Introduction
The John Cunningham virus (JCV) constitutes a family
of polyoma viruses, which are non-enveloped, have icosahedral
capsids and contain small, circular, double-stranded DNA genomes.
These viruses feature early and late coding regions, whose
transcription is initiated in opposite directions by an interposed
transcription control region. The early region is alternatively
spliced to produce small t antigen and large T
antigen, a large phosphoprotein that binds to the viral
replication region to promote double helix unwinding and
recruitment of cell proteins that are required for DNA synthesis.
The late region encodes the capsid structural proteins VP1, VP2 and
VP3 due to alternative splicing and the small regulatory element
known as agnoprotein. Viral proteins (VP) are essential for
assembly with viral DNA to form virions (1,2).
Serological studies have indicated asymptomatic JCV
infection in approximately 90% of the adult population. As JCV
replication is restricted to glial and lymphoid cells, which
contain the JCV transcriptional factors, the virus enters through
tonsillar stromal tissue and persists in a quiescent state in the
kidney and lymphoid tissue during latency. However, it may be
activated under immunosuppressive conditions, leading to the lethal
demyelinating disease, progressive multifocal leukoencephalopathy
(PML). JCV infection is initiated with binding to JCV-sensitive
cell surfaces. Additionally, JCV capsids undergo endocytosis and
are transported to the nucleus where the viral DNA is uncoated and
the early region begins to be transcribed. Under permissive
infection, replication of viral DNA may result in lytic infection
with viral amplification, but in non-permissive cells not allowing
viral replication, abortive infection or cell transformation is the
result (1–6).
JCV is capable of transforming cells, as manifested
by distinct phenotypic and morphological changes, such as growth in
agar, rapid division, prolongation of the life span, enhanced
production of plasminogen activator, anchorage-dependent growth,
unstable multicentric chromosomes, centric and acentric rings and
the ability to form dense foci in culture (7). Intravenous or intracranial inoculation
of JCV into experimental animals has been found to cause
astrocytomas, glioblastomas, neuroblastomas and medulloblastomas
(1). In addition, transgenic mice
expressing the JCV T antigen exhibit a 50% incidence of pituitary
adenomas by 1 year of age and some of the mice develop malignant
peripheral nerve sheath tumors (1,8). In
recent years, the presence of JCV has been suggested to correlate
with various types of human neoplasms, including tongue,
pharyngeal, gastric, esophageal, colorectal, anal and prostatic
cancer, brain tumors and lung carcinomas B (9–21). In
our laboratory, the transgenic model of lens tumors induced by JCV
T antigen was previously established for the first time (22). These reports indicate that JCV plays
a direct role in human tumorigenesis as an oncovirus.
Molecular mechanisms underlying oncogenesis by JCV
may center on its encoded products, T antigen, VP and agnoprotein.
The JCV T antigen serves as an ATPase, helicase and polymerase, and
orchestrates the assembly and function of cell proteins such as
replication protein A and DNA polymerase-α. It may also inactivate
p53 and members of the pRb family, and deregulate the Wnt signaling
pathway to promote uncontrolled proliferation and immortal
survival. The induction of structural chromosome aberrations and
genomic instability has been described (1–3,6,7).
It was found that T antigen inhibited AP2 binding to an AP2 site in
the BAG3 promoter, resulting in downregulation of the BAG3 promoter
expression and subsequent apoptotic inhibition, since BAG3
functions as a molecular co-chaperone through its interaction with
Hsc70/Hsp70 and is capable of initiating apoptosis (23). Reviriego-Mendoza et al
(24) have demonstrated that JCV
large T antigen binds the F-box proteins β-transducin-repeat
containing protein-1 and -2 (βTrCP1/2) and disrupts further
proteasomal degradation of β-catenin. JCV T antigen expression in
CRC associates with a metastatic phenotype, which may partly be
mediated through the AKT/MAPK signaling pathway.
