Introduction
Post-operative multi-organ dysfunction (MODF) and
systemic inflammatory reaction syndrome (SIRS) remain the main
complications of liver transplantation and are critical problems
that contribute to a high mortality rate (1,2).
Bacterial translocation and enterogenous endotoxemia following
intestinal injury may play a significant role in MODF and SIRS
(3). Zheyu and Lunan (4) demonstrated that the motility and
barriers of the intestine were destroyed following liver
transplantation in rats, as evidenced by the increase in plasma
endotoxin levels.
Liver transplantation requires the portal vein to be
blocked, causing intestinal congestion, hypoxia in the anhepatic
phase and the restoration of blood flow reperfusion, making
intestinal ischemia-reperfusion injury inevitable (4,5).
Oxidative damage induced by reactive oxygen species (ROS),
including the superoxide anion (O2−),
hydrogen peroxide (H2O2) and the hydroxyl
radical (•OH), play a crucial role in the pathogenesis
of ischemia-reperfusion injury (6,7). Due
to the existence of antioxidative enzymes, including superoxide
dismutase (SOD) and catalase (CAT), and antioxidants, including
glutathione (GSH), the redox balance is well maintained and the
clearance of ROS is promoted (8–10).
Several studies have shown that inflammatory mediators, including
tumor necrosis factor-α (TNF-α) and interleukin-1 (IL-1), are
involved in the intestinal injury that is induced by liver
transplantation (4,5). However, the exact alternative pattern
of the oxidant/antioxidant system in the process of intestinal
injury following liver transplantation remains obscure.
Nuclear factor NF-E2-related factor-2 (Nrf2) has
been demonstrated to regulate the synthesis of antioxidant enzymes
and to maintain the antioxidative capacity of the body (11,12). A
study using microarrays revealed that Nrf2 regulated the majority
of antioxidative enzyme gene expression in the lung (13). Numerous studies have reported that
Nrf2 plays a pivotal role in preventing oxidative damage. However,
the alterations to the expression Nrf2 remains unclear during an
intestinal injury following liver transplantation.
The present study was designed to investigate the
dynamic changes in the intestinal ROS levels, Nrf2 expression and
antioxidant enzymes activity (SOD, CAT and GSH) occurring 24 h
after orthotopic liver transplantation (OLAT), and to evaluate the
imbalance of oxidants and antioxidants.
Materials and methods
Animals and experimental groups
This study was approved by the Institutional Animal
Care and Use Committee of Sun Yat-Sen University (Guangzhou, China)
and followed the National Guidelines for the treatment of animals.
A total of 35 male rats weighing 220–280 g were randomly assigned
into five groups consisting of one sham surgery group (sham; n=7)
and four treatment groups that were tested at 4 (n=7), 8 (n=7), 16
(n=7) and 24 h (n=7) following OLAT, respectively. Subsequent to
being anesthetized, the rats belonging to the sham group were
subjected to a laparotomy and vascular dissection, without hepatic
vascular exclusion and perfusion. The rats in the other groups
underwent OLAT. The intestines, starting from 5 cm to the terminal
ileum, were removed at 4, 8, 16 and 24 h following OLAT.
Experimental model of rat OLAT
An OLAT model similar to that previously described
by Jin et al(14) was
applied with certain modifications (15). The rats were fasted for 12 h and
anesthetized using inhaled ether, which was administered through an
open face guard. Following the anesthesia, the abdomen was incised,
the falciform ligament of the liver was resected and the blood
vessels along the esophagus were ligated and disconnected. The
suprahepatic vena cava (SVC) was fully liberated and the liver was
replaced in its original position. Once the upper region of the
left renal vein was completely free, the inferior vena cava (IVC)
was dissociated. The first hepatic portal was drawn and the portal
vein (PV) was separated from the splenic veins and the convergence
of the inferior mesenteric vein. The hepatic artery and the biliary
tract were liberated. Subsequently, the portal hepatics were
ligated. Microvascular clamps were folded on the convergence of the
inferior mesenteric vein, hepatic artery, splenic vein, SVC and
IVC. The PV was punctured with a 4th needle in preparation for
reperfusion and fixed using a microvascular clamp. Ringer lactate
solution (pre-cooled, 4°C) was injected during reperfusion at a
speed of 2.5 ml/min and a 1-mm incision was cut in the wall of the
IVC as an outflow tract. The liver color progressively turned
yellow if the reperfusion was successful. The recovery of the liver
perfusion represented the accomplishment of OLAT. Finally, the
needle was extracted and the incision of the PV and IVC was
repaired using 8-0 sutures. The clamps on the PV, SVC, IVC and
hepatic artery were loosened. The whole anhepatic phase lasted for
20±1 min.
