Introduction
Gliomas are the most common primary tumors of the
central nervous system (CNS) and include a variety of histological
tumor types and World Health Organization (WHO) malignancy grades.
Histologically, astrocytic, oligodendroglial and mixed
oligoastrocytic tumors are the most common gliomas (1). Diffuse astrocytoma (WHO grade II) is
inherently predisposed to recur locally and spontaneously progress
to anaplastic astrocytoma (WHO grade III) and eventually secondary
glioblastoma (WHO grade IV) (2).
Over the last decades, the molecular mechanisms of tumorigenesis
and malignant transformation have been investigated and revealed to
be associated with genetic mutations. However, a limited number of
molecular alterations have attracted much attention, mainly
including TP53 mutations, loss of heterzygosity (LOH) on 17p,
isocitrate dehydrogenase 1 gene (IDH1) or IDH2 gene mutations and
deletions on chromosomes 6, 9p, 11p, 19q and 22q (3). Therefore, it is crucial to identify
novel biomarkers that are involved in the malignant transformation
in glioma.
microRNAs (miRNAs) are a class of non-coding short
RNAs from 17 to 25 nucleotides in length that play significant
roles in post-transcriptional gene regulation (4). The dysregulation of miRNAs has been
linked to a series of diseases, including various types of cancer.
They take part in complex regulatory networks that influence almost
every cellular process and may be involved in oncogenesis (5). The role of miRNAs in the tumorigenesis
of glioma is of great interest. It has been demonstrated that
miRNAs act as oncogenes or tumor suppressor genes and have
modulatory effects on tumor cell proliferation, differentiation,
metastasis and invasion (6). The
current study explored the role of a single miRNA, miR-544, in the
malignant progression of human gliomas by comparing expression
levels in primary low-grade gliomas and secondary high-grade
gliomas. The expression of miR-544 in tissue and serum samples of
glioma patients was analyzed to determine whether it may be used as
a biomarker of malignant transformation in glioma.
Patients and methods
Patients and tissue samples
The study was approved by the Ethics Committee of
Beijing Tiantan Hospital, Capital Medical University (Beijing,
China), and written informed consent was obtained from all
patients. In this study, a total of 198 patients with glioma (63
low-grade glioma, 44 anaplastic astrocytoma and 91 glioblastoma
tumors) were included in the microarray analysis to identify the
expression of miR-544 in tumor samples. Quantitative real-time
reverse transcription PCR (qRT-PCR) was used to validate the
expression levels of miR-544 in paired tissue and serum samples in
an independent validated cohort (25 low-grade glioma, 21 anaplastic
astrocytoma and 20 glioblastoma tumors).
Tissue samples were obtained from patients during
surgery at the Beijing Tiantan Hospital between January 2006 and
December 2009. Histological diagnoses were made according to the
2007 WHO classification. Samples were snap-frozen immediately
following surgery and stored at −80°C. Only specimens with a
histologically estimated tumor cell content of ≥80% were used for
the analyses.
Serum sample collection
Whole blood samples of glioma patients were
collected. The samples were allowed to stand at room temperature
for 30 min and then centrifuged at 2,500 x g for 15 min at 4°C. To
remove cellular contaminants, serum samples were subjected to
additional centrifugation at 15,000 x g for 10 min. The
supernatants to be used for RNA extraction were snap-frozen and
then stored at −80°C.
RNA extraction
Total RNA was extracted from tissues using an
mirVana PARIS kit (Ambion, Carlsbad, CA, USA) according to the
manufacturer’s instructions. The concentrations of all RNA samples
were quantified by NanoDrop 1000 (Nanodrop, Wilmington, Delaware,
USA).
RNA extraction from serum was performed using the
same kit mentioned previously. In initial experiments, a poor
correlation was observed between the RNA concentration, as measured
with a NanoDrop spectrophotometer, and the ability to amplify
miRNA. To overcome this problem, the RNA volume used in the
experiment was unified instead of the RNA concentration, as
described by McDonald et al (7). The volume of serum that was extracted
and the volume of extracted eluent used for qRT-PCR were consistent
throughout the study.
miRNA microarray
The miR-544 expression values of 198 glioma tissues
were obtained from Chinese Glioma Genome Atlas (CGGA), a database
that focuses on glioma. Briefly, 200 ng total RNA was
polyadenylated and then reverse transcribed into cDNA using a
biotin-labeled Oligo dT primer with a universal PCR sequence.
