Introduction
Cervical cancer is a major common cancer in women in
developing countries, and it is the second most frequent cause of
cancer-related death in women worldwide (1). Following standard treatment, a high
survival rate is noted in patients with cervical cancer. However,
the issue of recurrence and subsequent resistance to chemoradiation
therapy, as well as the mechanisms involved have yet to be
elucidated.
Rhein, one of the major bioactive constituents of
the rhizome of rhubarb (2,3), inhibits the proliferation of various
human cancer cells (4–8). Our previous study showed that rhein
lysinate (RHL; the salt of rhein and lysine, easily dissolved in
water) exhibits anti-cancer activity in breast and ovarian cancer
and in hepatocellular carcinoma in vivo and in vitro
(9–11).
RHL affects various cell signaling pathways, and
interest in its mechanisms has expanded to include a number of
protein kinase pathways. The mitogen-activated protein kinase
(MAPK) superfamily comprises a number of signaling pathways
involved in growth regulation (12,13).
These pathways include extracellular signal-regulated kinase 1
(ERK1) and ERK2 (or p42 MAPK and p44 MAPK), c-Jun NH2-terminal
kinases (JNKs) and p38 MAPK.
The potential role of RHL in the treatment of
cervical cancer remains to be investigated and the mechanism by
which RHL affects tumor growth remains to be determined. Therefore,
the effects of RHL on cell growth, apoptosis and various MAPKs in
cervical carcinoma HeLa cells were investigated.
Materials and methods
Chemicals and reagents
Rhein (98%) was purchased from Nanjing Qingze
Medicine Ltd. (Nanjing, Jiangsu, China). Lysine was purchased from
Beijing Solarbio Science and Technology Co. (Beijing, China). RHL
was created in our department (Patent No. 200810089025.8).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and dimethyl sulfoxide (DMSO) were obtained from Sigma Aldrich
(Shanghai, China). The remaining chemicals were of standard
analytical grade.
Cell culture
Human cervical carcinoma HeLa cell line was obtained
from the Cell Center of the Institute of Basic Medical Sciences,
Chinese Academy of Medical Sciences and Peking Union Medical
College, China. The HeLa cells were cultured in Dulbecco’s modified
Eagle’s medium (Gibco BRL, Grand Island, NY, USA) supplemented with
10% heat-inactivated fetal bovine serum (Sigma Chemical Co., St.
Louis, MO, USA), 2 mM glutamine, 100 U/ml penicillin and 100 μg/ml
streptomycin at 37°C in a humidified atmosphere containing 5%
CO2.
Cell proliferation assay
The cell proliferation assay was examined with the
MTT method, according to the manufacturer’s instructions. Cells
were seeded in 96-well plates (Costar, Cambridge, MA, USA) with
2,500 cells/well. Following overnight incubation, triplicate wells
were treated with various concentrations of RHL for 48 h. Then, 20
μl MTT solutions (5 mg/ml in PBS) were added to each well and
incubated for 4 h at 37°C. The MTT formazan was dissolved in 150 μl
DMSO and absorbance was measured using a Microplate Reader
(Multiskan MK3; Thermo Labsystem, USA) at a wavelength of 570
nm.
FITC-Annexin V/propidium iodide (PI)
apoptosis assay
Cells were collected and resuspended in 200 μl
binding buffer. Then, 10 μl FITC-labeled enhanced Annexin V (Baosai
Biotechnology Ltd., Beijing, China) and 100 ng PI were added.
Following incubation in the dark (15 min at room temperature or 30
min at 4°C), the samples were diluted with 300 μl binding buffer.
Flow cytometry was carried out using a FACScan instrument
(Becton-Dickinson) and the data were processed using
WinMDI/PC-software.
Western blot analysis
Cells were harvested and washed with PBS solution.
