1. Introduction
Breast cancer (BC) is the most commonly diagnosed
type of cancer among women globally, with >2.3 million cases
identified in 2020. In that same year, ~685,000 women succumbed due
to BC, making it the leading cause of cancer-related death in women
(1,2). BC is classified into four subtypes
based on the expression of human epidermal growth factor receptor 2
(HER2/neu), progesterone receptor and estrogen receptor
triple-negative breast cancer (TNBC), namely as HER2-positive,
Luminal A and Luminal B. The treatment options for BC primarily
involve surgical interventions, chemotherapy, radiation therapy,
hormonal therapy and immunotherapy (3). It is crucial to tailor the treatment
approach to the subtype, stage and specific characteristics of the
cancer, alongside considering the overall health and preferences of
the patient. During BC treatment, patients commonly face
challenges, such as side-effects, including nausea, hair loss,
fatigue, neuropathy and an elevated susceptibility to infections.
Chemotherapy (4,5) and certain treatments, such as
trastuzumab (Herceptin), may provoke adverse reactions with
potential cardiac toxicity (6,7). Despite
the increasing importance of immunotherapy in the treatment of
various types of cancer, its efficacy in BC remains limited,
benefiting only a small fraction of patients (8). Moreover, there have been reports of
immune-related adverse events linked to the use of immune
checkpoint inhibitors, often resembling autoimmune diseases, but
with a more acute onset and potential for severe, lasting effects.
The complexity and heterogeneity of BC pose significant challenges
for immunotherapies, particularly immune checkpoint inhibitors
(9,10). Without validated targeted therapies,
chemotherapy remains the standard treatment strategy for BC
(11). However, alongside known
adverse effects, the development of drug resistance is a
significant issue, leading to disease progression and increased
mortality rates. Recent research efforts have focused on overcoming
resistance by exploring combinations of cytotoxic, targeted and
immune-based therapies (12).
Recognizing the crucial role of the tumor microenvironment (TME) in
drug resistance underscores the importance of understanding tumor
immunity for developing effective immunotherapeutic strategies
against BC.
Myeloid-derived suppressor cells (MDSCs) are
critical components of the TME, playing a crucial role in
suppressing immune responses in cancer, infections and inflammatory
diseases. Originating from immature myeloid cells (IMCs), MDSCs
exhibit marked heterogeneity, comprising pathologically activated
neutrophils and macrophages (13).
Recent investigations underscore their dual function in suppressing
antitumor immune responses, while stimulating tumor progression.
MDSCs promote tumor angiogenesis, facilitate tumor cell invasion
and contribute to the formation of premetastatic niches (14,15).
MDSC levels are closely associated with clinical outcomes and
therapeutic efficacy in patients with BC (16).
In BC, MDSC accumulation in the TME and peripheral
circulation is notable, driven by the modulation of
immunosuppressive mechanisms, predominantly through T-cell
activation inhibition (17), along
with the secretion of multiple cytokines and non-immunosuppressive
pathways (18). This collective
action promotes tumor growth by enabling tumor angiogenesis,
enhancing invasion and metastasis, and modifying the TME to favor
tumor progression (19). MDSCs play
diverse roles in promoting tumor development by impeding the immune
system. Given their pivotal role in subverting the body's antitumor
defenses, MDSCs are increasingly recognized as promising targets
for therapeutic interventions, including innovative approaches such
as nanotechnology.
The present review discusses the immunosuppressive
functions of MDSCs, their role in BC, and strategies for targeting
them in cancer therapy.
2. MDSCs in BC
MDSCs derived from patients with BC exhibit
functional and phenotypic similarities to those originating from
bone marrow, indicating their myeloid precursor origin (13). These MDSCs are categorized into
monocytic (M)-MDSCs (CD11b+ Ly6G-
Ly6Chigh) and granulocytic (G)-MDSC (CD11b+
Ly6G+ Ly6Clow) subpopulations (20,21). In
humans, M-MDSCs are characterized by
CD11b+CD33+HLA-DR-/low
CD14+ CD15- markers, while G-MDSCs express
CD11b+ CD33+ HLA-DR-/low
CD14- CD15+ (or CD66b+) markers
(20,22).
MDSC development is regulated by a network of
signals that promote the growth of IMCs (23). Various signaling pathways and
regulators, such as the signal transducer and activator of
transcription (STAT) family, interferon (IFN) regulators, Notch,
adenosine receptor A2b and NLRP3, facilitate myelopoiesis, inhibit
the maturation and differentiation of progenitor cells, and expand
the IMCs. Additionally, signaling pathways and regulators,
including NF-κB, STAT1, STAT6, prostaglandin E2 (PGE2),
cyclooxygenase-2 and the endoplasmic reticulum stress response
contribute to the development of an immunosuppressive phenotype
that leads to the pathological activation of immature cells
(23,24).
Factors, such as granulocyte colony-stimulating
factor (G-CSF), granulocyte-macrophage colony-stimulating factor
(GM-CSF) (24-26),
vascular endothelial growth factor (VEGF), PGE2, interleukin (ILs)s
[IL-1 (27-29),
IL-6, IL-13, IL-17, IL-20, IL-33, IL-34 (30-35)],
macrophage migration inhibitory factor (MIF) (36), along with microRNAs (miRNAs/miRs),
contribute to MDSC amplification and activation in BC (37). Of note, patients with BC with MDSCs
often exhibit higher levels of psychological stress, likely
influenced by stress-related hormones and cytokines, such as
IL-1Ra, IFN-γ-induced protein 10, G-CSF and IL-6, further
stimulating MDSC production and accumulation (38) (Fig.
1).
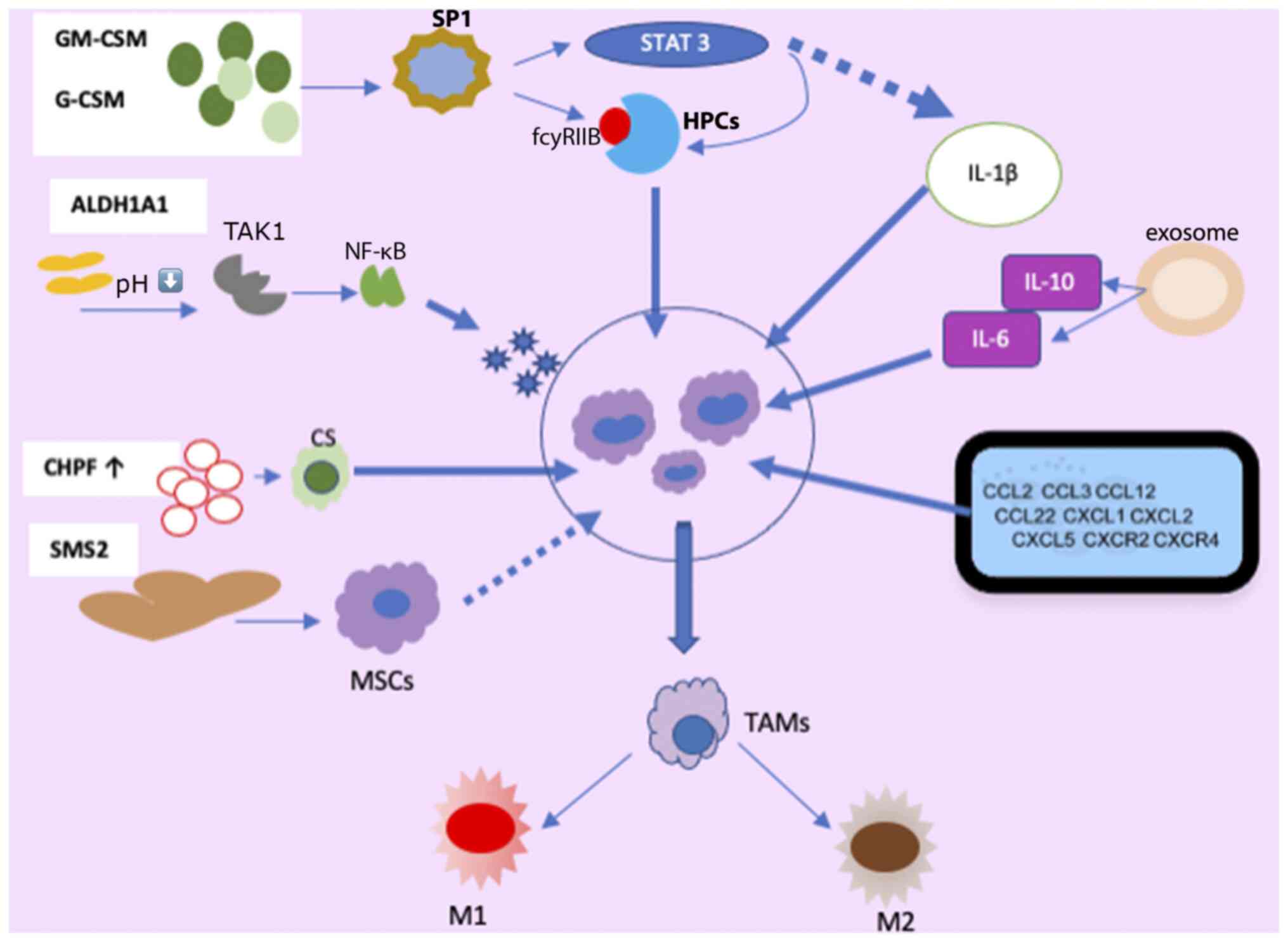 | Figure 1Mechanisms involved in the
production, activation, recruitment and differentiation of MDSC in
BC. MDSCs, myeloid-derived suppressor cells; HPCs, hematopoietic
progenitor cells; SP1, specific protein 1; GM-CS,
granulocyte-macrophage colony-stimulating factor; G-CSF,
granulocyte colony-stimulating factor; ALDH1A1, aldehyde
dehydrogenase 1A1; TAK1, TGF-β-activated kinase 1; CS, chondroitin
sulfate; CHPF, chondroitin polymerase factor; SMS2, sphingosine
synthase 2; MSCs, mesenchymal stem cells; TAMs, tumor-associated
macrophages; M1, M1 macrophages; M2, M2 macrophages. |
G-CSF and GM-CSF, derived from tumor cells, play
pivotal roles in MDSC accumulation (23,25).
