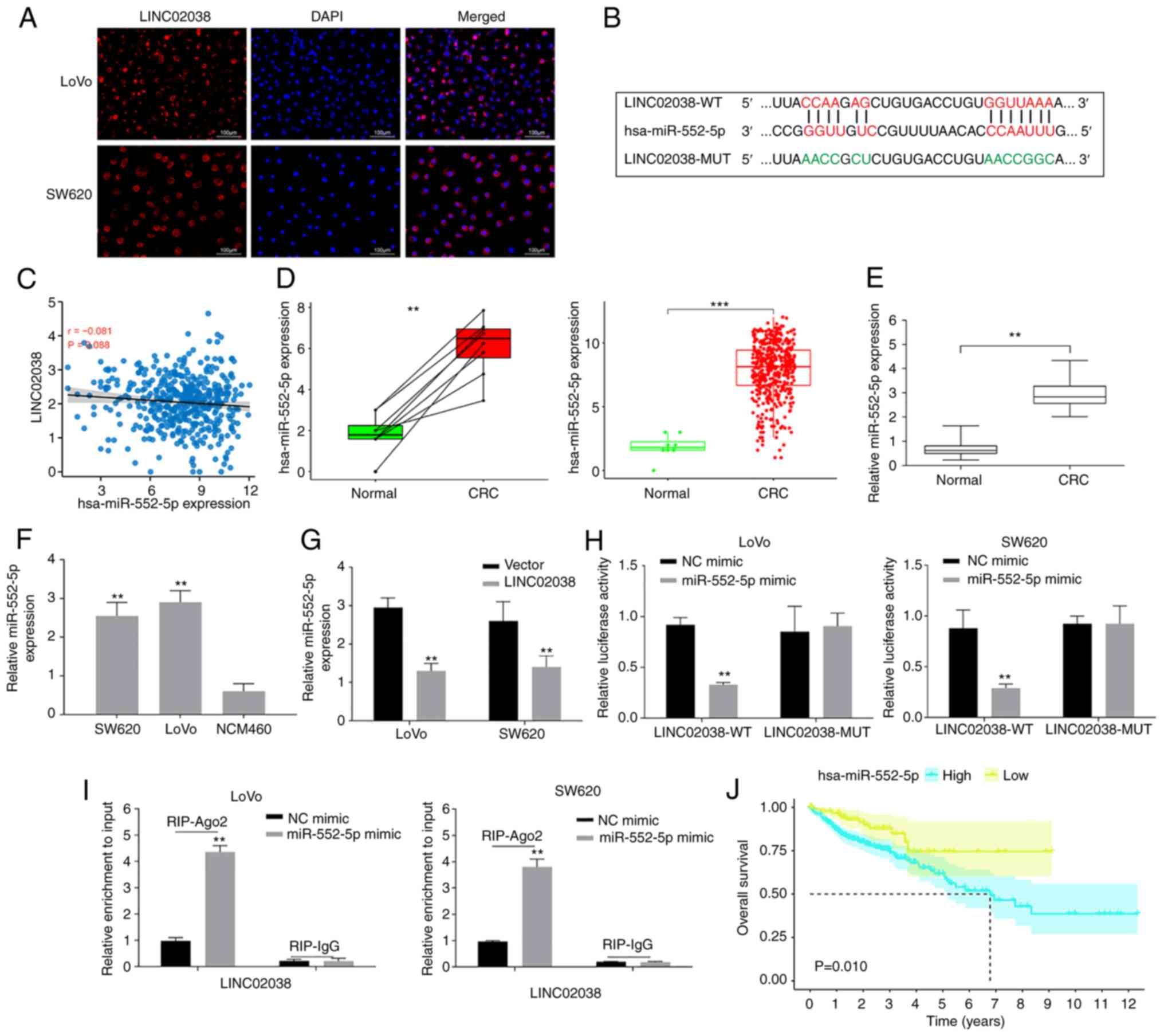 | Figure 3LINC02038 sponges miR-552-5p in the
CRC cells. (A) LINC08038 localized to the cytoplasm (red) and
nucleus (blue) in LoVo and SW620 cells using FISH test. (B) The
predicted binding site between LINC02038 and miR-552-5p. (C) Using
data obtained from TCGA-COAD dataset, it was shown there was a
negative association between LINC02038 and miR-552-5p expression in
CRC tissues. (D) TCGA-COAD database analysis confirmed that
miR-552-5p expression was higher in CRC tissues than in matched and
unpaired normal tissues. (E) RT-qPCR analysis of miR-552-5p
expression in CRC and normal tissues. (F) RT-qPCR detection of
miR-552-5p expression in CRC cells. (G) miR-552-5p expression in
LoVo and SW620 cells transfected with LINC02038 overexpression or
an empty vector was determined using RT-qPCR. (H) The relative
luciferase activity in LoVo and SW620 cells co-transfected with
LINC02038 WT or MUT and miR-552-5p mimics. (I) RIP assays were used
to examine LINC02038 enrichment in LoVo and SW620 cells
co-transfected with LINC0203 and miR-552-5p mimic. (J) The
relationship between miR-552-5p and overall survival in CRC
patients in TCGA-COAD dataset. **P<0.01,
***P<0.001. miR, microRNA; CRC, colorectal cancer;
FISH, Fluorescence in situ hybridization; TCGA, The Cancer
Genome Atlas; COAD, colon adenocarcinoma; RT-qPCR, reverse
transcription quantitative PCR; MUT, mutant; WT, wild-type; Ago2,
argonaute 2; RIP, RNA immunoprecipitation. |
To further confirm the interaction of LINC02038 with
miR-552-5p, dual-luciferase reporter plasmids with WT or MUT
putative binding sites of LINC02038 transcripts were constructed.
