Introduction
The long-term survival rate among patients with
breast cancer across Europe is >70% (1). This rate is, in part, due to the
widespread use of post-surgery adjuvant anthracycline-based
chemotherapy to treat early-stage breast cancer to reduce the risk
of relapse and mortality (2). The
major anthracycline chemotherapeutic reagent is doxorubicin (DOX),
a Streptomyces metabolite. DOX-based chemotherapy is used
against a wide range of cancers, including hematological
malignancies, soft-tissue sarcomas, lymphomas and various types of
carcinomas, as well as breast cancer, despite the clinical
limitations of the compound, such as cardiotoxicity and the
induction of multidrug resistance (3,4). In an
attempt to address these limitations, DOX therapy in the clinic is
often supplemented by DOX in combination with other
chemotherapeutic reagents. The cytotoxicity of DOX is, in part,
effected through the c-Jun N-terminal kinase (JNK); JNK-dependent
signaling plays a prominent role in DOX-induced cell cycle
withdrawal, differentiation and the control of apoptosis (5,6). The
JNK pathway has been demonstrated to be required for apoptosis
caused by chemotherapeutic agents (7). Thus, synergistic apoptotic responses
may require JNK signals and could consequently be manipulated for
the potentiation of cancer therapies. Our previous study showed
that an extract of an Indonesian marine sponge, Haliclona
sp., demonstrated potent cytotoxicity against multiple human solid
cancer cell lines (8). Previous
studies on flow cytometric analyses and nuclear morphological
changes have indicated that one of the active components of the
extract induced apoptosis; the major cytotoxic activity was
identified as being caused by papuamine (8). Our subsequent study showed that
papuamine inhibits MCF-7 cell survival through the activation of
JNK (9). The present study examined
the potential modulation of doxorubicin cytotoxicity by papuamine
in MCF-7, a human breast cancer cell line.
Materials and methods
Chemicals and cell cultures
Papuamine was isolated from Indonesian marine sponge
Haliclona sp. using our previously published methods
(8). Papuamine was dissolved in
dimethyl sulfoxide and stored as a 20-mM stock solution in
light-proof containers at −20°C. Doxorubicin (DOX) and all other
reagents, unless otherwise stated, were of the highest grade
available and were supplied by either Sigma (St. Louis, MO, USA) or
Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Exposure to
light was kept to a minimum for all drugs used. Immunoblotting
employed rabbit polyclonal antibodies (Cell Signaling Technology,
Inc., Danvers, MA, USA) against human proteins as follows: Anti-JNK
(to detect total JNK levels), anti-phospho-JNK (to detect levels of
phosphorylated JNK) and anti-β-actin (as the loading control). The
MCF-7 human breast cancer cell line was supplied by the Cell
Resource Center for Biomedical Research, Tohoku University (Sendai,
Japan). The cells were maintained in RPMI-1640 medium supplemented
with 10% fetal bovine serum, 100 U/ml penicillin G and 100 μg/ml
streptomycin at 37°C in a humidified 5% CO2-95% air
incubator under standard conditions. Viable cell counts were
determined using exclusion of staining by 0.2% trypan blue. To
maintain exponential growth, the cells were seeded at
5×104 cells/ml in standard tissue culture flasks and
passaged every 3–4 days. For other assays, the cells were cultured
in 2-ml aliquots in 35-mm dishes.
Soft agar colony formation assay
The effect of papuamine and DOX on the colony
formation of the MCF-7 cells was assessed by soft agar colony
formation assay. The assay was performed in 35-mm dishes; each
plate received 2 ml 0.8% agar (in culture medium), which then was
overlaid with a top layer of 1 ml 0.4% agar. Next, ~2,000 cells
were plated over the top layer. The cells were treated with
reagents and maintained at 37°C in a humidified 5% CO2
atmosphere. At day 14 post-treatment, the cells were stained by
exposure (60 min at 37°C) to 100 μl 5-mg/ml
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide. Each
dish then was assessed for the size and number of colonies
(original magnification, ×10).
Western blot analysis
The cells were washed with phosphate-buffered saline
(PBS) and lysed in CelLytic™ M (Sigma) to collect a total cell
lysate, according to the manufacturer’s instructions. Protein
concentration was measured using a Pierce™ BCA Protein Assay kit
(Thermo Fisher Scientific, Inc., Rockford, IL, USA), according to
the manufacturer’s instructions. Following electrophoresis of
protein samples (30 μg/lane) on 10% SDS-polyacrylamide gel, the
protein was transferred to a polyvinylidene difluoride membrane.