Following cDNA synthesis, miRNAs were individually interrogated
using specific oligonucleotides. A single miRNA-specific Oligo
(MSO) was designed against each mature miRNA sequence and
miRNA-specific primers were extended using DNA polymerase.
Universal primers were used to amplify the cDNA templates and the
primer complimentary to the array was fluorescently labeled.
Finally, the labeled, single-stranded PCR products were hybridized
to the Human v2.0 miRNA Expression BeadChip (Illumina, Inc., San
Diego, CA, USA) with 1,146 human miRNAs (97% coverage of the
miRBase 12.0 database).
Reverse transcription and qPCR
analyses
The TaqMan MicroRNA Reverse Transcription kit
(Applied Biosystems, Carlsbad, CA, USA) and miRNA-specific
stem-loop primers (Applied Biosystems) were used for miRNA reverse
transcription. qPCR was performed using TaqMan Gene Expression
Master Mix (Applied Biosystems) and individual miRNA primers and
hydrolysis probes (Applied Biosystems) according to the
manufacturer’s instructions. Relative quantification of miRNA
expression was calculated using the 2−ΔΔCT method. RNU6B
was used as endogenous control for tissue detection. For serum
qRT-PCR analysis, there was no consensus on the use of housekeeping
miRNA. Therefore, a healthy donor sample was used as control and
processed together with the testing samples as described previously