The whole cellular extracts were prepared by incubating cells on
ice in lysis buffer containing 50 mM Tris-HCl pH 7.5, 150 mM NaCl,
2 mM EDTA, 2 mM EGTA, 1 mM dithiothreitol, 1% Nonidet P-40, 0.1%
SDS, protease inhibitors (1 mM PMSF, 5 μg/ml aprotinin, 5 μg/ml
leupeptin and 5 μg/ml pepstatin) and phosphatase inhibitors (20 mM
β-glycerophosphate, 50 mM NaF and 1 mM
Na3VO4). The cell lysates were cleared by
centrifugation at 12,000 × g for 12 min. Protein concentrations
were determined using the Bradford assay. Equal amounts of lysate
(40 μg) were resolved by SDS-PAGE and transferred to polyvinylidene
difluoride membranes (Millipore Corp., Bedford, MA, USA). The
membranes were blocked in TBST containing 5% non-fat skim milk at
room temperature for 2 h and probed with primary antibodies
overnight at 4°C. Membranes were then blotted with an appropriate
horseradish peroxidase-linked secondary antibody (Santa Cruz
Biotechnology, Santa Cruz, CA, USA). Proteins were visualized using
enhanced chemiluminescence Western blot detection reagents
(Amersham Pharmacia Biotech, Inc., Piscataway, NJ, USA).
Results
Inhibition of proliferation by rhein
lysinate in HeLa cells
The growth inhibitory effect of RHL on HeLa cells
was examined using MTT assay. Cells were cultured for 48 h, with or
without different concentrations of RHL. A decreased cell
proliferation following treatment with various concentrations of
RHL is shown (Fig. 1A). The
IC50 value of RHL for the HeLa cells was ~80 μM. The
cells were cultured with RHL (50, 75, 100 and 125 μM) for the
indicated times and analyzed with the MTT assay (Fig. 1B). HeLa cells treated with the
indicated dose of RHL showed decreased cell proliferation in a
time-dependent manner.
Apoptosis induction of rhein lysinate in
HeLa cells
FITC-Annexin V/PI staining showed that RHL at 50 μM
induced early apoptosis in HeLa cells (Fig. 2A). The ratio of early apoptosis was
significantly enhanced when cells were incubated with 75 or 100 μM
RHL for 48 h. To determine whether RHL-induced apoptosis is
mediated by the activation of caspase and/or PARP, HeLa cells were
treated with RHL at various concentrations for 48 h, RHL-induced
caspase-3/7 and PARP cleavage in a dose-dependent manner (Fig. 2B). Taken together, these results
strongly suggest that RHL triggers caspase-dependent apoptosis in
HeLa cells.
Growth inhibition of rhein lysinate in
HeLa cells was associated with the activation of various MAPKs
MAPKs are a family of proteins that transduce
signals from the cell membrane to the nucleus in response to a wide
range of stimuli and regulate vital biological functions, including
gene expression, mitosis, differentiation, apoptosis, proliferation
and motility (12,13). Three major groups of MAPKs exist:
JNK, p38 MAPK and ERK.
To investigate the molecular mechanisms underlying
growth inhibition, the effect of RHL on the activities of MAPKs in
HeLa cells was examined. As shown in Fig. 3, Western blot analysis revealed that
RHL treatment increased the phosphorylation of JNK, p38 MAPK and
ERK1/2 in a dose- and time-dependent manner in HeLa cells. These
results suggest that RHL inhibits the growth of cervical cancer
cells by modulating the activation of various MAPKs.
Effects of mitogen-activated protein
kinase on growth inhibition induced by rhein lysinate in HeLa
cells
To further examine the role of various MAPKs in the
inhibition of cervical cancer cell growth, pharmacological
inhibitors of MAPKs (the JNK inhibitor SP600125, the p38 MAPK
inhibitor SB203580 and the MEK inhibitor U0126) were used in
combination with RHL.