The luciferase activity was substantially decreased in cells
co-transfected with LINC02038-WT and miR-552-5p mimics. However, no
statistical changes in luciferase activity were observed when cells
were co-transfected with LINC02038-MUT and miR-552-5p mimics
(Fig. 3H). In the cell cytoplasm,
AGO2, together with GW182 have a role in executing miRNA-mediated
repression. While miRNAs guide AGO2 to target mRNAs, a direct
interaction between AGO2 and GW182 proteins is required for the
assembly of ribonucleoprotein complexes, named RNA-induced
silencing complexes (RISCs) and the recruitment of additional
factors involved in gene silencing, which is ultimately achieved
through the degradation of target mRNAs or translational repression
(23). RNA-binding protein
immunoprecipitation can systematically identify RISC-bound miRNAs
and their target mRNA sequences in mammalian cells (24). LINC02038 was enriched when the Ago2
antibody was used in RIP studies in SW620 and LoVo cells (Fig. 3I), demonstrating that LINC02038
directly binds to miR-552-5p. Additionally, there was an inverse
correlation between LINC02038 expression and OS in the TCGA-COAD
dataset (Fig. 3J).
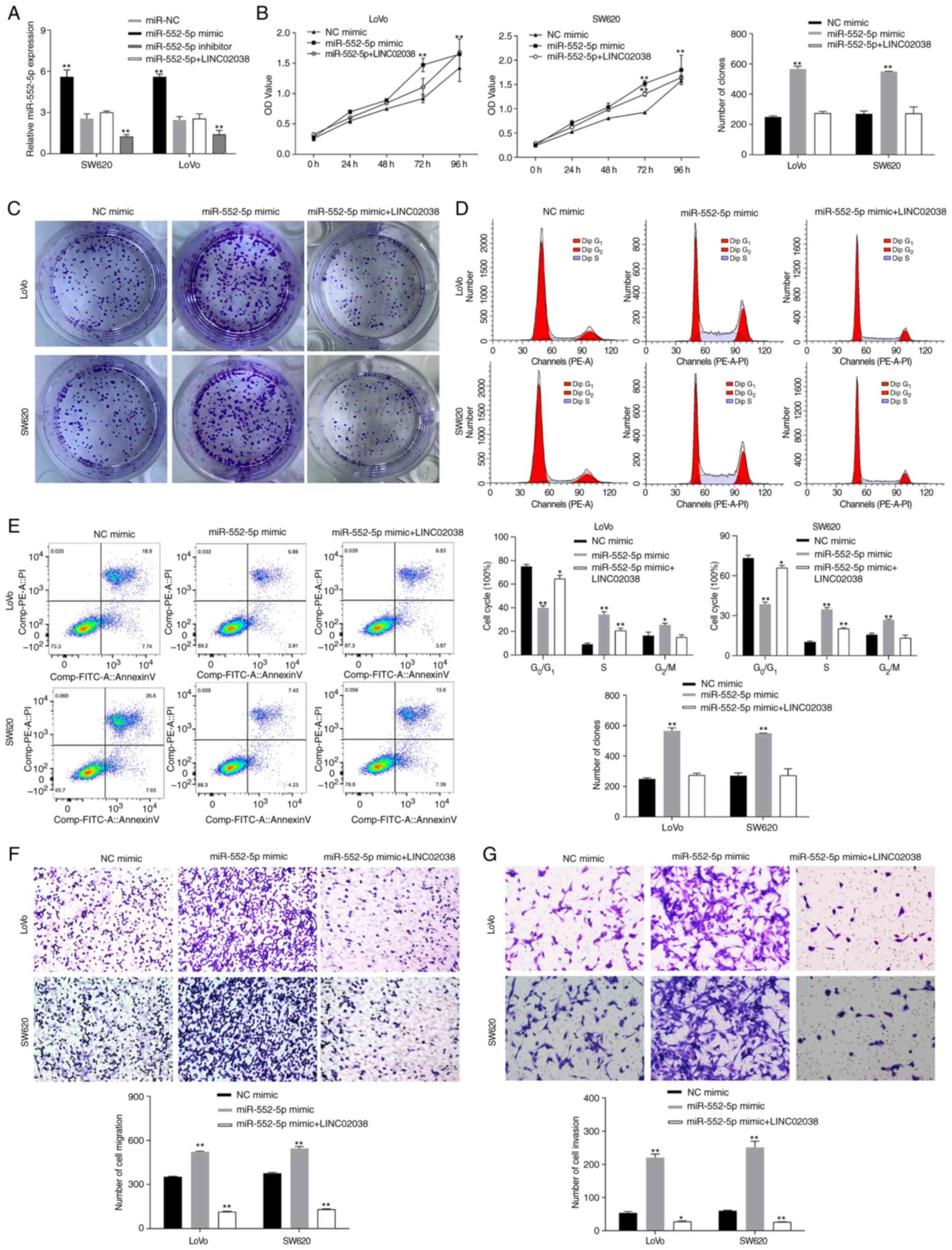
LINC02038 inhibits CRC progression by
targeting miR-552-5p
To study the effect of LINC02038 on miR-552-5p,
miR-552-5p mimic and inhibitor and LINC02038 overexpression vector
were transfected into SW620 and LoVo cells. The results indicated
that overexpression of LINC02038 significantly reduced miR-552-5p
expression levels (Fig. 4A). It
was shown that miR-552-5p promoted proliferation, migration and
invasiveness as well as reduced apoptosis. To evaluate whether
miR-552-5p regulated the molecular functions of LINC02038 in SW620
and LoVo cells, rescue experiments were conducted. The results
showed that transfection of LINC02038 overexpression vector
reversed the effect of miR-552-5p mimics on growth (Fig. 4B and C), migration (Fig. 4F) and invasiveness (Fig. 4G) and the inhibition of cell
apoptosis (Fig. 4E). Cell cycle
analysis showed that the proportion of SW620 and LoVo cells in the
S phase of the cycle was increased after miR-552-5p mimic
transfection, whereas LINC02038 led to an arrest of cell in the
G1 phase (Fig. 4D).
FAM172A is a direct target gene of
miR-552-5p
FAM172A binding to miR-552-5p was identified using