(8).
Statistical analysis
Comparisons of miR-544 levels among patients with
various tumor grades were performed using Student’s t-tests. The
Pearson correlation analysis was used to examine the correlation
between miR-544 levels of paired tissue and serum samples. All
statistical analyses were performed using GraphPad Prism 5. A
two-sided P<0.05 was considered to indicate a statistically
significant result.
Results
miR-544 expression is decreased in
anaplastic glioma and glioblastoma tissue samples
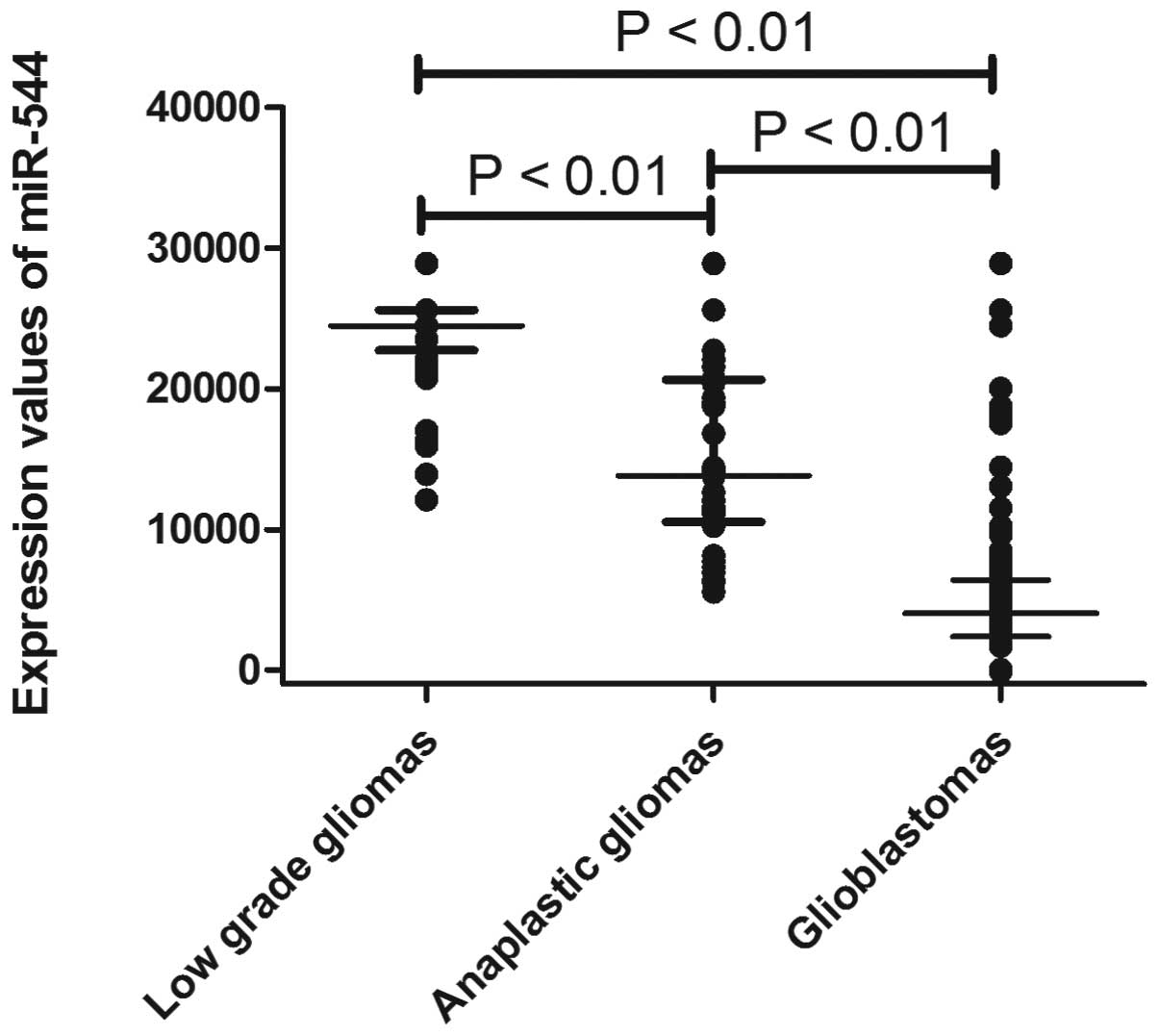
The expression of miR-544 was measured in a series
of 198 glioma samples via microarray. As shown in Fig. 1, miR-544 expression decreased
significantly in anaplastic gliomas (P<0.01) and glioblastomas
(P<0.01) compared with low-grade gliomas. To further confirm
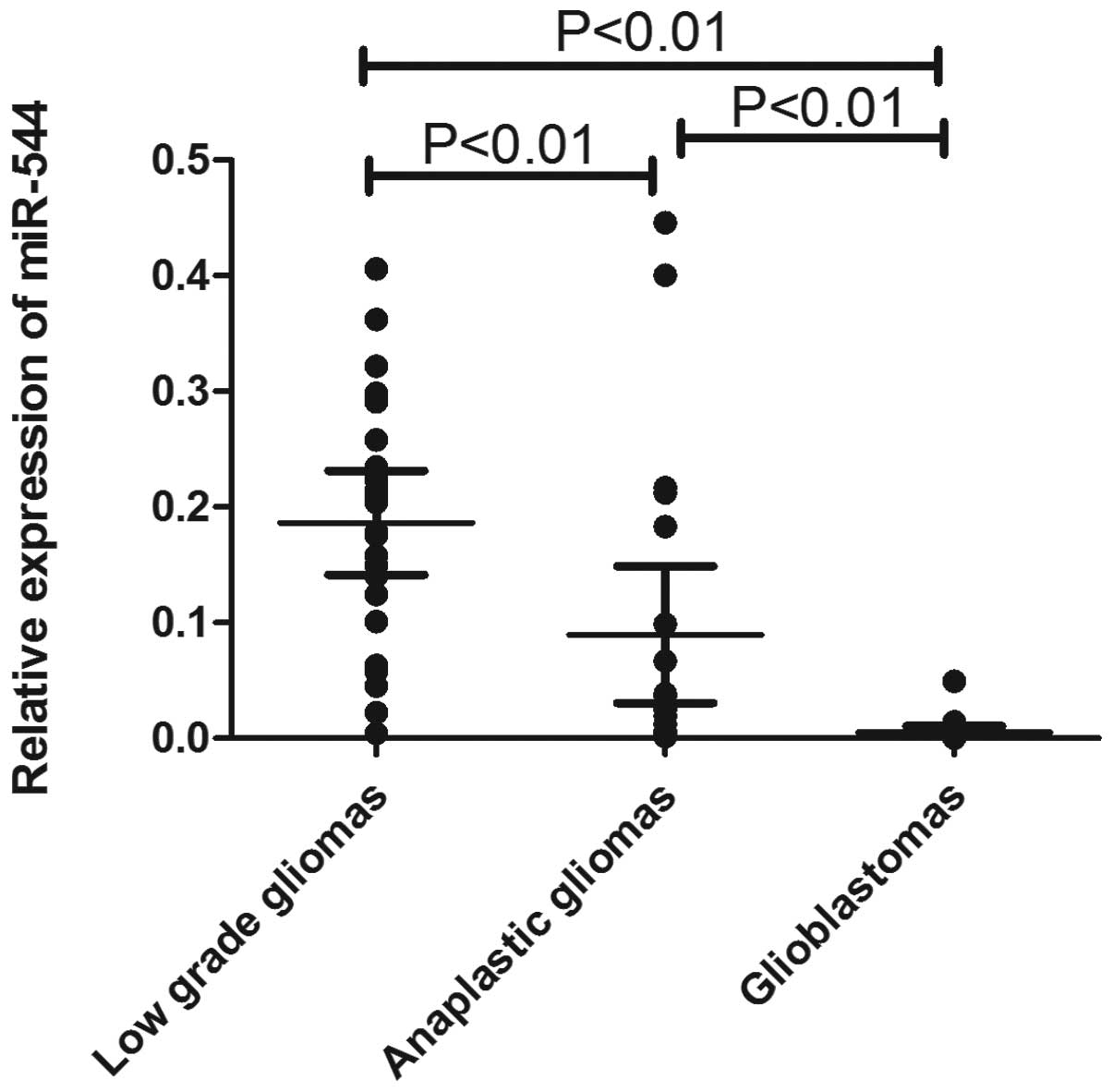
this result, qRT-PCR was performed to examine miR-544 expression
levels in an independent cohort containing 66 tumor samples (25
low-grade glioma, 21 anaplastic astrocytoma and 20 glioblastoma
tumors). As shown in Fig. 2,
miR-544 was significantly downregulated in anaplastic gliomas
(P<0.01) and glioblastomas (P<0.01) compared with low-grade
gliomas.
miR-544 expression is not significantly
different in glioma, anaplastic astrocytoma or glioblastoma serum
samples