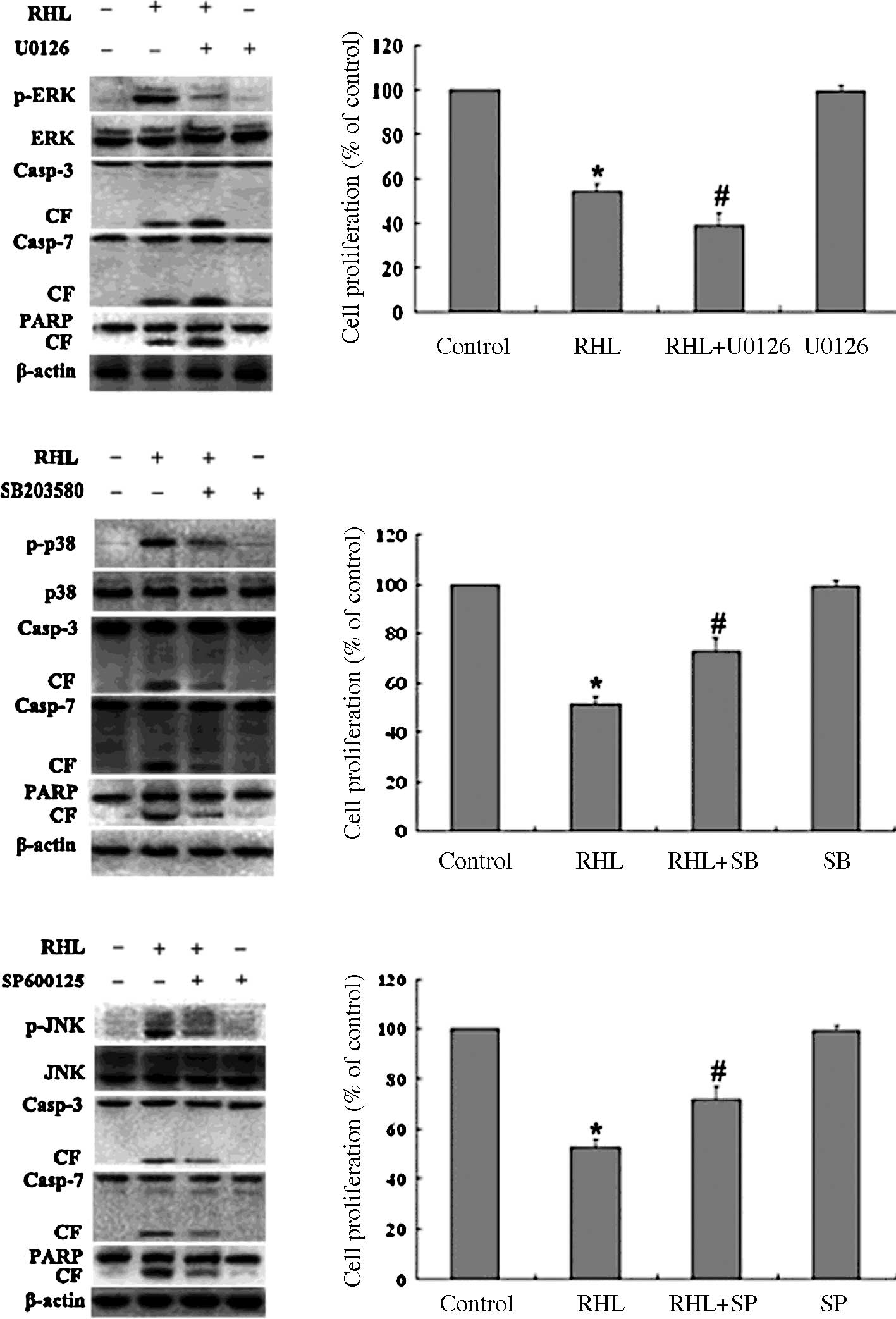
The specificity of each inhibitor for various
kinases on RHL-treated cells was assessed. Inhibitors were added
and incubated for 1 h, followed by RHL treatment (75 μM) for 48 h.
