Introduction
Bladder carcinoma (BCa) is the most common
genitourinary malignancy worldwide. The disease has been annually
diagnosed in 55,600 males and 17,910 females in the USA and is the
fourth most common type of cancer in males and the eighth most
common in females (1). Fifty to
seventy percent of initially diagnosed BCa patients experience a
recurrence within 5 years; however, 10% progress to invasive BCa
(2). The diverse biological
behavior of BCa has forced current guidelines to recommend intense
follow-up and invasive treatment, as no reliable method to
determine the recurrence and invasive potential of such tumors has
been identified (3). It is
important to predict the invasive behavior of BCa in clinical
studies, and specific molecular markers that may serve as credible
potential prognostic factors are required.
High mobility group box (HMGB) proteins are
ubiquitous, abundant nuclear proteins with diverse functions in the
cell. The proteins were first purified from nuclei in the 1970s and
termed ‘high mobility group’ (HMG) proteins to reflect their rapid
mobility in sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) gels (4,5). HMGB1
and HMGB2 are the main members of the HMGB protein family. Unlike
HMGB1, HMGB2 is highly expressed during embryogenesis, but has
limited expression in adult organs, mainly in the lymphoid organs
and testes. HMGB1 and HMGB2 are highly conserved with >80% amino
acid identity and similar molecular structures; both proteins have
a tripartite domain organization, consisting of two DNA-binding
domains, the HMG-boxes A and B, and acidic C-terminal tails of
variable length (6,7). HMGB1 and HMGB2 have indistinguishable
biological properties within the nucleus, such as binding to DNA
without sequence specificity and regulating transcription,
replication, DNA repair and recombination (8–10).
HMGB1 and HMGB2 are important in growth and development. An early
study demonstrated that HMGB1 knockout mice did not survive within
24 h after birth, due to hypoglycemia, and they exhibited a defect
in the transcriptional function of the glucocorticoid receptor.
Additionally, the HMGB2 knockout mice survived into adulthood,
although spermatogenesis was impaired (6).
In cancer chemotherapy, HMGB1 and HMGB2 may act as
sensors of DNA modification and facilitate p53 phospho rylation
following exposure to genotoxic stress, thus becoming newly
identified components of the DNA damage signaling cascade and
providing novel promising targets for chemotherapeutic intervention
(7,11). HMGB1 has also been demonstrated to
be released into the extra cellular medium and to exhibit important
effects in the mediation of tumor growth, angiogenesis and
metastasis (12,13). Overexpression of HMGB1 and HMGB2 has
been observed in several human cancer types, including
hepatocellular (14), skin squamous
cell (15), prostate (16), gastrointestinal (12,17)
and breast carcinomas (18,19). HMGB1 has been identified to be a
latent pro-angiogenic factor in cancer progression and
angiogenesis; it has been observed to initiate the production of
angiogenic factors, such as vascular endothelial growth factor
(VEGF) (20,21). As a surrogate marker for
angiogenesis, microvessel density (MVD) has been demonstrated to be
predictive of progression and a poor prognosis of BCa (22,23).
Although there has been extensive characterization of the various
roles of HMGB1 in cancer, considerably less is known regarding the
involvement of HMGB2 in carcinogenesis, including its angiogenic
effects and precise signaling pathways.
The present study investigated the
clinicopathological significance of HMGB1 and HMGB2 expression in
human BCa by quantitative real-time polymerase chain reaction (PCR)
and immunohistochemistry. Furthermore, the correlations between the
expression of HMGB1/HMGB2 and VEGF as well as MVD counts have been
analyzed, to explore the angiogenic role of HMGB1 and HMGB2 in
BCa.