MDSCs originate from immature, multipotent hematopoietic progenitor
cells (HPCs) and respond to signals from the host and tumor cells,
particularly through the secretion of GM-CSF, and are subsequently
recruited to the TME (39).
Among the receptors implicated in MDSC regulation,
FC gamma receptor IIB (FCγRIIB/CD32B) is the sole inhibitory member
expressed on B-cells, macrophages, dendritic cells (DCs) and
granulocytes, with an upregulation of its expression observed in
tumor-infiltrating MDSCs (40).
GM-CSF increases FcγRIIB expression in HPCs by activating
specificity protein family 1 (SP1) transcription factors, which
bind to GC-rich motifs, thereby regulating the expression of genes
involved in proliferation, apoptosis, differentiation, and immune
responses. The inhibition of SP1 dampens MDSC differentiation and
infiltration into the TME. However, when SP1 and FCγRIB are
activated, they promote MDSC generation from HPCs via STAT3, a
member of the STAT family of transcription factors activated by
tyrosine kinases in response to various cytokines and growth factor
receptors (41,42). Consequently, tumor cell-induced
activation of GM-CSF-driven Sp1 and STAT3 cooperatively trigger the
expression of target genes, facilitating the immunosuppressive
functions of MDSCs. Moreover, chondroitin polymerase factor,
frequently overexpressed in BC tissues, enhances G-CSF binding to
cell surface chondroitin sulfate, thereby promoting MDSC
accumulation (43).
The enzymatic activity of aldehyde dehydrogenase 1A1
(ALDH1A1) is crucial in reducing intracellular pH in BC cells. This
condition triggers increased TGF-β-activated kinase 1 (TAK1)
phosphorylation, subsequently activating the NF-κB pathway.
Consequently, GM-CSF secretion is induced, amplifying MDSCs and
fostering BC progression (44).
Exosomes released by 4T1 (syngeneic cell lines derived tumor
models) cells contain IL-6 and IL-10, which enhance MDSC
stimulation and proliferation by promoting STAT3 phosphorylation in
myeloid cells. This diminishes myeloid proliferation and death,
expediting differentiation into MDSCs (37).
The study by Jiang et al (45) revealed that tumor exosome-secreted
miR-9 and miR-181a targeted suppressor of cytokine signaling
protein 3 (SOCS3) and separately activated STAT3 (PIAS3),
triggering the JAK/STAT signaling cascade. Prolonged SOCS3
suppression and abnormal JAK/STAT pathway upregulation lead to
early-stage MDSC accumulation (46).
A number of molecules in signaling pathways form intricate
regulatory loops. In BC cells, the mammalian target of the
rapamycin pathway induces MDSC accumulation by modulating G-CSF
expression. MDSCs reciprocally increase tumor-initiating cell
frequency by activating the Notch signaling pathway in tumor cells,
which secretes G-CSF, establishing a feed-forward loop promoting
MDSC expansion (47). Autocrine
secretion of GM-CSF and IL-33 in the TME sustains MDSC viability by
inhibiting apoptosis, promoting a positive feedback loop that
induces MDSC accumulation (34).
Mechanisms of the recruitment of MDSCs
in BC
Several factors, including chemokines, cytokines,
and complements produced by both tumor and normal cells, induce the
recruitment of MDSCs into tumor tissue (48). Among these, lung fibroblasts secrete
chemokine (C-X-C motif) ligand (CXCL)1, which promotes an
immunosuppressive lung microenvironment by attracting granulocytic
MDSCs and facilitating the formation of BC metastatic niches in the
anterior lung (49).
BC exosomes carry elevated levels of miR-200b-3p,
which are taken up by alveolar epithelial type II cells, directly
impacting phosphatase and tensin homolog (PTEN). PTEN suppression
activates the AKT/NF-κB-p65 pathway, increasing chemokine (C-C
motif) ligand 2 (CCL2) expression and attracting MDSCs, ultimately
promoting lung metastasis in BC (50). Endoplasmic reticulum oxireductin 1a,
a disulfide oxidase located in the endoplasmic reticulum and
closely associated with tumors, participates in the oxidative
folding process, generating and attracting granulocytic MDSCs, and
contributes to G-CSF, CXCL1 and CXCL2 production (51). TGF-β1 upregulates miR-494 levels in
MDSCs, enhancing MDSC movement via CXCR4(52). The transcription factor, ΔNp63,
directly regulates CXCL2 and CCL22, facilitating MDSC recruitment
in TNBC (53).
Liver cells contribute to MDSC recruitment at
specific sites by producing S100A8, facilitating BC metastasis
(54). Chen et al (55) found that VEGF-C produced by breast
cancer cells was responsible for increasing the levels of
chemokines produced by lymphatic endothelial cells (LECs). This, in
turn, helped recruit MDSCs to the TME and lymph nodes (LNs) through
the CXCR2 pathway (55). Evidence
suggests that in the presence of 4T1 cells (breast cancer cell line
derived from the mammary gland tissue of a mouse BALB/c strain),
interstitial fluid migration alongside LECs aids MDSC
dissemination. Moreover, reduced levels of vascular VEGFR3 decrease
the flow response in MDSCs and 4T1 cells (56). The acetylation of the SMAD3 protein,
dependent on the epigenetic regulator KAT6A, contributes to MDSC
recruitment and TNBC metastasis through epigenetic regulation
(57).
The activation of the complement system,
particularly through C5a signaling, plays a pivotal role in
recruiting MDSCs into the TME and suppressing CD8+ T
cell-mediated tumor elimination. Consequently, lung angiogenesis is
fostered in tumor-bearing mice, facilitating BC metastasis to this
organ (58). Cheng et al
(59) demonstrated that periodontal
inflammation (PI) activation enhances the expression of chemokines
such as CCL5, CXCL12, CCL2 and CCL5, which recruit MDSCs, thereby
promoting the establishment of premetastatic niches at sites of
inflammation. MDSCs exhibit diverse differentiation pathways
regulated by various transcription factors, as depicted in Fig. 1. They can differentiate into
tumor-associated macrophages (TAMs) and dendritic cells, which
further stimulate the production of inflammatory DCs (60).
During BC progression, MDSCs transition into TAMs.
Under environmental pressures, such as hypoxia, M-MDSCs
differentiate into TAMs upon migration to specific tissues. TAMs,
in turn, can adopt either the M1 phenotype, characterized by
pro-inflammatory and antitumor properties, or the M2 phenotype,
which exhibits protumor functions, in response to stimuli such as
lipopolysaccharide (LPS), TNF-α and IFN-γ (61).
Macrophages and MDSCs are ubiquitous in the majority
of solid tumors, driving immune suppression and inflammation
(61). Their interaction increases
IL-10 production by MDSCs and decreases IL-12 production by
macrophages, polarizing the immune system toward a type 2 protumor
environment (62). IL-33 is known
for its stimulatory effects on myeloid and lymphoid cells,
promoting the production of type 2 cytokines. The activation of ST2
in type 2 innate lymphoid cells (ILC2) triggers the release of type
2 cytokines IL-33 and IL-13(63).
ILC2 are tissue-resident lymphocytes with various functional roles
in mucosal immunity. In tumor immunology, ILC2 is crucial in DC
recruitment via CCL5 production and activation through IL-9 and
IL-13 secretion (64). IL-33 further
stimulates ST2+ regulatory T-cells (Tregs) and
amphiregulin (AREG) expression, enhancing immune regulatory
functions and tissue repair (65).
The activation of ILC2 through IL-33 supports type 2 immune
responses and M2 reparative macrophages. ST2 is also expressed in
myeloid-derived antigen-presenting cells, such as macrophages and
CD11b+ CD11c+ DCs (66). Moreover, IL-33 triggers IL-2 release
and fosters Treg cell expansion. The type, density and spatial
distribution of these IL-33-modulated immune cells within tumors
profoundly influence tumor behavior (67). Thus, the cytokine IL-33 stimulates
the upregulation of IL-13 while concurrently suppressing IL-12
levels. This immune profile underlies the negative impact of M2
macrophages and Th2 cell polarization within the TME on antitumor
immunity (34).
TAMs are the most abundant immune cells involved in
regulating breast cancer progression. The TME contains many
immunosuppressive cells (68).
Macrophages exhibit heterogeneity, with at least two functionally
distinct subtypes responding to different stimuli: classically
activated M1 macrophages and alternatively activated M2 macrophages
(69). M1 macrophages eliminate
tumors directly by recognizing and phagocytizing cancer cells and
indirectly by producing pro-inflammatory cytokines, such as IFN-γ
and IL-12. Conversely, in the context of tumor development, M2
macrophages are regarded as ‘tumor promoters’. They facilitate
cancer progression, promote tumor cell metastasis and angiogenesis,
regulate energy metabolism and aid in immune system evasion
(70). During tumor progression, M2
macrophages become more prevalent, eventually dominating the TAM
population in the TME. The underlying mechanism suggests that TAMs
promote and sustain cancer stem cells, thereby supporting tumor
growth and self-renewal. Various cytokines and signaling pathways
within the TME influence the polarization of macrophages into M1 or
M2 types. When IL-4 binds to its receptor, it can promote the
phosphorylation of STAT6, leading to the polarization of M2-like
macrophages through the JAK/STAT6 signaling pathway (68,71).