As shown in Fig. 4A, the
specificity of each inhibitor was confirmed. Moreover, the cleavage
of caspase-3/7 and PARP triggered by RHL was blocked by SP600125
and SB203580, whereas the cleavage of caspase-3/7 and PARP
triggered by RHL was enhanced by U0126. As shown in Fig. 4B, in the absence of RHL, the
inhibitors of JNK, p38 MAPK and MEK alone did not significantly
alter proliferation in the HeLa cells. When the HeLa cells were
treated with RHL in the presence of three inhibitors, respectively,
the situation was altered. MTT results showed that inhibition of
the p38 MAPK activity with SB203580 and inhibition of the JNK
activity with SP600125 rescued RHL-mediated cell growth inhibition
in HeLa cells, whereas the inhibition of ERK activity with U0126
enhanced RHL-induced growth inhibition. These results suggest that
JNK and p38 MAPK play a partial role in mediating HeLa cell
apoptosis triggered by RHL.
Discussion
Cervical cancer frequently occurs in women in
developing countries, and it is the second most frequent cause of
cancer-related death in women worldwide (1). The survival rate after standard
treatment for cervical cancer is high. However, recurrence and
subsequent resistance to chemoradiation therapy remain a major
health issue and the mechanisms involved have yet to be elucidated.
Novel agents are required to offer long-term disease control or a
potential cure.
Rhein is one of the major bioactive constituents of
the rhizome of rhubarb (2,3), and inhibits the proliferation of
various human cancer cells (4–8). Our
previous study showed that RHL exhibits anti-cancer activity in
breast and ovarian cancer, and hepatocellular carcinoma in
vivo and in vitro (9–11). In
the present study, RHL was shown to inhibit the growth of human
cervical cancer cells. The IC50 value of RHL for HeLa
cells was 80 μM. HeLa cells treated with RHL showed an increasing
apoptotic ratio in a dose-dependent manner. This increase is
related to certain molecular changes involving the activation of
MAPKs, cleavage of caspase-3/7 and PARP. It is well known that
MAPKs are a family of proteins that transduce signals from the cell
membrane to the nucleus in response to a wide range of stimuli.
Moreover, MAPKs regulate vital biological functions, including gene
expression, mitosis, differentiation, apoptosis, proliferation and
motility (12,13). There are three major groups of
MAPKs, i.e., JNK, p38 MAPK and ERK. The involvement of JNK in
response to stress stimuli, cytokines and anti-cancer agents was
previously documented (14–17). In our previous study, JNK played a
crucial role in growth inhibition induced by lidamycin in human
multiple myeloma cells (18,19).
In the present study, RHL triggered JNK phosphorylation in a
dose-dependent manner in HeLa cells. The results showed that the
JNK inhibitor SP600125 markedly attenuated RHL-induced growth
inhibition in HeLa cells. The results suggest that the activation
of JNK plays a crucial role in growth inhibition induced by RHL in
HeLa cells.
p38 MAPK has been shown to mediate both
proapoptotic/ growth inhibitory and antiapoptotic/progrowth signals
in various systems, depending on the stimulus and cell type
involved (13,20–22).
Since RHL triggered the phosphorylation of p38 MAPK, we examined
the role of p38 MAPK activation in HeLa cells to RHL treatment. The
results showed that the p38 MAPK inhibitor SB203580 markedly
rescued RHL-induced growth inhibition in HeLa cells.
The results suggest that the activation of p38 MAPK
also plays a partial role in mediating HeLa cell growth inhibition
triggered by RHL in HeLa cells.
The decrease of HeLa cell growth inhibition by the
combination of RHL with SP600125 or SB203580 was accompanied by the
inactivation of caspase-3/7 and PARP, suggesting that the sustained
activation of JNK and p38 MAPK plays a critical role in RHL-induced
apoptosis in HeLa cells.