Histopathological examination
An intestinal segment of 0.5–1.0 cm in length was
cut from 5 cm to the terminal ileum and fixed in 10% formaldehyde.
Subsequent to being embedded in paraffin, the tissues were stained
with hematoxylin and eosin (HE) for light microscopy examination.
Intestinal mucosal damage was evaluated and graded according to the
following criteria suggested by Chiu et al(16): Grade 0, normal mucosa villi; grade
1, development of subepithelial Gruenhagen’s space at the tip of
the villi; grade 2, extension of the subepithelial space with
moderate epithelial lifting; grade 3, massive epithelial lifting,
possibly with a few denuded villi; grade 4, denuded villi with
lamina propria and exposed capillaries; and grade 5, disintegration
of the lamina propria, ulceration and hemorrhage.
Preparation of specimens and detection of
protein concentration in the small intestine
A segment of small intestine was washed with frozen
saline, dried with suction paper and frozen at −80ºC. Intestinal
protein levels were measured using a bicinchoninic acid assay
(BCA), according to the manufacturer’s instructions (Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China).
Detection of malondialdehyde (MDA)
content
MDA content was measured using the thiobarbituric
acid (TBA) method (Nanjing KeyGen Biotech Co., Ltd.). The
homogenate (0.1 ml) was used to detect the MDA content. The
condensation of MDA and TBA resulted in a red product, which had a
maximum absorption peak at 532 nm. The MDA content was calculated
by measuring the absorbance at 532 nm and expressed as nmol/mg
protein (nmol/mg prot).
Detection of H2O2
and •OH concentrations
In the acidic environment,
H2O2 is able to oxidize Fe2+ ions
to Fe3+ ions, which combine with dye molecules to
product Fe3+ dye complexes. This has a maximum
absorption wavelength that was proportional to the concentration of
H2O2 at 560 nm. The concentrations of
H2O2 were measured by detecting the
absorbance at 560 nm and were expressed as mmol/mgprot.
The levels of •OH were determined through
the Fenton reaction by detecting the absorbance at 550 nm. The
•OH level in the intestinal tissue was expressed as U/mg
prot.
Detection of SOD and CAT activities and
GSH content
The intestinal tissues were homogenized with normal
saline solution and centrifuged at 1,500 × g for 15 min to remove
the debris. The supernatant was transferred into fresh tubes for
the evaluation of the SOD and CAT activities and GSH content. The
measurements were performed using an assay kit (KeyGen Biotech Co.,
Ltd., Nanjing, China) following the manufacturer’s
instructions.
SOD activity was determined using the Xanthine
oxidase method by detecting the absorbance at 550 nm. SOD activity
was expressed as U/mg prot.
CAT was measured by the reaction of CAT scavenging
H2O2. In this reaction, ammonium molybdate
was added and a pale yellow complex was produced. The change in the
absorbance at 405 nm was monitored. CAT activity was expressed as
U/mg prot.
GSH reacted with dithiobis-nitrobenzoic acid to
generate 5-dithio-bis2-nitrobenzoic acid dithiobis-nitrobenzoic
acid anion, which was a yellow compound and had a maximum
absorption peak at 420 nm. The concentrations of GSH were measured
by detecting the absorbance at 420 nm and were expressed as
mg/gprot.