Phosphorylated STAT6 can bind to Krüppel-like factor 4 (KLF4) and
peroxisome proliferator-activated receptor γ, further promoting
this polarization. Furthermore, various signals, including TGF-β,
IL-10, bone morphogenetic protein-7 and IL-4 itself, induce M2
polarization through the PI3K/Akt signaling pathway. The complex
formed by CCAAT/enhancer binding protein α and KLF6 is also
associated with the switch from the M1 to the M2 phenotype
(71,72). The progression of BC can also be
observed through the promotion of monocytic MDSC differentiation
into immunosuppressive M2-polarized macrophages, facilitated by
both sphingosine synthase 2 and exosomes secreted by mesenchymal
stem cells (61,73). Doxorubicin (DOX)-resistant tumor
cells release PGE2, activating the EP2-EP4/cAMP/PKA signaling
cascade in MDSCs, fostering proliferation and an M2 polarized
phenotype shift. This polarization is mediated through miR-10a
induction (27). Studies have shown
that natural killer (NK) T-cells (NKT cells) help transform
CD11b+ HLA-DR MDSCs into CD11b low HLA-DR DCs (74).
Immunosuppressive effects of MDSCs on
BC progression
Fig. 2 shows how
MDSCs significantly impede the activity of tumor-fighting T- and
B-cells within the TME, particularly cytotoxic T-lymphocytes (CTLs)
and pro-inflammatory cells, such as NK cells. Additionally, MDSCs
can promote cancer progression by inducing the generation of Tregs
and T-helper 17 (Th17) cells, thereby altering the local
environment to promote tumor growth and enable immune evasion
(75).
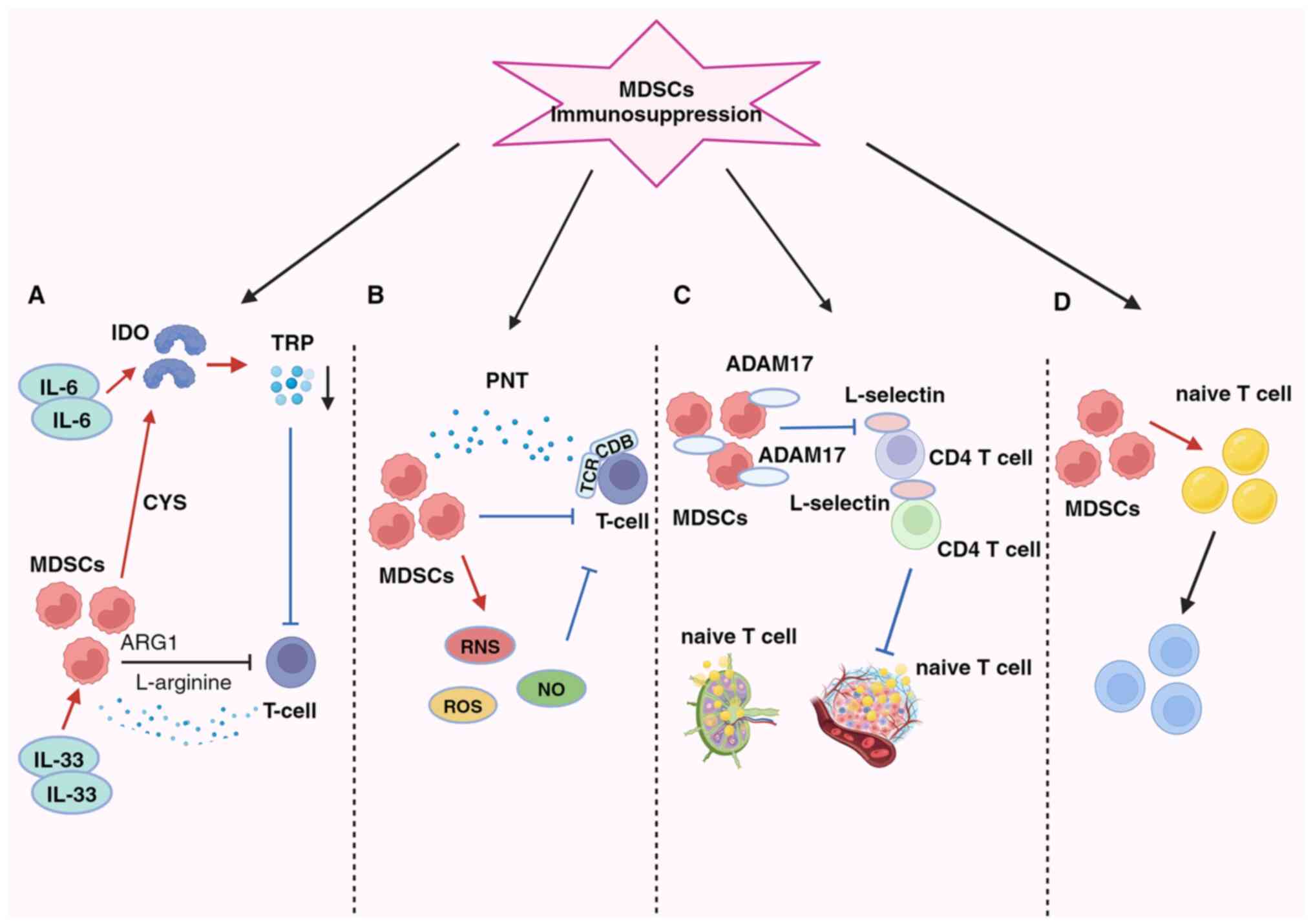 | Figure 2Presence of MDSCs in BC and their
diverse immunosuppressive effects (A) MDSCs suppress T-cells by
activating IDO, resulting in the depletion of essential nutrients,
such as ARG1, TRP and cysteine (CYS). (B) MDSCs induce oxidative
stress by generating ROS, RNS and NO, leading to T-cell
suppression. (C) MDSCs impede lymphocyte migration through direct
physical contact and the expression of ADAM17. (D) MDSCs disrupt
the conversion of naïve CD4+ T-cells into Tregs. MDSCs,
myeloid-derived suppressor cells; PNT, peroxynitrite; TCR, T-cell
receptor; IDO, indoleamine 2,3-dioxygenase; ARG1, arginase 1; TRP,
tryptophan; CYS, cysteine; ROS, reactive oxygen species; RNS,
reactive nitrogen species; NO, nitric oxide; ADAM17, a disintegrin
and metalloproteinase domain 17. |
The primary immunosuppressive mechanism in the BC
microenvironment involves the inhibition of T-cell function, which
is the main trigger process involving the depletion of vital
nutrients (Fig. 2A) (76). MDSCs exert inhibitory effects by
activating indoleamine 2,3-dioxygenase (IDO), reducing local
tryptophan availability, and generating cytotoxic metabolites, such
as kynurenine in the TME and lymphatic drainage regions. This leads
to an increase in Tregs, the inhibition of immune responses against
antigens and the suppression of tumor-specific CTLs (77). The activation of STAT3-dependent
NF-κB by IL-6 is responsible for maintaining IDO overexpression
(30,77). Additionally, MDSCs enhance the
suppressive action of IL-33 on T-cells by depleting L-arginine via
arginase 1 (ARG1) (33,75). Furthermore, MDSCs consume cysteine,
essential for T-cell activation and optimal function, leading to
its depletion and failure to replenish in their environment
(78). The generation of oxidative
stress is a key factor in the second mechanism (Fig. 2B).
By producing reactive oxygen species, reactive
nitrogen species and nitric oxide (NO), MDSCs suppress T-cells in
the TME. MDSCs induce immune tolerance by T-cell receptor and
CD8+ surface modifications and generate the free radical
peroxynitrite (79). According to
Stiff et al (80),
MDSC-generated NO also disrupts Fc receptor-mediated NK cell
activity, reducing monoclonal antibody efficacy and impeding the
immune response against tumors. The third mechanism occurs by
preventing lymphocyte migration (Fig.
2C). MDSCs reduce the immune response in peripheral lymphoid
organs and accumulate in sentinel LNs, where they impede
CD3/CD28-induced T-cell proliferation through contact-dependent
mechanisms. This process supports tumor progression and metastasis
(80,81). Hanson et al (82) linked decreased L-selectin expression
on CD4+ and CD8+ T-cells with a disintegrin
and metalloproteinase domain 17 generation on the plasma membrane.
Consequently, MDSCs in BC hinder immature T-cell activation and
migration into LNs and their subsequent transport to tumors,
ultimately compromising the immune system's capacity to combat
tumors (82).
The fourth aspect pertains to the expansion and
activation of Tregs, facilitated by MDSCs via promoting their
proliferation and differentiation of naïve CD4+ T cells
(Fig. 2D). Although this pathway
mechanism is not yet fully understood, it is known that Tregs can
infiltrate tumors and play a crucial role in the antitumor
immunosuppressive response. IL-34 triggers the conversion of bone
marrow stem cells into monocytic MDSCs, indirectly suppressing the
immune response by fostering Treg attraction via CCL22 secretion in
the TME, thus contributing to chemotherapy resistance (35). Additionally, BC-induced MDSCs can
stimulate effector T-cells to transition into Tregs through the IDO
mechanism (77). MDSCs suppress
T-cell activation, and once activated, T-cells trigger MDSC
apoptosis via the Fas-FasL pathway (83).
MDSCs in BC employ contact-dependent mechanisms and
indirect means, releasing NO, ARG and IL-1 to suppress the response
of B-cells against tumors (84).
Additionally, MDSCs can transform ordinary B-cells into specialized
immunomodulatory B-cell (Breg) phenotypes, which efficiently
suppress T-cell responses (85).
Furthermore, various mediators in the TME, including LPS, can
induce programmed cell death protein 1 (PD-1) expression in MDSCs
in BC (86). By activating the
PI3K/PKB/NF-κB signaling pathway in B-cells, MDSCs can enhance
immune evasion mediated by PD-1/PD-L1 Bregs through the PD-1
pathway (87,88).
