1. Introduction
With the development of medical science and
technology, the diagnostic efficiency and treatment of cancer has
markedly improved. There are three main types of systemic cancer
treatments: Chemotherapy, targeted therapy and immunotherapy
(1,2). There are notable individual
differences in the therapeutic effects of targeted therapy and
immunotherapy. Chemotherapy, which kills tumor cells
indiscriminately, is considered an irreplaceable cancer treatment.
Unfortunately, tumor multidrug resistance (MDR), resulting in the
lack of tumor sensitivity to a number of chemotherapy drugs, is an
obstacle limiting chemotherapy application (3). The mechanisms underlying MDR include
ATP-binding cassette (ABC) transporter overexpression, autophagy,
DNA damage repair, cancer stem cells, genetic mutations and DNA
methylation (4-8). Overexpressed ABC transporters, which
are the main cause of chemotherapy MDR, use the energy generated by
ATP hydrolysis to pump chemotherapy drugs out of the cell (9). MDR-related ABC transporters include
P-glycoprotein (P-gp), breast cancer resistance protein (BCRP),
multidrug resistance-associated protein (MRP) and lung
resistance-associated protein (LRP). P-gp is the main cause of MDR
in tumors according to numerous studies (10,11).
P-gp was the first drug resistance-related protein
to be discovered. In 1976, P-gp was identified in drug-resistant
Chinese hamster ovary cells as a membrane glycoprotein with a
molecular weight of ~170 kDa (12). P-gp, encoded by the ABCB1 gene, is
the most studied transmembrane transporter in the ABC transporter
family. The structure of P-gp consists of two nucleotide-binding
domains (NBDs) and two transmembrane domains (TMDs) (13). NBDs are located in the cytoplasm
and transfer energy through membranes to transport substrates. TMDs
consist of six transmembrane α-helical structures that provide
specificity for substrates. Chemotherapy drugs that act as P-gp
substrates are pumped out of tumor cells (14,15).
P-gp has been reported to be overexpressed in
various tumors, such as osteosarcoma, hepatocellular carcinoma
(HCC) (16), breast cancer
(17), gastric cancer (18), lung cancer (19,20)
and bowel cancer (21), resulting
in chemotherapy resistance. P-gp overexpression reduces
intracellular drug concentrations and cytotoxicity by using its
efflux pump function to expel cisplatin (20), paclitaxel (22), 5-fluorouracil (5-Fu) (23), doxorubicin (DOX) (24) and other chemotherapeutic drugs out
of cells, which results in inadequate proliferation-inhibiting
effects of chemotherapy drugs on tumor cells. In view of the common
phenomenon of clinical chemotherapy resistance, a large number of
studies have been carried out on the mechanism of P-gp-mediated
chemotherapy resistance and P-gp inhibitors (25,26).
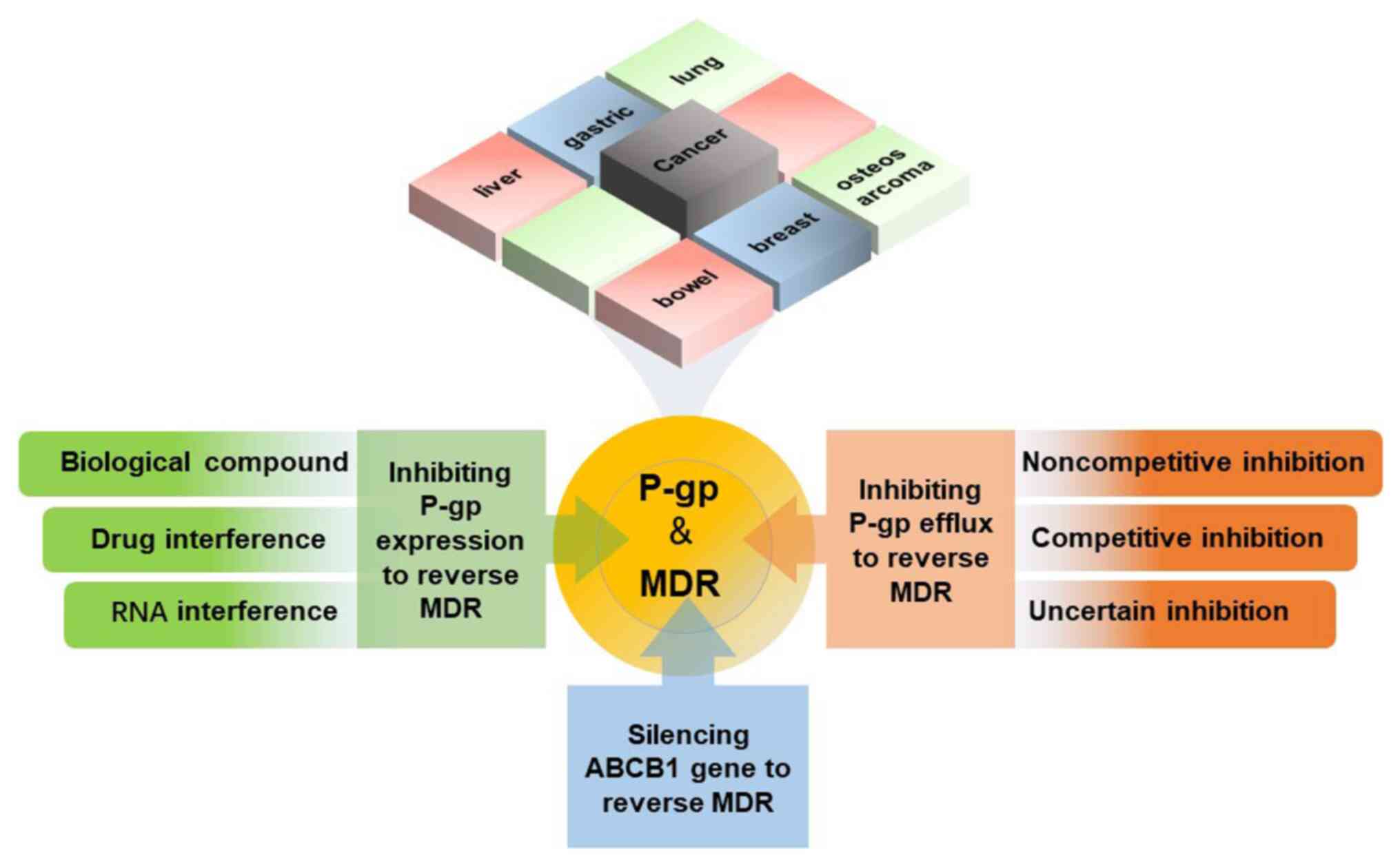
The present review aims to provide an overview of
the overexpression of P-gp in different types of cancer leading to
chemotherapy resistance (Fig. 1).
This review summarizes and elaborates on three current strategies
for overcoming P-gp-mediated chemotherapy resistance (Fig. 2). The first strategy involves
competitive or noncompetitive suppression of the P-gp efflux pump
function. The second strategy involves inhibition of the expression
of P-gp and efflux transporters on the cell membrane that bind to
chemotherapy drugs. The third approach involves knockout of the
ABCB1 gene. The active substances that reverse P-gp-mediated
chemotherapy resistance through these three pathways can also be
classified into groups. They are roughly divided into natural
products, synthetic compounds, endogenous RNAs and biological
compounds. The present review aims to alert researchers to the
important contribution of P-gp overexpression in chemotherapy
resistance and to provide a reference for P-gp-related chemotherapy
resistance research.
2. MDR caused by P-gp in various tumor
tissues
Before the advent of targeted therapy and
immunotherapy, chemotherapy was the most common clinical treatment
for a variety of malignant tumors; therefore, P-gp is a common
mechanism underlying MDR in multiple malignant tumors. In general,
ABC transporters, including P-gp, are expressed more highly in
resistant tumors, which increases the chance of drug efflux. The
high expression of P-gp in liver cancer, lung cancer, breast
cancer, bowel cancer, gastric cancer and osteosarcoma leads to
chemoresistance.
MDR caused by P-gp in liver cancer
Liver cancer is a type of malignant tumor that can
be caused by alcoholism, drug injury and viral infection (27). Before sorafenib, a small-molecule
targeted drug, was approved by the United States Food and Drug
Administration for the treatment of advanced HCC, chemotherapy was
the preferred treatment for unresectable liver cancer (28).
P-gp overexpression-mediated MDR is sometimes
induced by external factors and not the spontaneous behavior of
cancer cells following prolonged exposure to chemotherapy drugs.
HepG2 cells are resistant to DOX in an acute or chronic hypoxic
environment, and have high expression levels of nuclear factor
erythroid-2 related factor 2 (Nrf2) and P-gp. It has been
hypothesized that hypoxia can induce the expression of P-gp by
inducing the expression of Nrf2 (29).
The regulatory mechanism of P-gp is a multimolecular
process. In DOX-resistant liver cancer cells, the levels of NF-κB
and P-gp are relatively high, thus indicating that the expression
of NF-κB in HCC may be closely related to the ABCB1 gene (16). It has also been reported that the
resistance of HCC to epirubicin is associated with the high
expression of tripartite motif protein 25 (TRIM25).
Epirubicin-resistant HCC cells in which TRIM25 was knocked down
have been shown to exhibit increased sensitivity to epirubicin, an
increased apoptosis rate and downregulation of P-gp (30). Furthermore, Bcl-2-associated
transcription factor 1 can promote 5-Fu resistance, and the
expression of P-gp and MRP1 in HCC cells by directly binding to the
promoter region of long non-coding RNA (lncRNA) NEAT1 (31).
MDR caused by P-gp in lung cancer
Lung cancer is one of the most life-threatening
types of cancer (27).
P-gp-mediated cisplatin resistance is prominent in lung cancer.
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) was
first found in non-small cell lung cancer (NSCLC). It has been
reported that upregulated MALAT1 in A549 NSCLC cells promotes P-gp
expression and P-gp-mediated cisplatin resistance by increasing
signal transducer and activator of transcription 3 (STAT3)
phosphorylation levels (32). In
addition, it has been confirmed that the mRNA stability of ABCB1
and STAT3 phosphorylation in lung cancer cells can be enhanced by
sodium butyrate (SB); therefore, it can be inferred that P-gp
expression may be positively mediated by SB-induced expression or
phosphorylation of STAT3 (33).
Chloride channel-3 (ClC-3) promotes ABCB1 and P-gp expression by
activating the NF-κB signaling pathway, and the key point of this
process is the nuclear translocation of NF-κB P65 (19). Furthermore, upregulation of
hypoxia-inducible factor (HIF) in NSCLC may lead to P-gp
overexpression and promote cisplatin resistance (20); this result is similar to the
mechanism of hypoxia-induced P-gp overexpression in liver cancer.
Therefore, it may be suggested that modulating oxygen supply to
tumors could overcome P-gp-mediated MDR.
MDR caused by P-gp in osteosarcoma
Osteosarcoma is a highly malignant tumor with a poor
prognosis; however, chemotherapy before and after surgery can
greatly improve this prognosis (34). Previous studies have confirmed that
P-gp expression is positively associated with MDR in osteosarcoma
(11,35). The expression levels of P-gp in
cisplatin-resistant osteosarcoma cells have been shown to be much
higher than those in nondrug-resistant cells. Furthermore,
following knockdown of P-gp expression, the sensitivity of
osteosarcoma cells to cisplatin was restored (11). Pleiotrophin (PTN), a neurotrophic
growth factor, is linked to DOX resistance caused by P-gp
overexpression. PTN can upregulate ABCB1/P-gp by mediating
ALK/GSK3β/β-catenin signaling (35). LIM domain kinase 1 (LIMK1), a
member of the serine/threonine protein kinase family, can also
contribute to MDR by upregulating the levels of P-gp. This finding
was demonstrated by transfecting MG63/VCR osteosarcoma cells with
LIMK1-small interfering RNA (siRNA), wild-type-LIMK1 or empty
vector; the apoptosis rate of cells transfected with LIMK1-siRNA
was higher (36).
MDR caused by P-gp in breast cancer
P-gp is associated with drug resistance to a variety
of anticancer drugs used in the treatment of breast cancer. At
present, there are two types of breast cancer resistance mechanisms
related to P-gp: Activation of the P-gp efflux pump and enhancement
of P-gp expression.
The interaction of programmed cell death ligand 1
(PD-L1) and programmed cell death-1 (PD-1) on T cells can enhance
chemotherapeutic resistance in breast cancer cells by inhibiting
immune responses (37). Therefore,
the regulation of MDR by the tumor immune microenvironment was
investigated. From the perspective of tumor autoimmune regulation,
P-gp-mediated MDR may be an immune response to exogenous
chemotherapy drugs. Activation of multiple signaling pathways
promotes P-gp expression and MDR in breast cancer. Notably,
activating PI3K/AKT and MAPK/ERK pathways can increase the
expression of ABCB1/P-gp and enhance DOX resistance (37). ABCB1 transcription can also be
directly promoted by WW domain-binding protein 2 through binding to
ERα, which is a potential mechanism of P-gp-related MDR (38). Nkx-2.5, a transcription factor, can
bind to the +4 to +10 binding site of the upstream promoter of
ABCB1 in breast cancer cells, thereby transactivating ABCB1 for
transcription. Eventually, P-gp is upregulated, thus leading to
breast cancer resistance (39). In
addition to studies regarding the increased expression of P-gp,
β1-integrin binding to collagen type 1 has been reported to induce
chemoresistance of breast cancer cells by activating ABC efflux
transporters (40).
Furthermore, resistance to trastuzumab emtansine,
which is an antibody-conjugated drug used to treat metastatic
breast cancer, is related to the upregulation of P-gp (41).
MDR caused by P-gp in gastric cancer
A number of studies on gastric cancer resistance
have been conducted because of its high incidence (42,43).