Western blot analysis
Nuclear proteins were extracted from the frozen
intestinal tissues using protein extraction kits (KeyGen Biotech
Co., Ltd.) for Nrf2 measurements. The protein concentration was
measured using the Bradford method. Protein (60 μg) was loaded onto
a 4–20% SDS-PAGE pre-made gel (Invitrogen, Carlsbad, CA, USA) for
polyacrylamide gel electrophoresis and then transferred to a
polyvinylidene fluoride membrane that was pretreated with 100%
methanol. The membranes that were loaded with the protein of
interest, Nrf2, were incubated with 5% skimmed milk. Rabbit
monoclonal anti-Nrf2 antibody (dilution, 1:200; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) was added to the
supernatant and the mixture was incubated on a rotating wheel at
4°C overnight. On the second day, the membranes were washed with
TBS three times and incubated with a second antibody conjugated to
horseradish peroxidase (dilution, 1:2,000; Santa Cruz
Biotechnology, Inc.) for 1 h at room temperature. Densitometry was
used, and normalization to β-actin immunoreactivity was adopted to
correct the differences in the samples.
Statistical analysis
All the data are expressed as the mean ± SD and were
analyzed using SPSS 16.0 software (SPSS, Inc., Chicago, IL, USA).
Repeated measurements were applied for the intra-group comparison.
P<0.05 was considered to indicate a statistically significant
difference.
Results
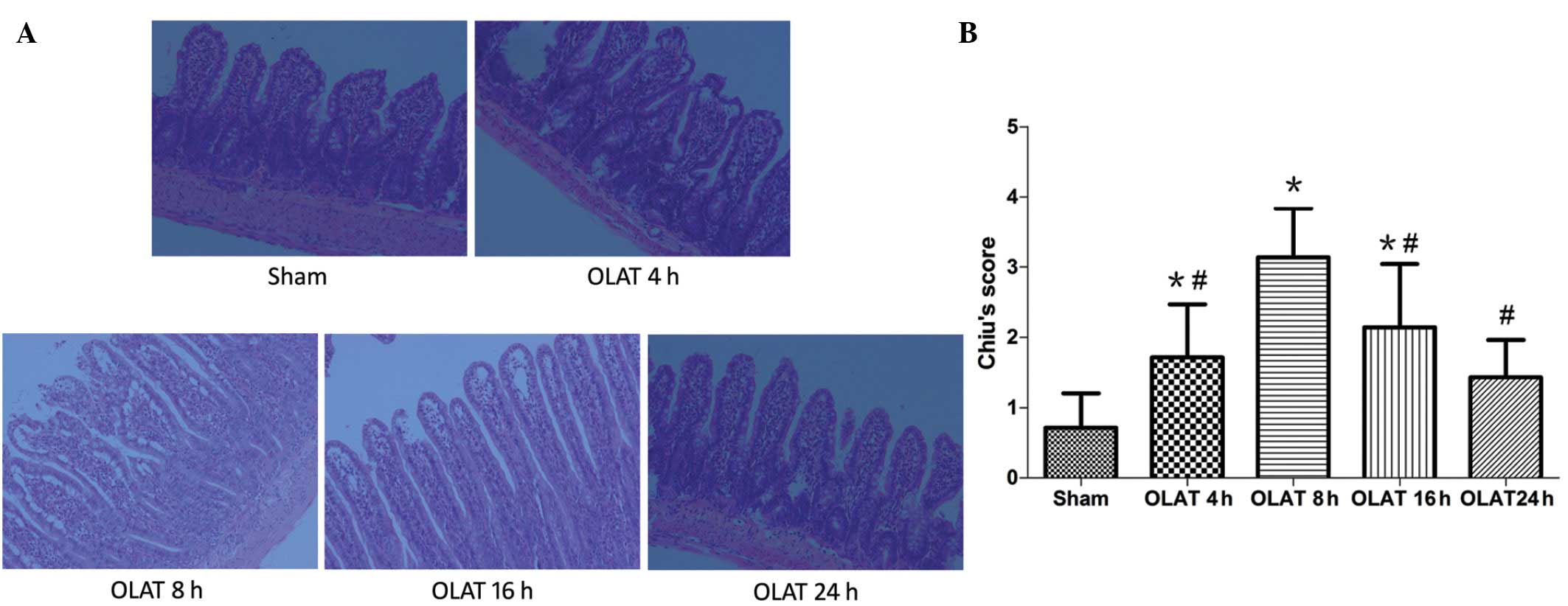
Intestinal pathology under light
microscopy
No injury was evident in the sham group, which
demonstrated normal mucosal villi and glands. However, in the
OLAT-induced intestinal injury groups (Fig. 1), the intestinal structure was
damaged most severely at 8 h following OLAT, where almost all the
animals demonstrated massive epithelial lifting down the sides of
the villi, with a few denuded villi. The intestinal structure was
less damaged at 4 and 16 h following OLAT compared with the animals
with a few denuded villi, accompanied with an extension of the
sub-epithelial space with moderate or massive epithelial lifting.