Materials and methods
Patients and tissue samples
Tumor tissue samples were collected from 64 patients
with BCa and 15 normal bladder tissue samples were obtained from
the Department of Urology, Huashan Hospital between January 2010
and July 2011. All tissues were confirmed by histological
examination of sequential sections. Tumor staging was determined
according to the sixth edition of the tumor node metastasis (TNM)
classification of the International Union Against Cancer. Clinical
information regarding the samples is described in detail in
Table I. Ethical approval was
obtained from the research ethics committee of Huashan Hospital,
and written informed consent was obtained from all patients.
 | Table ICorrelation between HMGB1 or HMGB2
protein expression and clinicopathological features of the 64
patients with BCa. |
Table I
Correlation between HMGB1 or HMGB2
protein expression and clinicopathological features of the 64
patients with BCa.
| Parameter | Total | High HMGB1
expression | P-value | High HMGB2
expression | P-value |
|---|
| Gender | | | 0.637 | | 0.154 |
| Female | 18 | 9 | | 3 | |
| Male | 46 | 26 | | 16 | |
| Age (years) | | | 0.451 | | 0.784 |
| ≤65 | 32 | 16 | | 10 | |
| >65 | 32 | 19 | | 9 | |
| T stage | | | 0.002 | | 0.010 |
| Ta-T1 | 42 | 17 | | 8 | |
| T2–T4 | 22 | 18 | | 11 | |
| Pathological
grade | | | <0.001 | | 0.016 |
| Low | 35 | 12 | | 6 | |
| High | 29 | 23 | | 13 | |
Real-time PCR
Total RNA was isolated from the tumor and normal
tissues using TRIzol reagent (Invitrogen Life Technologies;
Carlsbad, CA, USA). Total RNA (2 μg) was reverse transcribed
to cDNA using M-MLV reverse transcriptase (Promega Corp., Madison,
WI, USA). A typical 25 μl reaction mixture contained 12.5
μl 2X One Step SYBR RT-PCR buffer (Takara Bio. Inc., Shiga,
Japan), 11 μl of water, 0.5 μl of template and 1
μl of specific primers. The primer sequences were as
follows: forward: 5′-ATATGGCAAAAGCGGA CAAG-3′ and reverse:
5′-GCAACATCACCAATGGACAG-3′ for HMGB1; forward:
5′-CGTTCCTCCCAAAGGTGATA-3′ and reverse: 5′-TCTTTGGCTGACTGCTCAGA-3′
for HMGB2; forward: 5′-GGCGGCACCACCATGTACCCT-3′ and reverse:
5′-AGGGGCCGGACTCGTCATACT-3′ for β-actin. An initial
denaturation/activation step (15 sec at 95°C) was followed by 40
cycles (5 sec at 95°C and 30 sec at 60°C). The relative expression
of HMGB1 and HMGB2 mRNA was calculated with the 2−ΔΔCt
method and normalized using the β-actin mRNA expression level. All
experiments were performed in triplicate.
Immunohistochemistry and evaluation
Formalin-fixed and paraffin-embedded tissue sections
(5 mm) were dewaxed with xylene and rehydrated through an ethanol
gradient into water. Following blocking of endogenous peroxidase
activity with 0.3% hydrogen peroxide for 10 min, the sections were
washed with phosphate-buffered saline (PBS) and incubated overnight
with rabbit anti-HMGB1 antibody (ab92310; Abcam; Cambridge, MA,
USA) or HMGB2 antibody (ab11973; Abcam), VEGF antibody (ab1316;
Abcam) or CD34 antibody (ab81289; Abcam) at the dilution of 1:100
in a humidified chamber at at 4°C. After washing with PBS, sections
were incubated with biotinylated secondary antibody for 30 min at
37°C and then with horseradish peroxidase-labeled streptavidin for
30 min at 37°C. Diaminobenzidine (DAB) was used as chromogen and
the sections were subsequently counterstained with hematoxylin,
then dehydrated, cleared and mounted.
Sections were evaluated and scored as described
previously by Yang et al(24); the extent of staining was scored as
0 (0%), 1 (1–25%), 2 (26–50%), 3 (51–75%) or 4 (76–100%), according
to the percentage of positively stained areas. The staining
intensity was scored as 0 (negative), 1 (weak), 2 (medium) or 3
(strong). The first and second scores were then added together to
produce the final score (0–7) for HMGB1, HMGB2 or VEGF. Tumors with
a final staining score of >3 were considered to have high
protein expression and <2 were consi dered to have low protein
expression.