Notably, lncRNAs are closely related to the occurrence of
P-gp-mediated gastric cancer resistance. The high expression of
lncRNAs differentiation antagonizing non-protein coding RNA (DANCR)
and gastric carcinoma proliferation enhancing transcript 1 (GHET1)
in gastric cancer cells can increase the occurrence of drug
resistance by upregulating the expression of ABCB1, which has been
confirmed by silencing DANCR and GHET1 (44,45).
Zinc finger protein 139 inhibits the transcriptional
activity of the microRNA (miRNA/miR)-185 promoter to downregulate
miR-185, which also reduces the expression of ABCB1 (46). lncRNAs and miRNAs are non-coding
RNAs transcribed from the genome that regulate epigenetics and
signal transduction in vivo. Therefore, the involvement of
lncRNAs and miRNAs in P-gp-mediated MDR is not surprising.
MDR caused by P-gp in bowel cancer
Chemotherapeutic resistance is also a vital obstacle
in the clinical treatment of bowel cancer. Activation of the ABCB1
promoter region promotes P-gp expression and MDR. Visfatin
regulates ABCB1 transcription by regulating ABCB1 promoter activity
to mediate the DOX resistance of colorectal cancer cells. When
transfected with visfatin-specific siRNA, the expression of ABCB1
in colorectal cancer cells was significantly reduced and the
sensitivity to DOX was restored (47). ABCB1 mRNA and protein expression
can be stimulated by forkhead box O3 through binding to the
promoter region of ABCB1, and the proliferation of colon cancer
cells and MDR can be induced by the overexpression of forkhead box
O3 (48). In addition, IL-8 is
significantly upregulated in DOX-resistant colorectal cancer cells.
Notably, NF-κB (P65) may be activated by IL-8 via upregulation of
IKKβ phosphorylation, thus resulting in overexpression of P-gp
(49). P-gp expression can also be
promoted by SET and MYND domain-containing protein 2 through the
MEK/ERK/AP-1 signaling pathway, leading to the resistance of colon
cancer to oxaliplatin (21). A
relationship also exists between ABCB1 and adhesion G
protein-coupled receptor G1 (GPR56), a type of adhesion G-protein
coupled receptor. GPR56 can enhance drug resistance through
upregulation of ABCB1 levels via a RhoA-mediated signaling
mechanism in primary colon tumors (50).
3. Signaling pathways involved in P-gp
mediated MDR
P-gp exerting its efflux pump function is the final
step leading to chemotherapy MDR. Considering the large number of
molecules in the body, P-gp-mediated MDR is likely to be regulated
by a variety of signaling pathways (Fig. 3) P-gp overexpression mediates MDR
in various types of cancer and the signaling pathways involved are
similar. These pathways may seem intricate, but they all contain
the same key molecule, NF-κB, which acts as a hub. Simultaneous
overexpression of NF-κB and P-gp in drug-resistant tumors has
prompted studies on the regulation of P-gp by NF-κB (51,52).
The positive regulatory effect of NF-κB on P-gp has been verified
by comprehensive analysis of the signaling pathway of P-gp
inhibitors (16). It has been
reported that NF-κB regulates P-gp through the downstream STAT3
signaling pathway and the regulation of NF-κB on P-gp is influenced
by a number of upstream molecules in the body.
N-acetyl-glucosaminyl transferase III can inhibit DOX resistance by
negatively regulating P-gp expression through the TNFR2-NF/κB
signaling pathway (53).
Furthermore, melatonin inactivates the NF-κB pathway and
downregulates P-gp, thus increasing sensitivity to epirubicin
chemotherapy (54). By contrast,
IL-8 positively regulates P-gp expression in DOX-resistant cells by
activating the IKKβ/NF-κB signaling pathway (49). ClC-3 also increases ABCB1 mRNA and
P-gp levels by activating the NF-κB signaling pathway (19).
Dysregulation of the PI3K/Akt signaling pathway,
which is commonly seen in cancer, and regulates metabolism,
proliferation, cell survival, growth and angiogenesis, has also
been reported to serve an important role in regulating P-gp
expression and MDR (55,56). Notably, activation of the PI3K/Akt
pathway can upregulate P-gp in leukemia (56), ovarian cancer (51), liver cancer (57) and lung cancer (55). Compared with PI3K alone, the
CDK6-PI3K signal axis synergistically regulates ABCB1 expression
more strongly (25). Therefore,
the idea of multipathway and multitarget synergistic inhibition of
P-gp expression and efficient reversal of MDR was investigated.
MEK/ERK is an important downstream signaling pathway of PI3K/Akt.
Notably, the MEK/ERK signaling pathway positively regulates P-gp
expression, and mediates resistance to DOX, paclitaxel,
oxaliplatin, colchicine and vincristine (21,55).
Wnt/β-catenin, which regulates the high expression of P-gp in colon
cancer, HCC and leukaemia, is another major signaling pathway
associated with P-gp-mediated MDR. Moreover, the
MALAT1/STAT3/fucosyltransferase 4 (FUT4) (58), JNK/c-Jun/AP-1 (59) and PTN/β-catenin axes (60) are involved in regulating P-gp
expression and mediating MDR.
In conclusion, the expression of P-gp is the result
of the combined action of several signaling pathways. The discovery
of these signaling pathways and regulatory molecules could clarify
the role of P-gp and aid in identification of its inhibitors. These
signaling pathways and regulatory molecules that promote P-gp
expression and MDR can be used as potential targets to reverse
chemotherapy resistance; however, no related inhibitors are
currently used in clinical chemotherapy. In basic research, direct
or indirect inhibition of a target can appropriately alleviate
numerous drug resistance phenomena. The future application of these
targeted inhibitors in clinical treatment may alleviate
chemotherapy resistance and restore the therapeutic effect of
chemotherapy drugs.
4. Reversing MDR by inhibiting P-gp
Inhibition of the efflux pump, which can be divided
into noncompetitive inhibition and competitive inhibition according
to the mechanism, is a direct way to solve MDR induced by P-gp.
Noncompetitive inhibition
Noncompetitive inhibition means that the inhibitor
does not compete with the chemotherapeutic agent as a substrate for
binding P-gp. There are a number of natural products (61-63)
that have been reported as noncompetitive inhibitors of P-gp.
Acetylshikonin and acetoxyisovalerylshikonin, which are shikonin
derivatives, may be noncompetitive inhibitors of MDR-related
transport proteins. They can inhibit ABCB1, BCRP and MRP2
independent of concentration to increase the accumulation of
daunorubicin in carcinoma cells and chemotherapy sensitivity
(62). Phytic acid, a natural
phosphorylated inositol, can also noncompetitively suppress P-gp;
this may be related to reduced P-gp ATPase activity and altered
conformations of the P-gp molecule (61). A previous study reported that
(±)-30-O, 40-O-dicynnamoyl-cis-kellactone, a derivative of
(±)-praeruptorin A, noncompetitively inhibits P-gp-mediated DOX
efflux by changing the conformation of P-gp and inhibiting ATPase
activity (63).
Some synthetic substances can also be used as
noncompetitive inhibitors to inhibit P-gp. Quaternized thiourea
main-chain polymer, which is not a substrate for P-gp, has been
shown to induce DNA damage and overcome P-gp-associated tumor MDR
(64). The novel microtubule
inhibitor
5-(4-ethoxyphenyl)-1-(3,4,5-trimethoxyphenyl)-1H-1,2,4-triazol-3-amine
(YAN), which is not a substrate of P-gp, enhances the cytotoxicity
of paclitaxel in NSCLC by decreasing the expression and efflux
activity of P-gp (65).
It can be concluded from the aforementioned reports
that noncompetitive inhibitors do not bind to the efflux substrate
binding pockets of P-gp. However, they may combine with other
domains of P-gp to affect its conformation. This principle is
similar to the application of allosteric regulators that do not
bind to the active site of target proteins to induce protein
conformational changes.
Competitive inhibition
Competitive inhibitors binding to P-gp hinder
P-gp-drug binding and P-gp-mediated efflux pathways. This method of
replacing the drug with another substance as the target of efflux
significantly increases the intracellular concentration of the
drug.
Small molecule targeted drugs branebrutinib
(66), pyrazolo[3,4-d]pyrimidines
(67), almonertinib (68) and ribociclib (69) may be potential inhibitors of P-gp.
They can compete with P-gp while exerting their own tumor-killing
effects, thereby improving chemotherapy sensitivity. This means
that there is a dual antitumor effect of chemotherapy combined with
targeted therapy. Almonertinib can bind to the TMD of P-gp, whereas
ribociclib and branebrutinib interact with the substrate-binding
site of P-gp.
As well as existing drugs, a number of natural
products have also been shown to inhibit P-gp. Wilforine (14), betulin (70), betulinic acid (71) and lathyrane diterpenes (72), which are terpenoids, all
competitively inhibit P-gp efflux activity and overcome MDR.
Wilforine may bind to residues of P-gp, such as Leu884, Lys887,
Thr176 and Asm172 (14). Betulin
tightly binds to the TMD region of P-gp (70). Glabridin (73) and 5-hydroxy-7,8-dimethoxyflavanone
(74), which are flavonoid
compounds, have also been proven to competitively inhibit P-gp and
to increase the accumulation of chemotherapy drugs. Notably,
5-hydroxy-7,8-dimethoxyflavanone may slightly change the
conformation of P-gp. Min et al (75) proposed that emodin, which is an
anthraquinone derivative, can competitively bind to the R site of
P-gp to inhibit the efflux function of P-gp and reduce the
resistance to DOX. Voacamine, an alkaloid, has been validated as a
substrate of P-gp, which was revealed to competitively inhibit the
P-gp-mediated export of paclitaxel in ovarian cancer (76). Due to the development of computer
simulation techniques and protein structure prediction, precise
binding sites of these substances to P-gp have been reported. For
the exploration of competitive inhibitors of P-gp, there is a large
library of active compounds in natural products. Compared with
curcumin, the synthesized dimethyl curcumin pyrazole derivatives
with N-p-phenylcarboxyamide substitution have higher binding
affinity for P-gp and can effectively reverse MDR of chronic
myeloid leukaemia to DOX (77).
Further modification of natural products enhances the inhibition of
P-gp while reducing side effects. This is a future direction for
the study of natural products as lead compounds for P-gp
inhibition.
The sensitivity of adrenocortical carcinoma to
mitotane, DOX, etoposide, cisplatin and streptozotocin can be
significantly enhanced by verapamil and tariquidar, which are known
as competitive P-gp inhibitors and nontransporter P-gp inhibitors
(78). DHW-221, a dual PI3K/mTOR
inhibitor, has also been found to bind P-gp to competitively
inhibit efflux function and downregulate P-gp (15). The triazolo[1,5-a]pyrimidine
derivative WS-716 (79), some
1,4-substituted arylalkyl piperazine derivatives (80), pyrimidine aminobenzene derivatives
(81), 5-phenylfuran derivatives
characterized by alkyl-substituted phenols and
6,7-dimethoxy1,2,3,4-tetrahydroisoquinoline (82), and the 4-indolyl quinazoline
derivative YS-370 (83) have been
reported to be potential P-gp competitive inhibitors. They block
substrate efflux only by binding to P-gp, but do not affect P-gp
expression. The inhibitory activity of most synthetic compounds is
an accidental discovery distinct from their initial pharmacological
action. However, the initial pharmacological action on patients
cannot be ignored as a potential side effect; this may hinder the
use of existing drugs to inhibit P-gp.
At present, most recognized P-gp inhibitors are
competitive inhibitors. High-affinity compounds that can be pumped
out of the cell as P-gp substrates have the potential to reverse
P-gp-associated MDR as competitive inhibitors. However, it is
necessary to consider the physiological activity of these compounds
on the body, which makes it difficult to use competitive inhibitors
in clinical MDR. The application of computer aided simulation
technology has significantly improved the screening efficiency of
P-gp competitive inhibitors and provided the basis for protein
conformation binding in biological experiments.
Uncertain inhibition
Aside from definite competitive and noncompetitive
inhibitors, a number of substances that have not been demonstrated
to inhibit the efflux activity of P-gp in a clear way can also
increase the sensitivity to chemotherapy drugs in certain
experiments. According to the source, inhibitors with uncertain
mechanisms can be divided into chemically synthesized substances
and natural products.
Although it has been reported that decitabine
upregulates P-gp expression in resistant cells, it has been found
that the drug sensitivity of hematomas and solid tumors with high
P-gp expression could be restored by decitabine through activation
of the MAPK signaling pathway and inhibition of the efflux pump
activity of P-gp (84).
Methoxypolyethylene glycol (MPEG)-glycine-quinidine conjugates,
which couple MPEG to quinidine via a glycine linker, can also
inhibit P-gp in resistant cancer cells (85), and enequidar, a gut-specific P-gp
inhibitor, reduces DOX efflux by inhibiting P-gp activity (86).
There are more studies on natural P-gp inhibitors
than synthetic P-gp inhibitors. Celastrol has two possible
antitumor drug resistance targets, the sarcoplasmic/endoplasmic
reticulum Ca2+ATPase (SERCA) pump and P-gp. The sensitivity to
chemotherapy drugs can be increased by celastrol, which
simultaneously inhibits SERCA to induce autophagy in resistant
tumor cells and inhibits the efflux pump function of P-gp (87). Naringenin and dihydrokaempferol
extracted from Pistacia integerrima can bind to the
hydrophobic pocket of P-gp; therefore, naringenin and
dihydrokaempferol may be used to reverse MDR by inhibiting the
activity of P-gp (88). Moreover,
it has been reported that Camellia sinensis non-fermentative
extract Noviphenone can overcome Adriamycin resistance by
inhibiting the activity of P-gp. instead of its expression levels
(89). The sensitivity to DOX can
be restored in paclitaxel-resistant HepG2 and DOX-resistant
colon-26 cells without influencing P-gp expression when the cells
were treated with the 50% ethanol extract of Azadirachta
indica, thus it has been inferred that the 50% ethanol extract
of A. indica can inhibit the activity of P-gp based on the
accumulation of the substrate rhodamine 123 (Rh-123) (90).