The activation of ERK was crucial in mediating
proliferation in cancer cells (19,20,23–25).
In the present study, RHL triggered significant up-regulation of
ERK phosphorylation. We hypothesized that this up-regulation is a
compensatory positive feedback response to maintain HeLa cell
survival. Inhibiting MEK/ERK signaling would therefore enhance
RHL-induced growth inhibition in HeLa cells. To inhibit
up-regulation of MEK/ERK activity triggered by RHL, we utilized the
MEK1/2 inhibitor, U0126, which inhibits ERK activity. Growth
inhibition triggered by RHL was synergistically augmented in the
presence of U0126. The results suggest the potential of MEK/ERK
inhibition to enhance sensitivity to RHL and to overcome RHL
resistance, thereby improving the therapeutic efficacy of RHL in
HeLa cells. The increase of HeLa cell growth inhibition by the
combination of RHL with U0126 was accompanied by the enhancement of
the activation of caspase-3/7 and PARP, suggesting that ERK is
involved in regulating cell survival. The finding that RHL
inhibited ERK activation was shown in another study using human
mammary epithelial MCF cells (9).
However, in our study, the activation of ERK was increased
following RHL treatment in HeLa cells. These findings indicate that
ERK is activated or inhibited by RHL in a cell type-specific
manner.
RHL activates the three members of MAPKs, thus
modulating the balance between the survival and death signaling
cascades, thereby inducing growth inhibition. Taken together, these
results suggest that RHL inhibits cell growth via the activation of
p38 MAPK and JNK. The results provide the rationale for clinical
trials of RHL aimed at improving patient outcome in cervical
cancer.
Acknowledgements
We would like to thank Professor Yong-Shu Zhen
(Department of Oncology, Institute of Medicinal Biotechnology,
Chinese Academy of Medical Science, Beijing, China) for kindly
providing us with the rights to use Patent No. 200810089025.8.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
2
|
Kuo PL, Hsu YL, Ng LT and Lin CC: Rhein
inhibits the growth and induces the apoptosis of Hep G2 cells.
Planta Med. 70:12–16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang Q, Lu G, Shen HM, Chung MC and Ong
CN: Anti-cancer properties of anthraquinones from rhubarb. Med Res
Rev. 27:609–630. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lai WW, Yang JS, Lai KC, et al: Rhein
induced apoptosis through the endoplasmic reticulum stress,
caspase-and mitochondria-dependent pathways in SCC-4 human tongue
squamous cancer cells. In Vivo. 23:309–316. 2009.
|
|
5
|
Ip SW, Weng YS, Lin SY, et al: The role of
Ca2+ on rhein-induced apoptosis in human cervical cancer
Ca Ski cells. Anticancer Res. 27:379–389. 2007.
|
|
6
|
Lin ML, Chen SS, Lu YC, et al: Rhein
induces apoptosis through induction of endoplasmic reticulum stress
and Ca2+-dependent mitochondrial death pathway in human
nasopharyngeal carcinoma cells. Anticancer Res. 27:3313–3322.
2007.PubMed/NCBI
|
|
7
|
Cichewicz RH, Zhang Y, Seeram NP and Nair
MG: Inhibition of human tumor cell proliferation by novel
anthraquinones from daylilies. Life Sci. 74:1791–1799. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Floridi A, Gentile PF, Bruno T, et al:
Cytotoxic effect of the association of BCNU with rhein or
lonidamine on a human glioma cell line. Anticancer Res. 11:789–792.
1991.PubMed/NCBI
|
|
9
|
Lin YJ and Zhen YS: Rhein lysinate
suppresses the growth of breast cancer cells and potentiates the
inhibitory effect of Taxol in athymic mice. Anticancer Drugs.
20:65–72. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin YJ, Zhen YZ, Shang BY and Zhen YS:
Rhein lysinate suppresses the growth of tumor cells and increases
the anti-tumor activity of Taxol in mice. Am J Chin Med.