TargetScan and PicTar (Fig. 5A).
Thus, FAM172A expression was studied in TCGA datasets. FAM172A in
CRC tissues was lower than in the non-tumor tissues (Fig. 5B and C). Furthermore, RT-qPCR and
western blot analyses indicated that FAM172A was downregulated
compared to the normal colorectal cell line (Fig. 5D) and tissues (Fig. 5E). In addition, luciferase reporter
assay showed a substantial decrease in luciferase activity
following co-transfection with the FAM172A-WT and miR-552-5p
mimics, while there was no substantial change in cells
co-transfected with FAM172A-MUT and miR-552-5p mimics (Fig. 5F). RIP studies were performed using
the SW620 and LoVo cells. The results revealed that FAM172A
co-precipitated miR-552-5p (Fig.
5G). In addition, the GEPIA database indicated that low FAM172A
expression was associated with a lower progression-free survival
and OS (Fig. 5H and I).
 | Figure 5miR-552-5p targets and regulates
FAM172A. (A) Bioinformatics analysis revealed that miR-552-5p
directly targets the 3′-UTR of FAM172A. (B and C) Differential
FAM172A expression in CRC and normal tissues. FAM172A expression in
(D) several CRC cell lines was determined using RT-qPCR and (E) in
CRC tissues by western blotting. (F) The luciferase activity of the
reporter including miR-552-5p mimic and FAM172A WT or MUT was
evaluated. (G) RIP assay analysis showed FAM172A or miR-552-5p
expression pulled down from the lysates of SW620 and LoVo cells
with the anti-Ago2 antibody. Data obtained from TCGA was utilized
to investigate the association between FAM172A expression and the
(H) PFS and (I) overall survival in CRC patients. Data are
presented as the mean ± SD of three repeats.
**P<0.01, ***P<0.001. miR, microRNA;
UTR, untranslated region; CRC, colorectal cancer; RT-qPCR, reverse
transcription quantitative PCR; WT, wild-type; MUT, mutant; RIP,
RNA immunoprecipitation; TCGA, The Cancer Genome Atlas; PFS,
progression free survival; NC, negative control. |
FAM172A silencing counteracts the
inhibitory effects of LINC02038 overexpression in CRC
To ascertain whether the biological functions of
LINC02038 would change if FAM172A was silenced, a FAM172A knockdown
vector was transfected into CRC cells. The findings revealed that
FAM172A expression was reduced in SW620 and LoVo cells transfected
with siFAM172A (Fig. 6A).