Antibodies against ABCB1 seem to be the most direct
and targeted way to inhibit the activity of ABCB1. Monoclonal
antibody (mAb) ABCB1-modified chitosan nanoparticles loaded with
gefitinib and the autophagy inhibitor chloroquine have been
prepared to overcome acquired EGFR-TKI resistance. The binding of
mAb-ABCB1 to receptor (P-gp) suppresses the efflux pump activity of
ABCB1, leading to the accumulation of chemotherapeutics (91).
Compared with competitive and noncompetitive
inhibitors, uncertain inhibitors can suppress the efflux activity
of P-gp without clear mechanisms and are therefore not considered
safe candidates. Studies have shown that these inhibitors do not
affect the expression of P-gp. Understanding the mechanism of
action of uncertain inhibitors is the key to promote their clinical
application.
5. Reversing MDR by inhibiting P-gp
expression
RNA interference
lncRNAs
lncRNAs are a class of noncoding RNAs that are
involved in intracellular regulation, including the function of
overcoming MDR caused by P-gp (Table
I).
 | Table IExamples of RNA molecules reversing
P-gp-related chemotherapy resistance. |
Table I
Examples of RNA molecules reversing
P-gp-related chemotherapy resistance.
A, lncRNAs
|
|---|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|---|
| Chen, 2020 | lncRNA-GAS5 | Breast cancer | Adriamycin | Regulating
miR-221-3p/DKK2 axis and Wnt/β-catenin/ABCB1 pathway | (92) |
| Han, 2018 | lncRNA LUCAT1 | Osteosarcoma | Methotrexate | Regulating
miR-200c/ABCB1 pathway | (93) |
| Kang, 2020 | lncCRNDE | Acute myeloid
leukemia | Adriamycin | Inhibiting
Wnt/β-catenin pathway and P-gp expression | (94) |
| Zou, 2019 | lncRNA
LINC00152 | Ovarian cancer | Cisplatin | Inhibiting P-gp
expression | (95) |
|
| B, miRNAs |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|
| Zou, 2017 | miR-495 | Ovarian cancer,
gastric cancer | DOX, taxol | Inhibiting P-gp
expression by binding to the 3′-UTR of ABCB1 | (96) |
| Zhao, 2017 | miR-491-3p | Hepatocellular
Carcinoma | DOX,
vinblastin | Inhibiting P-gp
expression by binding to the 3′-UTR of ABCB1 | (97) |
| Deng, 2021 | hsa-miR-34a-5p | Gastric cancer | 5-Fu | Inhibiting
expression of SIRT1 and P-gp by binding to the 3′-UTR of SIRT1 | (43) |
| Yi, 2019 | miR-381 | Breast cancer | Cisplatin | Directly inhibiting
ABCB1 expression | (98) |
| Zheng, 2019 | miR-34a | Hepatocellular
carcinoma | DOX | Possibly inhibiting
P-gp expression by regulating p53 expression | (99) |
| Wei, 2020 | miR-451 | Bladder cancer | Adriamycin | Inhibiting P-gp
expression | (100) |
| Kenworthy,
2018 | miR-149 | Mesothelioma | Taxane | Inhibiting P-gp
expression | (101) |
| Cao, 2019 | miR-122 | Hepatocellular
carcinoma | Oxaliplatin | inhibiting
Wnt/β-catenin pathway and P-gp expression | (103) |
| Wang, 2022 | miR-200c | Gastric cancer | Vincristine | Inhibiting P-gp
expression | (102) |
| Sousa, 2020 | miR-204-5p | Lung cancer | DOX | Unreported | (104) |
| Sousa, 2020 | miR-139-5p | Lung cancer | DOX | Unreported | (104) |
| Sousa, 2020 | miR-29c-5p | Lung cancer | DOX | Unreported | (104) |
| Sousa, 2020 | miR-551b-3p | Lung cancer | DOX | Unreported | (104) |
| Sousa, 2020 | miR-29b-2-5p | Lung cancer | DOX | Unreported | (104) |
| Sousa, 2020 | miR-204-3p | Lung cancer | DOX | Unreported | (104) |
| Xie, 2021 | miR-1246 | Leukemia | Adriamycin | Inhibiting
AXIN2/GSK-3β/Wnt/β-catenin pathway and P-gp expression | (106) |
| Li, 2022 | miR-205 | Liver cancer | DOX | Regulating
PTEN/PI3K/Akt pathway and P-gp expression | (24) |
| Shi, 2020 | miR-29a | Colon cancer | DOX | Regulating
PTEN/PI3K/Akt pathway and P-gp expression | (105) |
|
| C, siRNAs |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|
| Peng, 2022 | si ABCB1 | Breast cancer | DOX | Silencing ABCB1
gene | (154) |
| Liu, 2022 | si ABCB1,
siBcl-2 | Breast cancer | DOX | Silencing ABCB1
gene | (112) |
| Wu, 2021 | si ABCB1 | Ovarian cancer | Paclitaxel | Silencing ABCB1
gene | (113) |
The overexpression of some lncRNAs can directly or
indirectly inhibit P-gp-mediated MDR. lncRNA-GAS5 can directly bind
to miR-221-3p and inhibit its expression, the downregulation of
which inhibits Dickkopf-related protein (DKK)2 and then activates
the Wnt/β-catenin signaling pathway; ultimately, P-gp expression is
reduced in DOX-resistant breast cancer cells (92). In methotrexate-resistant
osteosarcoma cells, miR-200c inhibition can upregulate the
expression of lncRNA LUCAT1 and P-gp. Upregulation of lncRNA LUCAT1
promotes P-gp expression and P-gp-mediated MDR (93). Knockout of some lncRNAs can also
reverse MDR. The expression of colorectal neoplasia differentially
expressed (CRNDE) in drug-resistant acute myeloid leukemia cells is
directly proportional to the expression of ABCB1 and the
sensitivity to Adriamycin has been reported to be enhanced after
lncRNA CRNDE knockout. These results may be related to the
inhibition of Wnt/β-catenin pathway by lncRNA CRNDE knockout
(94). Furthermore, the expression
of ABCB1is reduced when lncRNA LINC00152 is knocked down, which can
increase the chemosensitivity of epithelial ovarian cancer cells to
cisplatin (95).
miRNAs
miRNAs are another type of non-coding RNA associated
with tumors. These single-stranded RNA molecules can bind to mRNA
to regulate gene expression. In recent years, the regulatory role
of miRNAs in tumor resistance has gradually attracted attention
(Table I).
miRNAs alleviate chemoresistance mainly by directly
(96,97) or indirectly (43,60)
downregulating P-gp in various types of cancer. It has been
reported that miR-495 (96) and
miR-491-3p (97) can bind to two
sites on the 3′-untranslated region (UTR) of ABCB1 mRNA, thereby
inhibiting the expression of ABCB1 and sensitizing cancer cells to
chemotherapeutic drugs. The high expression of hsa-miR-34a-5p in
gastric cancer cells can inhibit uracil resistance by directly
binding to the 3′-UTR of sirtuin 1 (SIRT1), and downregulating
SIRT1, P-gp and MRP1 (43).
Numerous studies have shown that the expression of
miRNAs in cancer tissues is associated with the levels of P-gp. It
has been reported that upregulation of miR-381 (98), miR-34a (99), miR-451 (100), miR-149 (101) and miR-495 (96) can significantly reduce P-gp
expression, thereby enhancing the sensitivity of cancer tissues to
chemotherapeutic drugs. A retrospective clinical study reported the
negative association between miR-200c and the expression of P-gp in
gastric cancer tissue. miR-200c overexpression and P-gp
downregulation were shown to reverse MDR by increasing drug
sensitivity and predicted a better prognosis in patients with
gastric cancer (102). The
overexpression of miR-122, which has been reported as a negative
regulator, can also inhibit the Wnt/β-catenin signaling pathway and
further suppress ABCB1 expression (103). A previous study revealed that
there are a number of miRNAs (miR-204-5p, miR-139-5p, miR-29c-5p,
miR-551b-3p, miR-29b-2-5p and miR-204-3p) that are specific to lung
cancer cells with P-gp-mediated resistance and extracellular
vesicles. These miRNAs are likely markers of MDR or key molecules
that could be used to overcome P-gp-mediated MDR (104). It has also been reported that
miR-205 (24) and miR-29a
(105) significantly inhibit P-gp
expression by upregulating protein tyrosine phosphatase, thereby
enhancing the sensitivity of cancer tissues to DOX.
In addition to the association between the
overexpression of miRNAs and the downregulation of P-gp,
downregulation of certain miRNAs can also inhibit P-gp expression
and MDR through complex signaling pathways. The chemoresistance of
MDR leukaemia cells can be attenuated by the loss of miR-1246 via
negatively regulating axis inhibition protein 2 and glycogen
synthase kinase-3β to inactivate the Wnt/β-catenin pathway and
suppress P-gp expression (106).
Several miRNAs that inhibit P-gp expression have
been identified; however, the expression of P-gp is not the result
of a single miRNA. Although miRNAs do not translate proteins, they
are involved in regulating P-gp through a variety of signaling
pathways.
siRNAs
siRNAs are short non-coding RNAs that can regulate
gene expression in the body (107). siRNA-ABCB1 (si ABCB1) targets
ABCB1 mRNA to form a double-stranded RNA and hinder the translation
of ABCB1 mRNA, decreasing P-gp expression and enhancing
chemotherapy sensitivity. Studies on si ABCB1 in tumor resistance
have mainly focused on the effect of si ABCB1 delivered by
different delivery systems in reversing MDR (108) and the therapeutic effect of
siABCB1 in combination with different drugs (109-111).
There have been numerous research achievements
regarding the delivery of siRNAs and other materials via
nanoparticles or nanocarriers (109,112,113). Liu et al (112) prepared a targeted nanoplatform
based on biodegradable boronic acid modified ε-polylysine to
codeliver P-gp siRNA, Bcl-2 siRNA and DOX. In breast cancer,
compared with P-gp siRNA, Bcl-2 siRNA and/or DOX alone, the
combined application of P-gp siRNA, Bcl-2 siRNA and DOX supported
by nanocarriers has a stronger antitumor effect and ABCB1
gene-silencing effect (112).
Wang et al (109) designed
a biomimetic lipid/dextran hybrid nanocarrier with a diameter of
~100 nm. In this nanocarrier, the anionic si ABCB1 and the
hydrophobic drug paclitaxel were loaded into the cationic lipid
shell and the hydrophobic internal core of the hybrid nanocarriers,
respectively, in order to improve the effect of paclitaxel. In
another previous study, a multifunctional polymeric nanoparticle
called Py-TPE/siRNA@PMP was loaded with paclitaxel and si ABCB1;
this nanoparticle drug delivery system has been shown to enhance
chemotherapy sensitivity in ovarian cancer (113).
As well as nanoparticles, there are other types of
drug delivery systems. An EphA10-mediated pH-sensitive lipoplex has
been developed to codeliver ABCB1-targeting siABCB1 and DOX
(114). In another study, a
carrier-free system with MDR as the backbone, loaded with cisplatin
and divalent copper was constructed to treat drug-resistant tumors
(110).
Developments in the field of materials have improved
the delivery of siRNAs in cells. The use of materials to deliver
chemotherapeutic drugs and siRNA at the same time allows both
agents to be of maximum benefit. If the modified material can be
delivered to the target tumor, it will greatly attenuate the
downregulation of P-gp in normal tissue and the side effects of
chemotherapy.
Drugs that inhibit P-gp expression
Chemically synthesized products
In the process of exploring tumor MDR, a number of
chemically synthesized products have been detected to inhibit tumor
MDR. Studies conducted in recent years have shown that some
anticancer and non-anticancer drugs can downregulate P-gp, enhance
chemosensitivity and rescue tumor MDR.
According to a previous study, hypoxia increases
HIF1α expression resulting in P-gp overexpression and NSCLC MDR. By
contrast, the HIF1α inhibitor LW6 can attenuate cisplatin MDR by
downregulating P-gp (20). A novel
platinum complex can also effectively alleviate cisplatin
resistance by inhibiting the expression of P-gp (115). Ribociclib, a CDK4/6 inhibitor,
inhibits P-gp-mediated DOX resistance by downregulating the
expression and efflux activity of P-gp (69). The small molecule targeted
inhibitors sorafenib and apatinib also inhibit P-gp expression and
MDR while inducing anticancer effects (116). The killing effect of these drugs
may cause tumors to lose the self-protection mechanism of
chemotherapy-induced P-gp overexpression.
LSS-11, a novel naphthalimide derivative-based
topoisomerase inhibitor, may reduce paclitaxel resistance in A549
lung cancer cells through two different underlying pathways. One of
the mechanisms is that it could hinder the binding of STAT3 to the
ABCB1 promoters downregulating P-gp (117). A novel microtubule inhibitor,
YAN, can inhibit the resistance of NSCLC to paclitaxel by
inhibiting the expression of P-gp (65). BAY-1082439, a chemical molecule
undergoing phase I clinical trials in patients with advanced
cancer, can efficiently inhibit the expression of P-gp and BCRP by
selectively inhibiting PI3K 110α and 110β, and slightly suppress
the efflux effect of P-gp and BCRP (118). A tumor-targeted anticancer agent,
FFCLB, covalently linked by the delocalized lipophilic cation FF
and CLB, inhibits P-gp expression, thereby increasing the
intracellular concentration and cytotoxicity of DOX (119).
According to the aforementioned results, a number of
drugs or their derivatives used to treat other diseases can inhibit
P-gp expression and reverse chemotherapy resistance. It may be
hypothesized that there is an intersection between the signaling
pathway these drugs originally interact with to treat disease and
the signaling pathway that regulates P-gp expression. Although
these drugs can overcome P-gp-mediated chemotherapy resistance,
their originally discovered effects are not negligible. However,
the combination of small molecule targeted drugs with inhibitory
activity and chemotherapy drugs may play a dual role in cancer
suppression.