CD34-stained whole sections were used to evaluate
MVD as described previously by Ajili et al and Deniz et
al(22,23). Microvessel counts of five areas with
the most intense neovascularization were performed at a
magnification of ×200.
Statistical analysis
The χ2 test was used to analyze
differences in categorical variables. Statistical analyses were
performed using a Student’s two-tailed unpaired t-test for
comparisons between two groups. All computations were performed
with the Statistical Package for the Social Sciences (SPSS) 12.0
software (SPSS, Inc.; Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
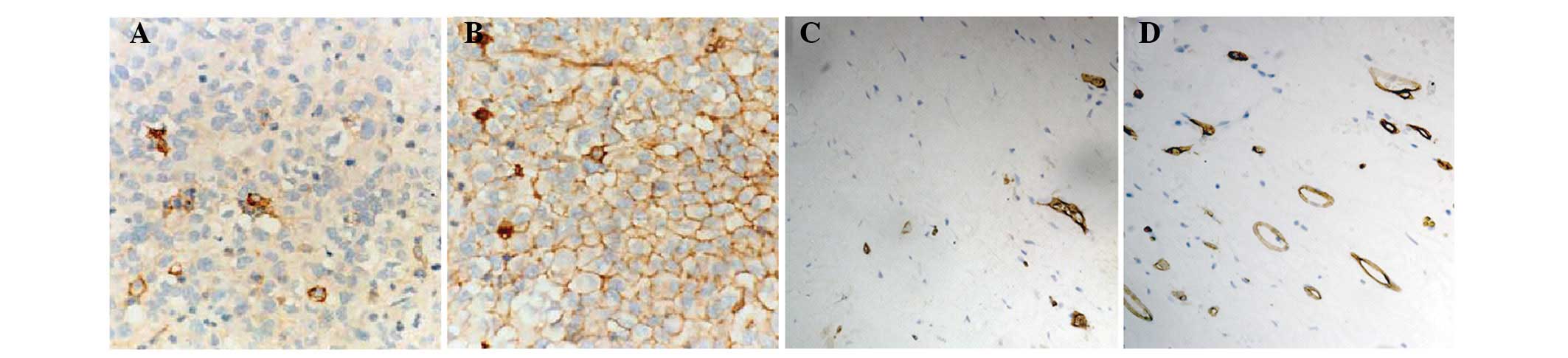
HMGB1 and HMGB2 are overexpressed in
BCa
Immunohistochemical analysis demonstrated that HMGB1
and HMGB2 were negative in the majority of normal bladder
urothelia, but were positive in BCa (Fig. 1). A total of 35 (55%) cases and 19
(30%) cases had high expression of HMGB1 and HMGB2 protein,
respectively (Table I). The
correlation between HMGB1 and HMGB2 protein expression and the
clinicopathological features of BCa demonstrated that levels of
both HMGB1 and HMGB2 protein expression were significantly
correlated with tumor grade and stage (Table I), and were not correlated with the
remaining clinicopathological features tested, including gender and
age.