Despite a worldwide decline in its incidence and
mortality since the second half of the 20th century, gastric
carcinoma still ranks as the fourth most common and the second most
frequent cause of mortality from cancer (25). Colorectal carcinoma is one of the
most common types of cancer in the world, accounting for almost 10%
of all new cases of cancer. These cancers continue to be a major
health concern, although sophisticated diagnostic and surgical
techniques are widely applied in clinical practice (26). JCV has been found in high
concentrations in urban sewage worldwide and is closely linked to
gastrointestinal carcinogenesis, leading some investigators to
suspect contaminated drinking water as a typical route of infection
(27). However, there is a lack in
the evidence regarding the correlation between JCV existence and
gastrointestinal carcinogenesis in China. Therefore, we employed
nested PCR and Southern blotting to detect JCV genomic DNA in the
frozen and paraffin-embedded samples of Chinese gastric and
colorectal carcinomas. Additionally, we aimed to clarify the
pathobiological features of JCV-related gastrointestinal
carcinomas.
Materials and methods
Subjects
Formalin-fixed and paraffin-embedded gastric and
colorectal carcinoma (n=40, 43) and adjacent non-neoplastic mucosa
(NNM) (n=42, 23) were collected from surgical materials in
Shengjing Hospital, China Medical University after obtaining
permission from the patients or their relatives. Frozen gastric
(n=22) and colorectal (n=18) carcinomas and matched NNM were also
collected here. The Ethics Committee of the Hospital gave approval
for the genetic experiments restricted to JCV.
DNA extraction and checking
Paraffin-embedded tissues were sectioned at 10 μm
and lesions were microdissected with reference to the hematoxylin
and eosin (HE) staining of consecutive sections and subjected to
deparaffinization. DNA was extracted from cell lines and the
deparaffinized samples by standard proteinase K digestion and
phenol/chloroform extraction. The DNA samples were amplified using
β-globin primers; sense: 5′-ACACAACTGTGTTCACTAGC-3′ and anti-sense:
5′-GTCTCCTTAAACCTGTCTTG-3′ (175 bp) by 30 cycles of denaturation at
95°C for 30 sec, annealing at 55°C for 30 sec, and extension at
72°C for 30 sec to confirm its integrity.
Nested PCR
PCR amplification was performed using three
individual sets of primers for T-antigen: T1 and T2 (nucleotides
3049-3069 of the Mad-1 strain, 5′-TGG CCTGTAAAGTTCTAGGCA-3′ and
3229-3207, 5′-GCAGAG TCAAGGGATTTACCTTC-3′, respectively), which
amplify sequences in the NH2-terminal region of the JCV T-antigen
were used for the first PCR, whereas T1 and T3 (nucleotides
3193-3171, 5′-AGCAACCTTGATTGCTTAAGAGA-3′) were used for the second
PCR (145 bp). For the VP capsid gene sequence, VP1 and VP2
(nucleotides 1828-1848, 5′-TGTGCACTC TAATGGGCAAGC-3′ and 2019-39,
5′-CTAGGTACGCCT TGTGCTCTG-3′, respectively) were used for the first
PCR, followed by VP1 and VP3 (nucleotides 2004-1982, 5′-GAT
TGCACTGTGGCATTCTTTGG-3′) for the second PCR (177 bp). Finally, for
JCV agnoprotein, AGNO1 and AGNO2 (nucleotides 280-298,
5′-GTCTGCTCAGTCAAACCACTG-3′; and 458-438,
5′-GTTCTTCGCCAGCTGTCAC-3′, respe- ctively), which amplify a region
within the coding region of the JCV agnoprotein, were used for the
first PCR and AGNO1 and AGNO3 (nucleotides 395-415, 5′-GCACAGG
TGAAGACAGTGTAG-3′) were used for the second PCR (64 bp). Reaction
mixtures (25 μl) contained 0.125 μl Takara Ex Taq HS (Takara,
Japan) with 2.0 mM MgCl2, 2.5 μl 10X PCR buffer, 2 μl
dNTP mixture, 1 μM of each primer set (external primers) and 100 ng
of template DNA. PCR conditions were denaturation at 95°C for 8
min, followed by 30 cycles of denaturation at 95°C for 15 sec,
annealing for 30 sec and extension at 72°C for 30 sec. The
annealing steps were carried out at temperatures of 55°C for the T
antigen primers, 57°C for the agnoprotein primers and 54°C for the
VP primers. As a termination step, the extension time of the last
cycle was increased to 7 min. The pBSK-T28 or pBSK-JCV plasmid
(28) was used as a positive
control. Samples amplified in the absence of template DNA were
employed as negative controls. Nested PCR was carried out as the
first PCR cycle, using 1% (volume) of the first PCR product with
the internal primers in each case.