Natural products
In the search for antitumor molecules to combat
tumor resistance, screening of natural products may yield safer
options than screening chemical compounds. To date, a variety of
natural product-derived compounds (120) and plant extracts (121) have been shown to inhibit ABCB1
and reverse tumor MDR in vitro and in vivo. These
research objects can be roughly divided into flavonoids, alkaloids,
coumarins, saponins, lignans, phenolic acids and terpenes. The
present review summarized these natural products that reverse tumor
drug resistance by inhibiting ABCB1 expression (Table II).
| Table IIExamples of natural products
reversing P-gp-related chemotherapy resistance. |
Table II
Examples of natural products
reversing P-gp-related chemotherapy resistance.
A, Flavonoids
|
|---|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|---|
| Kong, 2020 | Hesperetin | Lung cancer | Cisplatin | Inhibiting NF-κB
pathway and P-gp expression | (124) |
| Sui, 2021 | Cajanol | Ovarian cancer | Paclitaxel | Inhibiting
PI3K/Akt/NF-κB pathway and P-gp expression | (51) |
| Qin, 2022 | Luteolin | Osteosarcoma | Cisplatin and
DOX | Upregulate miR-384,
inhibiting PTN/β-catenin axis and P-gp expression | (60) |
| Wang, 2018 | Icaritin | Osteosarcoma | DOX | Inhibiting P-gp
expression | (127) |
| Satonaka, 2017 |
(-)-Epigallocatechin-3-gallate | Liver cancer | DOX | Inhibiting PI3K/Akt
and MEK/ERK pathway, P-gp expression | (57) |
| La, 2019 |
(-)-Epigallocatechin gallate | Colorectal
cancer | 5-Fu | Inhibiting
GRP78/NF-κB/miR-155-5p/ABCB1 pathway and P-gp expression | (125) |
| Tang, 2017 | Epigallocatechin
gallate | Gastric cancer | 5-Fu | Inhibiting
TFAP2A/VEGF pathway and P-gp expression | (42) |
| Qian, 2019 | Glabridin | Breast cancer | Paclitaxel,
DOX | Inhibiting P-gp
expression and P-gp ATPase activity, competitively inhibiting the
efflux activity of P-gp | (73) |
| Teng, 2021 |
5-Hydroxy-7,8-dimethoxyflavanone | Oral cavity
carcinomas | Vincristine | Competitively
inhibiting the efflux activity of P-gp | (74) |
|
| B, Alkaloids |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|
| Huang, 2017 | Cepharanthine | Ovarian cancer | Paclitaxel | Inhibiting PI3K/Akt
pathway and P-gp expression | (132) |
| Zhou, 2017 | Cepharanthine | Esophageal squamous
cell carcinoma | Cisplatin | Activating JNK/p53
pathway and inhibiting P-gp expression | (129) |
| Zhou, 2022 | Tetrandrine | Leukemia | Daunorubicin | Inhibiting the
efflux activity and expression of P-gp | (130) |
| Qian, 2021 | Berberine | Breast cancer | DOX | Inhibiting the
efflux activity and expression of P-gp | (131) |
| Liu, 2020 | Nuciferine | Lung cancer, bowel
cancer | Paclitaxel, DOX,
docetaxel, daunorubicin | Inhibiting
PI3K/AKT/ERK pathway and activation of Nrf2 and HIF-1α, P-gp
expression | (55) |
| Quarti, 2021 | Piperine | Leukemia | Vincristine | Inhibiting P-gp
expression | (134) |
|
| C, Coumarins |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|
| Wang, 2011 | Archangelicin | Leukemia | DOX | Inhibiting the
function and expression of P-gp | (135) |
| Wang, 2016 | Osthole | Leukemia | DOX | Inhibiting PI3K/Akt
pathway and expression of P-gp | (56) |
| Shen, 2006 | (±)-30-O,
40-O-dicynnamoyl-cis-kellactone | Hepatocellular
carcinoma, leukemia | DOX | Noncompetitively
inhibiting the efflux activity of P-gp | (63) |
|
| D, Saponins |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
| Huang, 2012 | Astragaloside
II | Hepatic cancer | 5-Fu | Inhibiting the
efflux activity and expression of P-gp via phosphorylation of
ERK1/2, p38 and JNK | (140) |
| Wang, 2017 | Astragaloside
IV | Hepatic cancer | 5-Fu | Inhibiting
JNK/c-Jun/AP-1 pathway and P-gp expression | (59) |
| Liu, 2018 | Ginsenoside
Rh2 | Colorectal
carcinoma | 5-Fu | Inhibiting P-gp
expression | (142) |
| Liu, 2018 | 20(S)-ginsenoside
Rg3 | Lung cancer | Cisplatin | Inhibiting P-gp
expression | (143) |
|
| E, Lignans |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|
| Sadava, 2013 | Silibinin | Small cell lung
carcinoma | Etoposide, DOX | Possibly inhibiting
P-gp expression | (144) |
| Dobiasová,
2020 | Silibinin
stereoisomer В | Ovarian cancer | DOX | Inhibiting P-gp
expression and P-gp ATPase activity | (145) |
| Dobiasová,
2020 |
2,3-Dehydrosilibinin A | Ovarian cancer | DOX | Unreported | (145) |
| Zhang, 2018 | Schisandrin A | Breast cancer | DOX | Inhibiting the
P-gp, NF-кВ and Stat3 pathway, and inhibiting P-gp expression | (146) |
|
| E, Phenolic
acids |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
| Liao, 2020 | Rosmarinic
acid | Non-small cell lung
cancer | Cisplatin | Possibly activating
JNK phosphorylation and inhibiting P-gp expression | (147) |
| Muthusamy,
2019 | Ferulic acid | Oral cavity
carcinomas | DOX,
vincristine | Inhibiting
PI3K/Akt/NF-κB pathway and P-gp expression | (148) |
| Gao, 2021 | Curcumol | Colon cancer | 5-Fu | Inhibiting
Wnt/β-catenin pathway and P-gp expression | (23) |
| Teng, 2020 | Caffeic acid | Oral cavity
carcinomas | DOX | Competitively
inhibiting efflux activity of P-gp | (155) |
|
| F, Terpenes |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
|
| Oh, 2020 | Cordycepin | Bladder cancer | Cisplatin | Inhibiting PI3K
pathway, ETS-1 phosphorylation and P-gp expression | (149) |
| Ma, 2020 | Asiaticoside | Hepatocellular
carcinoma | DOX, paclitaxel,
colchicine, vincristine | Inhibiting PI3K/Akt
and MAPK/ERK pathway | (156) |
| Ding, 2022 | Alantolactone | Lung cancer | Paclitaxel | Inhibiting
MALAT1/STAT3/FUT4 axis and P-gp expression | (58) |
| Ding, 2022 | Brevilin A | Lung cancer | Paclitaxel | Inhibiting
MALAT1/STAT3/FUT4 axis and P-gp expression | (58) |
| Chang, 2020 | Wilforine | Cervical
cancer | DOX | Competitively
inhibiting the efflux activity of P-gp | (14) |
| Laiolo, 2021 | Betulin | Leukemia | DOX | Competitively
inhibiting the efflux activity of P-gp | (70) |
| Liu, 2021 | Betulinic acid | Breast cancer | Adriamaycin,
vincristine | Competitively
inhibiting the efflux activity of P-gp | (71) |
| Yang, 2020 | Lathyrane
diterpenes | Hepatocellular
carcinoma | DOX | Competitively
inhibiting the efflux activity of P-gp | (72) |
|
| G, Quinones |
|
| First author,
year | Substance | Tumor type | Corresponding
drug | Anti-MDR
mechanism | (Refs.) |
| Mirzaei, 2018 | Acetylshikonin | Breast cancer | Daunorubicin | Noncompetitively
inhibiting the efflux activity of P-gp | (62) |
| Mirzaei, 2018 |
Acetoxyisovalerylshikonin | Breast cancer | Daunorubicin | Noncompetitively
inhibiting the efflux activity of P-gp | (62) |
Flavonoids
Flavonoids are a class of plant metabolites with a
2-phenylchromone structure that exist widely in nature. A variety
of flavonoids have been shown to have antitumor and anti-resistance
activities (122). Different
structures of flavonoids have different mechanisms of action and
inhibitory activities (123).
The NF-κB signaling pathway is involved in the
mechanism by which a variety of natural compound monomers inhibit
P-gp expression. Hesperetin, a derivative of hesperidin that is
extracted from tangerine peel, has been reported to inhibit tumor
growth in vivo and to restore the sensitivity of A549 lung
cancer cells to cisplatin by inhibiting the NF-κB signaling pathway
and expression of P-gp (124).
Cajanol, an isoflavone from pigeon pea, can inhibit the
transcription and translation of P-gp through the PI3K/Akt/NF-κB
pathway, and reverse the resistance of ovarian cancer cell lines to
paclitaxel in vitro and in vivo (51). It has also been reported that P-gp
expression and P-gp ATPase activity are suppressed by glabridin
(73).
The regulation of P-gp expression by various natural
products is related to miRNAs (60,125). Luteolin can enhance the
sensitivity of osteosarcoma to cisplatin and DOX; it has been
reported that luteolin can upregulate miR-384, which indirectly
inhibits P-gp expression by negatively regulating the PTN/β-catenin
axis (60).
Additionally, natural products that have been shown
to inhibit P-gp expression also include epigallocatechin gallate
(42), theaflavin, quercetin,
rutin, epicatechin 3 gallate, tamarixetin (126) and icaritin (127).
Alkaloids
Salvia miltiorrhiza (128), cepharanthine hydrochloride
(129), tetrandrine (130) and berberine (131) can inhibit the expression of P-gp
to reverse tumor MDR. Cepharanthine hydrochloride can serve a role
in reversing drug resistance in ovarian cancer via inhibiting the
expression of ABCB1, and reducing the mRNA and protein expression
levels of P-gp, which may be caused by inhibiting the PI3K/Akt
signaling pathway (132).
Tetrandrine (133), nuciferine
(55) and berberine (131) have been reported to increase the
intracellular aggregation of chemotherapeutic drugs via their dual
inhibitory effects of inhibiting the efflux activity of P-gp and
inhibiting the expression of P-gp. Tetrandrine and berberine are
isoquinoline alkaloids. Notably, inhibition of P-gp expression by
tetrandrine does not occur through the inhibition of ABCB1
transcription (133).
Furthermore, inhibition of P-gp expression by nuciferine is
regulated by a series of signaling pathways. Nuciferine can
effectively inhibit the PI3K/AKT/ERK signaling pathway, and the
activation of Nrf2 and HIF-1α, which can reduce P-gp expression and
enhance MDR sensitivity in tumor cells (55). Additionally, the ability of
piperine to overcome MDR in leukaemia cells may be related to
downregulation of P-gp (134).
Coumarins
Coumarins are a class of aromatic compounds
ubiquitous in nature with a benzopyrone structure. Some coumarins
have been reported to overcome MDR by inhibiting the activity and
expression of P-gp. A previous study revealed that archangelicin
can inhibit the function and expression of P-gp, and ultimately
reduce the resistance of the leukaemia cell line K562/A02 to DOX
(135). Osthole has also been
shown to inhibit the expression of P-gp by inhibiting the PI3K/Akt
signaling pathway, which can be induced by the uptake and efflux of
Rh-123 and the accumulation of DOX (56).
Saponins
Saponins, consisting of sapogenins and sugars, are
mainly found in plants. There are a number of types and complex
structures of saponins (136).
Numerous saponins, including Paris saponin VII (137), ginsenoside Rb1 (138) and sodium aescinate (139), have been reported to inhibit the
expression of P-gp and reverse tumor resistance. Astragaloside II
has been found to increase the intracellular accumulation of P-gp
substrates by inhibiting the efflux pump activity of P-gp. On the
other hand, it can also inhibit the expression of ABCB1 and
suppress the phosphorylation of ERK1/2, p38 and JNK; these
functions facilitate its role in reversing tumor MDR (140). Wang et al (141) investigated astragaloside IV and
verified that astragaloside IV can inhibit the expression of ABCB1,
increase the intracellular accumulation of 5-Fu, and ultimately
achieve the goal of inhibiting tumor resistance. In subsequent
experiments, it was further confirmed that the specific mechanism
by which astragaloside IV inhibits the expression of ABCB1 is by
inhibiting the JNK/c-Jun/AP-1 signaling pathway (59). In addition, the activity of
ginsenoside Rh2 in reversing tumor resistance may be related to its
inhibition of P-gp expression. In a previous study, it was revealed
that the expression levels of MRP1, ABCB1, LRP and glutathione
S-transferase are negatively correlated with ginsenoside Rh2
(142). In addition to
ginsenoside Rh2, 20(S)-ginsenoside Rg3 can also enhance the
cytotoxicity of chemotherapy drugs by inhibiting the expression of
drug resistance genes (143).
Lignans
Silibinin is considered to be the main bioactive
component of silymarin. In a previous report, it was speculated
that silibinin increased the chemosensitivity of small cell lung
cancer by downregulating P-gp rather than inhibiting the efflux
function of P-gp (144). However,
in recent years, silibinin stereoisomer B and its derivative
2,3-dehydrosilibinin A have also been found to sensitize ovarian
cancer cells to DOX, and silibinin stereoisomer B exhibits dual
inhibition of P-gp ATPase activity and P-gp expression (145). Schisandrin A can also inhibit the
expression of P-gp by reducing the levels of phosphorylated
(p)-IκB-α (Ser32) and p-Stat3 (Tyr705). Combined with its blocking
of P-gp efflux activity, it enhances the sensitivity of MDR breast
cancer cells to DOX (146).
Phenolic acids
Rosmarinic acid reverses cisplatin resistance in
NSCLC by activating MAPK and thereby inhibiting P-gp expression
(147). Ferulic acid has also
been reported to enhance the cytotoxicity of DOX and vincristine on
MDR cells with high P-gp expression, which is dependent on the
PI3K/Akt/NF-κB signaling pathway to inhibit P-gp expression
(148). Curcumin has also been
reported to downregulate the expression of P-gp (23).
