Introduction
Renal cell carcinoma, originating from the
epithelial system of the renal tubules, referred to as kidney
cancer, is one of the most prevalent malignant tumors of the
urinary system. It constitutes 3% of all adult malignancies and
accounts for an estimated 80-85% of renal malignancies (1).
According to the statistics, its incidence ranks the
third place in the urinary tumors, and second only to bladder
cancer and prostate cancer in China (2). The epidemiological survey revealed
that there were 63,000 new cases and 14,000 death cases of RCC in
US in 2016, rendering it the 9th most common cancer. Furthermore,
its incidence is still increasing every year (3). RCC can be divided into different
subtypes in histopathology, the most frequent of which is clear
cell RCC (ccRCC), which accounts for 70-80% of all renal
malignancies. As RCC lacks early clinical symptoms, nearly 20-30%
of patients have metastasized when diagnosed, and the 5-year
survival rate of these patients is only 10%, and the median
survival is 1 year (4). The
radical nephrectomy is the main treatment for localized RCC and has
a favorable clinical effect; however, metastatic RCC has lost the
indications for surgery, mainly based on the conservative
treatments (5). Unfortunately,
RCC is not sensitive to traditional radiotherapy and chemotherapy,
and the cytokines are also difficult to achieve the desired
therapeutic effect (6). Despite
the fact that targeted medicines can significantly prolong
progression-free survival and overall survival of patients with
metastatic RCC, as well as enhance their quality of life,
cytotoxicity and drug tolerance are still existing (7). Therefore, exploring a new effective
therapeutic method is becoming a hot topic in the research field of
RCC (especially for ccRCC).
In recent years, several studies suggested that
plant-derived food (such as polyphenols) intake is associated with
increased anticancer benefits for health (8). Gallic acid (3,4,5-trihydroxybenzoic
acid, GA) is an endogenous and naturally occurring plant polyphenol
which is widely distributed in various plants, fruits and foods
worldwide, including tea, oak, chestnut, grapes, strawberries,
bananas, red wine, vinegar and others as well as Chinese medicinal
herbs (9,10). GA can be absorbed very well by the
human organism. It is a yellow-white crystal with melting point at
250°C and water solubility 1.1% at 20°C. Its molecular mass is
170.12 g/mol and its chemical formula is
C6H2(OH)3COOH (11). GA was initially reported as a free
radical scavenger and cell differentiation inducer. It was
subsequently reported to exhibit a range of biological and
pharmacological properties, including antibacterial, antiviral,
antioxidant and anti-inflammatory activities, especially for its
antitumor activity (12). Of
note, GA was proved to have selective cytotoxicity on certain
cancer cells with very less damage to the normal cells (13,14). GA has diverse effects on various
tumors at different molecular levels. These features make it a
valuable supplement to reduce the risk of tumorigenesis.
Polyphenols were reported by numerous studies to exert the
anticancer effects via a pleiotropic molecular mechanism of action
on cell apoptosis and angiogenesis (15,16). However, with the deepening of the
research on the molecular mechanism of cancer development, certain
studies previously reported that polyphenols could induce autophagy
of cancer cells with dissimilar potencies (17-19). As GA is a well-known polyphenol,
it was hypothesized that GA plays a significant role in anticancer
activities by regulating autophagy.
To our knowledge, autophagy is a dynamic and
evolutionarily conserved process by which cells degrade their
long-lived proteins and damaged components (20). Briefly, it is one kind of
catabolic pathway and begins with the formation of autophagosomes
with double or multi-layer membrane structure. Subsequently, the
autophagosomes fuse with the lysosomes to form autolysosomes with
single layer membrane structure. Finally, the contents are degraded
by the hydrolases in lysosomes and recycled. As a result, cells
digest and recycle nutrients or damaged organelles via autophagy,
which improves cell survival in conditions of inadequate nutrition
or oxygen availability (21).
However, excessive autophagy may also result in autophagic cell
death (22). There is growing
evidence that autophagy plays a crucial role in the regulation of
tumorigenesis. Among them, certain studies indicated that autophagy
functions as tumor suppressor, while the others argued that
autophagy could promote tumor survival and progression (23). The issue, whether autophagy
represents a mechanism that allows tumor cells to survive from
therapy or a mechanism that initiates a non-apoptotic cell death
type is being debated. Additionally, there are limited studies with
regard to the effects of GA on RCC cells. Therefore, in order to
further address GA and its potential autophagy regulating
character, the roles of GA in RCC development were investigated in
the present study.
Materials and methods
Reagents
GA with 99% purity and 3-methyladenine (3-MA) were
obtained from MilliporeSigma. Roswell Park Memorial Institute-1640
medium (RPMI-1640), fetal bovine serum (FBS),
penicillin-streptomycin were purchased from Gibco; Thermo Fisher
Scientific, Inc. Cell Counting Kit-8 (CCK-8) was acquired from
Dojindo Laboratories, Inc. PI/RNase Staining Buffer kit and
Matrigel were obtained from BD Biosciences. The primary antibodies
against PI3K, phosphorylated (P-)PI3K, Akt, P-Akt, Atg16L1 and
P-Atg16L1 were purchased from Cell Signaling Technology, Inc. The
primary antibodies against LC3B, Beclin-1, P62 and GAPDH were
purchased from Abcam. The secondary antibody was obtained from
LI-COR Biosciences. PrimeScipt™ RT reagent Kit with gDNA Eraser and
SYBR@Premix Ex Taq™ II were obtained from Takara Bio, Inc.
Cell culture and drug preparation
The ccRCC cell lines (786-O and ACHN) and the normal
renal tubular epithelial cell (HK-2) were obtained from Cell Bank
of Wuhan University (Wuhan, China). The cells were cultured in
RPMI-1640 medium (Gibco; Thermo fisher scientific, Inc.) with 10%
FBS, 100 units/ml penicillin and 100 units/ml streptomycin, then
maintained in 37°C with 5% CO2 incubator (Binder GmbH).
The GA and 3-MA stock solution was respectively prepared at a
concentration of 73,477 µmol/l and 166 mmol/l: respectively
added 0.5 g GA and 2.5 g 3-MA into a EP tube, respectively added 40
and 100 ml FBS-free RPMI-1640 medium, respectively mixed on a
vortex oscillator to ensure the complete dissolution, and then
stored at −20°C. Before application, the GA and 3-MA stock solution
should be appropriately diluted and heated. To generate the working
solution (GA: 0, 5, 10 and 20 µmol/l; 3-MA: 5 mmol/l), the
GA and 3-MA stock solutions were taken out, dissolved at 37°C, and
then diluted to the designated concentration with FBS-free
RPMI-1640 medium. In addition, the light should be avoided in the
period of GA and 3-MA using.
Cell viability assay
The ccRCC cells (786-O and ACHN) and the normal
renal tubular epithelial cell (HK-2) were first plated in a 96-well
plate at 5×103 cells/well for 24 h of incubation at 37°C
for the CCK-8 assay. The preceding medium was then taken out, and
100 µl of 0, 5, 10, 20 µmol/l GA solution was placed
into each well for 24, 48 and 72 h of stimulation, respectively.
When the scheduled time arrived, the preceding medium was replaced
with 100 µl new RPMI-1640 medium containing 5% CCK-8. A
microplate reader (PerkinElmer, Inc.) was used to measure the
absorbance at 450 nm following 1 h of incubation at 37°C in the
dark. The half maximal inhibitory concentration (IC50)
and inhibition rate were then calculated for each group.
Plate clone formation assay
The ccRCC cells (786-O and ACHN) were initially
plated into the six-well plate and cultured for 48 h. The cells
were treated with varying concentrations of GA (0, 5, 10 and 20
µmol/l) for a duration of 24 h. Upon completion, the cells
in each well were digested, counted, and plated into a new six-well
plate at a seeding density of 1×103 cells/well with
complete medium. They were then cultured consecutively for one week
at 37°C. Subsequent to removal of the medium, the cells were washed
with phosphate buffered saline (PBS) for 3 times. Following this,
the cells were fixed using a 4% paraformaldehyde solution for 20
min at room temperature (RT), and subsequently stained with a 0.5%
crystal violet solution for 20 min at RT. The stained cells were
ultimately washed with PBS and air-dried, and the number of
colonies was counted manually. The minimum number of cells required
for colony formation was 50-100.
Cell cycle analysis
The ccRCC cells (786-O and ACHN) were firstly
stimulated with different concentrations of GA solution (0, 5, 10
and 20 µmol/l) for 24 h. The cells were then collected and
washed with PBS. Each tube received 1 ml 70% ethanol and was
incubated at −20°C overnight for fixing. Following that, the fixed
ccRCC cells were collected and washed with 4°C PBS, and 500
µl PI/RNase dye solution was added into each tube and
incubated for 10 min at 25°C in the dark. Finally, flow cytometry
(FACSCanto II cytometer; BD Biosciences) was used to measure the
fluorescence intensity of each group in real time and analyzed by
FlowJo software (Tree Star, Inc.).
Wound healing assay
The ccRCC cells (786-O and ACHN) from each group
were plated in six-well plates for 48 h. After the cell density
reached ~80%, the monolayer was wounded by scratching it lengthwise
along the plate surface with a 200-µl sterile pipette tip.
The medium was then removed, and the cells were washed twice with
PBS and cultured in serum-free RPMI-1640 medium. To capture images
of cell migration, an inverted microscope was used (Olympus
Corporation) and recorded at 0, 12 and 24 h. Finally, the area of
migration (µm2) was measured using the ImageJ
software (Version 1.53; National Institutes of health).
Transwell migration and Matrigel invasion
assays
The Transwell migration and invasion assays were
performed on ccRCC cells (786-O and ACHN) stimulated with varying
concentrations of GA solution (0, 5, 10 and 20 µmol/l).
After 24 h of stimulation, the cells were collected by trypsin and
suspended in FBS-free RPMI-1640 medium. The cell density was
adjusted and 100 µl of 2×105 cells/ml ccRCC cell
suspension was added to the upper Boyden chamber (8-µM pore
size; Corning, Inc.). The lower chamber was filled with 700
µl RPMI-1640 medium containing 20% FBS. The 24-well plate
was then placed in a 37°C incubator. After 24 h of incubation, the
Transwell chamber was removed from the incubator and non-migratory
cells were scraped with a cotton swab. The migratory cells were
fixed with 4% paraformaldehyde for 20 min at RT and dyed with 0.5%
crystal violet solution for another 20 min. The migratory cells
were observed and images were captured using an inverted light
microscope (Olympus Corporation). The cells in 5 randomly selected
fields were counted and the mean was calculated. Matrigel was
diluted 1:8 with FBS-free RPMI-1640 media on ice for the Matrigel
invasion assay. The upper chambers of the Transwell were coated
with 80 µl of the diluted Matrigel. The next processes were
identical to the migration experiments.