Southern blotting
Southern blotting was performed by resolving 10 μl
of each nested PCR product on 2% agarose gels stained with ethidium
bromide. After gels were denatured, neutralized and blotted onto
nylon membranes (Hybond N1, Amersham, Germany), hybridization was
performed using 20 pmol/ml of digoxygenin-labeled oligonucleotide
probes (nucleotide 3066-3101 for T antigen
5′-GGCACTGAATATTCA TTCATGGTTACAATTCCAGGT-3′; nucleotide 1872-1891
for VP 5′-AGCCAGTGCAGGGCACCAGC-3′ and nucleotide 395-415 for
agnoprotein, 5′-AAAGACAGAGACACAGTG GTT-3′) at 55°C overnight. After
washing the membranes with 2X SSC, 0.1% sodium dodecyl sulfate
(SDS), and 0.1X SSC and 0.1% SDS at the same temperature as the
hybrid temperature, the membrane was subjected to incubation of the
alkaline phosphatase (AP)-labeled anti-digoxygenin antibody for 30
min. Luminescence was detected with X film (Fujifilm, Japan) using
a Dig luminescent detection kit for nucleic acids (Boehringer
Mannheim, MA, USA).
Tissue microarrays (TMAs)
HE-stained sections of 103 lung carcinomas were
examined and representative areas of solid tumor were identified
for sampling. Two 2 mm-diameter tissue cores per donor block were
punched out and transferred to a recipient block holding a maximum
of 48 cores using a tissue microarrayer (AZUMAYA KIN-1, Japan).
Sections (4 μm) were consecutively incised from the recipient
blocks and transferred to poly-lysine-coated glass slides. HE
staining was performed for confirmation of tumor samples.
Immunohistochemistry
Serial sections were deparaffinized with xylene,
dehydrated with alcohol and subjected to antigen retrieval by
irradiation in target retrieval solution (TRS, Dako, Carpinteria,
CA, USA) for 5 min with a microwave oven (Oriental Rotor Ltd.,
Tokyo, Japan). Bovine serum albumin (5%) was then applied for 1 min
to prevent non-specific binding. The sections were incubated with
mouse anti-SV40 T antigen (Santa Cruz Biotechnology, Santa Cruz,
CA, USA; 1:100), rabbit anti-ki-67 (Dako, USA; 1:25), rabbit
anti-p53 (Dako, USA; 1:50), rabbit anti-caspase-3 (Dako, USA;
1:150) or mouse anti-human β-catenin (BD Biosciences, Transduction
Labs, Franklin Lake, NJ, USA, 1:200) for 15 min, then treated with
the anti-mouse or anti-rabbit Envison-PO (Dako, USA) antibody for
15 min. The incubations were performed in a microwave oven with
intermittent irradiation, as described previously (29). Following each treatment, the slides
were washed with TBST (10 mM Tris-HCl, 150 mM NaCl, 0.1% Tween-20)
three times for 5 min. The slides were developed for color with
3,3′-diaminobenzidine (DAB) and counterstained with Mayer’s
hematoxylin. Omission of the primary antibody was used as a
negative control.