Terpenes
Terpenoids are widely found in animals and plants,
such as essential oils, resins, pigments, hormones and vitamins.
Terpenoids have isoprene basic units in their structure. Cordycepin
inhibits the expression of ABCB1, thereby enhancing the sensitivity
of cisplatin-resistant bladder cancer cells to cisplatin.
Cordycepin may inhibit the PI3K signaling pathway and thereby
attenuate the phosphorylation of ETS-1 (Thr38), a transcription
factor that can activate the ABCB1 promoter after phosphorylation.
The reduction in ETS-1 (Thr38) phosphorylation hinders the
transcription and expression of ABCB1, downregulates the expression
of P-gp and reduces the resistance of bladder cancer cells
(149). Alantolactone and
brevilin A, which are sesquiterpene lactones, can inhibit the
resistance of lung cancer cells to paclitaxel (58). The mechanism involves the
inhibition of MALAT1 expression by alantolactone and brevilin A,
the subsequent activation of STAT3 and the upregulation of FUT4,
and ultimately the positive reduction of P-gp expression.
These natural products inhibit P-gp expression and
reverse chemotherapy resistance by regulating the NF-κB/STAT3,
PI3K/AKT, MEK/ERK and Wnt/β-catenin signaling pathways. These
involved signaling pathways are consistent with those that promote
P-gp-related chemotherapy resistance in different tumors.
Furthermore, these results confirm that these signaling pathways
are important and inhibitors targeting these signaling pathways
should be studied.
Biological compounds
In addition to some of their own biological
activities, a number of biological compounds in the human body,
including proteins, enzymes, nucleic acids, hormones, antibodies
and cytokines, have been proven to be effective in antitumor
MDR.
Tumor necrosis factor α-induced protein 8-like 2
(TIPE2) is a tumor suppressor that is related to P-gp. P-gp can be
inhibited by TIPE2 via inhibiting the TAK1-NF-κB-AP-1 signaling
pathway, thus sensitizing osteosarcoma cells to cisplatin (52). It may be hypothesized that the
overall inhibition of tumors by other targets may affect the
expression of P-gp in tumors. This seems to confirm the win-win
effect of chemotherapy in combination with other treatments. The
NF-κB signaling pathway has an important role in regulating P-gp
expression. Melatonin downregulates NF-κB/P65 expression in the
nucleus and disrupts the binding of P65 to the ABCB1 promoter.
Melatonin-mediated downregulation of P-gp expression increases the
chemosensitivity of diffuse large B-cell lymphoma cells to
epirubicin (54). Estrogen
2-methoxyestradiol can interact with cadmium in ovarian cancer to
reduce P-gp expression as a potential MDR regulator (150). γ-Tocotrienol inhibits P-gp
expression in breast cancer via the NF-κB signaling pathway
(151). The impact of tumors on
the body is systemic rather than local and the malignant
progression of tumors is accompanied by changes in hormone levels.
To overcome P-gp mediated MDR, systemic treatment, such as targeted
therapy combined with chemotherapy and targeted therapy combined
with immunotherapy, should be considered (1,2).
In addition to these endogenous substances,
biologics used to inhibit P-gp in the body are also effective. It
has been found that an adenovirus vector carrying DKK3 can augment
the antitumor effect of temozolomide in glioblastoma cells by
reducing the expression of P-gp, which is associated with
inhibition of Akt/NF- (152).
Active substances in humans have better biosafety
profiles than those derived from natural products or synthetic
compounds. These biological compounds can regulate the expression
of P-gp by inhibiting or activating certain signaling pathways and
molecules. Therefore, regulation of P-gp expression and
chemotherapy resistance can be affected by altering the expression
levels or activities of these biological compounds in vivo
and in vitro.
6. Reversing MDR by silencing ABCB1 DNA
The reversal of P-gp-mediated tumor MDR by
inhibiting the activity and expression of P-gp has been introduced
in the present review. If these two methods block the intermediate
steps of P-gp-mediated tumor resistance, then directly knocking out
or silencing the ABCB1 gene can completely inhibit P-gp expression
and curb resistance.
CRISPR/Cas9 is an efficient and convenient gene
editing technology. It can use single guide RNA (sgRNA) to identify
target gene DNA and guide Cas9 endonuclease to cut DNA to achieve
gene knockout. Three sgRNAs targeting the fourth and fifth exons of
the human ABCB1 gene were designed by Norouzi-Barough et al
(153). When all three Cas9-sgRNA
plasmids were transfected into Adriamycin-resistant cells, the
three sgRNAs bound to the target exons of ABCB1 inducing the Cas9
endonuclease to cut the target gene ABCB1. Finally, ABCB1 was
knocked out. Low-level expression of P-gp was detected in the
experiment, which verified that CRISPR/Cas9 technology successfully
knocked down the ABCB1 gene. In addition, the increased sensitivity
of ABCB1-knockdown cells to Adriamycin suggested that knocking out
drug resistance genes through gene editing may be a new way to
solve drug resistance (153).
However, how to safely and effectively apply this to the clinical
treatment of cancer is a difficult problem to be solved. The
urgency of the problem is alleviated by the continual discovery of
new drugs.
7. Conclusions and perspective
Chemotherapy drugs inhibit tumor cells by killing
them indiscriminately and thus have cytotoxic effects on normal
cells, leading to strong side effects. Targeted therapy and
immunotherapy have fewer side effects and are emerging therapies
for cancer. However, due to the difference in individual immune
status, the therapeutic effect of immunotherapy differs
significantly among individuals. Differences in the phenotype of
targeted genes also limit the use of targeted therapy. Therefore,
chemotherapy remains an irreplaceable therapy in cancer
treatment.
P-gp upregulation in cancer leads to
chemoresistance since P-gp pumps chemotherapeutic agents out of the
cell. Multiple signaling pathways, including PI3K/AKT (25), MEK/ERK (21), Wnt/β-catenin and NF-κB/STAT3,
promote the chemotherapy resistance of various tumor cells. The
mechanism of P-gp upregulation is consistent in different tumors
(16,20,37,49).
Multiple signaling pathways, including PI3K/AKT (37), MEK/ERK (21), Wnt/β-catenin and NF-κB/STAT3,
promote the chemotherapy resistance of various tumor cells.
Notably, the consistent mechanisms in different tumors reduces the
burden of exploring ways to overcome P-gp-mediated chemotherapeutic
resistance in different tumors.
Currently, the pathways found to reverse P-gp
mediated chemotherapeutic resistance can be divided into three
categories: Inhibition of the efflux function of P-gp, inhibition
of the expression of P-gp and knockdown of the ABCB1 gene.
Competitive or noncompetitive inhibition of P-gp efflux function
represents the most direct method. This class of inhibitors, which
mainly includes natural products and synthetic compounds, impedes
the binding of P-gp to chemotherapy drugs by binding to P-gp or
changing the conformation of P-gp. Computer-aided simulation
technology can be used to screen P-gp inhibitors. Downregulation of
P-gp is the way in which most inhibitors act. The regulation of
P-gp expression by natural products, synthetic compounds, RNA and
biological compounds in humans involves a variety of mechanisms and
molecular targets, such as the PI3K/AKT (51), MEK/ERK, Wnt/β-catenin and
NF-κB/STAT3 (58) signaling
pathways. Numerous small molecule targeted inhibitors not only
exert antitumor effects, but also inhibit P-gp expression and MDR,
reflecting the increased efficacy of the combination of targeted
therapy and chemotherapy. However, compared with inhibition of
signaling pathways, direct inhibition of the substrate-binding site
and efflux activity of P-gp is more effective in theory. To some
extent, some P-gp competitive and noncompetitive inhibitors act in
a manner similar to allosteric regulation, regulating protein
activity by changing the structure of the protein. At present,
there is a complete and feasible development process for allosteric
regulators of target proteins. Perhaps the research of P-gp efflux
inhibitors can refer to the exploration of allosteric
regulators.
In conclusion, the present review describes
P-gp-mediated chemotherapeutic resistance and related mechanisms in
various types of cancer to alert researchers to the importance of
P-gp in chemotherapeutic resistance. It may be possible to explore
the related mechanisms to overcome P-gp-mediated chemoresistance.
In addition, the present review summarized the active substances
that reverse P-gp-mediated chemotherapeutic resistance to provide
candidate compounds for screening clinical P-gp inhibitors and lead
compounds for the development of new P-gp inhibitors.
Availability of data and materials
Not applicable.
Authors' contributions
YT and FX designed the article. YT, YL, YW and JL
participated in the literature search. YT was the major contributor
in writing the manuscript. FX and JW participated in manuscript
revision. Data authentication is not applicable. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
This work was supported by the National Major Scientific
Research Instrument Development Project of NSFC (grant no.
21827812) and the Foundation and Advanced Research Project of CQ
CSTC (grant nos. cstc2018jscx-mszd0280 and
cstc2017shms-xdny0033).
References
|
1
|
Xu T, Wang X, Xin Y, Wang Z, Gong J, Zhang
X, Li Y, Ji C, Sun Y, Zhao F, et al: Trastuzumab combined with
irinotecan in patients with HER2-positive metastatic colorectal
cancer: A phase II single-arm study and exploratory biomarker
analysis. Cancer Res Treat. 55:626–635. 2023. View Article : Google Scholar :
|
|
2
|
Torka P, Groman A, Wong J, Nichols J,
Kader A, Mavis C, Anampa-Guzmán A, Sait SJ, Block A, Przespolewski
E, et al: Carfilzomib combined with rituximab, ifosfamide,
carboplatin, and etoposide for relapsed or refractory DLBCL. Blood
Adv. 7:1146–1155. 2023. View Article : Google Scholar
|
|
3
|
Yin Q, Shen J, Zhang Z, Yu H and Li Y:
Reversal of multidrug resistance by stimuli-responsive drug
delivery systems for therapy of tumor. Adv Drug Deliv Rev.
65:1699–1715. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ruan T, Liu W, Tao K and Wu C: A review of
research progress in multidrug-resistance mechanisms in gastric
cancer. Onco Targets Ther. 13:1797–1807. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu C, Wei Y, Wang X, Zhang Z, Yin J, Li W,
Chen L, Lyu X, Shi Z, Yan W and You Y: DNA-methylation-mediated
activating of lncRNA SNHG12 promotes temozolomide resistance in
glioblastoma. Mol Cancer. 19:282020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Galle E, Thienpont B, Cappuyns S, Venken
T, Busschaert P, Van Haele M, Van Cutsem E, Roskams T, van Pelt J,
Verslype C, et al: DNA methylation-driven EMT is a common mechanism
of resistance to various therapeutic agents in cancer. Clin
Epigenetics. 12:272020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leon G, MacDonagh L, Finn SP, Cuffe S and
Barr MP: Cancer stem cells in drug resistant lung cancer: Targeting
cell surface markers and signaling pathways. Pharmacol Ther.
158:71–90. 2016. View Article : Google Scholar
|
|
8
|
Vitale D, Kumar Katakam S, Greve B, Jang
B, Oh ES, Alaniz L and Götte M: Proteoglycans and
glycosaminoglycans as regulators of cancer stem cell function and
therapeutic resistance. FEBS J. 286:2870–2882. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang H, Xu H, Ashby CR Jr, Assaraf YG,
Chen ZS and Liu HM: Chemical molecular-based approach to overcome
multidrug resistance in cancer by targeting P-glycoprotein (P-gp).
Med Res Rev. 41:525–555. 2021. View Article : Google Scholar
|
|
10
|
Briz O, Perez-Silva L, Al-Abdulla R, Abete
L, Reviejo M, Romero MR and Marin JJG: What 'the cancer genome
atlas' database tells us about the role of ATP-binding cassette
(ABC) proteins in chemoresistance to anticancer drugs. Expert Opin
Drug Metab Toxicol. 15:577–593. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He C, Sun Z, Hoffman RM, Yang Z, Jiang Y,
Wang L and Hao Y: P-glycoprotein overexpression is associated with
cisplatin resistance in human osteosarcoma. Anticancer Res.
39:1711–1718. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Juliano RL and Ling V: A surface
glycoprotein modulating drug permeability in Chinese hamster ovary
cell mutants. Biochim Biophys Acta. 455:152–162. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ibrahim MAA, Abdeljawaad KAA,
Jaragh-Alhadad LA, Oraby HF, Atia MAM, Alzahrani OR, Mekhemer GAH,
Moustafa MF, Shawky AM, Sidhom PA and Abdelrahman AHM: Potential
drug candidates as P-glycoprotein inhibitors to reverse multidrug
resistance in cancer: An in silico drug discovery study. J Biomol
Struct Dyn. 1–16. 2023.Epub ahead of print.
|
|
14
|
Chang YT, Lin YC, Sun L, Liao WC, Wang
CCN, Chou CY, Morris-Natschke SL, Lee KH and Hung CC: Wilforine
resensitizes multidrug resistant cancer cells via competitive
inhibition of P-glycoprotein. Phytomedicine. 71:1532392020.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu M, Xu C, Qin X, Liu W, Li D, Jia H,
Gao X, Wu Y, Wu Q, Xu X, et al: DHW-221, a dual PI3K/mTOR
inhibitor, overcomes multidrug resistance by targeting
P-glycoprotein (P-gp/ABCB1) and Akt-mediated FOXO3a nuclear
translocation in non-small cell lung cancer. Front Oncol.
12:8736492022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu T, Wei R, Zhang Y, Chen W and Liu H:
Association between NF-κB expression and drug resistance of liver
cancer. Oncol Lett. 17:1030–1034. 2019.PubMed/NCBI
|
|
17
|
Liu Y, Cao F, Xia F, Li J, Dong X, Guo Y,
Zhang J, Zhao Q and Liu Y: Shc3 facilitates breast cancer drug
resistance by interacting with ErbB2 to initiate ErbB2/COX2/MDR1
axis. Cancer Med. 12:10768–10780. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jeddi F, Soozangar N, Sadeghi MR, Somi MH,
Shirmohamadi M, Eftekhar-Sadat AT and Samadi N: Nrf2 overexpression
is associated with P-glycoprotein upregulation in gastric cancer.