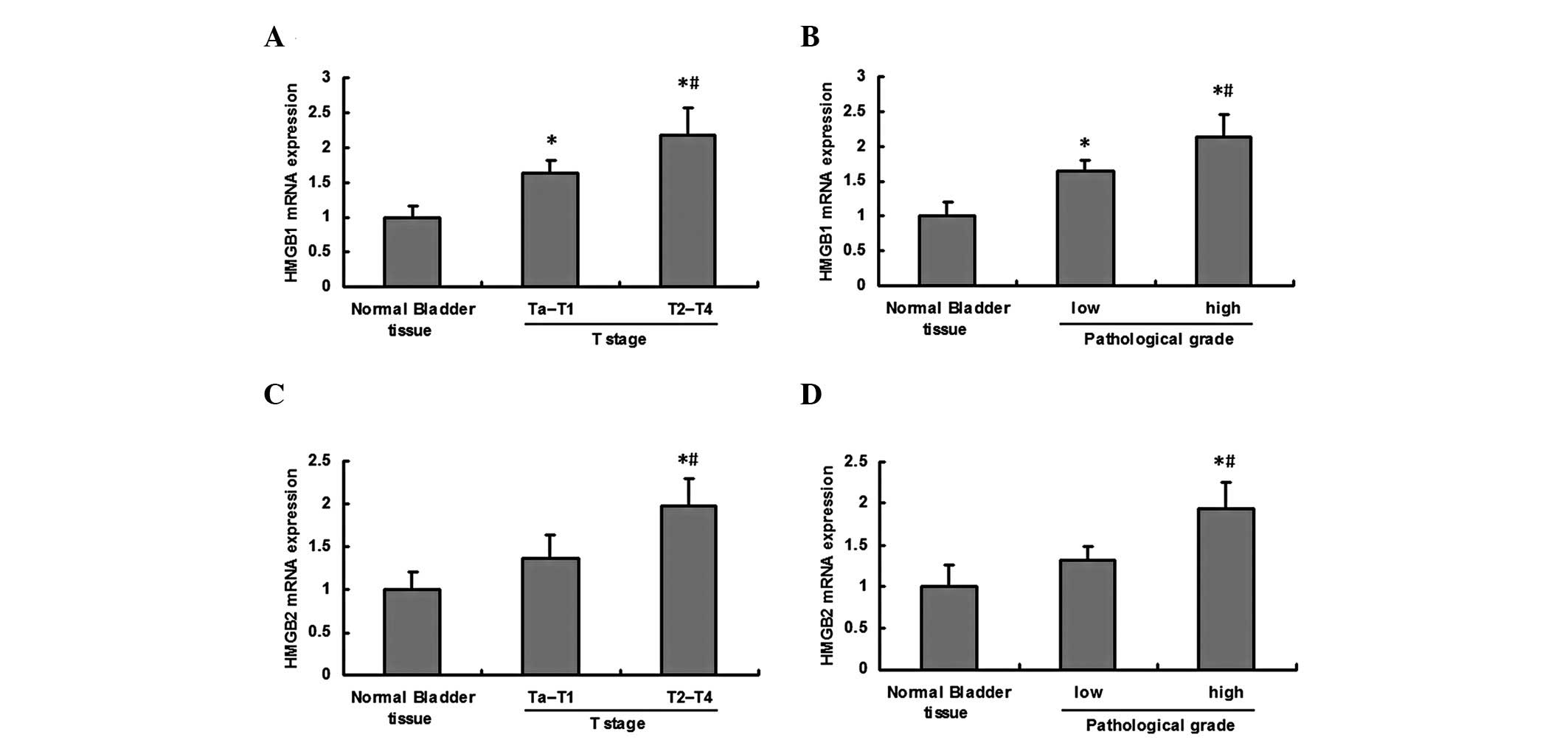
Upregulation of HMGB1 and HMGB2 mRNA in
BCa
Real-time PCR analysis of HMGB1 mRNA expression
revealed that the mRNA expression of HMGB1 was significantly
upregul ated in BCa, and the mRNA expression in T2–T4 stage or high
pathological grade tumor tissue was significantly higher than that
in Ta-T1 stage or low pathological grade tumor tissue (Fig. 2A and B). The mRNA expression of
HMGB2 was significantly upregulated in T2–T4 stage or high
pathological grade tumor tissue compared with normal bladder tissue
and Ta-T1 stage or low pathological grade tumor tissue; whereas no
marked differences were observed between normal bladder tissue and
low grade tumor tissue (Fig. 2C and
D).