Western blot analysis
The proteins of ccRCC cells were extracted using
RIPA lysis buffer (Thermo Fisher Scientific, Inc.) and phosphatase
inhibitors were added to prevent sample degradation. Protein
concentrations were measured with a bicinchoninic acid protein
assay kit (Beyotime Institute of Biotechnology), and
electrophoresed on sodium dodecyl sulfate-polyacrylamide gels with
varying concentrations of 8, 12, or 15%. The proteins were then
transferred onto polyvinylidene difluoride membranes
(MilliporeSigma). After blocking for 1 h at RT with 5% skimmed milk
in Tris-buffered saline containing 0.1% Tween-20 (TBST) and washing
three times with TBST, the membranes were incubated overnight at
4°C with the specified monoclonal primary antibodies
(1:1,000-2,000). The membranes were then washed 3 times with TBST
and incubated with the corresponding goat-anti-rabbit secondary
antibody (1:20,000) at RT for 1 h. After washing 3 times with TBST,
the membranes were scanned with a two-color infrared imaging system
(LI-COR, Biosciences). GAPDH was used as an internal reference.
Densitometric analysis was performed using ImageJ software.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from ccRCC cells (786-O and
ACHN) using TRIzol reagent (Invitrogen; Thermo fisher scientific,
Inc.). AB7500 detection system (Thermo fisher scientific, Inc.) was
used to measure the reverse transcription reactions, which were
carried out using a PrimeScriptTM RT reagent kit (Takara Bio, Inc.)
in accordance with the manufacturer's instructions. qPCR reactions
were prepared using a SYBR Green Mix (Takara Bio, Inc.) at a final
volume of 20 µl. The relative gene expression level was
calculated using the 2−ΔΔCq method. The data were
normalized using an internal control called GAPDH. Three duplicates
of each response were carried out. Primer sequences are displayed
in Table I.
 | Table IList of primers used for reverse
transcription-quantitative PCR. |
Table I
List of primers used for reverse
transcription-quantitative PCR.
| Gene name | Primer sequence
(5′→3′) |
|---|
| LC3B | F:
GGCTTTCAGAGAGACCCTGAG |
| R:
CCGTTTACCCTGCGTTTGTG |
| Beclin-1 | F:
CCACAGAAAGTGCCAACAGC |
| R:
GACGTTGAGCTGAGTGTCC |
| P62 | F:
GGTCGCGCTCACCTTTCT |
| R:
GGAGATGAGGCTCCGCAAT |
| GAPDH | F:
AAAGCCTGCCGGTGACTAAC |
| R:
TTCCCGTTCTCAGCCTTGAC |
Transmission electron microscopy
(TEM)
The ccRCC cells (786-O and ACHN) were collected and
fixed with 2.5% glutaraldehyde at 4°C for 2 h and 1% osmium
tetroxide for 30 min and then dehydrated in a graded series of
ethanol. The cells were embedded in epoxy resin and sectioned at
70-nm thickness using an ultramicrotome. The sections were stained
with 0.2% lead citrate and 2% uranyl acetate. The images were
captured with a transmission electron microscope (Hitachi,
Ltd.).
Xenografts and tumor metastasis
model
Female BALB/c nude 4-week-old mice (n=48) weighing
20±2 g were acquired from the Wuhan University Animal Experiment
Center (Wuhan, China). All animals were housed in a specific
pathogen-free environment (12/12-h light/dark cycle; ad
libitum access to food and water) with the right humidity and
temperature. Each mouse received an injection of 200 µl of
PBS containing ~1×107 786-O cells and ACHN cells in the
left flank. Every week after inoculation, tumor volume was measured
and computed using the formula (length x width2)/2. The
nude mice were separated into 4 groups of 3 mice each after the
average tumor volume reached ~100 mm3 (~10 days later).
The mice received freshly prepared solutions (the only source of
drinking water) every Monday, Wednesday, and Friday after the
weighted GA powder was separately dissolved in the water to form
solutions with varying concentrations of GA (w/v) (0.3, 0.6 and
0.9% in regular drinking water). The control mice were given access
to regular water during this time. All mice were administered an
excessive dose of carbon dioxide for euthanasia after two months,
and the weight of the tumors was measured immediately.
Additionally, the tumor metastasis model was created by injecting
786-O and ACHN cells into the tail veins of additional BALB/c nude
mice. To do this, 1×106 cells mixed with PBS were
injected into each mouse in 4 groups (n=3 per group), and the mice
were then administered drinks containing various concentrations of
GA solution. After 2 months, all mice were euthanized with an
overdose of carbon dioxide, and the weight of tumors was measured
immediately. Displacement rate of CO2 used for
euthanasia was 30%. All animal studies were approved by the Animal
Care and Use Committee of Anhui Medical University (approval no.
LLSC20232011; Hefei, China), and all animal experiments were
carried out in accordance with the National Institutes of Health's
Guide for the Care and Use of Laboratory Animals.
Hematoxylin-eosin (H&E) staining
The lung tissues were fixed in 4% paraformaldehyde
to cause coagulation and denaturing of proteins at RT for 8 h. All
samples were dehydrated in an ethanol gradient and xylene.
Subsequently, the tissues were embedded in paraffin, cooled and
solidified into blocks. Afterwards, paraffin blocks were cut into
4-µm-thick slices. The sections were then stained with
hematoxylin and eosin and images were captured by light microscopy
(Olympus Corporation).
Immunohistochemistry (IHC) assay
For antigen retrieval, the paraffin-embedded tumor
sections were heated at 105°C for 10 min in a citric acid buffer
(0.01 M) and then deparaffinized in xylene for 15 min. Following a
10-min PBS washing, the slides were put into a 5-min immersion in a
3% hydrogen peroxide solution to inhibit endogenous peroxidase
activity. The sections were then washed once more in PBS for 5 min,
blocked with the blocking solution for 1 h at RT, and then
incubated with the appropriate antibody for 1 h at RT. Finally, an
immunochemistry kit (OriGene Technologies, Inc.) was used to find
the immune complexes. A light microscope (Olympus Corporation) was
used to observe the sections.
Statistical analysis
At least three independent times of each experiment
were conducted. The mean ± standard deviation (SD) was used to
represent the data. The treatment groups were compared using
one-way analysis of variance (ANOVA) with SPSS 22.0 (SPSS, Inc.).
Bonferroni's post hoc test was used. *P<0.05 was considered to
indicate a statistically significant difference.
Results
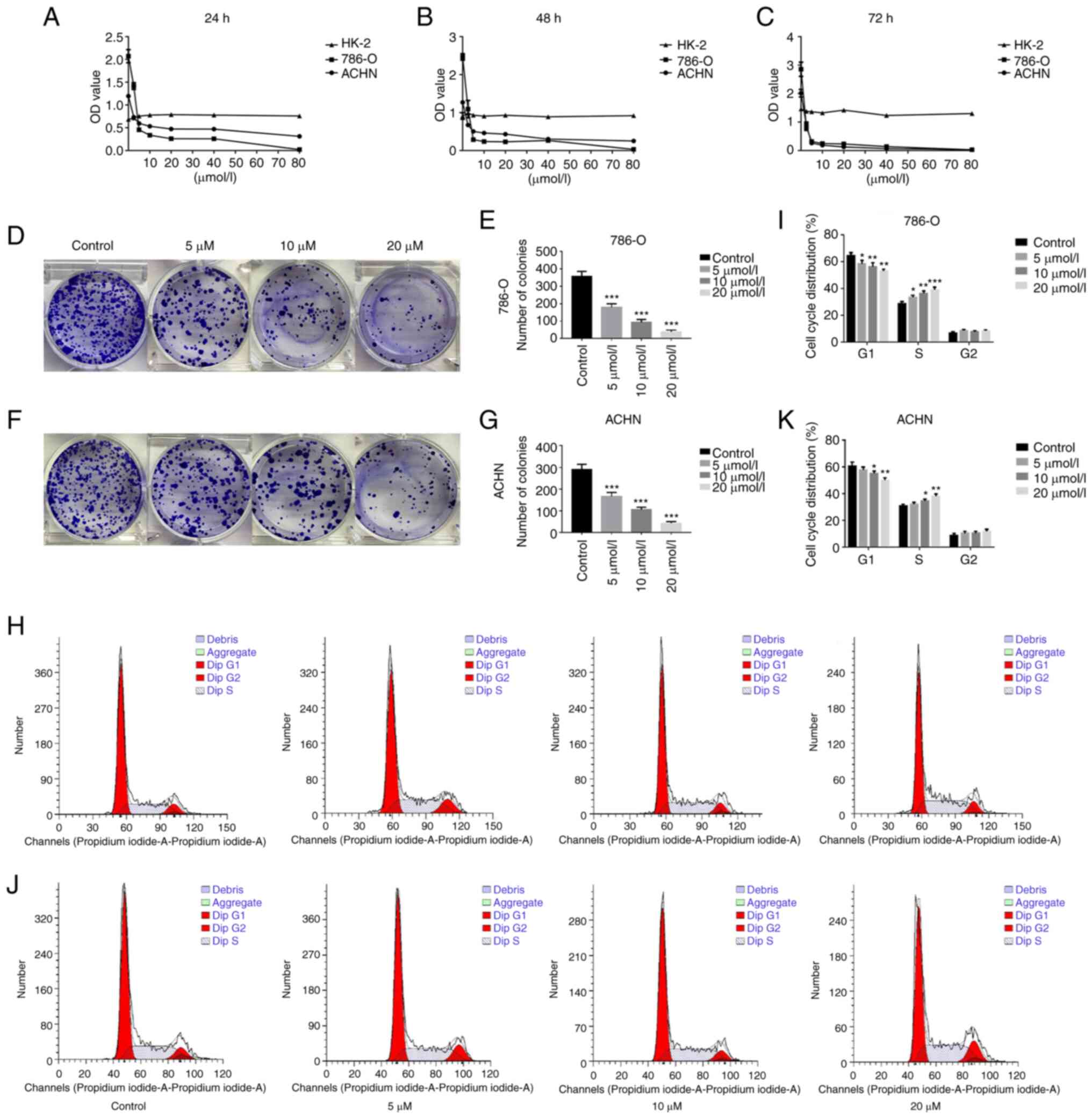
GA inhibits the viability of ccRCC
cells
In a previous study by the authors, the effects of
each monomer in pomegranate peel tannins on human bladder cancer
T24 cells viability were compared, and it was found the inhibition
of GA on T24 cells viability was significantly higher than the
other three monomers, which indicated that GA is the effective
anticancer component in pomegranate peel tannins (24). Therefore, in order to measure the
effects of GA on ccRCC cells (786-O and ACHN) and renal tubular
epithelial cell (HK-2) viability, CCK-8 assay was used. The
findings revealed that, when compared with the control group, the
vitality of the 786-O and ACHN cells in the GA-treated group was
considerably reduced (P<0.05), whereas that of the HK-2 cells
was not significantly affected. These findings demonstrated that GA
significantly decreased the viability of 786-O and ACHN cells in a
time- and concentration-dependent manner, but not HK-2 cells. As
demonstrated in Fig. 1A-C, the
IC50 of GA stimulating 786-O cells for 24, 48 and 72 h
were 4.20, 2.89 and 2.39 µmol/l respectively, while the
IC50 of GA stimulating ACHN cells for 24, 48 and 72 h
were 10.51, 7.05 and 2.75 µmol/l, respectively. These
findings revealed that GA significantly reduced the viability of
786-O and ACHN cells, especially when stimulated for 24 h
(P<0.05). As a consequence, GA with 0, 5, 10 and 20
µmol/l was identified as the suitable action concentration
and 24 h as the optimum action period, combining with the results
of CCK-8.