BC cells cultured under hypoxic conditions secrete
various cytokines, including monocyte chemotactic protein-1, which
triggers MDSC recruitment and suppresses the cytotoxic activity of
NK cells. These mechanisms collectively contribute to cancer
metastasis facilitation (89). NKT
cells can restore suppressed T-cell function by converting
CD11b+ HLA-DR MDSCs into CD11b low HLA-DR DCs through an
NKG2D-dependent signaling mechanism (74). The function of MDSCs is further
influenced by C5aR signaling, regulating CD4+ T-cell
polarization towards the Th2 phenotype in the lungs of
tumor-bearing mouse (58). The
administration of DOX increases miR-126 exosomes derived from
MDSCs, thereby suppressing T-cell functionality, inhibiting Th1
cell activation, and initiating Th2 cell responses in mouse lungs
(90).
Clinical aspects of MDSCs related to
BC
MDSC levels are associated with the progression of
BC, typically showing higher levels in more advanced cancer stages
(22). Additionally, surgical stress
induced by the excised primary tumor environment can trigger the
recruitment of MDSCs to lung and tumor tissues, underscoring the
importance of monitoring MDSC levels (91). Data indicate variations in MDSC
levels among patients undergoing antitumor treatment, potentially
reflecting individual responses to therapy. In neoadjuvant therapy,
circulating granulocytic MDSCs may initially increase, decrease
with DOX and cyclophosphamide administration, and return close to
baseline levels during paclitaxel treatment. Conversely, in
metastatic or recurrent BC, monocytic MDSCs undergo significant
expansion in the peripheral circulation, being associated with
increased degrees of metastases to LNs and other organs (92). Therefore, tracking M-MDSC levels in
patients with BC may be a valuable biomarker for monitoring cancer
progression and treatment response.
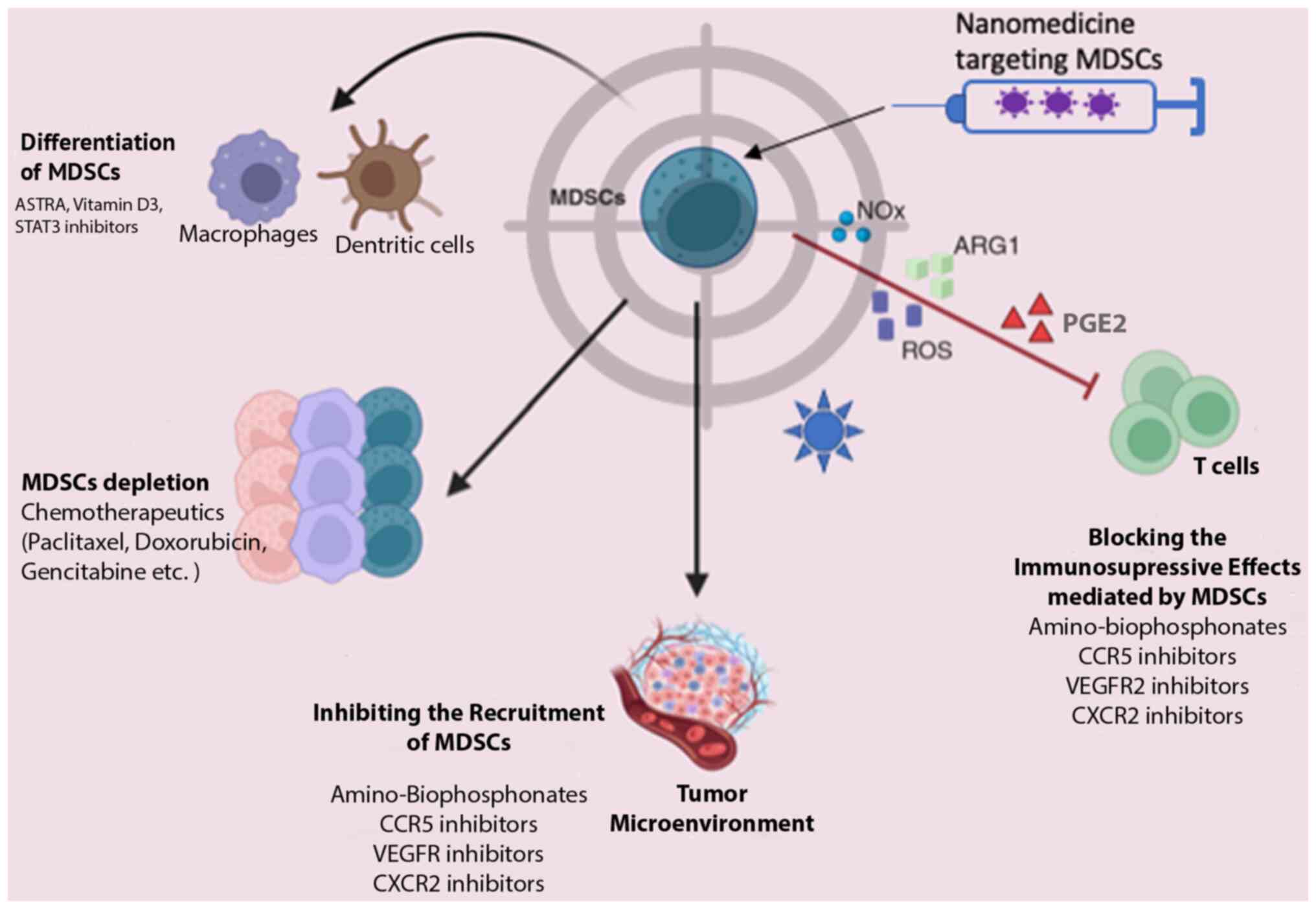
BC treatments targeting MDSCs
MDSCs play a pivotal role in BC progression and are
intricately linked to tumor immune evasion. Consequently, MDSCs are
a promising target for tumor immunotherapy, primarily aimed at
enhancing host immunity. Currently, therapeutic strategies
targeting MDSCs encompass four main approaches, as illustrated in
Fig. 3: MDSC depletion, the blockade
of MDSC recruitment, the suppression of MDSC immunosuppressive
function and the induction of MDSC differentiation into a
non-suppressive immune state (93-95).
While no specific selective inhibitors of MDSCs have
been identified to date, at least to the best of our knowledge,
several existing drugs exert indirect effects on MDSCs. For
instance, DNA methyltransferase inhibitors and histone deacetylase
(HDAC) inhibitors modulate systemic and intratumoral MDSCs,
enhancing the long-term response to immunotherapy (96). Other drugs with the potential to
suppress or deplete MDSCs and consequently enhance immunotherapy
efficacy include gemcitabine (97),
DOX (98) and 5-fluorouracil
(99). Additionally, combined
therapeutic strategies are under investigation, such as the
combination of Romidepsin (an HDAC inhibitor) with cisplatin and
nivolumab in TNBC (NCT02393794) (100) and IPI-549 (an inhibitor of PI3Kδ
and PI3Kγ isoforms that decreases MDSCs and enhances anti-PD-1
efficacy) with nivolumab in solid tumors (101).
Curcumin is known for its antitumor properties,
primarily attributed to its inhibition of IL-6. However, research
has revealed that its mechanism also entails the inhibition of
MDSCs in both blood and tissues, thus impeding tumor growth
(102). Numerous preclinical and
clinical studies are also dedicated to exploring strategies for
promoting MDSC maturation (103-105).
Sulforaphane, an inhibitor of the inflammatory cytokine MIF,
disrupts its protumor functions, including the facilitation of MDSC
differentiation in the TME. In vitro research has shown that
MIF inhibitors, such as sulforaphane inhibit the accumulation of
MDSCs in the TME, blocking their differentiation and restoring
antitumor immunological activity (36). Silibinin modulates CCR2 expression in
MDSCs, resulting in decreased MDSC accumulation in blood and tumor
tissue (106).
NG-monomethyl-L-arginine acetate, an inducible NO synthase
inhibitor, blocks MDSC differentiation into osteoclasts,
potentially preventing MDSC-mediated BC bone metastasis and
associated bone degradation (107).
Activated T-cells (ATCs) combined with bispecific anti-CD3 x
anti-Her2/neu antibodies (aATCs) effectively modulate MDSCs via INF
and IL-2, suppressing their activity and inhibiting tumor growth
(108). According to Thakur et
al (109), aATCs suppress the
actions and functions of MDSCs (via IFN and IL-2) and effectively
inhibit tumor growth and Treg differentiation.
Therapeutic combinations, such as adoptive cell
therapy involving sensitized immune and tumor cells reprogrammed
with CD25+ NKT, NK and memory T-cells, have achieved the
immunosuppression of MDSCs (110).
In other strategies tested in preclinical models, a vaccine
composed of Listeria monocytogenes has been investigated.
When MDSCs are infected with a highly attenuated bacterium,
Listeria monocytogenes (Listeriaat), their
immunosuppressive function is altered. Moreover,
Listeriaat-infected MDSCs, which primarily deliver Listeriaat to
the microenvironment of metastases and primary tumors, spread from
MDSCs to tumor cells. Consequently, Listeriaat immunotherapy
significantly reduces the population of MDSCs and can convert MDSCs
into an immunostimulatory phenotype that produces IL-12, while
concurrently reducing metastasis and tumor growth (111).
3. Potential nanotechnology-based
therapeutics through MDSC targeting
Nanotechnology-based drug delivery systems are the
emerging approaches in cancer therapy, characterized by intensive
exploration (112,113). Despite the biological and
functional barriers posed by the TME, nanoparticles (NPs) have
demonstrated efficacy in enhancing intertumoral drug accumulation
through passive or active targeting mechanisms. This optimized
biodistribution reduces side-effects and increases therapeutic
benefits (114,115). While drug delivery platforms
targeting MDSCs in cancer treatment (Fig. 3) are relatively new, their potential
is promising, particularly in BC.