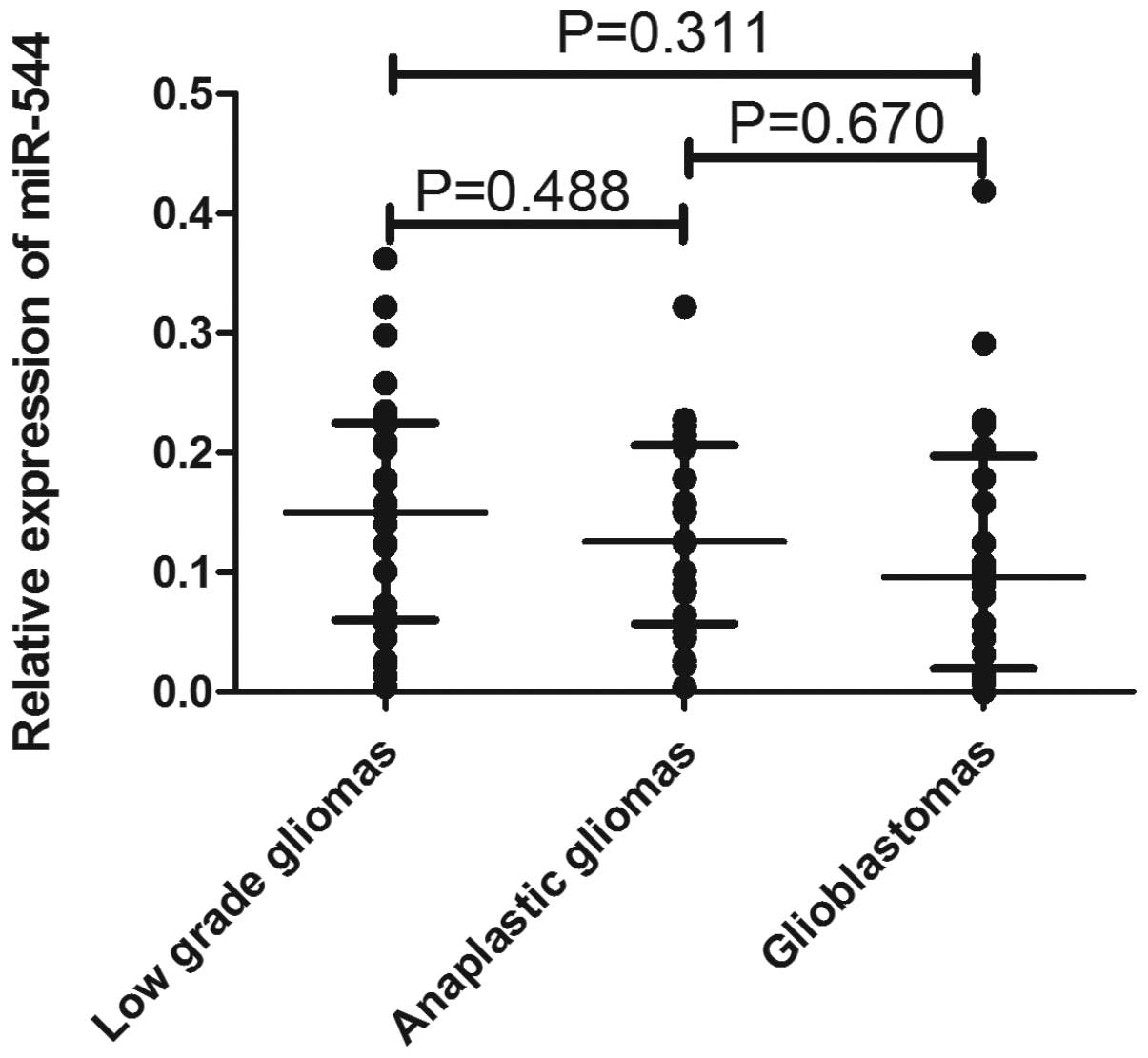
To explore whether miR-544 may be a soluble
biomarker of malignant transformation, the expression of miR-544
was detected in the validation cohort containing 66 serum samples
(25 low-grade glioma, 21 anaplastic astrocytoma and 20 glioblastoma
tumors), which were paired with tissue samples, by qRT-PCR assay.
As shown in Fig. 3, there was no
significant difference in miR-544 expression among the 3 groups,
although the levels tended to be lower in anaplastic and
glioblastoma patients.
miR-544 expression in tissue and serum
samples has a weakly positive correlation
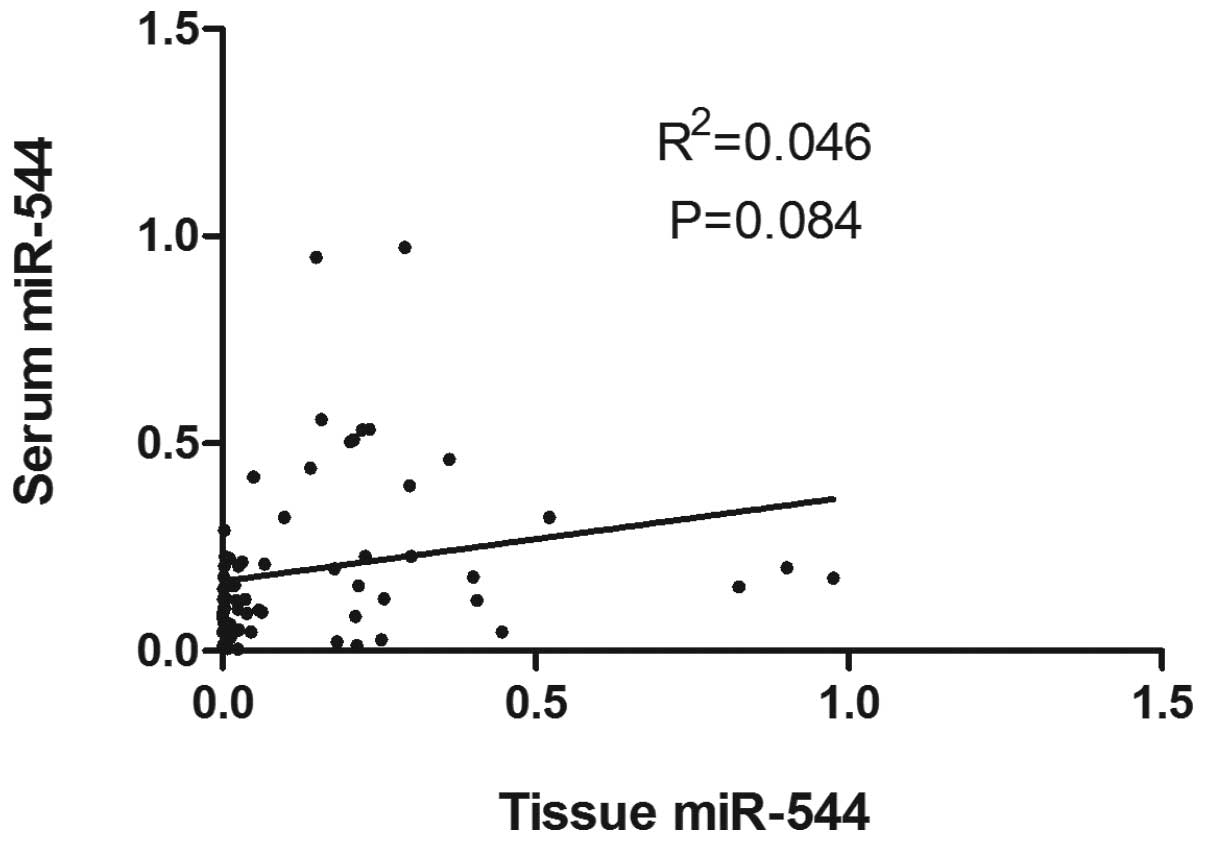
To further explore the correlation between miR-544
expression in paired tissue and serum samples, a Pearson
correlation analysis of miR-544 levels between 66 tissue and serum
samples of glioma patients was performed. As shown in Fig. 4, there was a weakly positive
correlation between tissue and serum levels of miR-544
(R2=0.046, P=0.084).
Discussion
Low-grade gliomas are well differentiated and grow
slowly but show a consistent tendency to diffusely infiltrate the
surrounding brain tissue. They almost invariably recur and this is
often associated with the progression to a higher malignancy
(9). The median survival of
low-grade glioma patients is between 5 and 10 years (10). However, the median survival time of