GA changes the morphology of ccRCC
cells
To study the effect of GA on ccRCC cell morphology,
an inverted microscope was utilized and it was observed that both
the 786-O and ACHN cells exhibited elongation, robust adhesion,
large cellular frames, clear cell outlines, and overall exhibited
signs of favorable condition. Following 24-h GA stimulation, the
786-O and ACHN cells demonstrated a notable decrease in size,
weakened adhesive properties, reduced intercellular connections,
blurred cellular outlines, and appeared to be in a state of
cellular death (Fig. S1A and
B).
GA inhibits proliferation of ccRCC
cells
To further evaluate the effect of GA on ccRCC cell
proliferation, a plate clone formation assay was utilized. The
results from the plate cloning assay indicated a significant
decrease in the number of colonies for both 786-O and ACHN cells in
the GA-treated groups, compared with the control group. Notably,
the number of colonies decreased consistently with increasing
concentrations of GA (Fig. 1D-G).
The results indicated that GA significantly suppressed
proliferation of ccRCC cells in a concentration-dependent way.
GA blocks the cycle process of ccRCC
cells
To evaluate the effect of GA on the cell cycle of
ccRCC cells, flow cytometry was employed. The results demonstrated
that the control group had 63.19% 786-O cells in G1 phase and
28.71% in S phase (Fig. 1H and
I). Additionally, the results for the control group revealed
that 60.53% of ACHN cells were in G1 phase, while 31.72% were in S
phase (Fig. 1J and K). In
addition, as the concentration of GA increased, there was a
significant increase in the number of 786-O and ACHN cells in S
phase, accompanied by a significant decrease in the number of cells
in G1 phase (P<0.05). In summary, the findings indicated that GA
effectively hindered the cell cycle of ccRCC cells specifically in
the S phase.
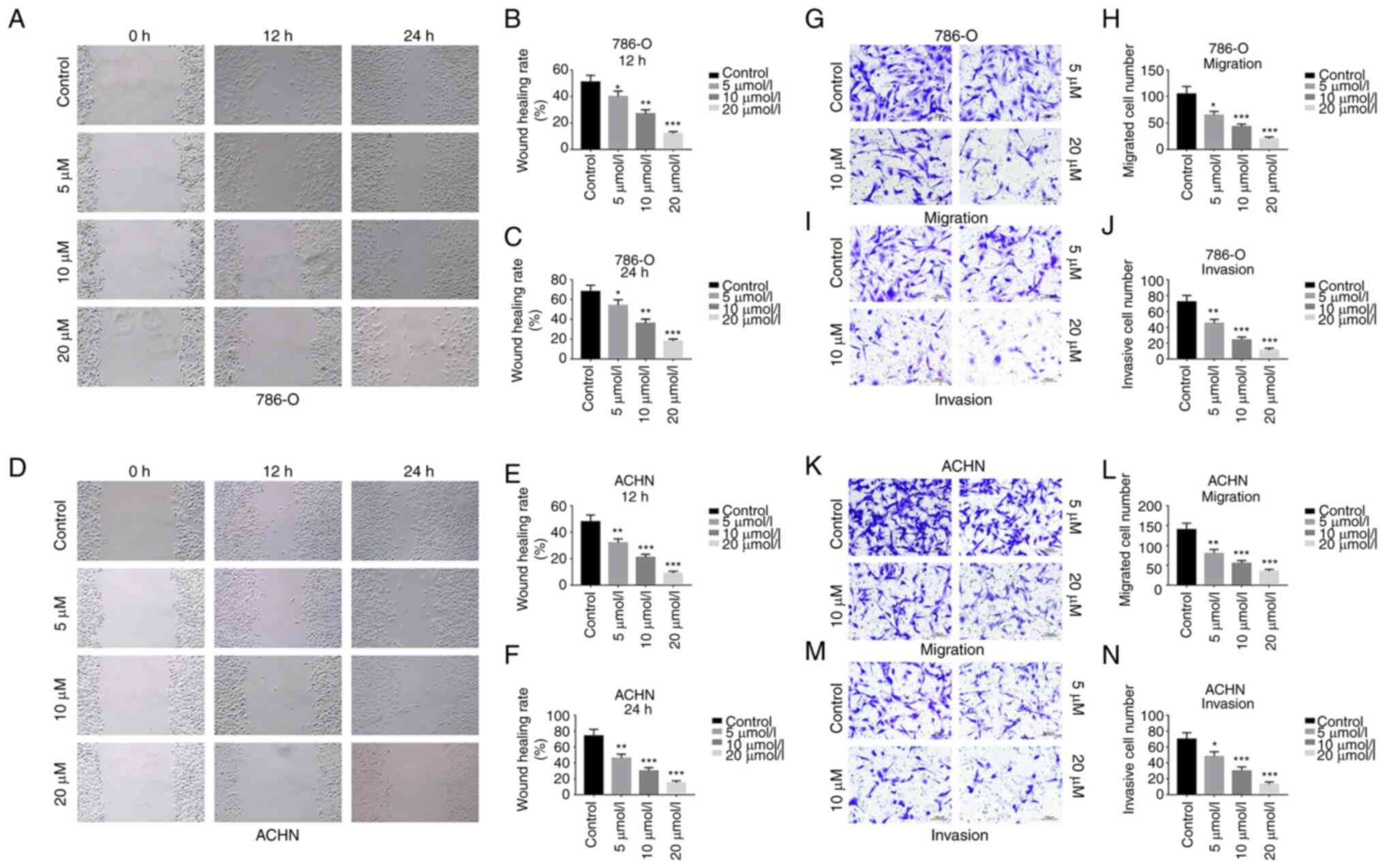
GA inhibits migration and invasion of
ccRCC cells
The wound healing and the Transwell assays were used
to evaluate the effects of GA on the migration and invasion of
ccRCC cells. It was identified that, in comparison with the control
group, the GA-treated group significantly inhibited the migration
of 786-O and ACHN cells, and that the inhibition rate was dependent
on the concentration of GA (Fig.
2A-F). Moreover, the Transwell migration assay demonstrated
that GA treatment led to a significant decrease in the number of
786-O cells (Fig. 2G and H) and
ACHN cells (Fig. 2K and L) that
had passed through the chamber membrane. Furthermore, it was
observed that the number of 786-O (Fig. 2I and J) and ACHN (Fig. 2M and N) cells that passed through
the Matrigel and chamber membrane decreased as the concentration of
GA increased in the Matrigel invasion assay. These results
indicated that GA significantly inhibited migration and invasion of
ccRCC cells in a concentration-dependent way.