Correlation between HMGB1 and HMGB2
protein expression and angiogenesis
To investigate angiogenesis in BCa,
immunohistochemistry of VEGF and CD34 was performed. As
demonstrated in Fig. 3 and Table II, levels of VEGF protein expression
and MVD were significantly correlated with levels of HMGB1 and
HMGB2 protein expression.
| Table IICorrelation between HMGB1 or HMGB2
protein expression and VEGF expression or MVD. |
Table II
Correlation between HMGB1 or HMGB2
protein expression and VEGF expression or MVD.
| Parameter | Total | Low VEGF
expression | High VEGF
expression | P-value | MVD | P-value |
|---|
| HMGB1 | | | | <0.001 | | <0.001 |
| Low
expression | 29 | 19 | 10 | | 23.0±6.4 | |
| High
expression | 35 | 7 | 28 | | 35.5±10.8 | |
| HMGB2 | | | | 0.038 | | <0.001 |
| Low
expression | 45 | 22 | 23 | | 26.6±9.6 | |
| High
expression | 19 | 4 | 15 | | 37.6±10.3 | |
Discussion
Our study found that HMGB1 and HMGB2 were
overexpressed in BCa tissues compared with normal tissues, and were
correlated with both the clinical stage and pathological grade of
the tumor. In addition, overexpression of HMGB1/HMGB2 was also
correlated with VEGF expression and MVD counts. This implies that
HMGB1 and HMGB2 are likely to have key roles in the progression of
BCa.
Overexpression of HMGB1 has been observed in several
human tumor types. HMGB1 may be involved in cancer progression and
prognosis, including apoptosis, angiogenesis, the inflammatory
microenvironment, mobility, invasion, metastasis and patient
survival (9,20). During cancer progression, HMGB1 has
been observed to modulate the NF-κB, PI3K/AKT and mitogen-activated
protein kinase (MAPK) signaling pathways by interacting with the
receptor for advanced glycation end products (RAGE), while blockade
of the HMGB1/RAGE interaction has been demonstrated to suppress
tumor growth and metastasis (25,26).
HMGB2 is highly homologous to HMGB1 and may exhibit similar effects
in neoplastic development (27).
Although HMGB1 has been studied more extensively, considerably less
is known regarding HMGB2 in the study of cancer, particularly its
relevance in carcinogenesis. To our knowledge, overexpression of
HMGB2 has been identified in several types of tumors, including
hepatocellular carcinoma (HCC) and skin cancer, and has been
involved in cancer progression and prognosis via interacting with
RAGE, steroid receptors, p53 and p73 (14,15).
In addition, HMGB2 has been demonstrated to be significantly
downregulated by the anti-human epidermal growth factor receptor 2
antibody through the AKT pathway in breast cancer cell lines
(28). The present study indicated
that HMGB1 and HMGB2 genes and proteins were overexpressed in BCa,
using real-time PCR and immunohistochemical examin ation. The
overexpression of HMGB/HMGB2 was correlated with both the clinical
stage and pathological grade of the tumor, and HMGB1 and HMGB2 may
be involved in BCa development and progression. Their involvement
in the regulation of the carcinogenesis of BCa requires further
investigation.
HMGB1 is a latent pro-angiogenic factor that has
been observed to act both directly and indirectly, and an antibody
targeting HMGB1 has been demonstrated to inhibit the angio-genesis
in vitro and in vivo(29). HMGB1 was able to directly induce
sprouting of endothelial cell spheroids in a collagen gel, and to
stimulate endothelial cell proliferation, chemotaxis and repair of
a wounded monolayer (30). HMGB1
exerts its angio-genetic effects via the RAGE pathway in tumor
cells; it has been demonstrated to increase the expression of
angiogenic growth factors, including VEGF, and to attract
macrophages, which also produce a number of potent angiogenetic
cytokines and growth factors (31).
Furthermore, HMGB1 has been observed to attract endothelial
progenitor cells (EPCs) and hematopoietic stem cells to sites of
tissue injury and tumors to improve neovascularization mediated by
RAGE, and anti-RAGE antibodies were able to inhibit this process
(32). The role of HMGB2 in
angiogenesis has not yet been studied. We found that overexpression
of HMGB1 and HMGB2 were connected with VEGF expression and MVD
counts in BCa tissues, indicating that they have important roles in
angiogenesis, partly through VEGF. However, the precise molecular
mechanisms require further study.