Evaluation of immunohistochemistry
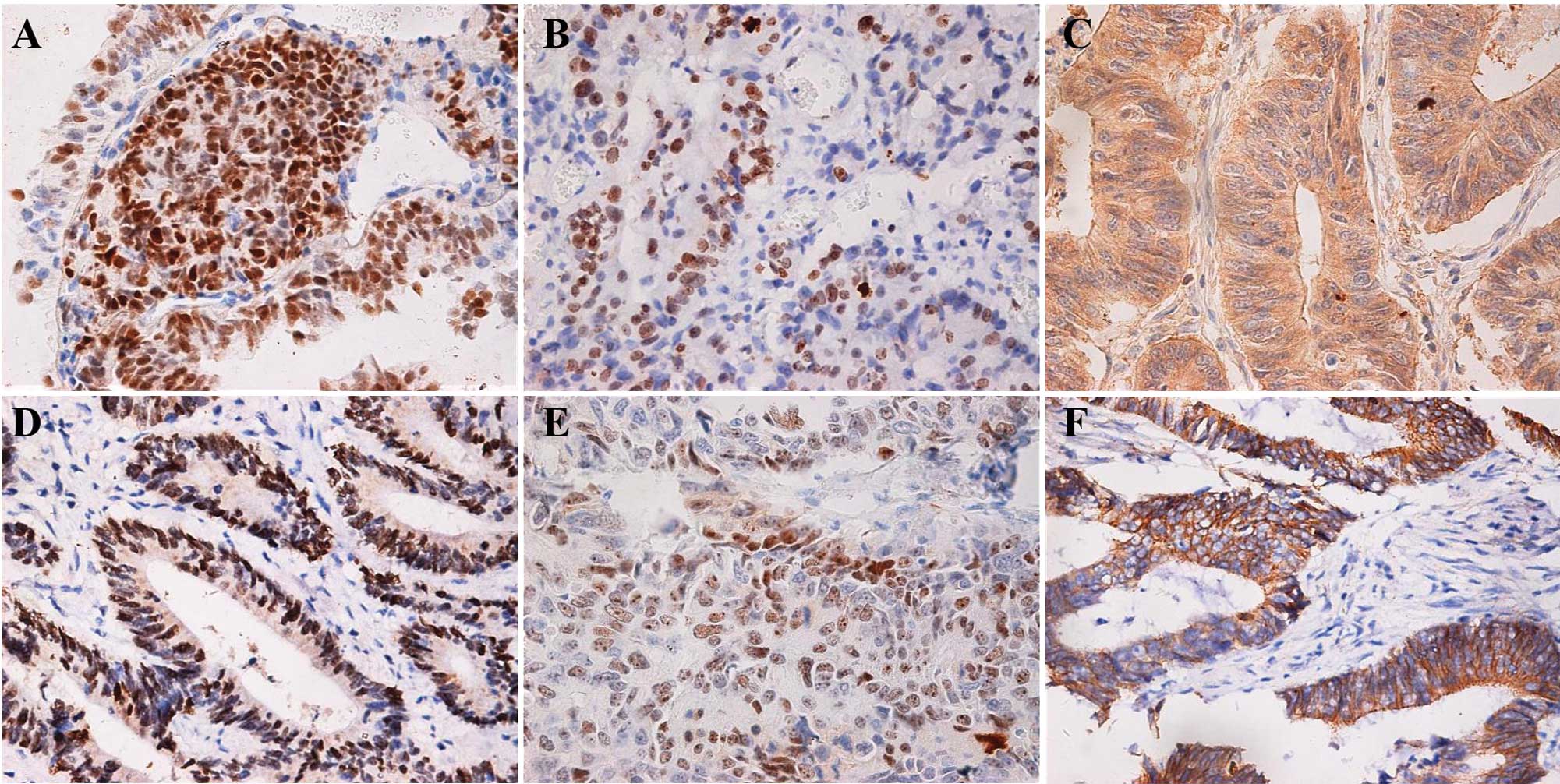
Immunoreactivity for JCV T antigen, ki-67, p53 and
Rb was localized in the nucleus, for caspase-3 in the cytoplasm,
and for β-catenin in the nucleus, cytoplasm or membrane (Fig. 1). A total of 100 cells were randomly
selected and counted from 5 representative fields of each section,
blindly, by two independent observers (Wang JP and Zheng HC). The
percentages of positive cells in total counted were graded
semi-quantitatively using a four-tier scoring system: negative (−),
0–5%; positive (+), 6–100%.
Statistical analysis
Statistical analysis was performed using Fisher’s
test to analyze the positive rates. P<0.05 was considered to be
statistically significant. SPSS 10.0 software was employed to
analyze all data.