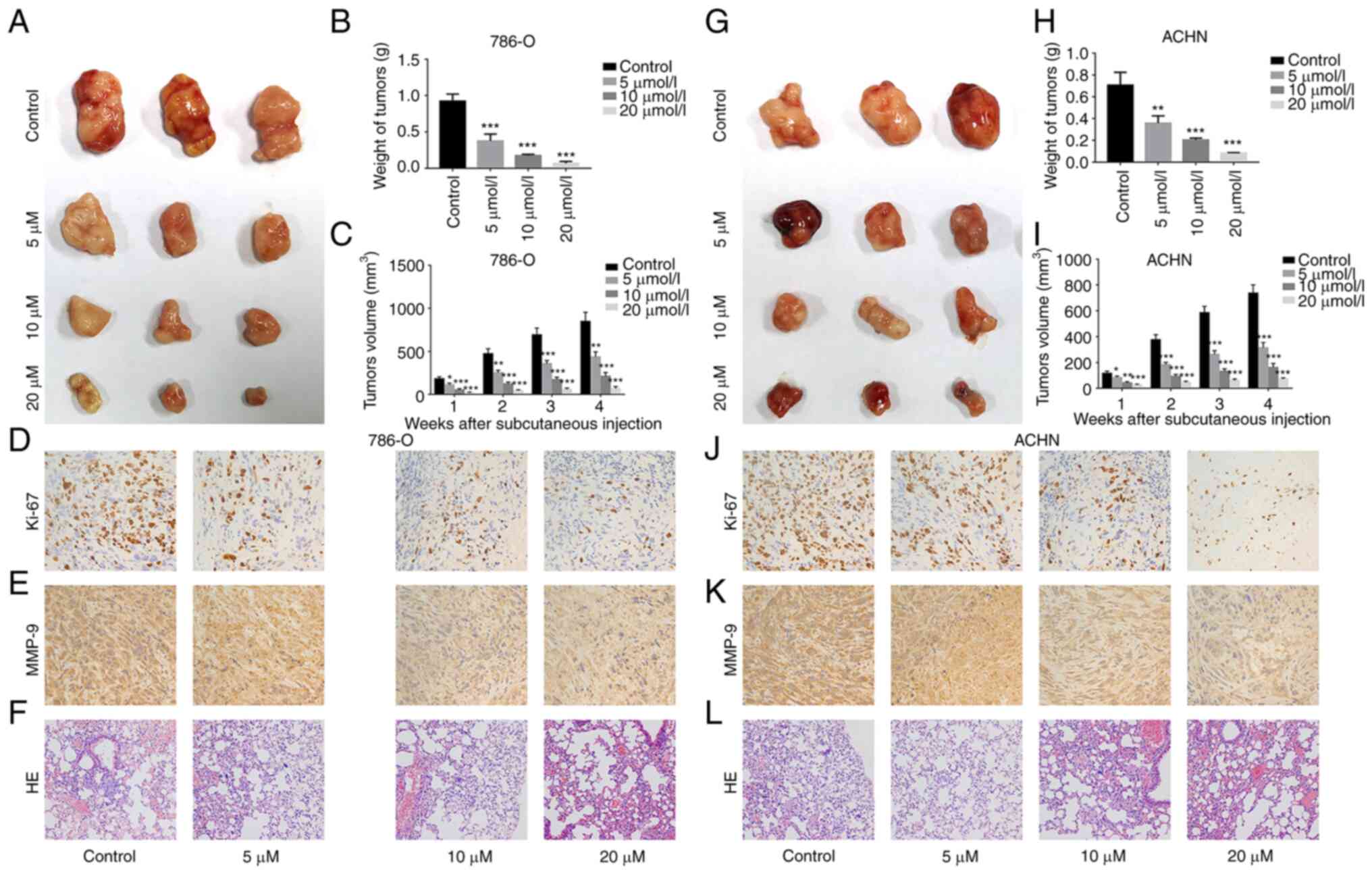
GA inhibits growth and metastasis of
ccRCC
To investigate the effect of GA on the growth and
metastasis of ccRCC, xenograft and tumor metastasis models were
established. In both models, 786-O and ACHN cells were used, and it
was found that tumors were successfully formed in all mice injected
with these cells (Fig. 3A-C and
G-I, respectively). Moreover, the weight and volume of tumors
were significantly lighter and smaller in the GA-treated group than
in the control group. The IHC results revealed a significant
decrease in the expression of Ki-67 (a marker of proliferation) and
MMP-9 (a marker of metastasis) in the GA-treated group compared
with the control group for both 786-O cells (Fig. 3D and E) and ACHN cells (Fig. 3J and K). Finally, the H&E
staining results demonstrated that compared with that in the
control group, 786-O cells (Fig.
3F) and ACHN cells (Fig. 3L)
metastasized into the lungs were significantly decreased in the
GA-treated group. The quantification of representative IHC and
H&E images is demonstrated in Fig. S2. These results indicated that
ccRCC growth and metastasis were significantly inhibited by GA in a
concentration-dependent way.
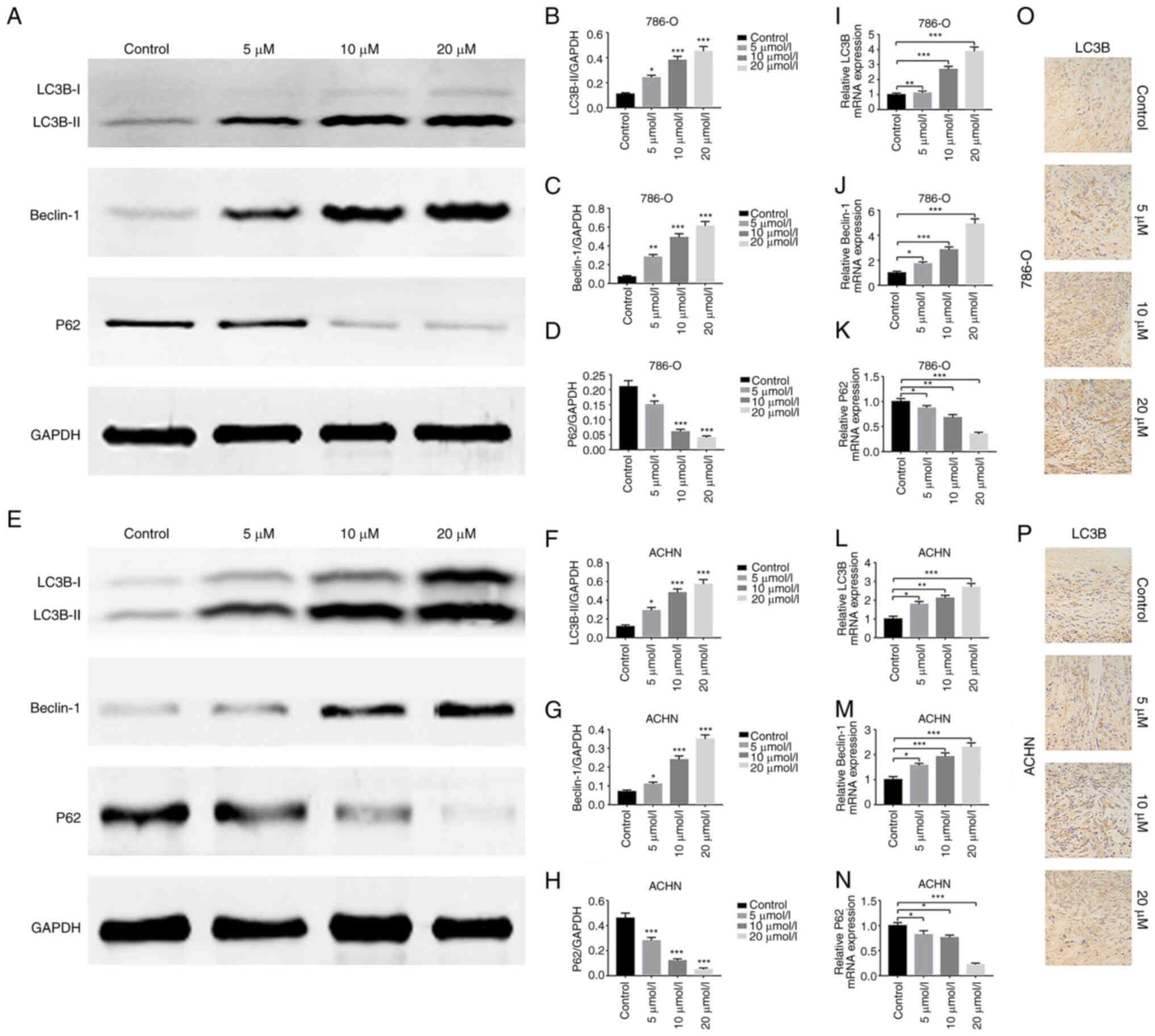
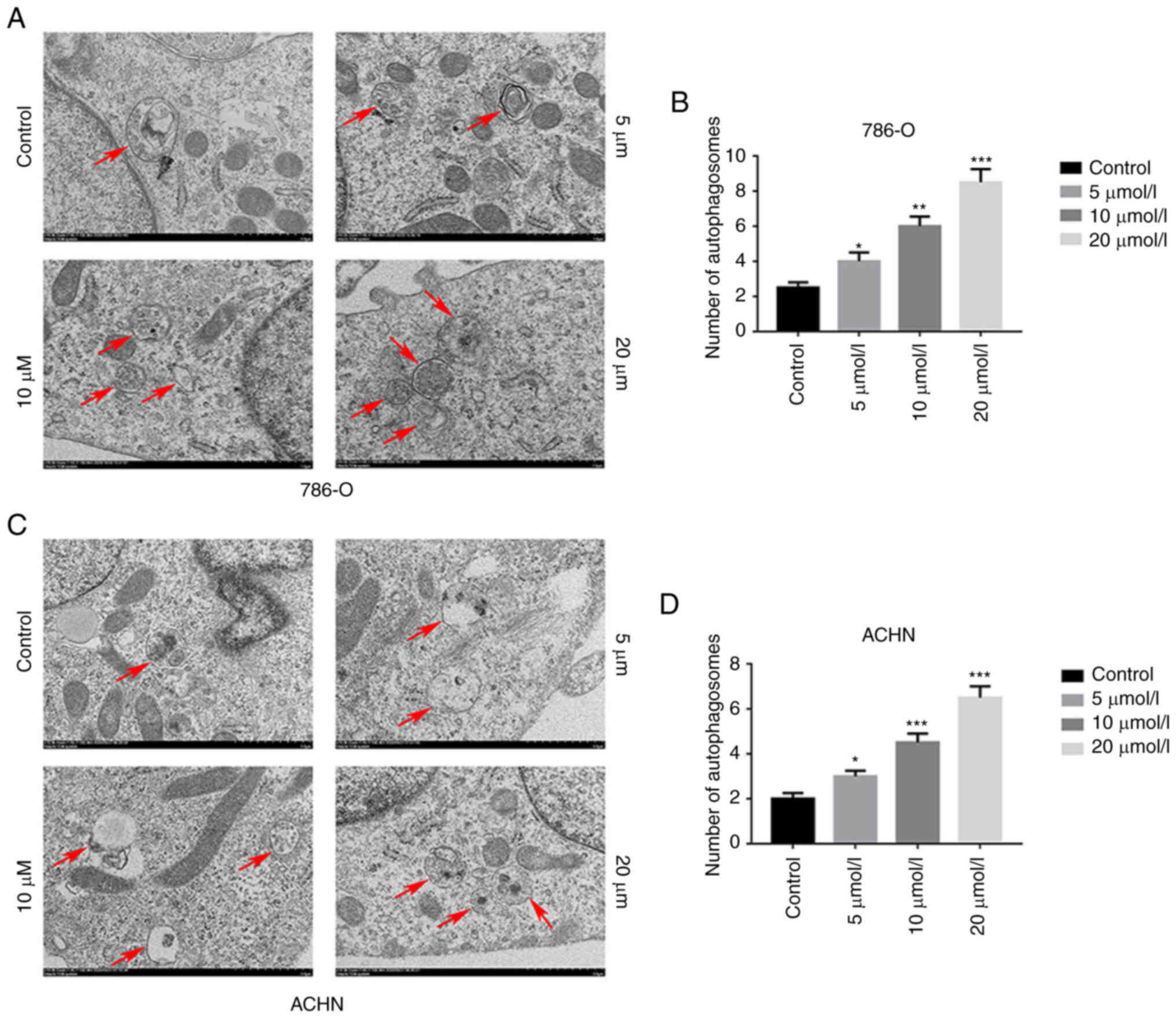
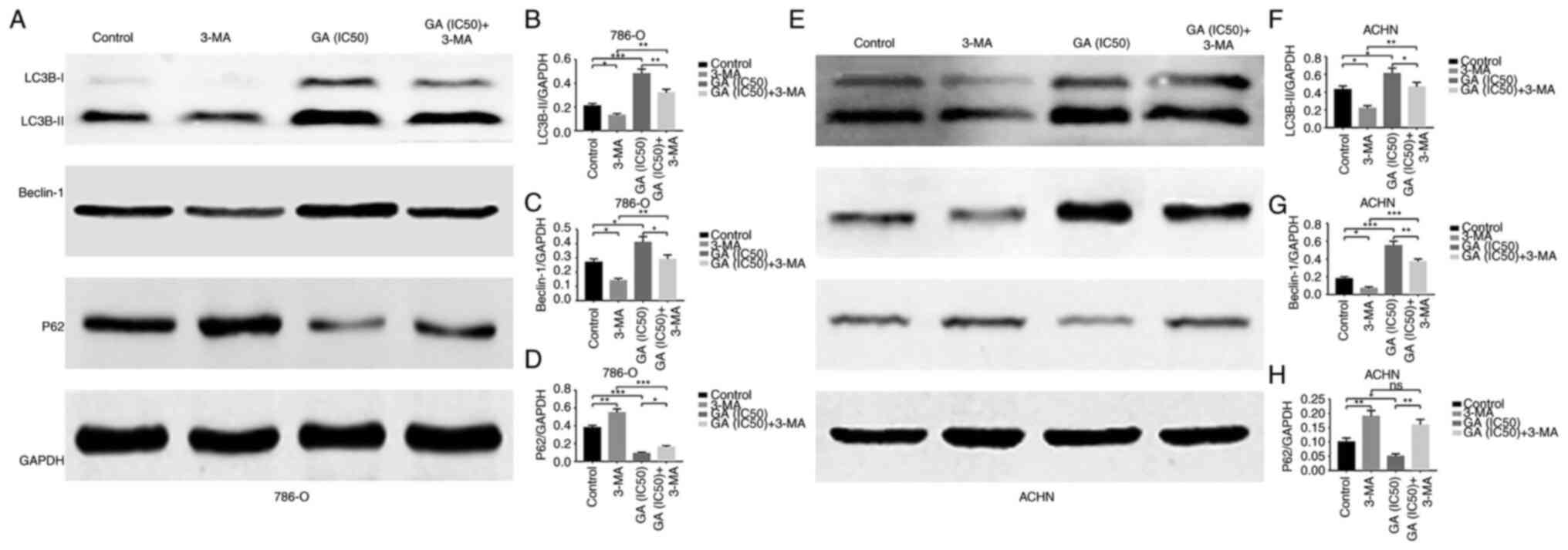
GA induces autophagy in ccRCC
To investigate the correlation between GA and
autophagy in ccRCC, the expression of autophagy-related markers was
examined. Firstly, the western blotting results revealed that
compared with that in the control group, LC3B-II and Beclin-1
protein expression was significantly increased, while P62 protein
expression was significantly decreased in the GA-treated groups
(Fig. 4A-H). Meanwhile, RT-qPCR
results demonstrated significantly increased LC3B and Beclin-1 gene
transcription and significantly decreased P62 gene transcription in
the GA-treated groups than in the control group (Fig. 4I-N). Subsequently, the TEM
revealed a significantly higher number of autophagosomes in the
groups treated with GA compared with the control group (Fig. 5A-D). Moreover, the IHC results
indicated that LC3B expression was significantly increased in the
tumors of GA-treated groups than in the tumors of control group