The protein was blocked with Blocking One® (Nacalai
Tesque, Inc., Kyoto, Japan) for 1 h and incubated with a first
antibody overnight at 4°C. The membrane was then washed with wash
buffer (PBS containing 0.05% Tween-20) and incubated with
anti-rabbit horseradish peroxidase-linked secondary antibody (Cell
Signaling Technology, Inc.) for 1 h. Subsequent to another wash
with wash buffer, the protein levels were analyzed by enhanced
chemiluminescence with Pierce Western Blotting substrate (Thermo
Fisher Scientific, Inc.).
DOX cellular accumulation assay
The cellular accumulation of DOX was ascertained
using a Tecan Infinite® M1000 microplate reader (Tecan
Group Ltd., Männedorf, Switzerland) at excitation and emission
wavelengths of 485 and 590 nm, respectively. Briefly, the cells
were seeded at a 5×103/well in Nunc™ MicroWell™ 96-well
optical-bottom plates (Thermo Fisher Scientific, Inc.) and
incubated overnight under standard culture conditions. Media was
replaced with culture medium supplemented with various
concentrations of papuamine (1, 2 or 5 μM) and doxorubicin (1, 3,
10 and 30 μM) and cultures were incubated for 4 h. The medium was
removed and washed twice with PBS, and the residual fluorescence
intensity was measured as aforementioned.
Statistical analysis
Statistical analysis was performed by one- or
two-way analysis of variance followed by Williams’ type multiple