Additionally, the protein and mRNA expression levels of FAM172A
were significantly suppressed in the si-FAM172A group, but these
effects were reversed after co-transfection of LINC02038
overexpression and si-FAM172A (Fig. 6B
and C). Downregulation of FAM172A increased proliferation
(Fig. 6D and E), migration and
invasiveness (Fig. 6H and I) and
inhibited apoptosis (Fig. 6G) of
the SW620 and LoVo cells, but these effects were abolished by
overexpression of LINC02038. Cell cycle distribution analysis
revealed that in SW620 and LoVo cells there was an increased
proportion of cells that had accumulated in the S phase of the cell
cycle following siFAM172A transfection. However, following
co-transfection of siFAM172A and LINC02038 overexpression in the
CRC cells, G1 phase arrest was observed (Fig. 6F). Furthermore, p-Akt and PI3K
protein expression levels were increased after transfection of
siFAM172A, whereas this was reversed in the CRC cells
co-transfected with LINC02038 overexpression and siFAM172A
(Fig. 6J).
Effect of LINC02038 on tumor growth in
vivo
In a xenograft model of tumor growth, LoVo cells
transfected with LINC02038 overexpression or an NC vector were
subcutaneously implanted into nude mice. Mice were euthanized after
4 weeks of injection and tumors were retrieved. Compared to the
control group, the tumor volume and weight were lower in the
LINC02038 overexpression group (Fig.
7A-C). Moreover, immunohistochemical staining of tumor
specimens showed that Ki-67 expression was significantly
downregulated but FAM172A expression was notably upregulated in the
LINC02038 overexpression group compared with the control group
(Fig. 7D). In vivo,
LINC02038 effectively inhibited tumor growth.
METTL3-mediated m6A
modification is associated with LINC02038 downregulation through a
YTHDF2-dependent mechanism in CRC
To further verify the regulatory mode of LINC02038,
SRAMP (http://www.cuilab.cn/sramp/) analysis
was utilized to predict m6A modification sites. It was
discovered that LINC02038 contained an abundance of m6A
modification sites, indicating that it was particularly susceptible
to m6A methylation (Fig.
8A). Moreover, the MeRIP results revealed that LINC02038 has a
number of fragments modified by RNA methylation (Fig. 8B). In addition, METTL3 was
expression was downregulated in LoVo and SW620 cells using two
shRNAs targeting METTL3 (shMETTL3-1 and shMETTL3-2), which markedly
attenuated the mRNA and protein expression of levels of METTL3
(Fig. 8C). According to the
bioinformatics analysis using TCGA, METTL3 expression was inversely
correlated with LINC02038 expression (Fig. 8D). Additionally, LINC02038
expression was significantly upregulated in CRC cells transfected
with shMETTL3 (Fig. 8E).
MeRIP-qPCR results suggested that the m6A abundance in
the LINC02038 mRNA was notably reduced following METTL3 knockdown
(Fig. 8F). Subsequently, the
findings of the RIP assays revealed that METTL3 and YTHDF2 could
enrich LINC02038 (Fig. 8G).
RT-qPCR results demonstrated that YTHDF2 was successfully knocked
down using two shRNA targeting YTHDF2 (shYTHDF2-1 and shYTHDF2-2)
in LoVo and SW620 cells, which markedly weakened YTHDF2 mRNA and
protein levels (Fig. 8H);
LINC02038 expression was upregulated in LoVo and SW620 cells
transfected with shYTHDF2 (Fig.
8I) and knockdown of YTHDF2 reduced the degradation of
LINC02038 (Fig. 8J).
 | Figure 8METTL3 knockdown reduces the
stability of LINC02038 via an m6A-YTHDF2-dependent
mechanism. (A) SRAMP was used to predict the possible
m6A modification locations of LINC02038. (B) The
enrichment of m6A-modified LINC02038 was measured using
a MeRIP-qPCR assay. (C) Using RT-qPCR and western blotting, METTL3
expression was determined in CRC cells following METTL3 knockdown.
(D) Correlation analysis of METTL3 and LINC02038 mRNA on data
obtained from TCGA. (E) LINC02038 expression in METTL3-deficient
LoVo and SW620 cells was detected using RT-qPCR. (F)
shMETTL3-mediated LINC02038 m6A modifications were
demonstrated in LoVo and SW620 cells. (G) The interaction between
METTL3, METTL14, FTO and YTHDF2. (H) YTHDF2 expression of LoVo and
SW620 cells transfected with shYTHDF2 were detected using RT-qPCR
and western blotting. (I) LINC02038 expression was determined using
RT-qPCR in YTHDF2-deficient LoVo and SW620 cells. (J) The
expression of LINC02038 was detected following treatment with
Actinomycin D (5 mg/ml) in SW620 cells using RT-qPCR at the
indicated time points. Data are presented as the mean ± SD of three
repeats. *P<0.05, **P<0.01.
m6A, N6-methyladenosine; MeRIP, methylated RNA
immunoprecipitation; qPCR, quantitative PCR; RT-qPCR, reverse
transcription qPCR; TCGA, The Cancer Genome Atlas; shRNA, short
hairpin RNA. |
Discussion
The aim of the present study was to determine the
clinical importance of LINC02038 and its role in the progression of
CRC, as well as to explore its potential molecular mechanisms. TCGA
and GEO datasets were analyzed and the results were validated in
CRC samples. It was discovered that LINC02038 played a significant
role in CRC. The findings showed that LINC02038 was notably
downregulated in CRC tissues and cell lines. Additionally, it was
found that low expression of LINC02038 was associated with poor
prognosis in CRC, suggesting that it could have been a potential
prognostic target for this disease.