patients with anaplastic astrocytomas is less than 3 years, while
for glioblastoma this is less than one year (11). In addition, resistance to
chemotherapeutic drugs and therapeutic irradiation often occurs in
the malignant transformation of gliomas (12). Thus, novel biomarkers for the
detection of malignant transformation are required to form
predictions and select the best possible treatment regimen for
glioma patients. Over the last few decades, several genes
associated with the malignant progression of gliomas have been
identified. The TP53 mutation appears to be a relatively early
event during the development of an astrocytoma, whereas the loss or
mutation of PTEN and amplification of EGFR are characteristic of
higher grade tumors (13–15). The loss of 9p21 is confined to WHO
grade III tumors, whereas mutations at codon 132 of IDH1 may occur
following the formation of a low-grade glioma and drive the
progression of the tumor to a glioblastoma (16,17).
The current study has identified that a non-coding RNA, miR-544,
exhibited a progression-associated downregulation in glioma tumors,
which may be used as a novel biomarker in malignant
transformation.
Previous studies have indicated that a variety of
miRNAs play significant roles in glioma. miR-21 was the first miRNA
to be linked with glioma malignancy and acted as an oncogene
(6). The expression levels of
miR-21 were elevated in human glioma cells and tissues and also
correlated with tumor grade (18).
miR-181a and miR-181b were observed as tumor suppressors that
inhibit growth and induce the apoptosis of glioma cells (19). The expression of miR-221 and 222
were upregulated in human glioblastoma and they induced an increase
in cell migration and growth by targeting the protein phosphate
PTPμ, whereas let-7 reduced the in vitro
proliferation and migration of glioma cells (20,21).
Using genome-wide miRNA profiling, a previous study
identified a 23-miRNA expression signature that may distinguish
glioblastoma from anaplastic astrocytoma with 95% accuracy
(22). In the current study, the
expression of miR-544 was measured using a microarray in a series
of glioma tissues with various tumor grades. It was revealed that
miR-544 expression decreased significantly in anaplastic gliomas or
glioblastoma compared with low-grade gliomas and the further
qRT-PCR assay confirmed the results. A previous study also reported
that miR-544 was significantly downregulated in osteosarcoma and
may affect its pathogenesis by targeting the cMYC transcript
(23). This is in accordance with
the finding that the dysregulation of miR-544 may be involved in
the process of tumorigenesis and malignant progression.