The Chiu’s scores significantly increased at 4, 8 and 16 h
following OLAT compared with the sham group (P<0.05 vs. sham
group). The Chiu’s scores peaked at 8 h (P<0.05 vs. the sham,
4-, 16- and 24-h groups) and then improved gradually (Fig. 1).
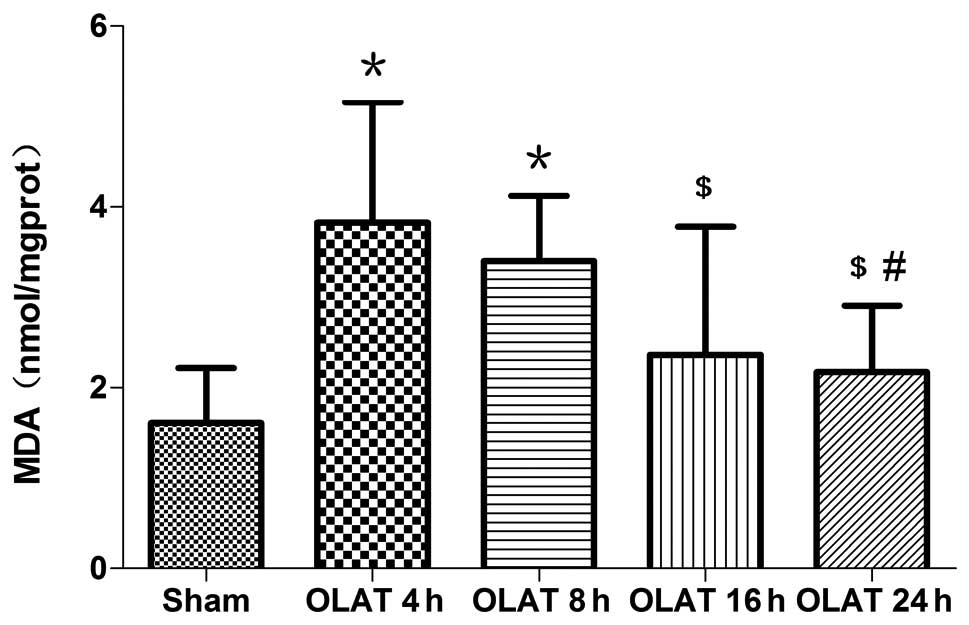
MDA content in the small intestine
The content of MDA increased significantly at 4 and
8 h following OLAT; the content was increased up to 2–2.4 times
(P<0.05 vs. sham group). The MDA content in the 16- and 24-h
groups gradually decreased (P<0.05 vs. 4-h group; Fig. 2).