Biomed Pharmacother. 97:286–292. 2018. View Article : Google Scholar
|
|
19
|
Chen Q, Liu X, Luo Z, Wang S, Lin J, Xie
Z, Li M, Li C, Cao H, Huang Q, et al: Chloride channel-3 mediates
multidrug resistance of cancer by upregulating P-glycoprotein
expression. J Cell Physiol. 234:6611–6623. 2019. View Article : Google Scholar
|
|
20
|
Mai L, Luo M, Wu JJ, Yang JH and Hong LY:
The combination therapy of HIF1α inhibitor LW6 and cisplatin plays
an effective role on anti-tumor function in A549 cells. Neoplasma.
66:776–784. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ren H, Wang Z, Chen Y, Liu Y, Zhang S,
Zhang T and Li Y: SMYD2-OE promotes oxaliplatin resistance in colon
cancer through MDR1/P-glycoprotein via MEK/ERK/AP1 pathway. Onco
Targets Ther. 12:2585–2594. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamamoto M, Suzuki S, Togashi K, Sanomachi
T, Seino S, Kitanaka C and Okada M: AS602801 sensitizes ovarian
cancer stem cells to paclitaxel by down-regulating MDR1. Anticancer
Res. 39:609–617. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gao J, Hou D, Hu P and Mao G: Curcumol
increases the sensitivity of colon cancer to 5-FU by regulating
Wnt/β-catenin signaling. Transl Cancer Res. 10:2437–2450. 2021.
View Article : Google Scholar
|
|
24
|
Li M, Li ZH, Song J, Li X, Zhai P, Mu X,
Qiu F and Yao L: miR-205 Reverses MDR-1 mediated doxorubicin
resistance via PTEN in human liver cancer HepG2 cells. Cell J.
24:112–119. 2022.PubMed/NCBI
|
|
25
|
Zhang L, Li Y, Hu C, Chen Y, Chen Z, Chen
ZS, Zhang JY and Fang S: CDK6-PI3K signaling axis is an efficient
target for attenuating ABCB1/P-gp mediated multi-drug resistance
(MDR) in cancer cells. Mol Cancer. 21:1032022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wan Y, Chen Z, Wang Y, Zhao W, Pei Z, Pu
L, Lv Y, Li J, Li J and Pei Y: A hyaluronic acid modified cuprous
metal-organic complex for reversing multidrug resistance via redox
dyshomeostasis. Carbohydr Polym. 311:1207622023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Abou-Alfa GK: Selection of patients with
hepatocellular carcinoma for sorafenib. J Natl Compr Canc Netw.
7:397–403. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xia X, Wang Q, Ye T, Liu Y, Liu D, Song S
and Zheng C: NRF2/ABCB1-mediated efflux and PARP1-mediated
dampening of DNA damage contribute to doxorubicin resistance in
chronic hypoxic HepG2 cells. Fundam Clin Pharmacol. 34:41–50. 2020.
View Article : Google Scholar
|
|
30
|
Yuan P, Zheng A and Tang Q: Tripartite
motif protein 25 is associated with epirubicin resistance in
hepatocellular carcinoma cells via regulating PTEN/AKT pathway.
Cell Biol Int. 44:1503–1513. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mou SJ, Yang PF, Liu YP, Xu N, Jiang WW
and Yue WJ: BCLAF1 promotes cell proliferation, invasion and
drug-resistance though targeting lncRNA NEAT1 in hepatocellular
carcinoma. Life Sci. 242:1171772020. View Article : Google Scholar
|
|
32
|
Fang Z, Chen W, Yuan Z, Liu X and Jiang H:
LncRNA-MALAT1 contributes to the cisplatin-resistance of lung
cancer by upregulating MRP1 and MDR1 via STAT3 activation. Biomed
Pharmacother. 101:536–542. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao L, Bin S, He HL, Yang JM, Pu YC, Gao
CH, Wang H and Wang BL: Sodium butyrate increases P-gp expression
in lung cancer by upregulation of STAT3 and mRNA stabilization of
ABCB1. Anticancer Drugs. 29:227–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu J, Hu W, Zhang Y, Du P, Xiao W and Li
Y: Comparison of survival outcomes of chemotherapy plus surgery vs
chemotherapy alone for patients with isolated liver metastases from
gallbladder carcinoma. Am Surg. 89:676–684. 2023. View Article : Google Scholar
|
|
35
|
Wu D, Liu L, Yan X, Wang C, Wang Y, Han K,
Lin S, Gan Z and Min D: Pleiotrophin promotes chemoresistance to
doxorubicin in osteosarcoma by upregulating P-glycoprotein.
Oncotarget. 8:63857–63870. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang JZ, Huang LH, Chen R, Meng LJ, Gao
YY, Ji QY and Wang Y: LIM kinase 1 serves an important role in the
multidrug resistance of osteosarcoma cells. Oncol Lett. 15:250–256.
2018.PubMed/NCBI
|
|
37
|
Liu S, Chen S, Yuan W, Wang H, Chen K and
Li D and Li D: PD-1/PD-L1 interaction up-regulates MDR1/P-gp
expression in breast cancer cells via PI3K/AKT and MAPK/ERK
pathways. Oncotarget. 8:99901–99912. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen S, Wang H, Li Z, You J, Wu QW, Zhao
C, Tzeng CM and Zhang ZM: Interaction of WBP2 with ERα increases
doxorubicin resistance of breast cancer cells by modulating MDR1
transcription. Br J Cancer. 119:182–192. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lim JS, Jung GY and Park SY: Nkx-2.5
regulates MDR1 expression via its upstream promoter in breast
cancer cells. J Korean Med Sci. 34:e1002019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Baltes F, Pfeifer V, Silbermann K, Caspers
J, Wantoch von Rekowski K, Schlesinger M and Bendas G:
β1-Integrin binding to collagen type 1 transmits breast
cancer cells into chemoresistance by activating ABC efflux
transporters. Biochim Biophys Acta Mol Cell Res. 1867:1186632020.
View Article : Google Scholar
|
|
41
|
Li G, Guo J, Shen BQ, Yadav DB, Sliwkowski
MX, Crocker LM, Lacap JA and Phillips GDL: Mechanisms of acquired
resistance to trastuzumab emtansine in breast cancer cells. Mol
Cancer Ther. 17:1441–1453. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tang H, Zeng L, Wang J, Zhang X, Ruan Q,
Wang J, Cui S and Yang D: Reversal of 5-fluorouracil resistance by
EGCG is mediate by inactivation of TFAP2A/VEGF signaling pathway
and down-regulation of MDR-1 and P-gp expression in gastric cancer.
Oncotarget. 8:82842–82853. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Deng XJ, Zheng HL, Ke XQ, Deng M, Ma ZZ,
Zhu Y and Cui YY: Hsa-miR-34a-5p reverses multidrug resistance in
gastric cancer cells by targeting the 3′-UTR of SIRT1 and
inhibiting its expression. Cell Signal. 84:1100162021. View Article : Google Scholar
|
|
44
|
Xu YD, Shang J, Li M and Zhang YY: LncRNA
DANCR accelerates the development of multidrug resistance of
gastric cancer. Eur Rev Med Pharmacol Sci. 23:2794–2802.
2019.PubMed/NCBI
|
|
45
|
Zhang X, Bo P, Liu L, Zhang X and Li J:
Overexpression of long non-coding RNA GHET1 promotes the
development of multidrug resistance in gastric cancer cells. Biomed
Pharmacother. 92:580–585. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tan B, Li Y, Zhao Q, Fan L and Wang D:
ZNF139 increases multidrug resistance in gastric cancer cells by
inhibiting miR-185. Biosci Rep. 38:BSR201810232018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yan X, Zhao J and Zhang R: Visfatin
mediates doxorubicin resistance in human colorectal cancer cells
via up regulation of multidrug resistance 1 (MDR1). Cancer
Chemother Pharmacol. 80:395–403. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gao Z, Li Z, Liu Y and Liu Z: Forkhead box
O3 promotes colon cancer proliferation and drug resistance by
activating MDR1 expression. Mol Genet Genomic Med. 7:e5542019.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Du J, He Y, Li P, Wu W, Chen Y and Ruan H:
IL-8 regulates the doxorubicin resistance of colorectal cancer
cells via modulation of multidrug resistance 1 (MDR1). Cancer
Chemother Pharmacol. 81:1111–1119. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang S, Chatterjee T, Godoy C, Wu L, Liu
QJ and Carmon KS: GPR56 drives colorectal tumor growth and promotes
drug resistance through upregulation of MDR1 expression via a
RhoA-mediated mechanism. Mol Cancer Res. 17:2196–2207. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sui M, Yang H, Guo M, Li W, Gong Z, Jiang
J and Li P: Cajanol sensitizes A2780/Taxol cells to paclitaxel by
inhibiting the PI3K/Akt/NF-κB signaling pathway. Front Pharmacol.
12:7833172021. View Article : Google Scholar
|
|
52
|
Zhao P, Wang S, Jiang J, Liu H, Zhu X,
Zhao N, Li J, Yin Y, Pan X, Yang X, et al: TIPE2 sensitizes
osteosarcoma cells to cis-platin by down-regulating MDR1 via the
TAK1-NF-κB and-AP-1 pathways. Mol Immunol. 101:471–478. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Song W, Liang C, Sun Y, Morii S, Yomogida
S, Isaji T, Fukuda T, Hang Q, Hara A, Nakano M and Gu J: Expression
of GnT-III decreases chemoresistance via negatively regulating
P-glycoprotein expression: Involvement of the TNFR2-NF-κB signaling
pathway. J Biol Chem. 299:1030512023. View Article : Google Scholar
|
|
54
|
Liu K, Song J, Yan Y, Zou K, Che Y, Wang
B, Li Z, Yu W, Guo W, Zou L, et al: Melatonin increases the
chemosensitivity of diffuse large B-cell lymphoma cells to
epirubicin by inhibiting P-glycoprotein expression via the NF-κB
pathway. Transl Oncol. 14:1008762021. View Article : Google Scholar
|
|
55
|
Liu RM, Xu P, Chen Q, Feng SL and Xie Y: A
multiple-targets alkaloid nuciferine overcomes paclitaxel-induced
drug resistance in vitro and in vivo. Phytomedicine. 79:1533422020.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang H, Jia XH, Chen JR, Wang JY and Li
YJ: Osthole shows the potential to overcome P-glycoprotein-mediated
multidrug resistance in human myelogenous leukemia K562/ADM cells
by inhibiting the PI3K/Akt signaling pathway. Oncol Rep.
35:3659–3668. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Satonaka H, Ishida K, Takai M, Koide R,
Shigemasa R, Ueyama J, Ishikawa T, Hayashi K, Goto H and Wakusawa
S: (-)-Epigallocatechin-3-gallate down-regulates
doxorubicin-induced overexpression of P-glycoprotein through the
coordinate inhibition of PI3K/Akt and MEK/ERK signaling pathways.
Anticancer Res. 37:6071–6077. 2017.PubMed/NCBI
|
|
58
|
Ding Y, Zhen Z, Nisar MA, Ali F, Din RU,
Khan M, Mughal TA, Alam G, Liu L and Saleem MZ: Sesquiterpene
lactones attenuate paclitaxel resistance via inhibiting
MALAT1/STAT3/FUT4 Axis and P-glycoprotein transporters in lung
cancer cells. Front Pharmacol. 13:7956132022. View Article : Google Scholar
|
|
59
|
Wang PP, Luan JJ, Xu WK, Wang L, Xu DJ,
Yang CY, Zhu YH and Wang YQ: Astragaloside IV downregulates the
expression of MDR1 in Bel-7402/FU human hepatic cancer cells by
inhibiting the JNK/c-Jun/AP-1 signaling pathway. Mol Med Rep.
16:2761–2766. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Qin T, Zhu W, Kan X, Li L and Wu D:
Luteolin attenuates the chemoresistance of osteosarcoma through
inhibiting the PTN/β-catenin/MDR1 signaling axis by upregulating
miR-384. J Bone Oncol. 34:100429–34. 2022. View Article : Google Scholar
|
|
61
|
Li L, Fu Q, Xia M, Xin L, Shen H, Li G, Ji
G, Meng Q and Xie Y: Inhibition of P-glycoprotein mediated efflux
in Caco-2 cells by phytic acid. J Agric Food Chem. 66:988–998.
2018. View Article : Google Scholar
|
|
62
|
Mirzaei SA, Reiisi S, Ghiasi Tabari P,
Shekari A, Aliakbari F, Azadfallah E and Elahian F: Broad blocking
of MDR efflux pumps by acetylshikonin and acetoxyisovalerylshikonin
to generate hypersensitive phenotype of malignant carcinoma cells.
Sci Rep. 8:34462018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shen X, Chen G, Zhu G and Fong WF:
(+/−)-3′-O, 4′-O-dicynnamoyl-cis-khellactone, a derivative of
(+/−)-praeruptorin A, reverses P-glycoprotein mediated multidrug
resistance in cancer cells. Bioorg Med Chem. 14:7138–7145. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Bai Z, Zhou Q, Zhu H, Ye X, Wu P and Ma L:
QTMP, a novel thiourea polymer, causes DNA damage to exert
anticancer activity and overcome multidrug resistance in colorectal
cancer cells. Front Oncol. 11:6676892021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gao M, Liu T, Li J, Guan Q, Wang H, Yan S,
Li Z, Zuo D, Zhang W and Wu Y: YAN, a novel microtubule inhibitor,
inhibits P-gp and MRP1 function and induces mitotic slippage
followed by apoptosis in multidrug-resistant A549/Taxol cells.