comparison or Bonferroni tests among multiple groups. Data are
expressed as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Inhibition of colony formation by
papuamine and DOX
First, the present study examined whether incubation
with papuamine and DOX, alone and in combination, affected the
ability of MCF-7 cells to form colonies in soft agar. Following
treatment with reagents, the cells were placed into medium with
soft agar, and colonies were counted subsequent to 2 weeks. As
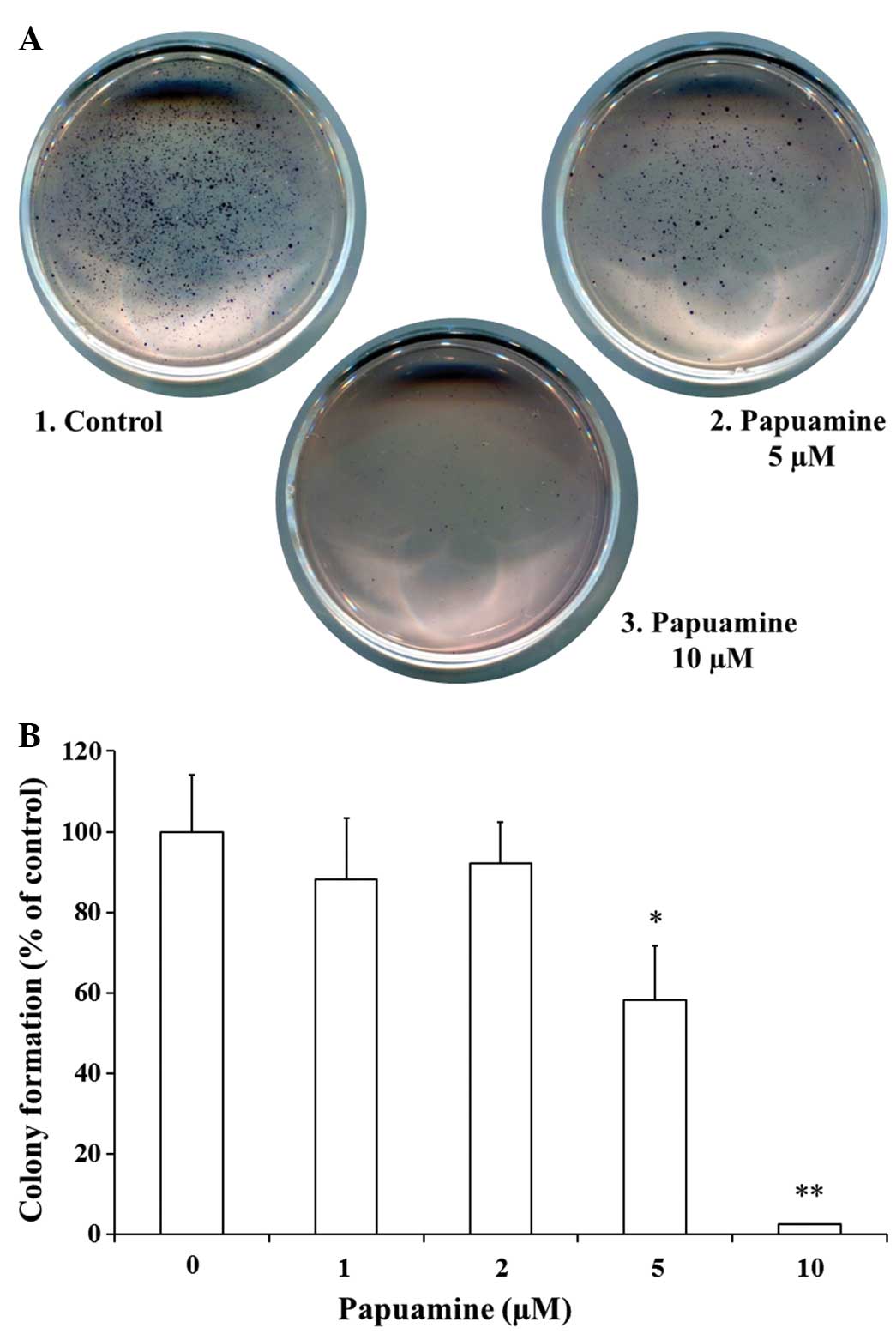
shown in Fig. 1A and B, incubation
with papuamine alone at 5 or 10 μM provided significant
concentration-dependent inhibition of colony formation (58.3±13.4%,
P<0.05 at 5 μM; 2.1±0.2%, P<0.01 at 10 μM) compared with the
control. Papuamine exhibited little effect at lower concentrations
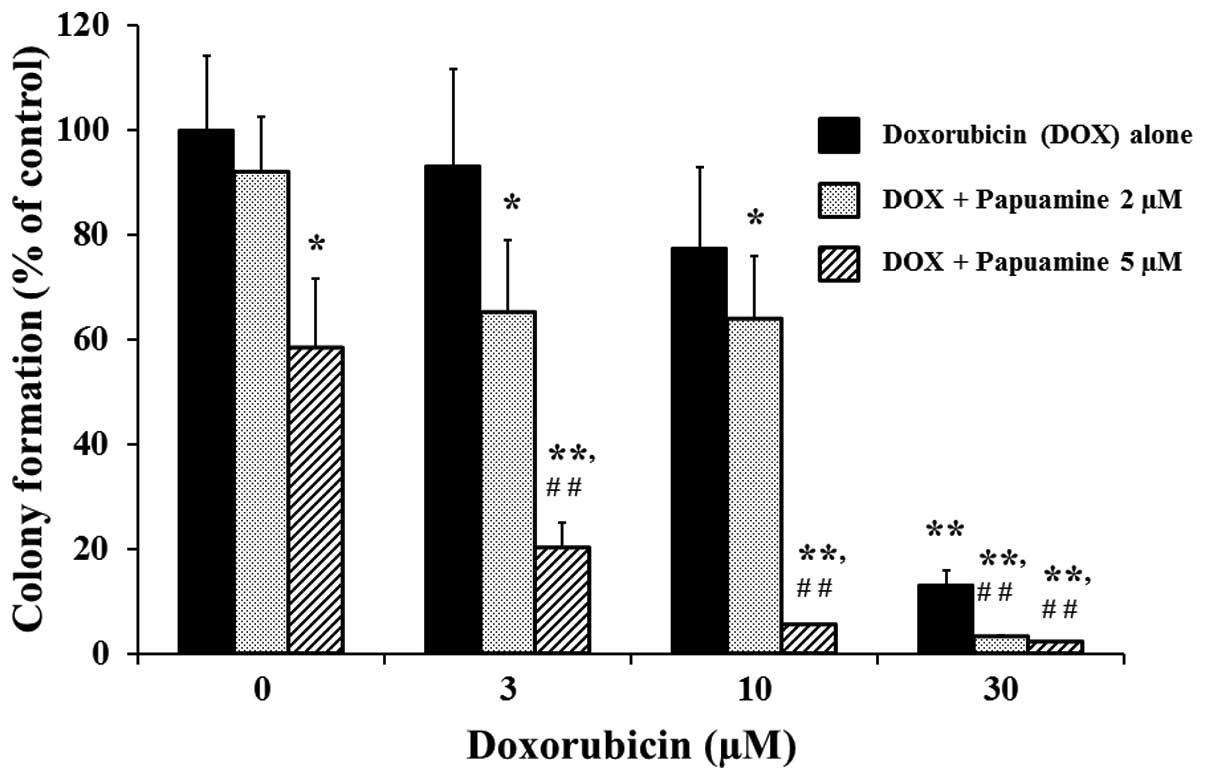
of ≤2 μM. As shown in Fig. 2,
incubation with DOX alone at 3, 10, or 30 μM provided significant
(P<0.01) concentration-dependent inhibition of colony formation
(93.2±18.4, 77.4±15.4 and 13.2±3.2%, respectively) compared with
the control. These effects were significantly enhanced by
co-incubation with papuamine at 2 or 5 μM (P<0.01 compared to
single incubation with DOX).
Detection of JNK activation by papuamine
and DOX
Next, the study examined the possible involvement of
the JNK pathway in the inhibition of cell growth in MCF-7 cells.