37:923–931. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin YJ, Huang YH, Zhen YZ, Liu XJ and Zhen
YS: Rhein lysinate induces apoptosis in breast cancer SK-Br-3 cells
by inhibiting HER-2 signal pathway. Yao Xue Xue Bao. 43:1099–1105.
2008.PubMed/NCBI
|
|
12
|
Schaeffer HJ and Weber MJ:
Mitogen-activated protein kinases: specific messages from
ubiquitous messengers. Mol Cell Biol. 19:2435–2444. 1999.PubMed/NCBI
|
|
13
|
Wen J, Cheng HY, Feng Y, et al: p38 MAPK
inhibition enhancing ATO-induced cytotoxicity against multiple
myeloma cells. Br J Haematol. 140:169–180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shtil AA, Mandlekar S, Yu R, et al:
Differential regulation of mitogen-activated protein kinases by
microtubule-binding agents in human breast cancer cells. Oncogene.
18:377–384. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kyriakis JM and Avruch J: Mammalian
mitogen-activated protein kinase signal transduction pathways
activated by stress and inflammation. Physiol Rev. 81:807–869.
2001.PubMed/NCBI
|
|
16
|
Wang TH, Wang HS, Ichijo H, et al:
Microtubule-interfering agents activate c-Jun N-terminal
kinase/stress-activated protein kinase through both Ras and
apoptosis signal-regulating kinase pathway. J Biol Chem.
273:4928–4936. 1998. View Article : Google Scholar
|
|
17
|
Stone AA and Chambers TC: Microtubule
inhibitors elicit differential effects on MAP kinase (JNK, ERK, and
p38) signaling pathways in human KB-3 carcinoma cells. Exp Cell
Res. 254:110–119. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhen YZ, Lin YJ, Li Y and Zhen YS:
Lidamycin shows highly potent cytotoxic to myeloma cells and
inhibits tumor growth in mice. Acta Pharmacol Sin. 30:1025–1032.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhen YZ, Lin YJ, Shang BY and Zhen YS:
Enediyne lidamycin induces apoptosis in human multiple myeloma
cells through activation of p38 mitogen-activated protein kinase
and c-Jun NH2-terminal kinase. Int J Hematol. 90:44–51. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hideshima T, Catley L, Yasui H, et al:
Perifosine, an oral bioactive novel alkylphospholipid, inhibits Akt
and induces in vitro and in vivo cytotoxicity in human multiple
myeloma cells. Blood. 107:4053–4062. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park WH, Seol JG, Kim ES, et al: Induction
of apoptosis by vitamin D3 analogue EB1089 in NCI-H929 myeloma
cells via activation of caspase 3 and p38 MAP kinase. Br J
Haematol. 109:576–583. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shimizu T, Nakazato T, Xian MJ, Sagawa M,
Ikeda Y and Kizaki M: Resveratrol induces apoptosis of human
malignant B cells by activation of caspase-3 and p38 MAP kinase
pathways. Biochem Pharmacol. 71:742–750. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pei XY, Dai Y, Tenorio S, et al: MEK1/2
inhibitors potentiate UCN-01 lethality in human multiple myeloma
cells through a Bim-dependent mechanism. Blood. 110:2092–2101.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lunghi P, Giuliani N, Mazzera L, et al:
Targeting MEK/MAPK signal transduction module potentiates
ATO-induced apoptosis in multiple myeloma cells through multiple
signaling pathways. Blood. 112:2450–2462. 2008. View Article : Google Scholar
|
|
25
|
Ozaki K, Kosugi M, Baba N, et al: Blockade
of the ERK or PI3K-Akt signaling pathway enhances the cytotoxicity
of histone deacetylase inhibitors in tumor cells resistant to
gefitinib or imatinib. Biochem Biophys Res Commun. 391:1610–1615.
2010. View Article : Google Scholar : PubMed/NCBI
|