Results
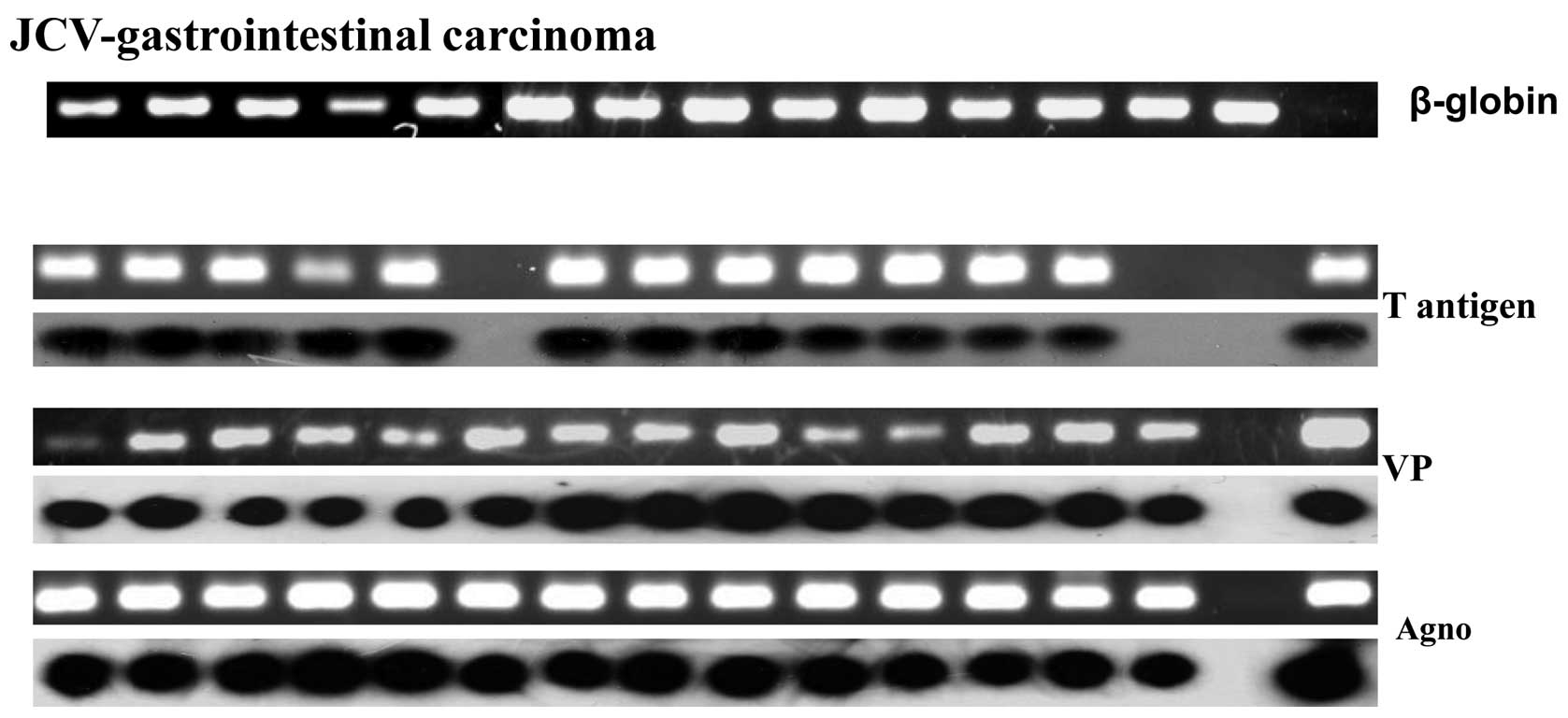
As shown in Fig. 2,
amplification of β-globin was of high quality in all the cases. The
PCR bands of T antigen, VP and agnoprotein were confirmed by the
specific probes in Southern blot analysis, respectively. The
positive rate of JCV T antigen was higher in paraffin-embedded
gastrointestinal carcinomas than adjacent NNM by nested-PCR
followed by Southern blot analysis (36.9 vs. 16.9%, P<0.05)
(Fig. 2 and Table I). However, no difference was
observed in other viral oncogenes, even in paraffin-embedded or
frozen samples. Immunohistochemically, T antigen was detectable in
9.6% (13/135) of carcinoma cases, higher than its positive rate in
NNM (0.8%, 1/126, P<0.01). There was genomic JCV DNA detected in
these T-antigen-positive cases. The genomic JCV DNA existence or
its T antigen expression was not correlated with
clinicopathological parameters of gastrointestinal carcinomas
(P>0.05), including age, gender, tumor size, histological types,
lymph node metastasis, expression of ki-67, caspase-3, p53, Rb and
β-catenin.
 | Table IJCV existence in lung samples by
nested PCR followed by Southern blot targeting T antigen. |
Table I
JCV existence in lung samples by
nested PCR followed by Southern blot targeting T antigen.