(Fig. 4O and P). In addition, the
western blotting results revealed that LC3B-II and Beclin-1 protein
expression was significantly decreased and P62 protein expression
was significantly increased in the GA (IC50) + 3-MA
treatment group compared with the GA (IC50) group
(Fig. 6A-H). These results
indicated that GA could induce ccRCC autophagy in vivo and
in vitro.
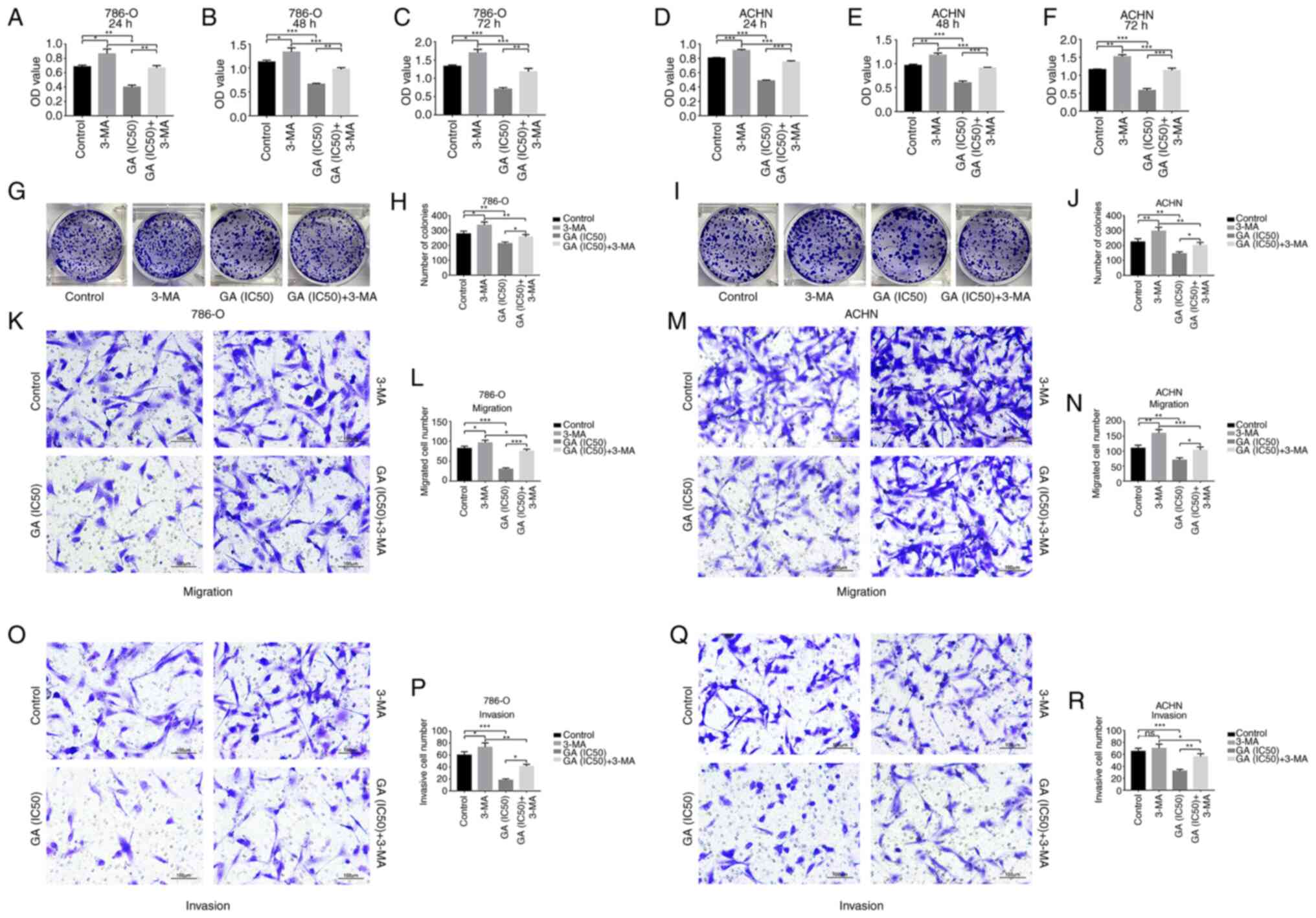
GA inhibits proliferation of ccRCC cells
by inducing autophagy
In order to assess the role of autophagy in the
inhibition of GA on ccRCC cells proliferation, 3-MA (a common
autophagy inhibitor) was added on the basis of GA (IC50)
stimulation. The CCK-8 results demonstrated that viability of 786-O
and ACHN cells was significantly inhibited in the GA
(IC50) group compared with the control group, while the
inhibitory effects of GA (IC50) on 786-O and ACHN cells
were significantly reversed by 3-MA (Fig. 7A-F). The plate clone formation
assay indicated that there were fewer colonies of 786-O and ACHN
cells in the GA (IC50) group compared with the control
group, while the reduction trend on the colonies of 786-O and ACHN
cells were significantly reversed by 3-MA (Fig. 7G-J). These results indicated that
GA could suppress proliferation of ccRCC cells by inducing
autophagy.
GA inhibits migration and invasion of
ccRCC cells by inducing autophagy
In order to further assess the role of autophagy in
the inhibition of GA on ccRCC cells migration and invasion, 3-MA
was added on the basis of GA (IC50) stimulation.
According to the results of the Transwell migration assay,
treatment with GA (IC50) led to a significant reduction
in the number of migratory 786-O and ACHN cells compared with the
control group. On the other hand, the migration of 786-O and ACHN
cells was significantly increased after treatment with 3-MA,
reversing the effect observed with GA (Fig. 7K-N). Moreover, the Matrigel
invasion assay results indicated that there were fewer invasive
786-O and ACHN cells number in the GA (IC50) group than
in the control group, while the invasion results of 786-O and ACHN
cells were significantly reversed by 3-MA (Fig. 7O-R). These results indicated that
GA could suppress migration and invasion of ccRCC cells by inducing
autophagy.