Toxicol In Vitro. 69:1049712020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wu CP, Murakami M, Wu YS, Chi YC, Hsiao
SH, Huang YH, Hung TH and Ambudkar SV: Branebrutinib (BMS-986195),
a bruton's tyrosine kinase inhibitor, resensitizes
P-glycoprotein-overexpressing multidrug-resistant cancer cells to
chemotherapeutic agents. Front Cell Dev Biol. 9:6995712021.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Podolski-Renić A, Dinić J, Stanković T,
Tsakovska I, Pajeva I, Tuccinardi T, Botta L, Schenone S and Pešić
M: New therapeutic strategy for overcoming multidrug resistance in
cancer cells with pyrazolo[3,4-d]pyrimidine tyrosine kinase
inhibitors. Cancers (Basel). 13:53082021. View Article : Google Scholar
|
|
68
|
Wu CP, Hung TH, Lusvarghi S, Chu YH, Hsiao
SH, Huang YH, Chang YT and Ambudkar SV: The third-generation EGFR
inhibitor almonertinib (HS-10296) resensitizes ABCB1-overexpressing
multidrug-resistant cancer cells to chemotherapeutic drugs. Biochem
Pharmacol. 188:1145162021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang L, Ye B, Lin Y, Li YD, Wang JQ, Chen
Z, Ping FF and Chen ZS: Ribociclib inhibits P-gp-mediated multidrug
resistance in human epidermoid carcinoma cells. Front Pharmacol.
13:8671282022. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Laiolo J, Barbieri CL, Joray MB, Lanza PA,
Palacios SM, Vera DMA and Carpinella MC: Plant extracts and betulin
from Ligaria cuneifolia inhibit P-glycoprotein function in leukemia
cells. Food Chem Toxicol. 147:1119222021. View Article : Google Scholar
|
|
71
|
Liu Z, Wen X, Wang G and Zhou Y:
Involvement of P-gp on reversing multidrug resistance effects of
23-hydroxybetulinic acid on chemotherapeutic agents. Front
Pharmacol. 12:7967452021. View Article : Google Scholar :
|
|
72
|
Yang T, Wang S, Li H, Zhao Q, Yan S, Dong
M, Liu D, Chen X and Li R: Lathyrane diterpenes from Euphorbia
lathyris and the potential mechanism to reverse the multi-drug
resistance in HepG2/ADR cells. Biomed Pharmacother. 121:1096632020.
View Article : Google Scholar
|
|
73
|
Qian J, Xia M, Liu W, Li L, Yang J, Mei Y,
Meng Q and Xie Y: Glabridin resensitizes
p-glycoprotein-overexpressing multidrug-resistant cancer cells to
conventional chemotherapeutic agents. Eur J Pharmacol. 852:231–243.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Teng YN, Lin KI, Lin YC, Thang TD, Lan YH
and Hung CC: A novel flavonoid from Fissistigma cupreonitens,
5-hydroxy-7,8-dimethoxyflavanone, competitively inhibited the
efflux function of human P-glycoprotein and reversed cancer
multi-drug resistance. Phytomedicine. 85:1535282021. View Article : Google Scholar
|
|
75
|
Min H, Niu M, Zhang W, Yan J, Li J, Tan X,
Li B, Su M, Di B and Yan F: Emodin reverses leukemia multidrug
resistance by competitive inhibition and downregulation of
P-glycoprotein. PLoS One. 12:e01879712017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Pellegrini E, Multari G, Gallo FR,
Vecchiotti D, Zazzeroni F, Condello M and Meschini S: A natural
product, voacamine, sensitizes paclitaxel-resistant human ovarian
cancer cells. Toxicol Appl Pharmacol. 434:1158162022. View Article : Google Scholar
|
|
77
|
Sagnou M, Novikov FN, Ivanova ES, Alexiou
P, Stroylov VS, Titov IY, Tatarskiy VV, Vagida MS, Pelecanou M,
Shtil AA and Chilov GG: Novel curcumin derivatives as
P-glycoprotein inhibitors: Molecular modeling, synthesis and
sensitization of multidrug resistant cells to doxorubicin. Eur J
Med Chem. 198:1123312020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Creemers SG, van Koetsveld PM, De Herder
WW, Dogan F, Franssen GJH, Feelders RA and Hofland LJ: MDR1
inhibition increases sensitivity to doxorubicin and etoposide in
adrenocortical cancer. Endocr Relat Cancer. 26:367–378. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wang SQ, Teng QX, Wang S, Lei ZN, Hu HH,
Lv HF, Chen BB, Wang JZ, Shi XJ, Xu WF, et al: Preclinical studies
of the triazolo[1,5-a]pyrimidine derivative WS-716 as a highly
potent, specific and orally active P-glycoprotein (P-gp) inhibitor.
Acta Pharm Sin B. 12:3263–3280. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Dei S, Coronnello M, Bartolucci G, Manetti
D, Romanelli MN, Udomtanakunchai C, Salerno M and Teodori E: Design
and synthesis of new potent N,N-bis(arylalkyl)piperazine
derivatives as multidrug resistance (MDR) reversing agents. Eur J
Med Chem. 147:7–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Qiu Q, Zou F, Li H, Shi W, Zhou D, Zhang
P, Li T, Yin Z, Cai Z, Jiang Y, et al: Structure-based discovery of
pyrimidine aminobenzene derivatives as potent oral reversal agents
against P-gp- and BCRP-mediated multidrug resistance. J Med Chem.
64:6179–6197. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Li YS, Yang X, Zhao DS, Cai Y, Huang Z, Wu
R, Wang SJ, Liu GJ, Wang J, Bao XZ, et al: Design, synthesis and
bioactivity study on 5-phenylfuran derivatives as potent reversal
agents against P-glycoprotein-mediated multidrug resistance in
MCF-7/ADR cell. Eur J Med Chem. 216:1133362021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Yuan S, Wang B, Dai QQ, Zhang XN, Zhang
JY, Zuo JH, Liu H, Chen ZS, Li GB, Wang S, et al: Discovery of new
4-indolyl quinazoline derivatives as highly potent and orally
bioavailable P-glycoprotein inhibitors. J Med Chem. 64:14895–14911.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wang H, Wang X, Liao A and Liu Z:
Hypomethylation agent decitabine restores drug sensitivity by
depressing P-glycoprotein activity through MAPK signaling pathway.
Mol Cell Biochem. 433:141–148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Snyder S, Murundi S, Crawford L and Putnam
D: Enabling P-glycoprotein inhibition in multidrug resistant cancer
through the reverse targeting of a quinidine-PEG conjugate. J
Control Release. 317:291–299. 2020. View Article : Google Scholar
|
|
86
|
Zhang H, Bian S, Xu Z, Gao M, Wang H,
Zhang J, Zhang M, Ke Y, Wang W, Chen ZS and Xu H: The effect and
mechanistic study of encequidar on reversing the resistance of
SW620/AD300 cells to doxorubicin. Biochem Pharmacol.
205:1152582022. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Xu SW, Law BYK, Qu SLQ, Hamdoun S, Chen J,
Zhang W, Guo JR, Wu AG, Mok SWF, Zhang DW, et al: SERCA and
P-glycoprotein inhibition and ATP depletion are necessary for
celastrol-induced autophagic cell death and collateral sensitivity
in multidrug-resistant tumor cells. Pharmacol Res. 153:1046602020.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Rauf A, Uddin G, Raza M, Ahmad B, Jehan N,
Siddiqui BS, Molnar J, Csonka A and Szabo D: Reversal of multidrug
resistance in mouse lymphoma cells by extracts and flavonoids from
Pistacia integerrima. Asian Pac J Cancer Prev. 17:51–55. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Giorgi C, Gasser UE, Lafont ME, Holzgang
HE and Thalhammer VF: Inhibition of chemoresistance in primary
tumor cells by Camellia sinensis non fermentatum extract
noviphenone (NPE®). Anticancer Res. 39:4101–4110. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kawami M, Yamada Y, Issarachot O,
Junyaprasert VB, Yumoto R and Takano M: P-gp modulating effect of
Azadirachta indica extract in multidrug-resistant cancer cell
lines. Pharmazie. 73:104–109. 2018.PubMed/NCBI
|
|
91
|
Zheng Y, Su C, Zhao L and Shi Y: mAb
MDR1-modified chitosan nanoparticles overcome acquired EGFR-TKI
resistance through two potential therapeutic targets modulation of
MDR1 and autophagy. J Nanobiotechnology. 15:662017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Chen Z, Pan T, Jiang D, Jin L, Geng Y,
Feng X, Shen A and Zhang L: The lncRNA-GAS5/miR-221-3p/DKK2 axis
modulates ABCB1-mediated adriamycin resistance of breast cancer via
the Wnt/β-catenin signaling pathway. Mol Ther Nucleic Acids.
19:1434–1448. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Han Z and Shi L: Long non-coding RNA
LUCAT1 modulates methotrexate resistance in osteosarcoma via
miR-200c/ABCB1 axis. Biochem Biophys Res Commun. 495:947–953. 2018.
View Article : Google Scholar
|
|
94
|
Kang Y, Zhang S, Cao W, Wan D and Sun L:
Knockdown of LncRNA CRNDE suppresses proliferation and
P-glycoprotein-mediated multidrug resistance in acute myelocytic
leukemia through the Wnt/β-catenin pathway. Biosci Rep.
40:BSR201934502020. View Article : Google Scholar
|
|
95
|
Zou H and Li H: Knockdown of long
non-coding RNA LINC00152 increases cisplatin sensitivity in ovarian
cancer cells. Exp Ther Med. 18:4510–4516. 2019.PubMed/NCBI
|
|
96
|
Zou Z, Zou R, Zong D, Shi Y, Chen J, Huang
J, Zhu J, Chen L, Bao X, Liu Y, et al: miR-495 sensitizes MDR
cancer cells to the combination of doxorubicin and taxol by
inhibiting MDR1 expression. J Cell Mol Med. 21:1929–1943. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Zhao Y, Qi X, Chen J, Wei W, Yu C, Yan H,
Pu M, Li Y, Miao L, Li C and Ren J: The miR-491-3p/Sp3/ABCB1 axis
attenuates multidrug resistance of hepatocellular carcinoma. Cancer
Lett. 408:102–111. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Yi D, Xu L, Wang R, Lu X and Sang J:
miR-381 overcomes cisplatin resistance in breast cancer by
targeting MDR1. Cell Biol Int. 43:12–21. 2019. View Article : Google Scholar
|
|
99
|
Zheng SZ, Sun P, Wang JP, Liu Y, Gong W
and Liu J: MiR-34a overexpression enhances the inhibitory effect of
doxorubicin on HepG2 cells. World J Gastroenterol. 25:2752–2762.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Wei S, Gao J, Zhang M, Dou Z, Li W and
Zhao L: Dual delivery nanoscale device for miR-451 and adriamycin
co-delivery to combat multidrug resistant in bladder cancer. Biomed
Pharmacother. 122:1094732020. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Kenworthy R, Bosco DB, DeLigio JT and
Zorio DAR: Micro-RNA149 confers taxane resistance to malignant
mesothelioma cells via regulation of P-glycoprotein expression.
Cancer Biol Ther. 19:181–187. 2018. View Article : Google Scholar :
|
|
102
|
Wang S, Guo J, Mo Z, Shi X and Qu C:
Clinical significance and correlation of miR-200c and P-gp
expression in gastric cancer and the effects on multidrug
resistance. J Gastrointest Oncol. 13:581–592. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Cao F and Yin LX: miR-122 enhances
sensitivity of hepatocellular carcinoma to oxaliplatin via
inhibiting MDR1 by targeting Wnt/β-catenin pathway. Exp Mol Pathol.
106:34–43. 2019. View Article : Google Scholar
|
|
104
|
Sousa D, Matthiesen R, Lima RT and
Vasconcelos MH: Deep sequencing analysis reveals distinctive
non-coding RNAs when comparing tumor multidrug-resistant cells and
extracellular vesicles with drug-sensitive counterparts. Cancers
(Basel). 12:2002020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Shi X, Valizadeh A, Mir SM, Asemi Z,
Karimian A, Majidina M, Safa A and Yosefi B: miRNA-29a reverses
P-glycoprotein-mediated drug resistance and inhibits proliferation
via up-regulation of PTEN in colon cancer cells. Eur J Pharmacol.
880:1731382020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Xie B, Li L, Zhang Z, Zhao L, Cheng J,
Zhou C, Cheng J, Yan J, Chen J, Yi J, et al: MicroRNA-1246 by
targeting AXIN2 and GSK-3β overcomes drug resistance and induces
apoptosis in chemo-resistant leukemia cells. J Cancer.
12:4196–4208. 2021. View Article : Google Scholar :
|
|
107
|
Kara G, Calin GA and Ozpolat B: RNAi-based
therapeutics and tumor targeted delivery in cancer. Adv Drug Deliv
Rev. 182:1141132022. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Yalamarty SSK, Filipczak N, Li X,
Pathrikar TV, Cotter C and Torchilin VP: Co-delivery of siRNA and
chemotherapeutic drug using 2C5 antibody-targeted dendrimer-based
mixed micelles for multidrug resistant cancers. Pharmaceutics.
14:14702022. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Wang C, Guan W, Peng J, Chen Y, Xu G and
Dou H: Gene/paclitaxel co-delivering nanocarriers prepared by
framework-induced self-assembly for the inhibition of highly
drug-resistant tumors. Acta Biomater. 103:247–258. 2020. View Article : Google Scholar
|
|
110
|
Jiang Y, Liu Y, Wang M, Li Z, Su L, Xu X,
Xing C, Li J, Lin L, Lu C and Yang H: siRNA-based carrier-free
system for synergistic chemo/chemodynamic/RNAi therapy of
drug-resistant tumors. ACS Appl Mater Interfaces. 14:361–372. 2022.