Circulating miRNAs are significant tumor markers due
to their invasiveness and ease of detection. To further explore
whether circulating miR-544 may be a non-invasive biomarker for
malignant transformation of gliomas, its expression levels were
measured in serum samples. The results revealed that the levels
tended to be lower in anaplastic and glioblastoma patients compared
with low-grade gliomas, but with no significant difference. There
was a weakly positive correlation between the tissue and serum
levels of miR-544. The majority of previous studies have
demonstrated the same trend of alteration between circulating
miRNAs and tissue miRNAs, either an increase or decrease, in
various types of cancer (24–26).
However, a different correlation between circulating miRNAs and
tissue miRNAs has also been observed. Wulfken et al observed
109 miRNAs at higher levels in the serum of patients with renal
cell carcinoma, but only 36 miRNAs were upregulated in tissue
samples (27). Another study
reported that approximately 66% of the released miRNAs reflect the
cellular miRNA content of malignant mammary epithelial cells
(28). These studies provide
potential evidence for the hypothesis that circulating miRNAs
reflect various aspects of human physiological status and only a
subset of circulating miRNAs have a tumor-specific origin (27,29).
Conversely, the field of detecting circulating miRNAs for use in
cancer diagnosis is new and thus the lack of standard protocol may
also contribute to the conflicting results. Therefore, certain
issues remain to be resolved, including type of samples, use of
appropriate normalization controls, standard protocols for sample
preparation and concentration measurement (7). Taken together, it is difficult for
circulating miR-544 to be a stable biomarker of the malignant
transformation of glioma.
In summary, miR-544 expression decreased
significantly in anaplastic gliomas or secondary glioblastomas
compared with low-grade gliomas. In an independent cohort of glioma
patients in validation studies, miR-544 exhibited a
progression-associated downregulation in glioma tumors. These data
support a significant role for miR-544 aberration in the malignant
transformation of glioma and thus the downregulation of miR-544 in
tissue may be used as a novel biomarker. Further studies are
required to characterize in detail the molecular mechanisms of
miR-544 action.
References
|
1.
|
O MartinhoS GranjaT
JaraquemadaDownregulation of RKIP is associated with poor outcome
and malignant progression in gliomasPLoS
One7e30769201210.1371/journal.pone.003076922292035
|
|
2.
|
DN LouisH OhgakiOD WiestlerThe 2007 WHO
classification of tumours of the central nervous systemActa
Neuropathol11497109200710.1007/s00401-007-0243-417618441
|
|
3.
|
B MalzkornM WolterF
LiesenbergIdentification and functional characterization of
microRNAs involved in the malignant progression of gliomasBrain
Pathol20539550201010.1111/j.1750-3639.2009.00328.x19775293
|
|
4.
|
L CastellanoG GiamasJ JacobThe estrogen
receptor-alpha-induced microRNA signature regulates itself and its
transcriptional responseProc Natl Acad Sci
USA1061573215737200910.1073/pnas.0906947106
|
|
5.
|
W FilipowiczSN BhattacharyyaN
SonenbergMechanisms of post-transcriptional regulation by
microRNAs: are the answers in sight?Nat Rev
Genet9102114200810.1038/nrg229018197166
|
|
6.
|
Y ZhangA DuttaR AbounaderThe role of
microRNAs in glioma initiation and progressionFront
Biosci17700712201210.2741/395222201769
|
|
7.
|
JS McDonaldD MilosevicHV ReddiSK GrebeA
Algeciras-SchimnichAnalysis of circulating microRNA: preanalytical
and analytical challengesClin
Chem57833840201110.1373/clinchem.2010.15719821487102
|
|
8.
|
Z HuX ChenY ZhaoSerum microRNA signatures
identified in a genome-wide serum microRNA expression profiling
predict survival of non-small-cell lung cancerJ Clin
Oncol2817211726201010.1200/JCO.2009.24.934220194856
|
|
9.
|
K WatanabeK SatoW BiernatIncidence and
timing of p53 mutations during astrocytoma progression in patients
with multiple biopsiesClin Cancer Res352353019979815715
|
|
10.
|
TY JungS JungJH MoonIY KimKS MoonWY
JangEarly prognostic factors related to progression and malignant
transformation of low-grade gliomasClin Neurol
Neurosurg113752757201110.1016/j.clineuro.2011.08.00221889256
|
|
11.