| Group | No. | T antigen | VP | Agno |
|---|
| |
|
|---|
| | + | PR (%) | + | PR (%) | + | PR (%) |
|---|
| Frozen |
| Non-neoplastic
mucosa | 40 | 4 | 10.0 | 1 | 2.5 | 3 | 7.5 |
| Carcinoma | 40 | 6 | 15.0 | 3 | 7.5 | 4 | 10.0 |
|
Paraffin-embedded |
| Non-neoplastic
mucosa | 65 | 11 | 16.9 | 11 | 16.9 | 16 | 24.6 |
| Carcinoma | 83 | 29 | 34.9a | 22 | 26.5 | 16 | 19.2 |
Discussion
In 1994, Theodoropoulos et al (30) for the first time employed real-time
PCR successfully to detect JCV in colonic cancer and adjacent
non-cancerous mucosa. At the same time, Schatzl et al
(31) also began to examine the JCV
in the brain and kidney tissue of PML using nested-PCR. In the
present study, we for the first time examined the oncogenic role of
genomic JCV DNA existence and its T antigen expression in Chinese
gastrointestinal carcinoma and found that the positive rates were
higher in carcinoma than that in matched NNM. In our previous
study, it was found that JCV T antigen existence or copies were
closely linked to the lung, tongue, pharyngeal and gastric
carcinogenesis (10,14,15,20).
Hori et al (9) reported that
T antigen was detected in 6 of 23 colorectal cancer cases (26.1%)
and 1 of 21 adenomas (4.8%), but none of the 20 samples of normal
colonic mucosa. These findings indicated that JCV T antigen may be
involved in the malignant transformation of epithelium as evidenced
by our transgenic lens tumor induced by T antigen, although we
failed to establish the animal model of gastric tumor induced by
the JCV T antigen (28).
Furthermore, previous studies have demonstrated the
presence of replicating JCV DNA in B lymphocytes from peripheral
blood, tonsils and spleen. Our previous study showed JCV T antigen
in the nuclei of tonsil lymphocytes by in situ PCR (20), in line with virus persistence in a
quiescent state in lymphoid tissue during latency and infection of
other cells upon immune suppression (32). Detection of viral gene products in
renal tubules and excretion of JC virions in the urine suggest JCV
persistence in the kidney (33). In
the present study, JCV DNA was detectable in gastrointestinal
carcinoma. Earlier surveys of raw sewage from urban areas have
shown the detection of JC viral particles in sewage samples from
widely divergent areas (27), and
the presence of JCV DNA sequences in the upper human
gastrointestinal tract (17),
suggesting a potential re-entry of JCV and/or viral DNA into the
human digestive tracts through the intake of virus-contaminated
water and food.
The JCV T antigen shares more than 70% homology with
the SV40 T antigen, particularly in the N-terminus. Based on SV40
studies, the N-terminal domain serves multiple functions, including
regulation of DNA replication, protein stability and cell
immortalization (34). The protein
region encoded by the large T/small t common exon is crucial in the
control of the cell cycle by interacting with key cell proteins
such as pRb and Rb family members including p107 and p130 (1–5). T
antigen may inhibit cellular apoptosis by binding to the AP2 site
of BAG3 and subsequently downregulate BAG3 promoter activity. In
addition, T antigen binds βTrCP1/2 and disrupts further proteasomal
degradation of β-catenin (23,24).
Therefore, we compared the JCV T antigen with the expression of
ki-67, caspase-3, p53, Rb and β-catenin. However, no significant
correlation has been found, possibly due to the lower rates of JCV
T antigen in Chinese patients with gastrointestinal carcinomas. By
contrast, it was documented that the lung carcinomas with high copy
numbers for the virus were closely associated with high
proliferation and downregulation of cell adhesion mediated by
β-catenin (20).
In the present study, we employed nested-PCR to
screen the JCV genes due to its low number of copies in human
tissue. Since there is some homology between polyomaviruses SV40,
BK and JC virus (4), a specific
probe for JCV T antigen was used to ensure the reality of
these amplicons in the Southern blot analysis. Additionally, we
used the anti-SV40 T antigen antibody to detect the JCV T antigen
with a positive control of transgenic lens tumor-induced JCV T
antigen (data not shown). To avoid false positivity, we evaluated
the T antigen immunoreactivity in combination with nested-PCR
followed by Southern blotting. Notably, JCV T antigen was higher in
paraffin-embedded samples in line with our report, although the JCV
T antigen copies were higher in frozen samples than those in
paraffin-embedded ones (15).