As examples of successful results, Zhang et
al (116) used ursolic acid, a
natural pentacyclic triterpenoid known for its antifungal,
antibacterial and recently discovered immunomodulatory properties,
encapsulated within liposomes to modulate the TME. Following five
administrations, treatment with this liposomal formulation led to a
significant reduction in biomarker levels across the bloodstream,
spleen, and tumor sites. This was coupled with enhanced cytotoxic
T-cell activity and consequent reductions in tumor volumes
(116). Chen et al (117) used gemcitabine-loaded nanocages in
combination with IDO-targeted small interfering RNA and PD-L1
antibody designed on the nanocarrier surface. This combinatorial
approach aims to improve immunosuppression and overall treatment
outcomes in patients with TNBC. The administration of these
tri-loaded nanocarriers (GSZMP) in TNBC mice was shown to result in
a significant decrease in MDSC proportions compared to the
controls, accompanied by increased T-cell infiltration.
Additionally, the GSZMP group exhibited significant tumor volume
reduction and increased survival rates compared to the control
group (118).
Using this targeted delivery strategy to the tumor
site, a nanoparticle formulation comprising c-peptide (RGDfk) in
low molecular weight heparin-retinoic acid (LMWH-ATRA) micelles
loaded with DOX and the immune adjuvant α-galactosyl ceramide (αGC)
(RLA/DOX/αGC NP) was proposed. The hydrophilic segment of LMWH
selectively bound to P-selectin present on vascular endothelial
cells impedes the recruitment of MDSCs. The hydrophobic ATRA
segment facilitated MDSC depletion, inducing their differentiation.
This multidimensional approach effectively modulated MDSCs,
significantly improving the inflammatory and immunosuppressive
microenvironment in the lungs and tumor sites while inhibiting NPM
formation. The micelles exhibited synergistic effects with other
components in their composition (notably αGC), enhancing overall
antitumor immunity. Thus, this formulation is a promising
therapeutic avenue for addressing BC and lung metastases (119).
4. Conclusions and future perspectives
There is evidence to suggest a heightened prevalence
of MDSCs in patients with BC, indicating their crucial role in the
immune-resistant characteristics of the disease. Given the diverse
nature of MDSCs, there is a pressing need for assays that can
accurately identify MDSC subtypes in patients with BC. It is
imperative to evaluate MDSC levels in both peripheral blood and the
TME across various stages and subtypes of BC. Such assessments
would provide insight into MDSC generation, expansion, and their
function in peripheral blood and the TME, thereby elucidating the
connection between MDSCs and the advancement of BC stages. The
present review aimed to facilitate the practical application of
these research findings and lay the foundation for the diagnosis
and treatment of MDSC-related aspects of BC. MDSCs are pivotal in
advancing tumor growth and metastasis through complex
mechanisms.
Immunotherapy is a promising therapeutic approach in
cancer treatment, demonstrating increased survival rates in
preclinical models and clinical settings. Until recently,
immunotherapy was not considered a viable option for BC treatment
as BC was long-viewed as a poorly immunogenic tumor type (120). However, increasing evidence in
recent years indicating immunogenic activity across various BC
subtypes has shifted this paradigm, highlighting immunotherapy as
an increasingly important tool in BC treatment (121).
Clinical trials are underway to combine
immunotherapy with other therapeutic modalities in BC to target
MDSCs (109,110). This approach is justified as MDSCs
are essential in the BC microenvironment, promoting tumor growth
and metastasis. Thus, MDSC-targeted therapies are promising as
potential treatments in clinical settings. Proposed strategies to
inhibit MDSCs include promoting differentiation, modulating
production flexibility, initiating recruitment in peripheral
organs, and direct elimination, aiming to circumvent the strong
toxicity and side effects associated with traditional non-specific
and sometimes ineffective chemotherapy.
Several therapies targeting MDSCs, either as
standalone immunotherapies or combined with standard methods, such
as chemotherapy and radiation, are undergoing preclinical trials to
enhance their antitumor capabilities. It is hoped that a more
in-depth understanding of the clinical significance of MDSCs will
prompt the development of MDSC-focused treatments, ultimately
improving the outcomes of patients with BC. The TME and MDSCs play
pivotal roles in tumor growth and survival, with their influence
particularly pronounced in TNBC. Emerging findings suggest that
targeting MDSCs may be a promising alternative therapeutic
approach, particularly in immunotherapy, reshaping the
immunosuppressive microenvironment and enhancing the efficacy of
cancer immunotherapy. In this context, nanotechnology is a valuable
tool. With its controlled drug release capabilities and increased
tumor accumulation, nanoparticles exhibit potential for cancer
treatment. Numerous studies on BC mouse models have shown promising
results with nanoparticle usage, including significant tumor
shrinkage and changes in the TME components. However, the
complexity of the microenvironment poses challenges in manipulating
it because changes to one cellular component can lead to cascading
effects on others. Despite the great promise of incorporating
nanotechnology in oncology, its application in clinical settings
still lacks successful results in clinical trials. Therefore,
comprehensive preclinical evaluations of various nanocarriers and a
thorough understanding of the strategies with which to most
effectively target MDSCs to alter the TME effectively are crucial
before they can be translated into clinical practice.
Acknowledgements
The authors gratefully acknowledge the Department of
Genetics and Morphology at the Institute of Biological Sciences,
University of Brasilia (GEM-IB-UnB) where the research related to
the present study work was conducted, the Coordinating Agency for
Advanced Training of Graduate Personnel (CAPES), and the National
Council for Technological and Scientific Development of Brazil
(CNPq) and the Federal District Research Support Foundation (FAPDF)
responsible for the financial support of research and
publications.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
DK conceptualized the study and was involved in the
collection of data from the literature for inclusion in the review,
in the writing and preparation of the original draft of the
manuscript, as well as in the editing of the manuscript, and in
figure creation. VCDS was involved in figure editing and in the
final reviewing of the manuscript. NLC was involved in the
conceptualization of the study, in the collection of data from the
literature for inclusion in the review, in the writing and
preparation of the original draft of the manuscript, and in the
editing and final reviewing of the manuscript. All authors have
reviewed and approved the submitted and final versions of the
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lei S, Zheng R, Zhang S, Wang S, Chen R,
Sun K, Zeng H, Zhou J and Wei W: Global patterns of breast cancer
incidence and mortality: A population-based cancer registry data
analysis from 2000 to 2020. Cancer Commun (Lond). 41:1183–1194.
2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Gradishar WJ, Moran MS, Abraham J, Aft R,
Agnese D, Allison KH, Anderson B, Burstein HJ, Chew H, Dang C, et
al: Breast cancer, version 3.2022, NCCN clinical practice
guidelines in oncology. J Natl Compr Canc Netw. 20:691–722.
2022.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Partridge AH, Burstein HJ and Winer EP:
Side effects of chemotherapy and combined chemohormonal therapy in
women with early-stage breast cancer. J Natl Cancer Inst Monogr.
(30):135–142. 2001.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Langeh U, Kumar V, Ahuja P, Singh C and
Singh A: An update on breast cancer chemotherapy-associated
toxicity and their management approaches. Health Sci Re.
9(100119)2023.
|
|
6
|
Slamon D, Eiermann W, Robert N, Pienkowski
T, Martin M, Press M, Mackey J, Glaspy J, Chan A, Pawlicki M, et
al: Adjuvant trastuzumab in HER2-positive breast cancer. N Engl J
Med. 365:1273–1283. 2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Darby SC, Ewertz M, McGale P, Bennet AM,
Blom-Goldman U, Brønnum D, Correa C, Cutter D, Gagliardi G, Gigante
B, et al: Risk of ischemic heart disease in women after
radiotherapy for breast cancer. N Engl J Med. 368:987–998.
2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Adams S, Gatti-Mays ME, Kalinsky K, Korde
LA, Sharon E, Amiri-Kordestani L, Bear H, McArthur HL, Frank E,
Perlmutter J, et al: Current landscape of immunotherapy in breast
cancer: A review. JAMA Oncol. 5:1205–1214. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Verheijden RJ, van Eijs MJM, May AM, van
Wijk F and Suijkerbuijk KPM: Immunosuppression for immune-related
adverse events during checkpoint inhibition: An intricate balance.
NPJ Precis Oncol. 7(41)2023.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Martins F, Sofiya L, Sykiotis GP, Lamine
F, Maillard M, Fraga M, Shabafrouz K, Ribi C, Cairoli A,
Guex-Crosier Y, et al: Adverse effects of immune-checkpoint
inhibitors: epidemiology, management and surveillance. Nat Rev Clin
Oncol. 6:563–580. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Trimboli RM, Giorgi Rossi P, Battisti NML,
Cozzi A, Magni V, Zanardo M and Sardanelli F: Do we still need
breast cancer screening in the era of targeted therapies and
precision medicine? Insights Imaging. 11(105)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Elemam NM, Talaat IM, Assal RA and Youness
RA: Understanding the crosstalk between immune cells and the tumor
microenvironment in cancer and its implications for immunotherapy.
Front Med (Lausanne). 10(1202581)2023.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Cha YJ and Koo JS: Role of
tumor-associated myeloid cells in breast cancer. Cells.
9(1785)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ortiz ML, Lu L, Ramachandran I and
Gabrilovich DI: Myeloid-derived suppressor cells in the development
of lung cancer. Cancer Immunol Res. 2:50–58. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Srivastava MK, Zhu L, Harris-White M, Kar
UK, Huang M, Johnson MF, Lee JM, Elashoff D, Strieter R, Dubinett S
and Sharma S: Myeloid suppressor cell depletion augments antitumor
activity in lung cancer. PLoS One. 7(e40677)2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yang Z, Guo J, Weng L, Tang W, Jin S and
Ma W: Myeloid-derived suppressor cells-new and exciting players in
lung cancer. J Hematol Oncol. 13(10)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Blaye C, Boyer T, Peyraud F, Domblides C
and Larmonier N: Beyond immunosuppression: The multifaceted
functions of tumor-promoting myeloid cells in breast cancers. Front
Immunol. 13(838040)2022.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li L, Li M and Jia Q: Myeloid-derived
suppressor cells: Key immunosuppressive regulators and therapeutic
targets in cancer. Pathol Res Pract. 248(154711)2023.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Parker KH, Beury DW and Ostrand-Rosenberg
S: Myeloid-derived suppressor cells: critical cells driving immune
suppression in the tumor microenvironment. Adv Cancer Res.