The present study examined whether LINC02038 was a
unique tumor-suppressing lncRNA that regulated the growth and
aggressiveness of CRC. The results of gain-of-function assays
showed that LINC02038 suppressed CRC cell proliferation, migration
and invasiveness while promoting apoptosis. In vivo
experiments revealed that LINC02038 inhibited CRC tumor growth,
demonstrating its tumor suppressor function in CRC. Studies have
shown the importance of lncRNAs in carcinogenesis and antitumor
behavior, with dysregulation of lncRNAs being closely associated
with tumor formation (25,26). CRC-specific diagnosis, prognosis
and therapies may target specific lncRNAs (27). Potentially viable treatments for
advanced CRC may involve regulating lncRNAs with specific
expression patterns. Therefore, understanding the function and
regulation processes of lncRNAs is critical. LINC02038 is a 2,434
nucleotide-long intergenic non-protein coding RNA involved in CRC
prognosis (14). However, its
ability to regulate cellular processes through interactions with
other cellular components remains to be elucidated. To understand
the function of an lncRNA, knowledge about its subcellular
distribution within the cell is required (28). The present study found that
LINC02038 was mainly located in the cytoplasm of CRC cells.
Theoretically, LINC02038 may affect fundamental biological
processes such as methylation and post-transcriptional
regulation.
The majority of cytoplasmic lncRNAs act as ceRNAs to
modulate downstream mRNA expression by competitively suppressing
miRNA expression through the presence of miRNA response elements
(29). For example, LINC01133 can
elicit biological behaviors as a ceRNA by interacting with the
target sequence of miR-106a-3p, thereby expanding the roles of APC
mRNA targets (30). Xu et
al (31) reported that lncRNA
SNHG6 was elevated in colorectal cancer (CRC) and enhanced the
invasiveness of CRC cells by sponging miR-26a/b and miR-214.
Dual-luciferase reporter and RIP assays indicated that LINC02038
absorbed miR-552-5p. Previous studies have shown that miR-552-5p is
an oncogene in various types of human cancer and promotes cell
proliferation and metastasis (32,33).
Mechanistically, LINC02038 functions as a ceRNA for miR-552-5p,
inhibiting CRC cell proliferation, migration and invasion while
promoting apoptosis. These findings may offer novel therapeutic
strategies for CRC. The miRNA target gene is essential for the
function of lncRNAs as ceRNAs. Using TargetScan and PicTar online
databases, FAM172A was identified as a potential target of
miR-552-5p. Luciferase reporter and RIP assay results showed that
FAM172A was a downstream target gene of miR-552-5p. Previous
studies have indicated that FAM172A may play an important role in
inhibiting human cancer cell growth by regulating different
signaling pathways. For example, FAM172A inhibited CRC
proliferation and invasion by being regulated at the
post-transcriptional level by miR-27a via NF-κB pathways (34). Chen et al (35) reported that FAM172A was
downregulated in human pancreatic cancer tissues and inhibited
epithelial-mesenchymal transition (EMT) in pancreatic cancer cells
via MAPK/ERK and PI3K/Akt signaling.
In the present study, overexpression of LINC02038
partly rescued the effects of FAM172A silencing on malignant
phenotypes of CRC cells, suggesting that FAM172A is essential for
the biological effects mediated by the LINC02038/miR-552-5p axis in
CRC cells. Emerging evidence reveals that RNA methylation plays a
crucial role in the dysfunction of lncRNAs (36). The most prevalent type of RNA
modification is m6A modification (37,38).
The present study aimed to determine whether m6A
modification sites existed in the LINC02038 sequence. Notably,
several sites with high confidence for m6A modification
were found on LINC02038. Therefore, it was hypothesized that
abnormal expression of LINC02038 may be linked to m6A
modification. RNA pull-down experiments showed that LINC02038
specifically binds to METTL3 and YTHDF2. METTL3, an m6A
writer RNA methyltransferase, was upregulated in CRC and regulated
mRNA stability (39). Knockdown of
METTL3 enhanced LINC02038 expression in this study, rendering
LINC02038 incapable of m6A modification due to METTL3
deficiency. Furthermore, the m6A reader YTHDF2 has been
implicated in regulating methylated mRNA stability and tumor
progression in CRC (40,41). The present study found that YTHDF2
directly bound to specific m6A sites on LINC02038 and
regulated it in an m6A-dependent manner. The results
confirmed elevated expression of YTHDF2 in CRC and its regulatory
effect on LINC02038 instability to promote CRC.