View Article : Google Scholar
|
|
111
|
Yang X, Wang Y, Chen S, Zhang S and Cui C:
Cetuximab-modified human serum albumin nanoparticles co-loaded with
doxorubicin and MDR1 siRNA for the treatment of drug-resistant
breast tumors. Int J Nanomedicine. 16:7051–7069. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Liu H, Ma D, Chen J, Ye L, Li Y, Xie Y,
Zhao X, Zou H, Chen X, Pu J and Liu P: A targeted nanoplatform
co-delivery of pooled siRNA and doxorubicin for reversing of
multidrug resistance in breast cancer. Nano Res. 15:6306–6314.
2022. View Article : Google Scholar
|
|
113
|
Wu J, Wang Q, Dong X, Xu M, Yang J, Yi X,
Chen B, Dong X, Wang Y, Lou X, et al: Biocompatible
AIEgen/p-glycoprotein siRNA@reduction-sensitive paclitaxel
polymeric prodrug nanoparticles for overcoming chemotherapy
resistance in ovarian cancer. Theranostics. 11:3710–3724. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Zhang J, Du Z, Pan S, Shi M, Li J, Yang C,
Hu H, Qiao M, Chen D and Zhao X: Overcoming multidrug resistance by
codelivery of MDR1-targeting siRNA and doxorubicin using
EphA10-mediated pH-sensitive lipoplexes: In vitro and in vivo
evaluation. ACS Appl Mater Interfaces. 10:21590–21600. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Cao X, Li R, Xiong H, Su J, Guo C, An T,
Zong H and Zhao R: Novel Pt(IV) complexes to overcome multidrug
resistance in gastric cancer by targeting P-glycoprotein. Eur J Med
Chem. 221:1135202021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Liu ZL, Jin BJ, Cheng CG, Zhang FX, Wang
SW, Wang Y and Wu B: Apatinib resensitizes cisplatin-resistant
non-small cell lung carcinoma A549 cell through reversing multidrug
resistance and suppressing ERK signaling pathway. Eur Rev Med
Pharmacol Sci. 21:5370–5377. 2017.PubMed/NCBI
|
|
117
|
Ji L, Liu X, Zhang S, Tang S, Yang S, Li
S, Qi X, Yu S, Lu L, Meng X and Liu Z: The novel
triazolonaphthalimide derivative LSS-11 synergizes the
anti-proliferative effect of paclitaxel via STAT3-dependent MDR1
and MRP1 downregulation in chemoresistant lung cancer cells.
Molecules. 22:18222017. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Zhang L, Li Y, Wang Q, Chen Z, Li X, Wu Z,
Hu C, Liao D, Zhang W and Chen ZS: The PI3K subunits, P110α and
P110β are potential targets for overcoming P-gp and BCRP-mediated
MDR in cancer. Mol Cancer. 19:102020. View Article : Google Scholar
|
|
119
|
Liu T, Peng Y, Li X, Liu L, Liu F and He
L: A novel delocalized lipophilic cation-chlorambucil conjugate
inhibits P-glycoprotein in HepG2/ADM cells. Bioorg Med Chem.
25:5461–5467. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Ganesan M, Kanimozhi G, Pradhapsingh B,
Khan HA, Alhomida AS, Ekhzaimy A, Brindha GR and Prasad NR:
Phytochemicals reverse P-glycoprotein mediated multidrug resistance
via signal transduction pathways. Biomed Pharmacother.
139:1116322021. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Li Y, Li X, Lu Y, Chaurasiya B, Mi G, Shi
D, Chen D, Webster TJ, Tu J and Shen Y: Co-delivery of Poria cocos
extract and doxorubicin as an 'all-in-one' nanocarrier to combat
breast cancer multidrug resistance during chemotherapy.
Nanomedicine. 23:1020952020. View Article : Google Scholar
|
|
122
|
Ye Q, Liu K, Shen Q, Li Q, Hao J, Han F
and Jiang RW: Reversal of multidrug resistance in cancer by
multi-functional flavonoids. Front Oncol. 9:4872019. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Fang Y, Liang F, Xia M, Cao W, Pan S, Wu T
and Xu X: Structure-activity relationship and mechanism of
flavonoids on the inhibitory activity of P-glycoprotein
(P-gp)-mediated transport of rhodamine123 and daunorubicin in P-gp
overexpressed human mouth epidermal carcinoma (KB/MDR) cells. Food
Chem Toxicol. 155:1123812021. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Kong W, Ling X, Chen Y, Wu X, Zhao Z, Wang
W, Wang S, Lai G and Yu Z: Hesperetin reverses
P-glycoprotein-mediated cisplatin resistance in DDP-resistant human
lung cancer cells via modulation of the nuclear factor-κB signaling
pathway. Int J Mol Med. 45:1213–1224. 2020.PubMed/NCBI
|
|
125
|
La X, Zhang L, Li Z, Li H and Yang Y:
(-)-Epigallocatechin gallate (EGCG) enhances the sensitivity of
colorectal cancer cells to 5-FU by inhibiting
GRP78/NF-κB/miR-155-5p/MDR1 pathway. J Agric Food Chem.
67:2510–2518. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Mohana S, Ganesan M, Rajendra Prasad N,
Ananthakrishnan D and Velmurugan D: Flavonoids modulate multidrug
resistance through wnt signaling in P-glycoprotein overexpressing
cell lines. BMC Cancer. 18:11682018. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Wang ZD, Wang RZ, Xia YZ, Kong LY and Yang
L: Reversal of multidrug resistance by icaritin in
doxorubicin-resistant human osteosarcoma cells. Chin J Nat Med.
16:20–28. 2018.PubMed/NCBI
|
|
128
|
Wang F, Huang N, Yang Q, Liu J and Chen J:
Reversal of doxorubicin-resistance by Salvia miltiorrhiza
ligustrazine in the SHG44/doxorubicin glioma drug-resistant cell
line. Oncol Lett. 14:4708–4714. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Zhou P, Zhang R, Wang Y, Xu D, Zhang L,
Qin J, Su G, Feng Y, Chen H, You S, et al: Cepharanthine
hydrochloride reverses the mdr1 (P-glycoprotein)-mediated
esophageal squamous cell carcinoma cell cisplatin resistance
through JNK and p53 signals. Oncotarget. 8:111144–111160. 2017.
View Article : Google Scholar
|
|
130
|
Zhou X, Jin N and Chen B: Tetrandrine
overcomes drug resistance mediated by bone marrow microenvironment
by regulating the expression of P-glycoprotein in acute leukemia.
Hematology. 27:274–279. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Qian K, Tang CY, Chen LY, Zheng S, Zhao Y,
Ma LS, Xu L, Fan LH, Yu JD, Tan HS, et al: Berberine reverses
breast cancer multidrug resistance based on fluorescence
pharmacokinetics in vitro and in vivo. ACS Omega. 6:10645–10654.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Huang CZ, Wang YF, Zhang Y, Peng YM, Liu
YX, Ma F, Jiang JH and Wang QD: Cepharanthine hydrochloride
reverses P-glycoprotein-mediated multidrug resistance in human
ovarian carcinoma A2780/Taxol cells by inhibiting the PI3K/Akt
signaling pathway. Oncol Rep. 38:2558–2564. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Liao D, Zhang W, Gupta P, Lei ZN, Wang JQ,
Cai CY, Vera AA, Zhang L, Chen ZS and Yang DH: Tetrandrine
interaction with ABCB1 reverses multidrug resistance in cancer
cells through competition with anti-cancer drugs followed by
downregulation of ABCB1 expression. Molecules. 24:43832019.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Quarti J, Torres DNM, Ferreira E, Vidal
RS, Casanova F, Chiarini LB, Fialho E and Rumjanek VM: Selective
cytotoxicity of piperine over multidrug resistance leukemic cells.
Molecules. 26:9342021. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Wang SQ, Li X, Wang XN and Lou HX:
Reversal of P-glycoprotein-mediated multidrug resistance in human
leukemia cell line K562/A02 by archangelicin. Chin J Nat Med.
9:146–150. 2011.
|
|
136
|
Sharma P, Tyagi A, Bhansali P, Pareek S,
Singh V, Ilyas A, Mishra R and Poddar NK: Saponins: Extraction,
bio-medicinal properties and way forward to anti-viral
representatives. Food Chem Toxicol. 150:1120752021. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Li Y, Sun Y, Tang T, Niu Y, Li X, Xie M,
Jin H and Mei Q: Paris saponin VII reverses chemoresistance in
breast MCF-7/ADR cells. J Ethnopharmacol. 232:47–54. 2019.
View Article : Google Scholar
|
|
138
|
Niu X, Shi Y, Li Q, Chen H, Fan X, Yu Y,
Lv C and Lu J: Ginsenoside Rb1 for overcoming
cisplatin-insensitivity of A549/DDP cells in vitro and vivo through
the dual-inhibition on two efflux pumps of ABCB1 and PTCH1.
Phytomedicine. 115:1547762023. View Article : Google Scholar
|
|
139
|
Li J, Zheng L, Wang R, Sun D, Liang S, Wu
J, Liu Y, Tian X, Li T, Yang Y and Han L: Synergistic combination
of sodium aescinate-stabilized, polymer-free, twin-like
nanoparticles to reverse paclitaxel resistance. Int J Nanomedicine.
15:5839–5853. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Huang C, Xu D, Xia Q, Wang P, Rong C and
Su Y: Reversal of P-glycoprotein-mediated multidrug resistance of
human hepatic cancer cells by astragaloside II. J Pharm Pharmacol.
64:1741–1750. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Wang PP, Xu DJ, Huang C, Wang WP and Xu
WK: Astragaloside IV reduces the expression level of P-glycoprotein
in multidrug-resistant human hepatic cancer cell lines. Mol Med
Rep. 9:2131–2137. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Liu GW, Liu YH, Jiang GS and Ren WD: The
reversal effect of ginsenoside Rh2 on drug resistance in human
colorectal carcinoma cells and its mechanism. Hum Cell. 31:189–198.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Liu C, Gong Q, Chen T, Lv J, Feng Z, Liu P
and Deng Z: Treatment with 20(S)-ginsenoside Rg3 reverses multidrug
resistance in A549/DDP xenograft tumors. Oncol Lett. 15:4376–4382.
2018.PubMed/NCBI
|
|
144
|
Sadava D and Kane SE: Silibinin reverses
drug resistance in human small-cell lung carcinoma cells. Cancer
Lett. 339:102–106. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Dobiasová S, Řehořová K, Kučerová D,
Biedermann D, Káňová K, Petrásková L, Koucká K, Václavíková R,
Valentová K, Ruml T, et al: Multidrug resistance modulation
activity of silybin derivatives and their anti-inflammatory
potential. Antioxidants (Basel). 9:4552020. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Zhang ZL, Jiang QC and Wang SR:
Schisandrin A reverses doxorubicin-resistant human breast cancer
cell line by the inhibition of P65 and Stat3 phosphorylation.
Breast Cancer. 25:233–242. 2018. View Article : Google Scholar
|
|
147
|
Liao XZ, Gao Y, Sun LL, Liu JH, Chen HR,
Yu L, Chen ZZ, Chen WH and Lin LZ: Rosmarinic acid reverses
non-small cell lung cancer cisplatin resistance by activating the
MAPK signaling pathway. Phytother Res. 34:1142–1153. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Muthusamy G, Gunaseelan S and Prasad NR:
Ferulic acid reverses P-glycoprotein-mediated multidrug resistance
via inhibition of PI3K/Akt/NF-κB signaling pathway. J Nutr Biochem.
63:62–71. 2019. View Article : Google Scholar
|
|
149
|
Oh SS, Lee KW, Madhi H, Jeong JW, Park S,
Kim M, Lee Y, Han HT, Hwangbo C, Yoo J and Kim KD: Cordycepin
resensitizes T24R2 cisplatin-resistant human bladder cancer cells
to cisplatin by inactivating Ets-1 dependent MDR1 transcription.
Int J Mol Sci. 21:17102020. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Sawicka E, Saczko J, Kulbacka J, Szydełko
M, Szymańska B and Piwowar A: The influence of interaction between
cadmium with 17β-estradiol, 2-methoxyestradiol and
16α-hydroxyestrone on viability and P-glycoprotein in ovarian
cancer cell line. Int J Mol Sci. 23:26282022. View Article : Google Scholar
|
|
151
|
Ding Y, Fan J, Fan Z and Zhang K:
γ-Tocotrienol reverses multidrug resistance of breast cancer cells
through the regulation of the γ-Tocotrienol-NF-κB-P-gp axis. J
Steroid Biochem Mol Biol. 209:1058352021. View Article : Google Scholar
|
|
152
|
Fujihara T, Mizobuchi Y, Nakajima K,
Kageji T, Matsuzaki K, Kitazato KT, Otsuka R, Hara K, Mure H,
Okazaki T, et al: Down-regulation of MDR1 by Ad-DKK3 via Akt/NFκB
pathways augments the anti-tumor effect of temozolomide in
glioblastoma cells and a murine xenograft model. J Neurooncol.
139:323–332. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Norouzi-Barough L, Sarookhani M, Salehi R,
Sharifi M and Moghbelinejad S: CRISPR/Cas9, a new approach to
successful knockdown of ABCB1/P-glycoprotein and reversal of
chemosensitivity in human epithelial ovarian cancer cell line. Iran
J Basic Med Sci. 21:181–187. 2018.PubMed/NCBI
|
|
154
|
Peng H, Qiao L, Shan G, Gao M, Zhang R, Yi
X and He X: Stepwise responsive carboxymethyl chitosan-based
nanoplatform for effective drug-resistant breast cancer
suppression. Carbohydr Polym. 291:1195542022. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Teng YN, Wang CCN, Liao WC, Lan YH and
Hung CC: Caffeic acid attenuates multi-drug resistance in cancer
cells by inhibiting efflux function of human P-glycoprotein.
Molecules. 25:2472020. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Ma Y, Wen J, Wang J, Wang C, Zhang Y, Zhao
L, Li J and Feng X: Asiaticoside antagonizes proliferation and
chemotherapeutic drug resistance in hepatocellular carcinoma (HCC)
cells. Med Sci Monit. 26:e9244352020. View Article : Google Scholar : PubMed/NCBI
|