128:95–139. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Bronte V, Brandau S, Chen SH, Colombo MP,
Frey AB, Greten TF, Mandruzzato S, Murray PJ, Ochoa A,
Ostrand-Rosenberg S, et al: Recommendations for myeloid-derived
suppressor cell nomenclature and characterization standards. Nat
Commun. 7(12150)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Cassetta L, Baekkevold ES, Brandau S,
Bujko A, Cassatella MA, Dorhoi A, Krieg C, Lin A, Loré K, Marini O,
et al: Deciphering myeloid-derived suppressor cells: isolation and
markers in humans, mice, and non-human primates. Cancer Immunol
Immunother. 68:687–697. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang PF, Song SY, Wang TJ, Ji WJ, Li SW,
Liu N and Yan CX: Prognostic role of pretreatment circulating MDSCs
in patients with solid malignancies: A meta-analysis of 40 studies.
Oncoimmunology. 7(e1494113)2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Condamine T, Mastio J and Gabrilovich DI:
Transcriptional regulation of myeloid-derived suppressor cells. J
Leukoc Biol. 98:913–922. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Alshetaiwi H, Pervolarakis N, McIntyre LL,
Ma D, Nguyen Q, Rath JA, Nee K, Hernandez G, Evans K, Torosian L,
et al: Defining the emergence of myeloid-derived suppressor cells
in breast cancer using single-cell transcriptomics. Sci Immunol.
5(eaay6017)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Millrud CR, Bergenfelz C and Leandersson
K: On the origin of myeloid-derived suppressor cells. Oncotarget.
8:3649–3665. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Sun HW, Wu WC, Chen HT, Xu YT, Yang YY,
Chen J, Yu XJ, Wang Z, Shuang ZY and Zheng L: Glutamine deprivation
promotes the generation and mobilization of MDSCs by enhancing
expression of G-CSF and GM-CSF. Front Immunol.
11(616367)2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Rong Y, Yuan CH, Qu Z, Zhou H, Guan Q,
Yang N, Leng XH, Bu L, Wu K and Wang F: Doxorubicin-resistant
cancer cells activate myeloid-derived suppressor cells by releasing
PGE2. Sci Rep. 6(23824)2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ching MM, Reader J and Fulton AM:
Eicosanoids in cancer: prostaglandin E2 receptor 4 in cancer
therapeutics and immunotherapy. Front Pharmacol.
11(530199)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pradhan AK, Maji S, Bhoopathi P, Talukdar
S, Mannangatti P, Guo C, Wang XY, Cartagena LC, Idowu M, Landry JW,
et al: Pharmacological inhibition of MDA-9/Syntenin blocks breast
cancer metastasis through suppression of IL-1β. Proc Natl Acad Sci
USA. 118(e2103180118)2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Jiang M, Chen J, Zhang W, Zhang R, Ye Y,
Liu P, Yu W, Wei F, Ren X and Yu J: Interleukin-6 trans-signaling
pathway promotes immunosuppressive myeloid-derived suppressor cells
via suppression of suppressor of cytokine signaling 3 in breast
cancer. Front Immunol. 8(1840)2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhao N, Zhu W, Wang J, Liu W, Kang L, Yu R
and Liu B: Group 2 innate lymphoid cells promote TNBC lung
metastasis via the IL-13-MDSC axis in a murine tumor model. Int
Immunopharmacol. 99(107924)2021.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Popović M, Dedić Plavetić N, Vrbanec D,
Marušić Z, Mijatović D and Kulić A: Interleukin 17 in early
invasive breast cancer. Front Oncol. 13(1171254)2023.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Gao W, Wen H, Liang L, Dong X, Du R, Zhou
W, Zhang X, Zhang C, Xiang R and Li N: IL20RA signaling enhances
stemness and promotes the formation of an immunosuppressive
microenvironment in breast cancer. Theranostics. 11:2564–2580.
2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Xiao P, Wan X, Cui B, Liu Y, Qiu C, Rong
J, Zheng M, Song Y, Chen L, He J, et al: Interleukin 33 in tumor
microenvironment is crucial for the accumulation and function of
myeloid-derived suppressor cells. Oncoimmunology.
5(e1063772)2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kajihara N, Kobayashi T, Otsuka R,
Nio-Kobayashi J, Oshino T, Takahashi M, Imanishi S, Hashimoto A,
Wada H and Seino KI: Tumor-derived interleukin-34 creates an
immunosuppressive and chemoresistant tumor microenvironment by
modulating myeloid-derived suppressor cells in triple-negative
breast cancer. Cancer Immunol Immunother. 72:851–864.
2023.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Simpson KD, Templeton DJ and Cross JV:
Macrophage migration inhibitory factor promotes tumor growth and
metastasis by inducing myeloid-derived suppressor cells in the
tumor microenvironment. J Immunol. 189:5533–5540. 2012.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Liu QW, Chen Y, Li JY, Xiao L, Zhang WJ,
Zhao JL, Gu HC, Wu HY, Zuo GS, Deng KY and Xin HB: Bone marrow
cells are differentiated into MDSCs by BCC-Ex through
down-regulating the expression of CXCR4 and activating the STAT3
signalling pathway. J Cell Mol Med. 25:5497–5510. 2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Mundy-Bosse BL, Thornton LM, Yang HC,
Andersen BL and Carson WE: Psychological stress is associated with
altered levels of myeloid-derived suppressor cells in breast cancer
patients. Cell Immunol. 270:80–87. 2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
He K, Liu X, Hoffman RD, Shi RZ, Lv GY and
Gao JL: G-CSF/GM-CSF-induced hematopoietic dysregulation in the
progression of solid tumors. FEBS Open Bio. 12:1268–1285.
2022.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Smith KG and Clatworthy MR: FcγRIIB in
autoimmunity and infection: Evolutionary and therapeutic
implications. Nat Rev Immunol. 10:328–343. 2010.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wu L, Xu Y, Zhao H, Zhou Y, Chen Y, Yang
S, Lei J, Zhang J, Wang J, Wu Y and Li Y: FcγRIIB potentiates
differentiation of myeloid-derived suppressor cells to mediate
tumor immunoescape. Theranostics. 12:842–858. 2022.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hillmer EJ, Zhang H, Li HS and Watowich
SS: STAT3 signaling in immunity. Cytokine Growth Factor Rev.
31:1–15. 2016.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Liao WC, Yen HR, Chen CH, Chu YH, Song YC,
Tseng TJ and Liu CH: CHPF promotes malignancy of breast cancer
cells by modifying syndecan-4 and the tumor microenvironment. Am J
Cancer Res. 11:812–826. 2021.PubMed/NCBI
|
|
44
|
Liu C, Qiang J, Deng Q, Xia J, Deng L,
Zhou L, Wang D, He X, Liu Y, Zhao B, et al: ALDH1A1 activity in
tumor-initiating cells remodels myeloid-derived suppressor cells to
promote breast cancer progression. Cancer Res. 81:5919–5934.
2021.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Jiang M, Zhang W, Zhang R, Liu P, Ye Y, Yu
W, Guo X and Yu J: Cancer exosome-derived miR-9 and miR-181a
promote the development of early-stage MDSCs via interfering with
SOCS3 and PIAS3 respectively in breast cancer. Oncogene.
39:4681–4694. 2020.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Zhang W, Jiang M, Chen J, Zhang R, Ye Y,
Liu P, Yu W and Yu J: SOCS3 suppression promoted the recruitment of
CD11b+ Gr-1-F4/80-MHCII-early-stage myeloid-derived suppressor
cells and accelerated interleukin-6-related tumor invasion via
affecting myeloid differentiation in breast cancer. Front Immunol.
9(1699)2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Welte T, Kim IS, Tian L, Gao X, Wang H, Li
J, Holdman XB, Herschkowitz JI, Pond A, Xie G, et al: Oncogenic
mTOR signalling recruits myeloid-derived suppressor cells to
promote tumour initiation. Nat Cell Biol. 18:632–644.
2016.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Ozga AJ, Chow MT and Luster AD: Chemokines
and the immune response to cancer. Immunity. 54:859–874.
2021.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Huang YC, Hou MF, Tsai YM, Pan YC, Tsai
PH, Lin YS, Chang CY, Tsai EM and Hsu YL: Involvement of ACACA
(acetyl-CoA carboxylase α) in the lung pre-metastatic niche
formation in breast cancer by senescence phenotypic conversion in
fibroblasts. Cell Oncol (Dordr). 46:643–660. 2023.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Gu P, Sun M, Li L, Yang Y, Jiang Z, Ge Y,
Wang W, Mu W and Wang H: Breast tumor-derived exosomal
microRNA-200b-3p promotes specific organ metastasis through
regulating CCL2 expression in lung epithelial cells. Front Cell Dev
Biol. 9(657158)2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Tanaka T, Kajiwara T, Torigoe T, Okamoto
Y, Sato N and Tamura Y: Cancer-associated oxidoreductase ERO1-α
drives the production of tumor-promoting myeloid-derived suppressor
cells via oxidative protein folding. J Immunol. 194:2004–2010.
2015.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Liu Y, Lai L, Chen Q, Song Y, Xu S, Ma F,
Wang X, Wang J, Yu H, Cao X and Wang Q: MicroRNA-494 is required
for the accumulation and functions of tumor-expanded
myeloid-derived suppressor cells via targeting of PTEN. J Immunol.