In conclusion, the T antigen of JCV may contribute
to gastrointestinal carcinogenesis and the gastrointestinal tract
may lead to JCV infection. Possible molecular mechanisms of
oncogenesis of the JCV T antigen in gastrointestinal carcinogenesis
and the reasons for high detection rates of JCV genes should be
elucidated in the future.
Acknowledgements
This study was supported by the Shenyang Outstanding
Talent Foundation of China, Liaoning BaiQianWan Talents Program,
Scientific and Technological Projects for Overseas Returned
persons, Ministry of Personnel; Shenyang Science and Technology
Grant (1091175-1-00); Scientific Research Foundation for the
Returned Overseas Chinese Scholars.
References
|
1
|
Reiss K and Khalili K: Viruses and cancer:
lessons from the human polyomavirus, JCV. Oncogene. 22:6517–6523.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Frisque RJ, Bream GL and Cannella MT:
Human polyomavirus JC virus genome. J Virol. 51:458–469.
1984.PubMed/NCBI
|
|
3
|
Zheng HC, Yan L, Cui L, Guan YF and Takano
Y: Mapping the history and current situation of research on John
Cunningham virus - a bibliometric analysis. BMC Infect Dis.
9:282009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
White MK and Khalili K: Expression of JC
virus regulatory proteins in human cancer: potential mechanisms for
tumourigenesis. Eur J Cancer. 41:2537–2548. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
White MK and Khalili K: Polyomaviruses and
human cancer: molecular mechanisms underlying patterns of
tumorigenesis. Virology. 324:1–16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khalili K, Gordon J and White MK: The
polyomavirus, JCV and its involvement in human disease. Adv Exp Med
Biol. 577:274–287. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Neel JV: The Colonel Harlan D. Sanders
Award Address for 1998: JC virus and its possible role in
oncogenesis. Am J Med Genet. 83:152–156. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gordon J, Del Valle L, Otte J and Khalili
K: Pituitary neoplasia induced by expression of human neurotropic
polyomavirus, JCV, early genome in transgenic mice. Oncogene.
19:4840–4846. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hori R, Murai Y, Tsuneyama K, Abdel-Aziz
HO, Nomoto K, Takahashi H, Cheng CM, Kuchina T, Harman BV and
Takano Y: Detection of JC virus DNA sequences in colorectal cancers
in Japan. Virchow Arch. 447:723–730. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zheng Y, Xia P, Zheng HC, Takahashi H,
Masuda S and Takano Y: The screening of viral risk factors in
tongue and pharyngolaryngeal squamous carcinoma. Anticancer Res.
30:1233–1238. 2010.PubMed/NCBI
|
|
11
|
Del Valle L, White MK, Enam S, Oviedo SP,
Bromer MQ, Thomas RM, Parkman HP and Khalili K: Detection of JC
virus DNA sequences and expression of viral T antigen and
agnoprotein in esophageal carcinoma. Cancer. 103:516–527.
2005.PubMed/NCBI
|
|
12
|
Enam S, Del Valle L, Lara C, Gan DD,
Ortiz-Hidalgo C, Palazzo JP and Khalili K: Association of human
polyomavirus JCV with colon cancer: evidence for interaction of
viral T-antigen and beta-catenin. Cancer Res. 62:7093–7101.
2002.PubMed/NCBI
|
|
13
|
Zambrano A, Kalantari M, Simoneau A,
Jensen JL and Villarreal LP: Detection of human polyomaviruses and
papillomaviruses in prostatic tissue reveals the prostate as a
habitat for multiple viral infections. Prostate. 53:263–276. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kutsuna T, Zheng H, Abdel-Aziz HO, Murai
Y, Tsuneyama K, Furuta I and Takano Y: High JC virus load in tongue
carcinomas may be a risk factor for tongue tumorigenesis. Virchows
Arch. 452:405–410. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Murai Y, Zheng HC, Abdel Aziz HO, Mei H,
Kutsuna T, Nakanishi Y, Tsuneyama K and Takano Y: High JC virus
load in gastric cancer and adjacent non-cancerous mucosa. Cancer
Sci. 98:25–31. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Link A, Shin SK, Nagasaka T, Balaguer F,
Koi M, Jung B, Boland CR and Goel A: JC virus mediates invasion and
migration in colorectal metastasis. PLoS One. 4:e81462009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ricciardiello L, Laghi L, Ramamirtham P,
Chang CL, Chang DK, Randolph AE and Boland CR: JC virus DNA
sequences are frequently present in the human upper and lower
gastrointestinal tract. Gastroenterology. 119:1228–1235. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Niv Y, Goel A and Boland CR: JC virus and
colorectal cancer: a possible trigger in the chromosomal
instability pathways. Curr Opin Gastroenterol. 21:85–89.