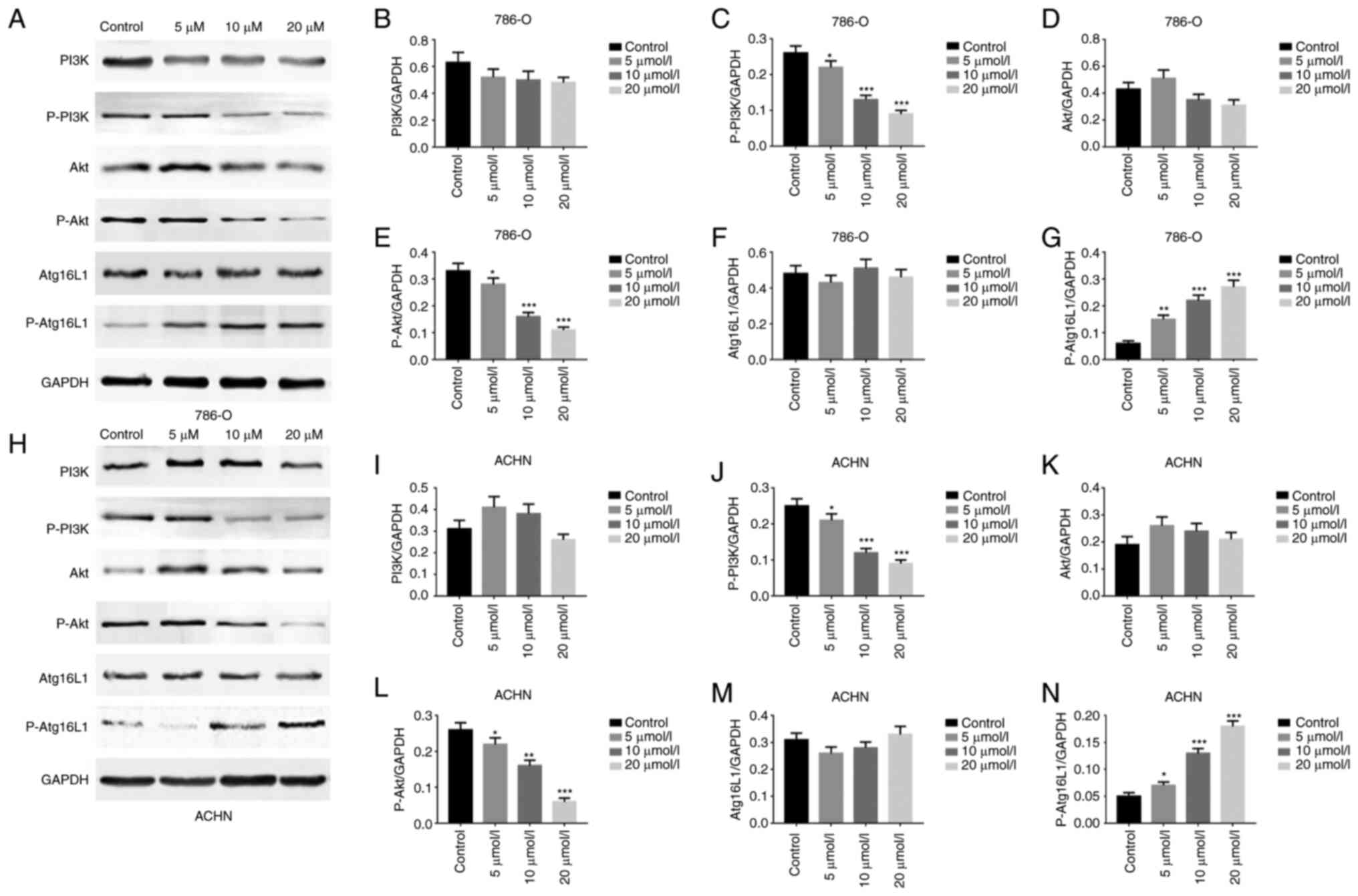
GA induces autophagy of ccRCC cells via
the PI3K/Akt/Atg16L1 signaling pathway
In order to explore the mechanism involved in the
induction of ccRCC cells autophagy by GA, the transcription of
related genes and the expression of critical protein were assessed
in ccRCC cells (786-O and ACHN). Firstly, the RT-qPCR results
revealed that Atg16L1 gene expression was significantly increased
in both 786-O and ACHN cells (Fig. 8A
and B). Subsequently, the western blotting results demonstrated
that compared with that in the control group, P-PI3K and P-Akt
protein expression levels were significantly decreased, while
Atg16L1 protein expression was significantly increased in the
GA-treated groups (Fig. 9A-N).
These results indicated that GA could induce autophagy in ccRCC
cells via the PI3K/Akt/Atg16L1 signaling pathway.
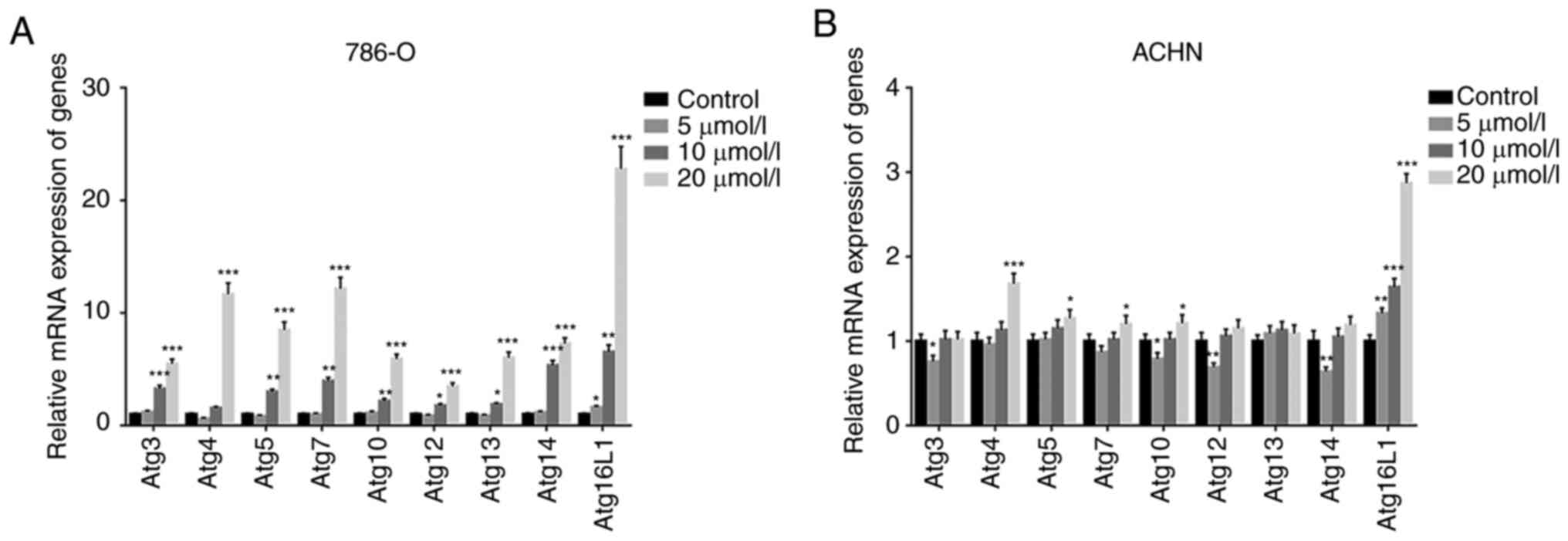 | Figure 8GA regulates the expression of Atgs
in ccRCC cells. (A and B) Reverse transcription-PCR analyses of
autophagy-related genes, including Atg3, Atg4, Atg5, Atg7, Atg10,
Atg12, Atg13, Atg14, Atg16L1, in ccRCC cells with different
concentrations of GA treatment. *P<0.05,
**P<0.01 and ***P<0.001 compared with
the control group. ccRCC, clear cell renal cell carcinoma; Atg,
autophagy-related gene; GA, gallic acid. |
Discussion
RCC is a leading cause of cancer-related death as
previously reported, especially for ccRCC (25). Therefore, it is particularly
important to find an effective and anti-resistance treatment with
minor side effects for it. Modern drug research mainly focuses on
the identification of effective and safe drugs. Although there are
numerous cancer projects at present, the choices in replacing the
traditional treatments for cancer patients are very limited. It is
estimated that ~49% of drugs that are involved in the treatment of
cancers derive from either natural products or their derivatives
(26). GA, as a predominant
polyphenol, its various biological activities have been reported;
however, the major interest in GA is its antitumoral activity. GA
has been demonstrated to inhibit the development of several cancer
types including leukemia, prostate, lung, oral, pancreatic, glioma,
gastric, colon, breast, cervical and esophageal cancer via animal
models in vivo or via cell lines in vitro (27-32). Specifically, the inhibitory
effects of GA on cancer progression involve in cell cycle, growth,
metastasis, angiogenesis and apoptosis. For example, Lee et
al (33) reported that GA
caused an increase in G0/G1 and sub-G1 phase ratio in breast cancer
cells, and found that GA downregulated cyclin D1/CDK4 and cyclin
E/CDK2, and upregulated p21 and p27. Lima et al (34) presented that GA decreased
proliferation of hepatocellular carcinoma cells in a dose-dependent
manner without causing necrosis; moreover, GA did not cause
regrowth of cancer cells in a long period. Ho et al
(35) supported that GA inhibited
metastasis and invasive growth of gastric cancer cells via RhoB
increase, AKT/small GTPase signaling downregulation and NF-κB
activity inhibition. Zhao and Hu (9) indicated that GA significantly
reduced proliferation of human cervical cancer cells and the tube
formation in human umbilical vein endothelial cells. Subramanian
et al (36) displayed that
GA induced the apoptosis of colon cancer cells with the
morphological changes. These actions of GA against cancers make it
an important biological molecule for therapeutic use.
Based on the aforementioned discussion, it is
concluded that the inhibition of cell proliferation and induction
of cell death are the crucial focus for cancers therapy. Chemical
drug therapy typically involves the induction of caspase-mediated
apoptosis, which is a well-established fact. However, a major
obstacle associated with this approach is the development of
resistance by cancer cells to drug-induced apoptosis over time
(18). Therefore, agents that can
induce alternative type of programmed cell death (PCD) may help in
breaking this apoptosis-resistance of cancer cells to improve the
therapeutic efficiency. Autophagy is a type of PCD, and the
existence of this alternative cell death may make the
aforementioned inference to be the truth. Autophagy is a cellular
process that is conserved throughout evolution. It involves the
degradation and recycling of cellular components. The molecular
machinery of autophagy has been clarified in yeast and mammals.
There are 11 autophagy-related genes (Atgs) and 8 orthologs in
mammals, which play a critical role in the autophagy pathway. Atg3,
Atg4 and Atg8/LC3 are involved in phagophore expansion, while Atg5
is crucial for generating the Atg12-Atg5 conjugate required for
this process. Atg6/Beclin-1 and Atg14 are components of PI3K
complexes, which are involved in the induction of autophagy, while
Atg7 and Atg10 are E1-like and E2-like enzymes, respectively, that
mediate conjugation systems involved in phagophore expansion. Atg9
is a transmembrane protein involved in autophagy induction and
Atg13 serves as a binding partner and regulator of Atg1/ULK.