|
R StuppM ReniG GattaE MazzaC
VechtAnaplastic astrocytoma in adultsCrit Rev Oncol
Hematol637280200710.1016/j.critrevonc.2007.03.003
|
|
12.
|
M WellerU MalipieroA AguzziJC ReedA
FontanaProtooncogene bcl-2 gene transfer abrogates Fas/APO-1
antibody-mediated apoptosis of human malignant glioma cells and
confers resistance to chemotherapeutic drugs and therapeutic
irradiationJ Clin Invest9526332643199510.1172/JCI117965
|
|
13.
|
H OhgakiP KleihuesGenetic pathways to
primary and secondary glioblastomaAm J
Pathol17014451453200710.2353/ajpath.2007.07001117456751
|
|
14.
|
FB FurnariT FentonRM BachooMalignant
astrocytic glioma: genetics, biology, and paths to treatmentGenes
Dev2126832710200710.1101/gad.159670717974913
|
|
15.
|
RG WeberM SabelJ
ReifenbergerCharacterization of genomic alterations associated with
glioma progression by comparative genomic
hybridizationOncogene1398399419968806688
|
|
16.
|
SH BignerMR MatthewsBK RasheedMolecular
genetic aspects of oligodendrogliomas including analysis by
comparative genomic hybridizationAm J
Pathol155375386199910.1016/S0002-9440(10)65134-6
|
|
17.
|
DW ParsonsS JonesX ZhangAn integrated
genomic analysis of human glioblastoma
multiformeScience32118071812200810.1126/science.116438218772396
|
|
18.
|
A ContiM AguennouzD La TorremiR-21 and 221
upregulation and miR-181b downregulation in human grade II–IV
astrocytic tumorsJ Neurooncol93325332200919159078
|
|
19.
|
L ShiZ ChengJ Zhanghsa-mir-181a and
hsa-mir-181b function as tumor suppressors in human glioma
cellsBrain
Res1236185193200810.1016/j.brainres.2008.07.08518710654
|
|
20.
|
C QuintavalleM GarofaloC ZancamiR-221/222
over-expession in human glioblastoma increases invasiveness by
targeting the protein phosphate
PTPmuOncogene31858868201210.1038/onc.2011.28021743492
|
|
21.
|
ST LeeK ChuHJ OhLet-7 microRNA inhibits
the proliferation of human glioblastoma cellsJ
Neurooncol1021924201110.1007/s11060-010-0286-620607356
|
|
22.
|
SA RaoV SantoshK SomasundaramGenome-wide
expression profiling identifies deregulated miRNAs in malignant
astrocytomaMod
Pathol2314041417201010.1038/modpathol.2010.13520711171
|
|
23.
|
V ThayanithyAL SarverRV KarthaPerturbation
of 14q32 miRNAs-cMYC gene network in
osteosarcomaBone50171181201210.1016/j.bone.2011.10.01222037351
|
|
24.
|
JC BraseM JohannesT SchlommCirculating
miRNAs are correlated with tumor progression in prostate cancerInt
J Cancer128608616201110.1002/ijc.2537620473869
|
|
25.
|
EK NgWW ChongH JinDifferential expression
of microRNAs in plasma of patients with colorectal cancer: a
potential marker for colorectal cancer
screeningGut5813751381200910.1136/gut.2008.16781719201770
|
|
26.
|
CC YangPS HungPW WangmiR-181 as a putative
biomarker for lymph-node metastasis of oral squamous cell
carcinomaJ Oral Pathol
Med40397404201110.1111/j.1600-0714.2010.01003.x21244495
|
|
27.
|
LM WulfkenR MoritzC OhlmannMicroRNAs in
renal cell carcinoma: diagnostic implications of serum miR-1233
levelsPLoS One6e25787201110.1371/journal.pone.002578721984948
|
|
28.
|
L PigatiSC YaddanapudiR IyengarSelective
release of microRNA species from normal and malignant mammary
epithelial cellsPLoS
One5e13515201010.1371/journal.pone.001351520976003
|
|
29.
|
K ZenCY ZhangCirculating MicroRNAs: a
novel class of biomarkers to diagnose and monitor human cancersMed
Res Rev32326348201210.1002/med.2021522383180
|