2005.PubMed/NCBI
|
|
19
|
Shin SK, Li MS, Fuerst F, Hotchkiss E,
Meyer R, Kim IT, Goel A and Boland CR: Oncogenic T-antigen of JC
virus is present frequently in human gastric cancers. Cancer.
107:481–488. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zheng H, Abdel Aziz HO, Nakanishi Y,
Masuda S, Saito H, Tsuneyama K and Takano Y: Oncogenic role of JC
virus in lung cancer. J Pathol. 212:306–315. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ramamoorthy S, Devaraj B, Miyai K, Luo L,
Liu YT, Boland CR, Goel A and Carethers JM: John Cunningham virus
T-antigen expression in anal carcinoma. Cancer. (Epub ahead of
print) in press. 2010.
|
|
22
|
Wang JP, Zhang MY, Xu XY, Wang W, Xia P,
Zheng ZH, Wang LZ and Takano Y: The establishment of transgenic
mouse model of lens epithelial tumors induced by JC Virus T
Antigen. J Chin Med U. 40:27–29. 2011.
|
|
23
|
Basile A, Darbinian N, Kaminski R, White
MK, Gentilella A, Turco MC and Khalili K: Evidence for modulation
of BAG3 by polyomavirus JC early protein. J Gen Virol.
90:1629–1640. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Reviriego-Mendoza MM and Frisque RJ:
Interaction and co-localization of JC virus large T antigen and the
F-box protein β-transducin-repeat containing protein. Virology.
410:119–128. 2011.PubMed/NCBI
|
|
25
|
Kelley JR and Duggan JM: Gastric cancer
epidemiology and risk factors. J Clin Epidemiol. 56:1–9. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
American Cancer Society. Colorectal cancer
facts and figures. 3rd edition. American Cancer Society; 2009
|
|
27
|
Calgua B, Barardi CR, Bofill-Mas S,
Rodriguez-Manzano J and Girones R: Detection and quantitation of
infectious human adenoviruses and JC polyomaviruses in water by
immunofluorescence assay. J Virol Methods. 171:1–7. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xia P, Xu XY, Jia BP, Wang W, Guan YF,
Takano Y and Zheng HC: The construction and expression confirmation
of JC virus T antigen expression plasmid in gastric mucosa. J Chin
Med U. 39:18–21. 2010.
|
|
29
|
Kumada T, Tsuneyama K, Hatta H, Ishizawa S
and Takano Y: Improved 1-h rapid immunostaining method using
intermittent microwave irradiation: practicability based on 5 years
application in Toyama Medical and Pharmaceutical University
Hospital. Mod Pathol. 17:1141–1149. 2004.
|
|
30
|
Theodoropoulos G, Panoussopoulos D,
Papaconstantinou I, Gazouli M, Perdiki M, Bramis J and Lazaris ACh:
Assessment of JC polyoma virus in colon neoplasms. Dis Colon
Rectum. 48:86–91. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schätzl HM, Sieger E and Jäger G, Nitschko
H, Bader L, Ruckdeschel G and Jäger G: Detection by PCR of human
polyomaviruses BK and JC in immunocompromised individuals and
partial sequencing of control regions. J Med Virol. 42:138–145.
1994.PubMed/NCBI
|
|
32
|
Monaco MC, Jensen PN, Hou J, Durham LC and
Major EO: Detection of JC virus DNA in human tonsil tissue:
evidence for site of initial viral infection. J Virol.
72:9918–9923. 1998.PubMed/NCBI
|
|
33
|
Boldorini R, Veggiani C, Barco D and Monga
G: Kidney and urinary tract polyomavirus infection and
distribution. Molecular biology investigation of 10 consecutive
autopsies. Arch Pathol Lab Med. 129:69–73. 2005.
|
|
34
|
Cozen SD and Cole CN: The three
transforming regions of SV40 T antigens are required for
immortalization of primary mouse embryo fibroblasts. Oncogene.
11:2295–2302. 1995.PubMed/NCBI
|