Lastly, Atg12 and Atg16L1 are involved in directing LC3 lipidation
on autophagosomal membranes, which is essential for phagophore
expansion (37).
Recently, the relationship between autophagy and
cancers has been extensively studied, indicating that autophagy can
promote or inhibit cancers progression under different conditions.
Therefore, the implication of autophagy in tumorigenesis remains
controversial (38). Meanwhile,
an increasing number of studies have shown that polyphenols can
suppress the development of cancers by activating the death program
of cancer cells, including autophagy. For example, Zhao et
al (17) found that urolithin
A, a major polyphenol metabolite, induced both autophagy and
apoptosis in colorectal cancer cells, and mainly reflected in the
decreased cell proliferation, delayed cell migration and inhibited
cell metastasis. Tedesco et al (39) indicated that the polyphenols in
the lyophilized red wine induced apoptotic and autophagic cell
death in human osteosarcoma in a dose-dependent manner with a
maximum effect in the range of 100-200 µg/ml equivalents of
GA. Liu et al (40)
identified that Sanguisorba officinalis radix, a chief
constituent of which is GA, inhibited colorectal cancer cell
proliferation and sensitized the cancer cells to 5-FU therapy by
activating a reactive oxygen species-mediated and
mitochondria-caspase-dependent apoptotic pathway. Wang et al
(19) displayed that punicalagin,
a polyphenol isolated from Punica granatum, induced death of
human glioma cells through apoptotic and autophagic pathways, and
the suppression of autophagy dose-dependently alleviated the cell
death caused by punicalagin. Dong et al (41) demonstrated that
penta-1,2,3,4,6-O-galloyl-β-D-glucose, a naturally existing
gallotannin polyphenol compound found in oriental herbs, promoted
senescence of cancer cells, and this process was mediated by
autophagy.
However, whether GA could also inhibit the
progression of ccRCC through inducing autophagy remains elusive.
The present study solved this query to a certain extent. In the
present study, it was firstly found that GA inhibited
proliferation, migration and invasion of ccRCC cells (786-O and
ACHN), with showing no or minor damage to the normal renal tubular
epithelial cells (HK-2). In case of GA treatment, HK-2 cells
exhibited elliptical morphology with relatively regular surfaces.
During GA stimulation, there was no significant change in cell
morphology with the increase of GA concentration or the extension
of culture time. The vast majority of cells did not elongate into
irregular shapes, and the intercellular connections remained
intact. Overall, GA stimulation did not significantly destroy the
morphology of HK-2 cells. To some extent, this evaluates the safety
of the drug.
Moreover, GA arrested ccRCC cell cycle in S phase.
The results indicated that GA could selectively inhibit ccRCC
progression in a concentration-dependent manner in vitro. In
addition, the proteins and genes' expression of autophagy markers
as well as autophagosomes formation were further assessed. The
results indicated that GA significantly induced autophagy of ccRCC
cells. Subsequently, the nude mouse xenograft assay and the tumor
metastasis assay revealed that GA significantly inhibited tumor
growth and metastasis, and induced tumor autophagy in a
concentration-dependent manner in vivo.
To further confirm the present findings, 3-MA was
utilized to suppress autophagy and a remarkable reversal was
observed in ccRCC cell proliferation, migration and invasion. The
process of autophagy can be divided into several stages:
Initiation, autophagosome formation, autophagosome maturation and
lysosomal degradation. PI3K plays a crucial role in both
autophagy-mediated lysosomal degradation and vesicular trafficking
during the process of autophagy (42). Knocking down autophagy-related
genes or blocking class III PI3-Kinase is the most common method of
inhibiting autophagy.
3-MA is a classic autophagy inhibitor that exerts
its inhibitory effect by blocking key steps in the formation of
autophagosomes and autophagic vacuoles (43). It was initially discovered by
screening purine-related substances using liver cells isolated from
rats in 1982 (44). It is also
considered the most extensively utilized autophagy inhibitor owing
to its ability to impede the interaction between class III
PI3-Kinase and several ATG partners (45). The potential value of
3-MA-mediated autophagy inhibition in tumor therapy has been
demonstrated in multiple studies (46,47).
The activation of the PI3K/Akt pathway is a known
causal mechanism for tumorigenesis and antitumor therapy. This is
also true in the progression of RCC (48). Finally, in order to explore the
specific mechanism involved in GA inducing ccRCC cells autophagy,
the expression of autophagy-related genes and several critical
proteins was measured, and it was found that GA induced autophagy
of ccRCC cells via the PI3K/Akt/Atg16L1 signaling pathway. All
these results suggested that GA could selectively inhibit
proliferation, migration, and invasion of ccRCC cells as well as
suppress ccRCC growth and metastasis by inducing autophagy via the
PI3K/Akt/Atg16L1 signaling pathway. 3-MA acts as a pharmacological
blocker of class III PI3-Kinase, preventing the formation of
autophagosomes, blocking the autophagy pathway, finally reversing
these effects.
Apoptosis induction is one of the targeted
approaches used in cancer therapy, and serves as the primary
mechanism targeted by chemotherapy drugs and polyphenols. Several
natural compounds that have been previously reported to induce
apoptosis have shown promising potential as candidates, as they
possess the ability to regulate apoptosis through diverse signaling
pathways (49,50).
Mitochondria exert critical roles in in apoptotic
cell death. In a previous study by the authors, it was demonstrated
that GA significantly inhibited viability of T24 bladder cancer
cells in a concentration and time-dependent manner. GA induces
apoptosis of T24 cells via the mitochondrial pathway. GA can
inhibit T24 cells proliferation, metastasis and promote apoptosis,
and the pro-apoptotic activity is closely associated with
mitochondrial dysfunction and PI3K/Akt/NF-κB signaling suppression
(24). In future studies, it
shall be continued to explore the role of GA in apoptosis and to
determine the pathway and relevant components by which GA-induced
apoptosis is mediated through.
In conclusion, the present study identified that
autophagy of ccRCC cells was induced in response to GA treatment.
Subsequently, the in vitro results were translated into an
animal model. The findings in cell research were consistent with
those in animal study, namely, the key markers of autophagy were
all observed in the in vitro and in vivo studies.
Furthermore, the signaling pathway involved in GA-induced autophagy
of ccRCC cells was indeed activated. Hence, it was firstly
presented that GA could inhibit ccRCC progression by inducing
autophagy. GA will be a promising agent, and its application may
provide an effective, independent of apoptosis-resistance treatment
for patients with ccRCC.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TZ, CZL and YS conceived and designed the present
study. TZ, XZ and JL supervised and analyzed the results. YS, TZ
and YF performed the experiments. LZ, DZ, JL and SF analyzed the
data. YS, LZ and JZ contributed analysis tools. TZ, XZ, YS, YF and
SF contributed significantly in writing the manuscript. TZ, CZL and
YS confirm the authenticity of all the raw data. All authors read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved (approval no.
LLSC20232011) by the Ethical Committee of Anhui Medical University
(Hefei, China). All applicable international, national, and/or
institutional guidelines for the care and use of animals were
followed.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The present study was supported by Anhui Medical University
Basic and Clinical Cooperative Research Promotion Project (grant
no. 2020xkjT031) and the National Level Innovation Training Program
for Chinese College Students of the Ministry of Education of the
People's Republic of China (grant no. 202110366035).
References
|
1
|
Petejova N and Martinek A: Renal cell
carcinoma: Review of etiology, pathophysiology and risk factors.
Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 160:183–194.
2016. View Article : Google Scholar
|
|
2
|
Qu Y, Feng J, Wu X, Bai L, Xu W, Zhu L,
Liu Y, Xu F, Zhang X, Yang G, et al: A proteogenomic analysis of
clear cell renal cell carcinoma in a Chinese population. Nat
Commun. 13:20522022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Capitanio U, Bensalah K, Bex A, Boorjian
SA, Bray F, Coleman J, Gore JL, Sun M, Wood C and Russo P:
Epidemiology of renal cell carcinoma. Eur Urol. 75:74–84. 2019.
View Article : Google Scholar
|
|
5
|
Sun Z, Tao W, Guo X, Jing C, Zhang M, Wang
Z, Kong F, Suo N, Jiang S and Wang H: Construction of a
lactate-related prognostic signature for predicting prognosis,
tumor microenvironment, and immune response in kidney renal clear
cell carcinoma. Front Immunol. 13:8189842022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bi K, He MX, Bakouny Z, Kanodia A,
Napolitano S, Wu J, Grimaldi G, Braun DA, Cuoco MS, Mayorga A, et
al: Tumor and immune reprogramming during immunotherapy in advanced
renal cell carcinoma. Cancer Cell. 39:649–661.e5. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wolf MM, Kimryn Rathmell W and Beckermann
KE: Modeling clear cell renal cell carcinoma and therapeutic
implications. Oncogene. 39:3413–3426. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Maiuolo J, Gliozzi M, Carresi C, Musolino
V, Oppedisano F, Scarano F, Nucera S, Scicchitano M, Bosco F, Macri
R, et al: Nutraceuticals and cancer: Potential for natural
polyphenols. Nutrients. 13:38342021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao B and Hu M: Gallic acid reduces cell
viability, proliferation, invasion and angiogenesis in human
cervical cancer cells. Oncol Lett. 6:1749–1755. 2013. View Article : Google Scholar
|
|
10
|
Tuli HS, Mistry H, Kaur G, Aggarwal D,
Garg VK, Mittal S, Yerer MB, Sak K and Khan MA: Gallic acid: A
dietary polyphenol that exhibits anti-neoplastic activities by
modulating multiple oncogenic targets. Anticancer Agents Med Chem.
22:499–514. 2022. View Article : Google Scholar
|
|
11
|
Al Zahrani NA, El-Shishtawy RM and Asiri
AM: Recent developments of gallic acid derivatives and their
hybrids in medicinal chemistry: A review. Eur J Med Chem.
204:1126092020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bai J, Zhang Y, Tang C, Hou Y, Ai X, Chen
X, Zhang Y, Wang X and Meng X: Gallic acid: Pharmacological
activities and molecular mechanisms involved in
inflammation-related diseases. Biomed Pharmacother. 133:1109852021.