In conclusion, our data revealed that the expression
levels of HMGB1 and HMGB2 were highly increased in BCa, and that
HMGB1/HMGB2 overexpression was significantly correlated with
malignant tumor progression and angiogenesis. As potential
diagnostic and therapeutic targets, further studies are required to
understand the molecular mechanism of the involvement of HMGB1 and
HMGB2 in BCa.
Acknowledgements
This study was supported by a grant
from the Science and Technology Commission of Shanghai Municipality
(Project No. 074119519).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar
|
|
2
|
Ghoneim MA, Abdel-Latif M, el-Mekresh M,
et al: Radical cystectomy for carcinoma of the bladder: 2,720
consecutive cases 5 years later. J Urol. 180:121–127.
2008.PubMed/NCBI
|
|
3
|
Vishnu P, Mathew J and Tan WW: Current
therapeutic strategies for invasive and metastatic bladder cancer.
Onco Targets Ther. 4:97–113. 2011.PubMed/NCBI
|
|
4
|
van Beijnum JR, Buurman WA and Griffioen
AW: Convergence and amplification of toll-like receptor (TLR) and
receptor for advanced glycation end products (RAGE) signaling
pathways via high mobility group B1 (HMGB1). Angiogenesis.
11:91–99. 2008.PubMed/NCBI
|
|
5
|
Zhang J, McCauley MJ, Maher LJ III, et al:
Mechanism of DNA flexibility enhancement by HMGB proteins. Nucleic
Acids Res. 37:1107–1114. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stros M, Polanská E, Struncová S and
Pospísilová S: HMGB1 and HMGB2 proteins up-regulate cellular
expression of human topoisome rase IIalpha. Nucleic Acids Res.
37:2070–2086. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Krynetskaia NF, Phadke MS, Jadhav SH and
Krynetskiy EY: Chromatin-associated proteins HMGB1/2 and PDIA3
trigger cellular response to chemotherapy-induced DNA damage. Mol
Cancer Ther. 8:864–872. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Balani P, Boulaire J, Zhao Y, et al: High
mobility group box2 promoter-controlled suicide gene expression
enables targeted glioblastoma treatment. Mol Ther. 17:1003–1011.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Naglova H and Bucova M: HMGB1 and its
physiological and pathological roles. Bratisl Lek Listy.
113:163–171. 2012.PubMed/NCBI
|
|
10
|
Franklin S, Chen H, Mitchell-Jordan S, et
al: Quantitative analysis of the chromatin proteome in disease
reveals remodeling principles and identifies high mobility group
protein b2 as a regulator of hypertrophic growth. Mol Cell
Proteomics. 11:M111.0142582012. View Article : Google Scholar
|
|
11
|
Dong Xda E, Ito N, Lotze MT, et al: High
mobility group box I (HMGB1) release from tumor cells after
treatment: implications for development of targeted
chemoimmunotherapy. J Immunother. 30:596–606. 2007.PubMed/NCBI
|
|
12
|
Yao X, Zhao G, Yang H, et al:
Overexpression of high-mobility group box 1 correlates with tumor
progression and poor prognosis in human colorectal carcinoma. J
Cancer Res Clin Oncol. 136:677–684. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shen X, Hong L, Sun H, et al: The
expression of high-mobility group protein box 1 correlates with the
progression of non-small cell lung cancer. Oncol Rep. 22:535–539.
2009.PubMed/NCBI
|
|
14
|
Kostova N, Zlateva S, Ugrinova I and
Pasheva E: The expression of HMGB1 protein and its receptor RAGE in
human malignant tumors. Mol Cell Biochem. 337:251–258. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sharma A, Ray R and Rajeswari MR:
Overexpression of high mobility group (HMG) B1 and B2 proteins
directly correlates with the progression of squamous cell carcinoma
in skin. Cancer Invest. 26:843–851. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gnanasekar M, Thirugnanam S and Ramaswamy
K: Short hairpin RNA (shRNA) constructs targeting high mobility
group box-1 (HMGB1) expression leads to inhibition of prostate
cancer cell survival and apoptosis. Int J Oncol. 34:425–431.