188:5500–5510. 2012.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Guo L, Kong D, Liu J, Zhan L, Luo L, Zheng
W, Zheng Q, Chen C and Sun S: Breast cancer heterogeneity and its
implication in personalized precision therapy. Exp Hematol Oncol.
12(3)2023.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Vrakas CN, O'Sullivan RM, Evans SE, Ingram
DA, Jones CB, Phuong T and Kurt RA: The Measure of DAMPs and a role
for S100A8 in recruiting suppressor cells in breast cancer lung
metastasis. Immunol Invest. 44:174–188. 2015.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Chen JY, Lai YS, Chu PY, Chan SH, Wang LH
and Hung WC: Cancer-derived VEGF-C increases chemokine production
in lymphatic endothelial cells to promote CXCR2-dependent cancer
invasion and MDSC recruitment. Cancers (Basel).
11(1120)2019.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Roberts LM, Perez MJ, Balogh KN,
Mingledorff G, Cross JV and Munson JM: Myeloid derived suppressor
cells migrate in response to flow and lymphatic endothelial cell
interaction in the breast tumor microenvironment. Cancers (Basel).
14(3008)2022.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Yu B, Luo F, Sun B, Liu W, Shi Q, Cheng
SY, Chen C, Chen G, Li Y and Feng H: KAT6A acetylation of SMAD3
regulates myeloid-derived suppressor cell recruitment, metastasis,
and immunotherapy in triple-negative breast cancer. Adv Sci
(Weinh). 8(e2100014)2021.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Vadrevu SK, Chintala NK, Sharma SK, Sharma
P, Cleveland C, Riediger L, Manne S, Fairlie DP, Gorczyca W,
Almanza O, et al: Complement c5a receptor facilitates cancer
metastasis by altering T-cell responses in the metastatic niche.
Cancer Res. 74:3454–3465. 2014.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Cheng R, Billet S, Liu C, Haldar S,
Choudhury D, Tripathi M, Hav M, Merchant A, Hu T, Huang H, et al:
Periodontal inflammation recruits distant metastatic breast cancer
cells by increasing myeloid-derived suppressor cells. Oncogene.
39:1543–1556. 2020.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Tcyganov E, Mastio J, Chen E and
Gabrilovich DI: Plasticity of myeloid-derived suppressor cells in
cancer. Curr Opin Immunol. 51:76–82. 2018.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Mehta AK, Kadel S, Townsend MG, Oliwa M
and Guerriero JL: Macrophage biology and mechanisms of immune
suppression in breast cancer. Front Immunol.
12(643771)2021.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Ostrand-Rosenberg S and Fenselau C:
Myeloid-derived suppressor cells: immune-suppressive cells that
impair antitumor immunity and are sculpted by their environment. J
Immunol. 200:422–431. 2018.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Cayrol C and Girard JP: Interleukin-33
(IL-33): A critical review of its biology and the mechanisms
involved in its release as a potent extracellular cytokine.
Cytokine. 156(155891)2022.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Mattiola I and Diefenbach A: Enabling
anti-tumor immunity by unleashing ILC2. Cell Res. 30:461–462.
2020.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Halvorsen EC, Franks SE, Wadsworth BJ,
Harbourne BT, Cederberg RA, Steer CA, Martinez-Gonzalez I, Calder
J, Lockwood WW and Bennewith KL: IL-33 increases ST2+ Tregs and
promotes metastatic tumour growth in the lungs in an
amphiregulin-dependent manner. Oncoimmunology.
8(e1527497)2018.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Gurram RK and Zhu J: Orchestration between
ILC2s and Th2 cells in shaping type 2 immune responses. Cell Mol
Immunol. 16:225–235. 2019.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Choi MR, Sosman JA and Zhang B: The janus
face of IL-33 signaling in tumor development and immune escape.
Cancers (Basel). 13(3281)2021.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Huang X, Cao J and Zu X: Tumor-associated
macrophages: An important player in breast cancer progression.
Thorac Cancer. 13:269–276. 2022.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Hao NB, Lü MH, Fan YH, Cao YL, Zhang ZR
and Yang SM: Macrophages in tumor microenvironments and the
progression of tumors. Clin Dev Immunol.
2012(948098)2012.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Boutilier AJ and Elsawa SF: Macrophage
polarization states in the tumor microenvironment. Int J Mol Sci.
22(6995)2021.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Wang S, Wang J, Chen Z, Luo J, Guo W, Sun
L and Lin L: Targeting M2-like tumor-associated macrophages is a
potential therapeutic approach to overcome antitumor drug
resistance. NPJ Precis Oncol. 8(31)2024.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Chen S, Saeed AFUH, Liu Q, Jiang Q, Xu H,
Xiao GG, Rao L and Duo Y: Macrophages in immunoregulation and
therapeutics. Signal Transduct Target Ther. 8(207)2023.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Biswas S, Mandal G, Roy Chowdhury S,
Purohit S, Payne KK, Anadon C, Gupta A, Swanson P, Yu X,
Conejo-Garcia JR and Bhattacharyya A: Exosomes produced by
mesenchymal stem cells drive differentiation of myeloid cells into
immunosuppressive M2-polarized macrophages in breast cancer. J
Immunol. 203:3447–3460. 2019.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Payne KK, Zoon CK, Wan W, Marlar K, Keim
RC, Kenari MN, Kazim AL, Bear HD and Manjili MH: Peripheral blood
mononuclear cells of patients with breast cancer can be
reprogrammed to enhance anti-HER-2/neu reactivity and overcome
myeloid-derived suppressor cells. Breast Cancer Res Treat.
142:45–57. 2013.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Gabrilovich DI, Ostrand-Rosenberg S and
Bronte V: Coordinated regulation of myeloid cells by tumours. Nat
Rev Immunol. 12:253–268. 2012.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Pansy K, Uhl B, Krstic J, Szmyra M,
Fechter K, Santiso A, Thüminger L, Greinix H, Kargl J, Prochazka K,
et al: Immune regulatory processes of the tumor microenvironment
under malignant conditions. Int J Mol Sci. 22(13311)2021.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Li F, Zhao Y, Wei L, Li S and Liu J:
Tumor-infiltrating Treg, MDSC, and IDO expression associated with
outcomes of neoadjuvant chemotherapy of breast cancer. Cancer Biol
Ther. 19:695–705. 2018.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Srivastava MK, Sinha P, Clements VK,
Rodriguez P and Ostrand-Rosenberg S: Myeloid-derived suppressor
cells inhibit T-cell activation by depleting cystine and cysteine.
Cancer Res. 70:68–77. 2010.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Lu T, Ramakrishnan R, Altiok S, Youn JI,
Cheng P, Celis E, Pisarev V, Sherman S, Sporn MB and Gabrilovich D:
Tumor-infiltrating myeloid cells induce tumor cell resistance to
cytotoxic T cells in mice. J Clin Invest. 121:4015–4029.
2011.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Stiff A, Trikha P, Mundy-Bosse B,
McMichael E, Mace TA, Benner B, Kendra K, Campbell A, Gautam S and
Abood D: , et al: Nitric oxide production by myeloid-derived
suppressor cells plays a role in impairing Fc receptor-mediated
natural killer cell function. Clin Cancer Res. 24:1891–1904.
2018.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Sceneay J, Griessinger CM, Hoffmann SHL,
Wen SW, Wong CSF, Krumeich S, Kneilling M, Pichler BJ and Möller A:
Tracking the fate of adoptively transferred myeloid-derived
suppressor cells in the primary breast tumor microenvironment. PLoS
One. 13(e0196040)2018.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Hanson EM, Clements VK, Sinha P, Ilkovitch
D and Ostrand-Rosenberg S: Myeloid-derived suppressor cells
down-regulate L-selectin expression on CD4+ and CD8+ T cells. J
Immunol. 183:937–944. 2009.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Sinha P, Chornoguz O, Clements VK,
Artemenko KA, Zubarev RA and Ostrand-Rosenberg S: Myeloid-derived
suppressor cells express the death receptor Fas and apoptose in
response to T cell-expressed FasL. Blood. 117:5381–5390.
2011.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Lelis FJ, Jaufmann J, Singh A, Fromm K,
Teschner AC, Pöschel S, Schäfer I, Beer-Hammer S, Rieber N and
Hartl D: Myeloid-derived suppressor cells modulate B-cell
responses. Immunol Lett. 188:108–115. 2017.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Shen M, Wang J, Yu W, Zhang C, Liu M, Wang
K, Yang L, Wei F, Wang SE, Sun Q and Ren X: A novel MDSC-induced
PD-1- PD-L1+ B-cell subset in breast tumor microenvironment
possesses immuno-suppressive properties. Oncoimmunology.
7(e1413520)2018.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Nam S, Lee A, Lim J and Lim JS: Analysis
of the expression and regulation of PD-1 protein on the surface of
myeloid-derived suppressor cells (MDSCs). Biomol Ther (Seoul).
27:63–70. 2019.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Liu M, Wei F, Wang J, Yu W, Shen M, Liu T,
Zhang D, Wang Y, Ren X and Sun Q: Myeloid-derived suppressor cells
regulate the immunosuppressive functions of PD-1- PD-L1+ Bregs
through PD-L1/PI3K/AKT/NF-κB axis in breast cancer. Cell Death Dis.