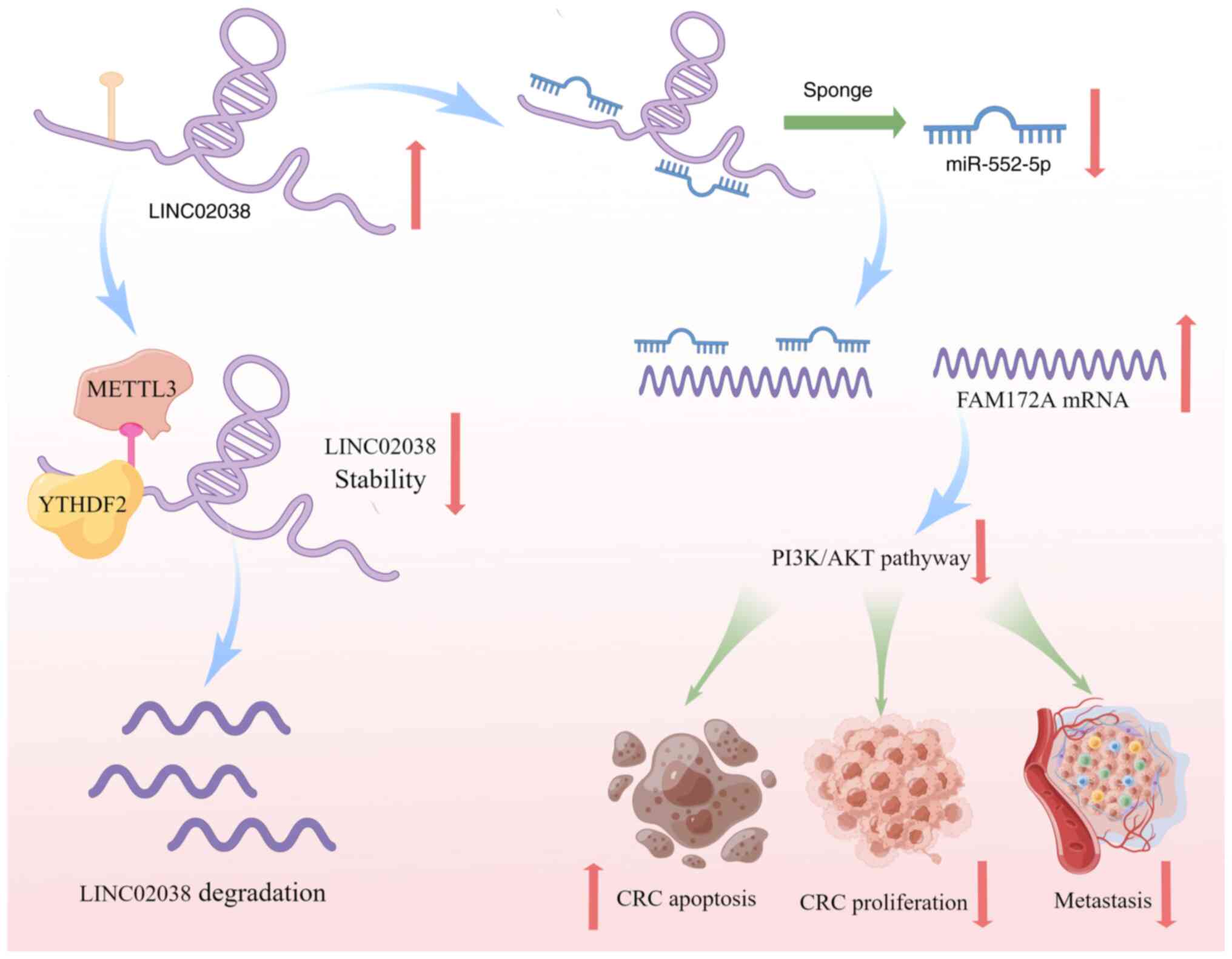
In summary, the present study demonstrated that
LINC02038 is a novel tumor suppressor in CRC. Decreased levels of
LINC02038 in CRC patients were associated with increased tumor
metastasis and worse survival due to m6A modification.
LINC02038 sponged miR-552-5p to impede CRC development, which was
mediated by FAM172A and PI3K/Akt signaling in CRC cells. These
results shed new light on the mechanisms underlying the function of
the LINC02038 in CRC. Therefore, m6A modification of the
LINC02038/miR-552-5p/FAM172A axis may represent a promising
therapeutic approach and prognostic biomarker for CRC (Fig. 9).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL, ZZ and CY conceived and designed the present
study. WL, ZZ and JL performed the experiments. XL and BH acquired
and collected tissue samples and the clinical data. KQ and ZM
performed the in vivo experiments. JD and WL analyzed and
interpreted the data. WL and ZZ wrote the manuscript. CY edited the
manuscript. WL and JD confirm the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study involving human tissues was
approved by the Ethics Committee of the First People's Hospital of
Foshan, Guangdong (approval no. AF-SOP-18-1.6-2). All patients
provided written informed consent prior to their inclusion within
the study. All animal experiments were approved by the Committee of
the Ethics of Animal Experiments of Southern Medical University
(approval no. 2032101).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was funded by the Natural Science Foundation
of Guangdong Province (grant no. 2018A0303130312), the clinical
characteristic technology project of Guangzhou (grant no.