2009.PubMed/NCBI
|
|
17
|
Song B, Song WG, Li ZJ, et al: Effect of
HMGB1 silencing on cell proliferation, invasion and apoptosis of
MGC-803 gastric cancer cells. Cell Biochem Funct. 30:11–17. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rhodes DR, Yu J, Shanker K, et al:
Large-scale meta-analysis of cancer microarray data identifies
common transcriptional profiles of neoplastic transformation and
progression. Proc Natl Acad Sci USA. 101:9309–9314. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiao Y, Wang HC and Fan SJ: Growth
suppression and radio-sensitivity increase by HMGB1 in breast
cancer. Acta Pharmacol Sin. 28:1957–1967. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang D, Kang R, Zeh HJ III and Lotze MT:
High-mobility group box 1 and cancer. Biochim Biophys Acta.
1799:131–140. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sitohy B, Nagy J and Dvorak H:
Anti-VEGF/VEGFR therapy for cancer: reassessing the target. Cancer
Res. 72:1909–1914. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ajili F, Kacem M, Tounsi H, et al:
Prognostic impact of angio-genesis in nonmuscle invasive bladder
cancer as defined by microvessel density after immunohistochemical
staining for CD34. Ultrastruct Pathol. 36:336–342. 2012. View Article : Google Scholar
|
|
23
|
Deniz H, Karakök M, Yagci F and Güldür ME:
Evaluation of relationship between HIF-1alpha immunoreactivity and
stage, grade, angiogenic profile and proliferative index in bladder
urothelial carcinomas. Int Urol Nephrol. 42:103–107. 2012.
View Article : Google Scholar
|
|
24
|
Yang GL, Zhang LH, Bo JJ, et al: Increased
expression of HMGB1 is associated with poor prognosis in human
bladder cancer. J Surg Oncol. 106:57–61. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang D, Kang R, Cheh CW, et al: HMGB1
release and redox regulates autophagy and apoptosis in cancer
cells. Oncogene. 29:5299–5310. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Todorova J and Pasheva E: High mobility
group B1 protein interacts with its receptor RAGE in tumor cells
but not in normal tissues. Oncol Lett. 3:214–218. 2012.PubMed/NCBI
|
|
27
|
Pusterla T, de Marchis F, Palumbo R and
Bianchi ME: High mobility group B2 is secreted by myeloid cells and
has mitogenic and chemoattractant activities similar to high
mobility group B1. Autoimmunity. 42:308–310. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Le XF, Lammayot A, Gold D, et al: Genes
affecting the cell cycle, growth, maintenance, and drug sensitivity
are preferentially regulated by anti-HER2 antibody through
phosphatidylinositol 3-kinase-AKT signaling. J Biol Chem.
280:2092–2104. 2005.
|
|
29
|
van Beijnum JR, Nowak-Sliwinska P, van den
Boezem E, et al: Tumor angiogenesis is enforced by autocrine
regulation of high-mobility group box 1. Oncogene. Mar 5–2012.(Epub
ahead of print). View Article : Google Scholar
|
|
30
|
Sachdev U, Cui X, Hong G, et al: High
mobility group box 1 promotes endothelial cell angiogenic behavior
in vitro and improves muscle perfusion in vivo in response to
ischemic injury. J Vasc Surg. 55:180–191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Urquidi V, Goodison S, Kim J, et al:
Vascular endothelial growth factor, carbonic anhydrase 9, and
angiogenin as urinary bio markers for bladder cancer detection.
Urology. 79:1185.e1–6. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chavakis E, Hain A, Vinci M, et al:
High-mobility group box 1 activates integrin-dependent homing of
endothelial progenitor cells. Circ Res. 100:204–212. 2007.
View Article : Google Scholar : PubMed/NCBI
|