12(465)2021.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Spallanzani RG, Dalotto-Moreno T, Raffo
Iraolagoitia XL, Ziblat A, Domaica CI, Avila DE, Rossi LE, Fuertes
MB, Battistone MA, Rabinovich GA, et al: Expansion of CD11b+ Ly6G+
Ly6C int cells driven by medroxyprogesterone acetate in mice
bearing breast tumors restrains NK cell effector functions. Cancer
Immunol Immunother. 62:1781–1795. 2013.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Sceneay J, Chow MT, Chen A, Halse HM, Wong
CS, Andrews DM, Sloan EK, Parker BS, Bowtell DD, Smyth MJ and
Möller A: Primary tumor hypoxia recruits CD11b+/Ly6Cmed/Ly6G+
immune suppressor cells and compromises NK cell cytotoxicity in the
premetastatic niche. Cancer Res. 72:3906–3911. 2012.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Deng Z, Rong Y, Teng Y, Zhuang X,
Samykutty A, Mu J, Zhang L, Cao P, Yan J, Miller D and Zhang HG:
Exosomes miR-126a released from MDSC induced by DOX treatment
promotes lung metastasis. Oncogene. 36:639–651. 2017.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Ma X, Wang M, Yin T, Zhao Y and Wei X:
Myeloid-derived suppressor cells promote metastasis in breast
cancer after the stress of operative removal of the primary cancer.
Front Oncol. 9(855)2019.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Bergenfelz C, Roxå A, Mehmeti M,
Leandersson K and Larsson AM: Clinical relevance of systemic
monocytic-MDSCs in patients with metastatic breast cancer. Cancer
Immunol Immunother. 69:435–448. 2020.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Liu H, Wang Z, Zhou Y and Yang Y: MDSCs in
breast cancer: An important enabler of tumor progression and an
emerging therapeutic target. Front Immunol.
14(1199273)2023.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Veglia F, Perego M and Gabrilovich D:
Myeloid-derived suppressor cells coming of age. Nat Immunol.
19:108–119. 2018.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Gatti-Mays ME, Balko JM, Gameiro SR, Bear
HD, Prabhakaran S, Fukui J, Disis ML, Nanda R, Gulley JL, Kalinsky
K, et al: If we build it they will come: targeting the immune
response to breast cancer. NPJ Breast Cancer. 5(37)2019.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Kim K, Skora AD, Li Z, Liu Q, Tam AJ,
Blosser RL, Diaz LA Jr, Papadopoulos N, Kinzler KW, Vogelstein B
and Zhou S: Eradication of metastatic mouse cancers resistant to
immune checkpoint blockade by suppression of myeloid-derived cells.
Proc Natl Acad Sci USA. 111:11774–11779. 2014.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Le HK, Graham L, Cha E, Morales JK,
Manjili MH and Bear HD: Gemcitabine directly inhibits myeloid
derived suppressor cells in BALB/c mice bearing 4T1 mammary
carcinoma and augments expansion of T cells from tumor-bearing
mice. Int Immunopharmacol. 9:900–909. 2009.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Alizadeh D, Trad M, Hanke NT, Larmonier
CB, Janikashvili N, Bonnotte B, Katsanis E and Larmonier N:
Doxorubicin eliminates myeloid-derived suppressor cells and
enhances the efficacy of adoptive T-cell transfer in breast cancer.
Cancer Res. 74:104–118. 2014.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Vincent J, Mignot G, Chalmin F, Ladoire S,
Bruchard M, Chevriaux A, Martin F, Apetoh L, Rébé C and
Ghiringhelli F: 5-Fluorouracil selectively kills tumor-associated
myeloid-derived suppressor cells resulting in enhanced T
cell-dependent antitumor immunity. Cancer Res. 70:3052–3061.
2010.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Sharma P, Abramson V, O’Dea A, Nye L,
Mayer I, Crane G, Elia M, Yoder R, Staley J, Schwensen K, et al:
Romidepsin (HDACi) plus cisplatin and nivolumab triplet combination
in patients with metastatic triple negative breast cancer (mTNBC).
J Clin Oncol. 39(10.1200/JCO.2021.39.15_suppl.1076)2021.
|
|
101
|
Davis RJ, Moore EC, Clavijo PE, Friedman
J, Cash H, Chen Z, Silvin C, Van Waes C and Allen C: Anti-PD-L1
efficacy can be enhanced by inhibition of myeloid-derived
suppressor cells with a selective inhibitor of PI3Kδ/γ. Cancer Res.
77:2607–2619. 2017.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Tu SP, Jin H, Shi JD, Zhu LM, Suo Y, Lu G,
Liu A, Wang TC and Yang CS: Curcumin induces the differentiation of
myeloid-derived suppressor cells and inhibits their interaction
with cancer cells and related tumor growth. Cancer Prev Res
(Phila). 5:205–215. 2012.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Sánchez-León ML, Jiménez-Cortegana C,
Silva Romeiro S, Garnacho C, de la Cruz-Merino L, García-Domínguez
DJ, Hontecillas-Prieto L and Sánchez-Margalet V: Defining the
emergence of new immunotherapy approaches in breast cancer: Role of
myeloid-derived suppressor cells. Int J Mol Sci.
24(5208)2023.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Kusmartsev S, Cheng F, Yu B, Nefedova Y,
Sotomayor E, Lush R and Gabrilovich D: All-trans-retinoic acid
eliminates immature myeloid cells from tumor-bearing mice and
improves the effect of vaccination. Cancer Res. 63:4441–4449.
2003.PubMed/NCBI
|
|
105
|
Iclozan C, Antonia S, Chiappori A, Chen DT
and Gabrilovich D: Therapeutic regulation of myeloid-derived
suppressor cells and immune response to cancer vaccine in patients
with extensive stage small cell lung cancer. Cancer Immunol
Immunother. 62:909–918. 2013.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Forghani P, Khorramizadeh MR and Waller
EK: Silibinin inhibits accumulation of myeloid-derived suppressor
cells and tumor growth of murine breast cancer. Cancer Med.
3:215–224. 2014.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Sawant A, Deshane J, Jules J, Lee CM,
Harris BA, Feng X and Ponnazhagan S: Myeloid-derived suppressor
cells function as novel osteoclast progenitors enhancing bone loss
in breast cancer. Cancer Res. 73:672–682. 2013.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Kugler A, Stuhler G, Walden P, Zöller G,
Zobywalski A, Brossart P, Trefzer U, Ullrich S, Müller CA, Becker
V, et al: Regression of human metastatic renal cell carcinoma after
vaccination with tumor cell-dendritic cell hybrids. Nat Med.
6:332–336. 2000.PubMed/NCBI View
Article : Google Scholar
|
|
109
|
Thakur A, Schalk D, Sarkar SH, Al-Khadimi
Z, Sarkar FH and Lum LG: A Th1 cytokine-enriched microenvironment
enhances tumor killing by activated T cells armed with bispecific
antibodies and inhibits the development of myeloid-derived
suppressor cells. Cancer Immunol Immunother. 61:497–509.
2012.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Kmieciak M, Basu D, Payne KK, Toor A,
Yacoub A, Wang XY, Smith L, Bear HD and Manjili MH: Activated NK T
cells and NK cells render T cells resistant to MDSC and result in
an effective adoptive cellular therapy against breast cancer in the
FVBN202 transgenic mouse. J Immunol. 187:708–717. 2011.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Chandra D, Jahangir A, Quispe-Tintaya W,
Einstein MH and Gravekamp C: Myeloid-derived suppressor cells have
a central role in attenuated Listeria monocytogenes-based
immunotherapy against metastatic breast cancer in young and old
mice. Br J Cancer. 108:2281–2290. 2013.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Chaves NL, Amorim DA, Lopes CAP,
Estrela-Lopis I, Böttner J, de Souza AR and Báo SN: Comparison of
the effect of rhodium citrate-associated iron oxide nanoparticles
on metastatic and non-metastatic breast cancer cells. Cancer Nano.
10:1–12. 2019.
|
|
113
|
Yao Y, Zhou Y, Liu L, Xu Y, Chen Q, Wang
Y, Wu S, Deng Y, Zhang J and Shao A: Nanoparticle-based drug
delivery in cancer therapy and its role in overcoming drug
resistance. Front Mol Biosci. 7(193)2020.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Chaves NL, Estrela-Lopis I, Böttner J,
Lopes CA, Guido BC, de Sousa AR and Báo SN: Exploring cellular
uptake of iron oxide nanoparticles associated with rhodium citrate
in breast cancer cells. Int J Nanomedicine. 12:5511–5523.
2017.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Figueiro Longo JP and Muehlmann LA:
Nanomedicine beyond tumor passive targeting: What next?
Nanomedicine (Lond). 15:1819–1822. 2020.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Zhang N, Liu S, Shi S, Chen Y, Xu F, Wei X
and Xu Y: Solubilization and delivery of Ursolic-acid for
modulating tumor microenvironment and regulatory T cell activities
in cancer immunotherapy. J Control Release. 320:168–178.
2020.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Chen C, Li A, Sun P, Xu J, Du W, Zhang J,
Liu Y, Zhang R, Zhang S, Yang Z, et al: Efficiently restoring the
tumoricidal immunity against resistant malignancies via an immune
nanomodulator. J Control Release. 324:574–585. 2020.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Ali R, Shao H and Varamini P: Potential
Nanotechnology-Based Therapeutics to Prevent Cancer Progression
through TME Cell-Driven Populations. Pharmaceutics.
15(112)2022.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Lu Z, Liu H, Ma L, Ren K, He Z, Li M and
He Q: Micellar nanoparticles inhibit breast cancer and pulmonary
metastasis by modulating the recruitment and depletion of
myeloid-derived suppressor cells. Nanoscale. 14:17315–17330.
2022.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Debien V, De Caluwé A, Wang X,
Piccart-Gebhart M, Tuohy VK, Romano E and Buisseret L:
Immunotherapy in breast cancer: An overview of current strategies
and perspectives. NPJ Breast Cancer. 9(7)2023.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Teschendorff AE, Miremadi A, Pinder SE,
Ellis IO and Caldas C: An immune response gene expression module
identifies a good prognosis subtype in estrogen receptor negative
breast cancer. Genome Biol. 8(R157)2007.PubMed/NCBI View Article : Google Scholar
|