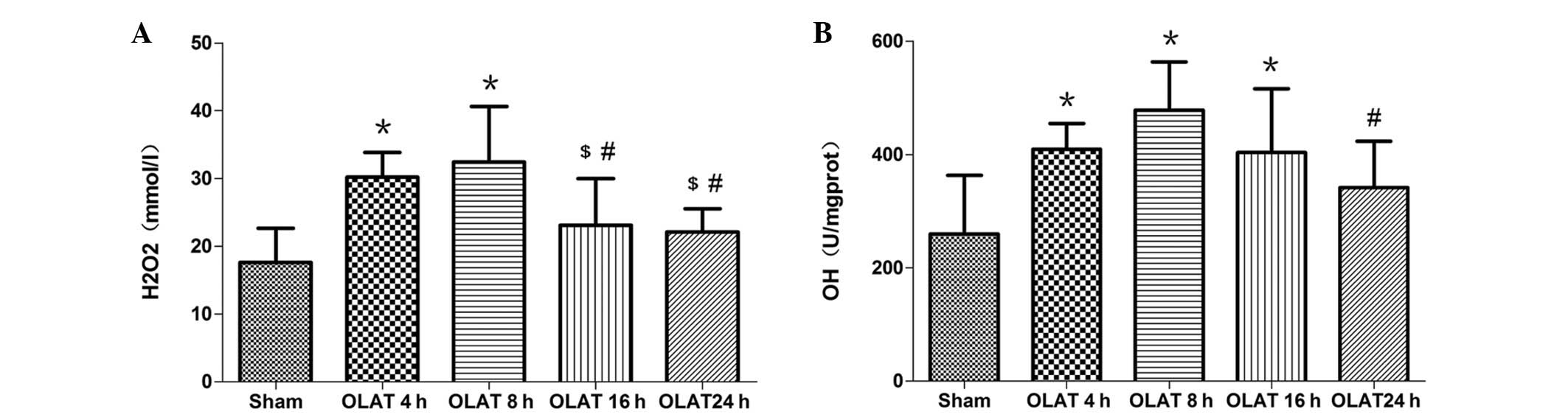
H2O2 content and
•OH level in the small intestine
The content of H2O2 increased
by 1.7–1.8 times at 4 and 8 h following OLAT (P<0.05 vs. sham
group; Fig. 3). The content of
H2O2 gradually decreased at 16-and 24-h
(P<0.05 vs. 4- and 8-h groups). Compared with the sham group,
the levels of •OH increased at 4, 8 and 16 h following
OLAT. The increase ranged between 1.6–1.8 times (P<0.05 vs. sham
group). The levels of •OH at 24 h were lower than those
at 8 h (P<0.05; Fig. 3).
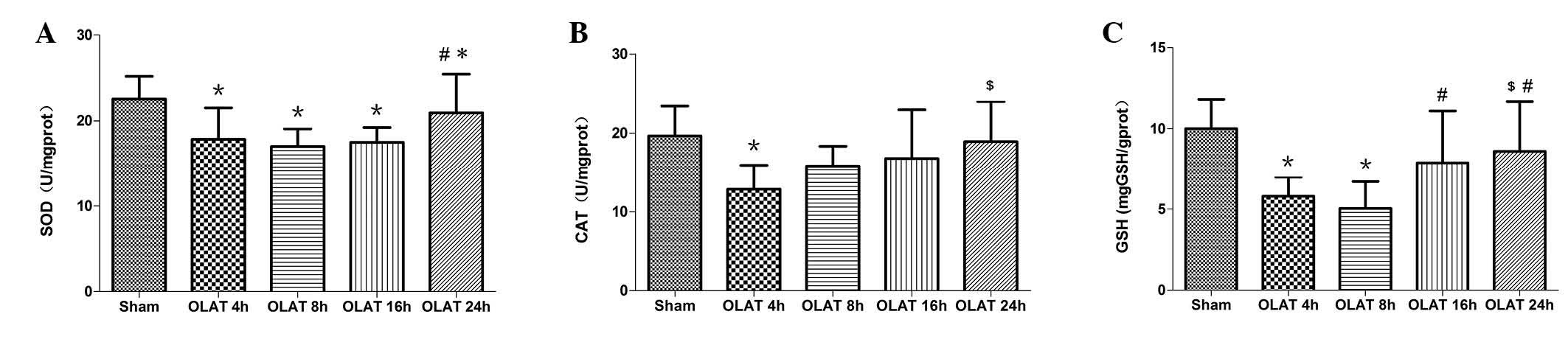
SOD and CAT activity and GSH content in
the small intestine
The SOD activity at 4, 8 and 16 h markedly decreased
by 20–25% (P<0.05 vs. sham group). The SOD activity at 24 h
gradually recovered to the sham group levels (Fig. 4).