View Article : Google Scholar
|
|
13
|
Ashrafizadeh M, Zarrabi A, Mirzaei S,
Hashemi F, Samarghandian S, Zabolian A, Hushmandi K, Ang HL, Sethi
G, Kumar AP, et al: Gallic acid for cancer therapy: Molecular
mechanisms and boosting efficacy by nanoscopical delivery. Food
Chem Toxicol. 157:1125762021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Deng B, Yang B, Chen J, Wang S, Zhang W,
Guo Y, Han Y, Li H, Dang Y, Yuan Y, et al: Gallic acid induces
T-helper-1-like Treg cells and strengthens immune
checkpoint blockade efficacy. J Immunother Cancer. 10:e0040372022.
View Article : Google Scholar
|
|
15
|
Zhang HH, Zhang Y, Cheng YN, Gong FL, Cao
ZQ, Yu LG and Guo XL: Metformin incombination with curcumin
inhibits the growth, metastasis, and angiogenesis of hepatocellular
carcinoma in vitro and in vivo. Mol Carcinog. 57:44–56. 2018.
View Article : Google Scholar
|
|
16
|
Lin Y, Luo T, Weng A, Huang X, Yao Y, Fu
Z, Li Y, Liu A, Li X, Chen D and Pan H: Gallic acid alleviates
gouty arthritis by inhibiting NLRP3 inflammasome activation and
pyroptosis through enhancing Nrf2 signaling. Front Immunol.
11:5805932020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao W, Shi F, Guo Z, Zhao J, Song X and
Yang H: Metabolite of ellagitannins, urolithin A induces autophagy
and inhibits metastasis in human sw620 colorectal cancer cells. Mol
Carcinog. 57:193–200. 2018. View Article : Google Scholar :
|
|
18
|
Cao Y, Chen J, Ren G, Zhang Y, Tan X and
Yang L: Punicalagin prevents inflammation in LPS-induced RAW264.7
macrophages by inhibiting FoxO3a/autophagy signaling pathway.
Nutrients. 11:27942019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang SG, Huang MH, Li JH, Lai FI, Lee HM
and Hsu YN: Punicalagin induces apoptotic and autophagic cell death
in human U87MG glioma cells. Acta Pharmacol Sin. 34:1411–1419.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Klionsky DJ, Petroni G, Amaravadi RK,
Baehrecke EH, Ballabio A, Boya P, Bravo-San Pedro JM, Cadwell K,
Cecconi F, Choi AMK, et al: Autophagy in major human diseases. EMBO
J. 40:e1088632021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ferro F, Servais S, Besson P, Roger S,
Dumas JF and Brisson L: Autophagy and mitophagy in cancer metabolic
remodelling. Semin Cell Dev Biol. 98:129–138. 2020. View Article : Google Scholar
|
|
22
|
Moon S and Jung HS: Endoplasmic reticulum
stress and dysregulated autophagy in human pancreatic beta cells.
Diabetes Metab J. 46:533–542. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li X, He S and Ma B: Autophagy and
autophagy-related proteins in cancer. Mol Cancer. 19:122020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zeng M, Su Y, Li K, Jin D, Li Q, Li Y and
Zhou B: Gallic acid inhibits bladder cancer T24 cell progression
through mitochondrial dysfunction and PI3K/Akt/NF-κB signaling
suppression. Front Pharmacol. 11:12222020. View Article : Google Scholar
|
|
25
|
Khadirnaikar S, Kumar P, Pandi SN, Malik
R, Dhanasekaran SM and Shukla SK: Immune associated LncRNAs
identify novel prognostic subtypes of renal clear cell carcinoma.
Mol Carcinog. 58:544–553. 2019. View Article : Google Scholar
|
|
26
|
Sales MS, Roy A, Antony L, Banu SK,
Jeyaraman S and Manikkam R: Octyl gallate and gallic acid isolated
from Terminalia bellarica regulates normal cell cycle in human
breast cancer cell lines. Biomed Pharmacother. 103:1577–1584. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gu R, Zhang M, Meng H, Xu D and Xie Y:
Gallic acid targets acute myeloid leukemia via Akt/mTOR-dependent
mitochondrial respiration inhibition. Biomed Pharmacother.
105:491–497. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pham HNT, Sakoff JA, Vuong QV, Bowyer MC
and Scarlett CJ: Comparative cytotoxic activity between kaempferol
and gallic acid against various cancer cell lines. Data Brief.
21:1033–1036. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rosman R, Saifullah B, Maniam S, Dorniani
D, Hussein MZ and Fakurazi S: Improved anticancer effect of
magnetite nanocomposite formulation of GALLIC Acid
(Fe3O4-PEG-GA) against lung, breast and colon
cancer cells. Nanomaterials (Basel). 8:832018. View Article : Google Scholar
|
|
30
|
Hong Z, Tang P, Liu B, Ran C, Yuan C,
Zhang Y, Lu Y, Duan X, Yang Y and Wu H: Ferroptosis-related genes
for overall survival prediction in patients with colorectal cancer
can be inhibited by gallic acid. Int J Biol Sci. 17:942–956. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cai L, Wei Z, Zhao X, Li Y, Li X and Jiang
X: Gallic acid mitigates LPS-induced inflammatory response via
suppressing NF-κB signalling pathway in IPEC-J2 cells. J Anim
Physiol Anim Nutr (Berl). 106:1000–1008. 2022. View Article : Google Scholar
|
|
32
|
Zhang P, Ye J, Dai J, Wang Y, Chen G, Hu
J, Hu Q and Fei J: Gallic acid inhibits osteoclastogenesis and
prevents ovariectomy-induced bone loss. Front Endocrinol
(Lausanne). 13:9632372022. View Article : Google Scholar
|
|
33
|
Lee HL, Lin CS, Kao SH and Chou MC: Gallic
acid induces G1 phase arrest and apoptosis of triple-negative
breast cancer cell MDA-MB-231 via p38 mitogen-activated protein
kinase/p21/p27 axis. Anticancer Drugs. 28:1150–1156. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lima KG, Krause GC, Schuster AD, Catarina
AV, Basso BS, De Mesquita FC, Pedrazza L, Marczak ES, Martha BA,
Nunes FB, et al: Gallic acid reduces cell growth by induction of
apoptosis and reduction of IL-8 in HepG2 cells. Biomed
Pharmacother. 84:1282–1290. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ho HH, Chang CS, Ho WC, Liao SY, Lin WL
and Wang CJ: Gallic acid inhibits gastric cancer cells metastasis
and invasive growth via increased expression of RhoB,
downregulation of AKT/small GTPase signals and inhibition of NF-κB
activity. Toxicol Appl Pharmacol. 266:76–85. 2013. View Article : Google Scholar
|
|
36
|
Subramanian AP, Jaganathan SK, Mandal M,
Supriyanto E and Muhamad II: Gallic acid induced apoptotic events
in HCT-15 colon cancer cells. World J Gastroenterol. 22:3952–3961.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Boya P, Reggiori F and Codogno P: Emerging
regulation and functions of autophagy. Nat Cell Biol. 15:713–720.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Das S, Shukla N, Singh SS, Kushwaha S and
Shrivastava R: Mechanism of interaction between autophagy and
apoptosis in cancer. Apoptosis. 26:512–533. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tedesco I, Russo M, Bilotto S, Spagnuolo
C, Scognamiglio A, Palumbo R, Nappo A, Iacomino G, Moio L and Russo
GL: Dealcoholated red wine induces autophagic and apoptotic cell
death in an osteosarcoma cell line. Food Chem Toxicol. 60:377–384.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu MP, Liao M, Dai C, Chen JF, Yang CJ,
Liu M, Chen ZG and Yao MC: Sanguisorba officinalis L
synergistically enhanced 5-fluorouracil cytotoxicity in colorectal
cancer cells by promoting a reactive oxygen species-mediated,
mitochondria-caspase-dependent apoptotic pathway. Sci Rep.
6:342452016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dong Y, Yin S, Jiang C, Luo X, Guo X, Zhao
C, Fan L, Meng Y, Lu J, Song X, et al: Involvement of autophagy
induction in penta-1, 2,3,4,6-O-galloyl-β-D-glucose-induced
senescence-like growth arrest in human cancer cells. Autophagy.
10:296–310. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Iershov A, Nemazanyy I, Alkhoury C, Girard
M, Barth E, Cagnard N, Montagner A, Chretien D, Rugarli EI, Guillou
H, et al: The class 3 PI3K coordinates autophagy and mitochondrial
lipid catabolism by controlling nuclear receptor PPARα. Nat Commun.
10:15662019. View Article : Google Scholar
|
|
43
|
Zhao F, Feng G, Zhu J, Su Z, Guo R, Liu J,
Zhang H and Zhai Y: 3-Methyladenine-enhanced susceptibility to
sorafenib in hepatocellular carcinoma cells by inhibiting
autophagy. Anticancer Drugs. 32:386–393. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Seglen PO and Gordon PB: 3-Methyladenine:
Specific inhibitor of autophagic/lysosomal protein degradation in
isolated rat hepatocytes. Proc Natl Acad Sci USA. 79:1889–1892.
1982. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Vinod V, Padmakrishnan CJ, Vijayan B and
Gopala S: 'How can I halt thee?' The puzzles involved in autophagic
inhibition. Pharmacol Res. 82:1–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li J, Hou N, Faried A, Tsutsumi S,
Takeuchi T and Kuwano H: Inhibition of autophagy by 3-MA enhances
the effect of 5-FU-induced apoptosis in colon cancer cells. Ann
Surg Oncol. 16:761–771. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li J, Yang D, Wang W, Piao S, Zhou J,
Saiyin W, Zheng C, Sun H and Li Y: Inhibition of autophagy by 3-MA
enhances IL-24-induced apoptosis in human oral squamous cell
carcinoma cells. J Exp Clin Cancer Res. 34:972015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Guo H, German P, Bai S, Barnes S, Guo W,
Qi X, Lou H, Liang J, Jonasch E, Mills GB and Ding Z: The PI3K/AKT
pathway and renal cell carcinoma. J Genet Genomics. 42:343–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rajabi S, Maresca M, Yumashev AV, Choopani
R and Hajimehdipoor H: The most competent plant-derived natural
products for targeting apoptosis in cancer therapy. Biomolecules.
11:5342021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chimento A, De Luca A, D'Amico M, De
Amicis F and Pezzi V: The involvement of natural polyphenols in
molecular mechanisms inducing apoptosis in tumor cells: A promising
adjuvant in cancer therapy. Int J Mol Sci. 24:16802023. View Article : Google Scholar : PubMed/NCBI
|