2023C-TS45).
References
|
1
|
Kanth P and Inadomi JM: Screening and
prevention of colorectal cancer. BMJ. 374:n18552021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dekker E, Tanis PJ, Vleugels JLA, Kasi PM
and Wallace MB: Colorectal cancer. Lancet. 394:1467–1480. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Biller LH and Schrag D: Diagnosis and
treatment of metastatic colorectal cancer: A review. JAMA.
325:669–685. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li N, Lu B, Luo C, Cai J, Lu M, Zhang Y,
Chen H and Dai M: Incidence, mortality, survival, risk factor and
screening of colorectal cancer: A comparison among China, Europe,
and northern America. Cancer Lett. 522:255–268. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou L, Zhu Y, Sun D and Zhang Q: Emerging
roles of long non-coding RNAs in the tumor microenvironment. Int J
Biol Sci. 16:2094–2103. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chi Y, Wang D, Wang J, Yu W and Yang J:
Long non-coding RNA in the pathogenesis of cancers. Cells.
8:10152019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao Z, Sun W, Guo Z, Zhang J, Yu H and
Liu B: Mechanisms of lncRNA/microRNA interactions in angiogenesis.
Life Sci. 254:1169002020. View Article : Google Scholar
|
|
9
|
Winkle M, El-Daly SM, Fabbri M and Calin
GA: Noncoding RNA therapeutics - challenges and potential
solutions. Nat Rev Drug Discov. 20:629–651. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chan JJ and Tay Y: Noncoding RNA: RNA
regulatory networks in cancer. Int J Mol Sci. 19:13102018.
View Article : Google Scholar
|
|
11
|
Tang J, Yan T, Bao Y, Shen C, Yu C, Zhu X,
Tian X, Guo F, Liang Q, Liu Q, et al: LncRNA GLCC1 promotes
colorectal carcinogenesis and glucose metabolism by stabilizing
c-Myc. Nat Commun. 10:34992019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin X, Zhuang S, Chen X, Du J, Zhong L,
Ding J, Wang L, Yi J, Hu G, Tang G, et al: lncRNA ITGB8-AS1
functions as a ceRNA to promote colorectal cancer growth and
migration through integrin-mediated focal adhesion signaling. Mol
Ther. 30:688–702. 2022. View Article : Google Scholar :
|
|
13
|
Hu XT, Xing W, Zhao RS, Tan Y, Wu XF, Ao
LQ, Li Z, Yao MW, Yuan M, Guo W, et al: HDAC2 inhibits EMT-mediated
cancer metastasis by downregulating the long noncoding RNA H19 in
colorectal cancer. J Exp Clin Cancer Res. 39:2702020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen J, Song Y, Li M, Zhang Y, Lin T, Sun
J, Wang D, Liu Y, Guo J and Yu W: Comprehensive analysis of ceRNA
networks reveals prognostic lncRNAs related to immune infiltration
in colorectal cancer. BMC Cancer. 21:2552021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karreth FA and Pandolfi PP: ceRNA
cross-talk in cancer: When ce-bling rivalries go awry. Cancer
Discov. 3:1113–1121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu AM, He CJ, Tuerxun Z and Anikezi A:
FAM172A affects cell proliferation and apoptosis not by targeting
β-tubulin in HepG2 cells. Transl Cancer Res. 9:5637–5644. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Narayanankutty A: PI3K/Akt/mTOR pathway as
a therapeutic target for colorectal cancer: A review of preclinical
and clinical evidence. Curr Drug Targets. 20:1217–1226. 2019.
View Article : Google Scholar
|
|
18
|
Vu LP, Cheng Y and Kharas MG: The biology
of m6A RNA methylation in normal and malignant
hematopoiesis. Cancer Discov. 9:25–33. 2019. View Article : Google Scholar
|
|
19
|
Ianniello Z, Paiardini A and Fatica A:
N6-Methyladenosine (m6A): A promising new
molecular target in acute myeloid leukemia. Front Oncol. 9:2512019.
View Article : Google Scholar
|
|
20
|
Xiang S, Liang X, Yin S, Liu J and Xiang
Z: N6-methyladenosine methyltransferase METTL3 promotes colorectal
cancer cell proliferation through enhancing MYC expression. Am J
Transl Res. 12:1789–1806. 2020.PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Li HB, Tong J, Zhu S, Batista PJ, Duffy
EE, Zhao J, Bailis W, Cao G, Kroehling L, Chen Y, et al:
m6A mRNA methylation controls T cell homeostasis by
targeting the IL-7/STAT5/SOCS pathways. Nature. 548:338–342. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pfaff J, Hennig J, Herzog F, Aebersold R,
Sattler M, Niessing D and Meister G: Structural features of
Argonaute-GW182 protein interactions. Proc Natl Acad Sci USA.