The CAT activity greatly decreased by ~40% at 4 h
following OLAT (Fig. 4; P<0.05
vs. sham group). The activity of CAT at 8, 16 and 24 h
progressively recuperated the baseline levels (P>0.05 vs. sham
group).
The content of GSH decreased at 4 and 8 h. The
decrease was 40–50% (P<0.05 vs. sham group). The content of GSH
at 16 and 24 h was higher than the content at 4 and 8 h (P<0.05;
Fig. 4).
Expression of Nrf2 in the small
intestine
OLAT induced a ~50% higher expression of Nrf2
(Fig. 5). Differences were observed
at 4, 16 and 24 h following OLAT (P<0.05 vs. sham group).
Discussion
Bacterial translocation and enterogenous endotoxemia
following intestinal injury have been known to play a significant
role in MODF and SIRS (3).
Oxidative damage that is induced by ROS has a crucial role in the
pathogenesis of intestinal injury. However the correlation between
perioperative intestinal injury and oxidative stress in liver
transplantation remains unclear. The present study used the OLAT
model, which is able to simulate the key surgical procedures and
pathophysiological processes that occur in liver transplantation,
including hemodynamic changes, congestion, hypoxia, intestinal
reperfusion subsequent to unclamping the portal vein and hepatic
ischemia-reperfusion injury (17).
The present study identified that OLAT causes serious injury to the
pathological structure in the intestines at 8 h following the
procedure. The tissues recover gradually, accompanied with an
imbalance in oxidation and antioxidation. The characteristics of
the intestinal damage within 24 h following OLAT were aggravated
progressively and then recovered gradually.
Oxidative stress has been known to play a crucial
role in organ damage, with O2−,
H2O2 and •OH being the major ROS
(6,7). Antioxidant enzymes and antioxidants,
including SOD, CAT and GSH, which constitute the defense system of
the body, play a major role in scavenging oxygen free radicals,
dealing with oxidative stress and maintaining the redox balance
(8–10). The increased production of ROS and
the decreased production or excessive consumption of antioxidant
enzymes or antioxidants may cause an imbalance between oxidants and
antioxidants. The MDA content may reflect the degree of lipid
peroxidation damage (18).
Karabulut et al(19)
investigated the changes in the levels of ileum MDA, GSH and nitric
oxide (NO) in rats that underwent 45 min of hepatic ischemia and 30
min reperfusion, and the results demonstrated that oxidative damage
in the ileum was demonstrated by a significant decrease in GSH and
an increase in MDA and NO levels. More recently, Tanrikulu et
al(20) revealed that hepatic
ischemia for 60 min and a reperfusion time of 90 min caused
histological injury of the intestinal mucosa accompanied by an
increase in intestinal MDA and a decrease in GSH-peroxidase. These
studies demonstrated that the oxidant and antioxidant imbalance
participates in intestinal damage induced by hepatic
ischemia-reperfusion.
Hepatic ischemia-reperfusion and intestinal
congestion or hypoxia-reperfusion are the main procedures of liver
transplantation, but it is uncertain whether or not these
procedures change the balance of intestinal oxidants and
antioxidants. The present study demonstrated that all the observed
oxidative indexes changed most significantly at 8 h following OLAT,
showing that OLAT induced a 1.6–1.8-fold increase in
H2O2 and •OH production, as well
as a 2–2.4-fold increase in MDA, a decrease (20–40%) in SOD and CAT
activity and a significant decrease (40–50%) in GSH. The amplitude
of the oxidative and antioxidative indexes decreased at 16 and 24 h
following OLAT. The results of the present study indicated that the
imbalance between oxidants and antioxidants contributed to
intestinal oxidative injury induced by liver transplantation,
particularly in the early phase, while the gradual restoration of
the oxidation/antioxidation system balance was beneficial to
promoting the repair of the intestine. This investigation indicates
that antioxidative intervention is required to alleviate intestinal
oxidative injury. Based on the results of the present study, severe
complications, including SIRS or MODS following liver
transplantation, may be effectively prevented.
Nrf2 is an oxidative-stress sensitive transcription
factor. Following the phosphorylation of serine/threonine,
stimulated by active oxygen, Nrf2 combines with antioxidative
response element to form a complex that regulates the genetic
expression of antioxidase, therefore protecting against oxidative
injury by enhancing antioxidant capacity (11–13,21,22).
Lu et al(23) used
Nrf2-knockout mice and observed that pyrazole caused oxidative
liver damage, which was severe in the Nrf2-knockout mice compared
with the wild-type mice. Jin et al(24) revealed that Nrf2-deficient mice were
more susceptible to traumatic brain injury-induced acute intestinal
mucosal injury than wild-type Nrf2+/+ mice, as
characterized by decreased intestinal mRNA expression and
antioxidative and detoxifying enzyme activities, including
NAD(P)H:quinone acceptor oxidoreductase 1 and glutathione
S-transferase A1. The present study identified that the
expression of intestinal Nrf2 increased at each time point
following OLAT, demonstrating that ROS may promote the activation
of Nrf2 and therefore initiate the process of endogenous
antioxidation. The extent of alteration to all the parameters at 8
h following OLAT was further identified; Nrf2 expression was
increased (~40%), the ROS level was raised >82% and the MDA
levels were increased by 2.4-fold. However, the activity of the
antioxidative enzymes decreased. Nrf2 did not offer enough
antioxidative enzymes against the intestinal injury that was caused
by ROS in the early phase following the graft. However, it may be
hypothesized that the intestine may be repaired gradually with an
increase of the antioxidase activation regulated by Nrf2.
In conclusion, OLAT results in intestinal damage,
which is consistent with the imbalance between oxidation and
antioxidation. The upregulation of Nrf2 following OLAT is not
sufficient to withstand the oxidative injury in the intestine.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (NSFC; no. 30972858) and the Planned
Science and Technology Project of Guangzhou (no. 2011J4300056).
References
|
1
|
Echániz A, Pita S, Otero A, Suárez F,
Gómez M and Guerrero A: Incidence, risk factors and influence on
survival of infectious complications in liver transplantation.
Enferm Infect Microbiol Clin. 21:224–231. 2003.(In Spanish).
|
|
2
|
Hepp J, Zapata R, Buckel E, et al: General
considerations, indications and contraindications for liver
transplantation in Chile: a multicenter consensus development
document. Rev Med Chil. 136:793–804. 2008.(In Spanish).
|
|
3
|
Swank GM and Deitch EA: Role of the gut in
multiple organ failure: bacterial translocation and permeability
changes. World J Surg. 20:411–417. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zheyu C and Lunan Y: Early changes of
small intestine function in rats after liver transplantation.
Transplant Proc. 38:1564–1568. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bedirli A, Sakrak O, Soyuer I and
Muhtarogul S: Portosystemic shunt prevents apoptosis in rat
intestinal mucosa caused by total hepatic ischemia. Eur Surg Res.
36:293–299. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems and apoptosis. Free Radic Biol Med.
48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
He SQ, Zhang YH, Venugopal SK, et al:
Delivery of antioxidative enzyme genes protects against
ischemia/reperfusion-induced liver injury in mice. Liver Transpl.