110:E3770–E3779. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hicks JA, Li L, Matsui M, Chu Y, Volkov O,
Johnson KC and Corey DR: Human GW182 paralogs are the central
organizers for RNA-mediated control of transcription. Cell Rep.
20:1543–1552. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Smolarz B, Zadrożna-Nowak A and Romanowicz
H: The Role of lncRNA in the development of tumors, including
breast cancer. Int J Mol Sci. 22:84272021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ghafouri-Fard S, Abak A, Tondro Anamag F,
Shoorei H, Majidpoor J and Taheri M: The emerging role of
non-coding RNAs in the regulation of PI3K/AKT pathway in the
carcinogenesis process. Biomed Pharmacother. 137:1112792021.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He Q, Long J, Yin Y, Li Y, Lei X, Li Z and
Zhu W: Emerging roles of lncRNAs in the formation and progression
of colorectal cancer. Front Oncol. 9:15422019. View Article : Google Scholar
|
|
28
|
Statello L, Guo CJ, Chen LL and Huarte M:
Gene regulation by long non-coding RNAs and its biological
functions. Nat Rev Mol Cell Biol. 22:96–118. 2021. View Article : Google Scholar
|
|
29
|
Smillie CL, Sirey T and Ponting CP:
Complexities of post-transcriptional regulation and the modeling of
ceRNA crosstalk. Crit Rev Biochem Mol Biol. 53:231–245. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang XZ, Cheng TT, He QJ, Lei ZY, Chi J,
Tang Z, Liao QX, Zhang H, Zeng LS and Cui SZ: LINC01133 as ceRNA
inhibits gastric cancer progression by sponging miR-106a-3p to
regulate APC expression and the Wnt/β-catenin pathway. Mol Cancer.
17:1262018. View Article : Google Scholar
|
|
31
|
Xu M, Chen X, Lin K, Zeng K, Liu X, Xu X,
Pan B, Xu T, Sun L, He B, et al: lncRNA SNHG6 regulates EZH2
expression by sponging miR-26a/b and miR-214 in colorectal cancer.
J Hematol Oncol. 12:32019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen T, Lei S, Zeng Z, Zhang J, Xue Y, Sun
Y, Lan J, Xu S, Mao D and Guo B: Linc00261 inhibits metastasis and
the WNT signaling pathway of pancreatic cancer by regulating a
miR-552-5p/FOXO3 axis. Oncol Rep. 43:930–942. 2020.PubMed/NCBI
|
|
33
|
Cai W, Xu Y, Yin J, Zuo W and Su Z:
miR-552-5p facilitates osteosarcoma cell proliferation and
metastasis by targeting WIF1. Exp Ther Med. 17:3781–3788.
2019.PubMed/NCBI
|
|
34
|
Liu W, Qian K, Wei X, Deng H, Zhao B, Chen
Q, Zhang J and Liu H: miR-27a promotes proliferation, migration,
and invasion of colorectal cancer by targeting FAM172A and acts as
a diagnostic and prognostic biomarker. Oncol Rep. 37:3554–3564.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen Y, Liu P, Shen D, Liu H, Xu L, Wang
J, Shen D, Sun H and Wu H: FAM172A inhibits EMT in pancreatic
cancer via ERK-MAPK signaling. Biol Open. 9:bio0484622020.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu HT, Zou YX, Zhu WJ, Sen-Liu, Zhang GH,
Ma RR, Guo XY and Gao P: lncRNA THAP7-AS1, transcriptionally
activated by SP1 and post-transcriptionally stabilized by
METTL3-mediated m6A modification, exerts oncogenic properties by
improving CUL4B entry into the nucleus. Cell Death Differ.
29:627–641. 2022. View Article : Google Scholar :
|
|
37
|
Deng X, Su R, Weng H, Huang H, Li Z and
Chen J: RNA N6-methyladenosine modification in cancers:
Current status and perspectives. Cell Res. 28:507–517. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Roundtree IA, Evans ME, Pan T and He C:
Dynamic RNA modifications in gene expression regulation. Cell.
169:1187–1200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peng W, Li J, Chen R, Gu Q, Yang P, Qian
W, Ji D, Wang Q, Zhang Z, Tang J and Sun Y: Upregulated METTL3
promotes metastasis of colorectal cancer via miR-1246/SPRED2/MAPK
signaling pathway. J Exp Clin Cancer Res. 38:3932019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhou D, Tang W, Xu Y, Xu Y, Xu B, Fu S,
Wang Y, Chen F, Chen Y, Han Y and Wang G: METTL3/YTHDF2 m6A axis
accelerates colorectal carcinogenesis through epigenetically
suppressing YPEL5. Mol Oncol. 15:2172–2184. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu W, Liu C, You J, Chen Z, Qian C, Lin
W, Yu L, Ye L, Zhao L and Zhou R: Pan-cancer analysis identifies
YTHDF2 as an immunotherapeutic and prognostic biomarker. Front Cell
Dev Biol. 10:9542142022. View Article : Google Scholar : PubMed/NCBI
|