12:1869–1879. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matés JM: Effects of antioxidant enzymes
in the molecular control of reactive oxygen species toxicology.
Toxicology. 153:83–104. 2000.PubMed/NCBI
|
|
9
|
Sasaki M and Joh T: Oxidative stress and
ischemia-reperfusion injury in gastrointestinal tract and
antioxidant, protective agents. J Clin Biochem Nutr. 40:1–12. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gałecka E, Jacewicz R, Mrowicka M,
Florkowski A and Gałecki P: Antioxidative enzymes - structure,
properties, functions. Pol Merkur Lekarski. 25:266–268. 2008.(In
Polish).
|
|
11
|
Itoh K, Wakabayashi N, Katoh Y, Ishii T,
Igarashi K, Engel JD and Yamamoto M: Keap1 represses nuclear
activation of antioxidant responsive elements by Nrf2 through
binding to the amino-terminal Neh2 domain. Genes Dev. 13:76–86.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shah ZA, Li RC, Thimmulappa RK, Kensler
TW, Yamamoto M, Biswal S and Doré S: Role of reactive oxygen
species in modulation of Nrf2 following ischemic reperfusion
injury. Neuroscience. 147:53–59. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cho HY, Reddy SP and Kleeberger SR: Nrf2
defends the lung from oxidative stress. Antioxid Redox Signal.
8:76–87. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin C, Zhang PJ, Wu XM, et al: Impact of
hypoxic preconditioning on apoptosis and its possible mechanism in
orthotopic liver autotransplantation in rats. Hepatobiliary
Pancreat Dis Int. 8:40–45. 2009.PubMed/NCBI
|
|
15
|
Chi X, Zhang A, Luo G, et al: Knockdown of
myeloid differentiation protein-2 reduces acute lung injury
following orthotopic autologous liver transplantation in a rat
model. Pulm Pharmacol Ther. 26:380–387. 2013. View Article : Google Scholar
|
|
16
|
Chiu CJ, McArdle AH, Brown R, Scott HJ and
Gurd FN: Intestinal mucosal lesion in low-flow states. I. A
morphological, hemodynamic, and metabolic reappraisal. Arch Surg.
101:478–483. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kashfi A, Mehrabi A, Pahlavan PS, et al: A
review of various techniques of orthotopic liver transplantation in
the rat. Transplant Proc. 37:185–188. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bela P, Bahl R, Sane AS, Sawant PH, Shah
VR, Mishra VV and Trivedi HL: Oxidative stress status: possible
guideline for clinical management of critically ill patients.
Panminerva Med. 43:27–31. 2001.PubMed/NCBI
|
|
19
|
Karabulut AB, Kirimlioglu V, Kirimlioglu
H, Yilmaz S, Isik B and Isikgil O: Protective effects of
resveratrol on spleen and ileum in rats subjected to
ischemia-reperfusion. Transplant Proc. 38:375–377. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tanrikulu Y, Kismet K, Serin Kilicoglu S,
et al: Diosmin ameliorates intestinal injury induced by hepatic
ischemia reperfusion in rats. Bratisl Lek Listy. 112:545–551.
2011.PubMed/NCBI
|
|
21
|
Kaspar JW, Niture SK and Jaiswal AK:
Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic Biol
Med. 47:1304–1309. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ishii T, Itoh K, Takahashi S, et al:
Transcription factor Nrf2 coordinately regulates a group of
oxidative stress-inducible genes in macrophages. J Biol Chem.
275:16023–16029. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu YK, Gong PF and Cederbaum AI: Pyrazole
induced oxidative liver injury independent of CYP2E1/2A5 induction
due to Nrf2 deficiency. Toxicology. 252:9–16. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jin W, Wang HD, Hu ZG, Yan W, Chen G and
Yin HX: Transcription factor Nrf2 plays a pivotal role in
protection against traumatic brain injury-induced acute intestinal
mucosal injury in mice. J Surg Res. 157:251–260. 2009. View Article : Google Scholar
|