JNK activation was determined by western blot analysis following a
24-h treatment of the cells with reagents (Fig. 3). Phosphorylated JNK (phospho-JNK),
an activated form of the protein, was detected at low levels
following incubation with either papuamine (5 μM) or DOX (3 μM)
alone. Phospho-JNK levels exhibited concentration-dependent
increases upon co-incubation with papuamine and DOX (Fig. 3, upper panel). In control blots, the
levels of total JNK and β-actin were not changed under these same
conditions (Fig. 3, lower and
middle panels).
Effect of papuamine on DOX cellular
accumulation
The effect of papuamine may be exerted by changes in
the cellular uptake or efflux of DOX in MCF-7 cells. Therefore, the
effect of papuamine on DOX cellular accumulation was examined using
a fluorescence assay (Fig. 4). DOX
autofluoresces with λex=485 nm and λem=590 nm. Incubation with DOX
alone (at 1 to 30 μM) for 4 h showed concentration-dependent
increases in fluorescence intensity. Co-incubation with papuamine
at 1, 2 or 5 μM resulted in small (≤25%) decreases in DOX
fluorescence. Notably, however, these decreases were not
statistically significant or concentration-dependent for
papuamine.
Discussion
To evaluate the effects of papuamine and DOX on cell
growth, colony formation assays were performed in soft agar in the
present study. Soft agar colony formation assays are commonly used
as a monitoring system for anchorage-independent
(three-dimensional) growth, measuring proliferation by the counting
of colonies subsequent to 3–4 weeks in a semi-solid culture medium.
This traditional method has been widely published (10–12).
As the technique delivers results that are comparable to those
obtained when injecting tumorigenic cells into nude mice, the soft
agar colony formation assay is regarded as the gold standard for
testing the tumorigenicity of cells in vitro (13). Notably, in the present study,
incubation with 10 μM papuamine alone provided significant
inhibition of colony formation by the MCF-7 cells, while incubation
with the same concentration (10 μM) of DOX did not yield
significant inhibition, indicating that papuamine was more potent
than DOX. Co-incubation with papuamine and DOX enhanced the
inhibition of colony formation. Thus, these results indicate
potential efficacy for the use of a combined papuamine and DOX
chemotherapy regime in the treatment of breast cancer. To confirm
the possible mechanisms of the combinatory effects of papuamine and
DOX in MCF-7 cells, the activation of JNK and the cellular
accumulation of DOX was examined in the treated cells. JNK has
previously been indicated to be a key player in the DOX-induced
apoptotic cascade (14), and our
previous study showed that papuamine-induced cytotoxicity involved
JNK activation (9). In the present
study, it was demonstrated that these reagents concurrently
increased JNK activation, consistent with their additive action in
inhibiting colony formation. Previous studies have shown that DOX
is a substrate for P-glycoprotein (P-gp), meaning that the
inhibition of P-gp could increase the cellular accumulation of DOX
and enhance its chemotherapeutic effect (15–17).
However, the present study demonstrated that papuamine did not
significantly alter DOX cellular accumulation in MCF-7 cells. These
results indicate that papuamine does not enhance DOX efficacy by
increasing DOX accumulation (as may apply for changes in P-gp),
suggesting that papuamine represents a novel type of modulator for
DOX chemotherapy.
In conclusion, papuamine and DOX exhibit synergy
when used as a combination treatment in MCF-7 cells. This
combination effect is not mediated by changes in the cellular
accumulation of DOX, but appears to reflect a shared activation of
JNK phosphorylation. The present study may therefore aid in the
development of therapeutic strategies for breast cancer
treatments.
References
|
1
|
Sant M, Allemani C, Santaquilani M, Knijn
A, Marchesi F and Capocaccia R; EUROCARE Working Group. EUROCARE-4.
Survival of cancer patients diagnosed in 1995–1999 Results and
commentary. Eur J Cancer. 45:931–991. 2009.
|
|
2
|
Smith LA, Cornelius VR, Plummer CJ, et al:
Cardiotoxicity of anthracycline agents for the treatment of cancer:
systematic review and meta-analysis of randomised controlled
trials. BMC Cancer. 10:3372010.
|
|
3
|
Minotti G, Menna P, Salvatorelli E, Cairo
G and Gianni L: Anthracyclines: molecular advances and
pharmacologic developments in antitumor activity and
cardiotoxicity. Pharmacol Rev. 56:185–229. 2004.
|
|
4
|
Gewirtz DA: A critical evaluation of the
mechanisms of action proposed for the antitumor effects of the
anthracycline antibiotics adriamycin and daunorubicin. Biochem
Pharmacol. 57:727–741. 1999.
|
|
5
|
Kim J and Freeman MR: JNK/SAPK mediates
doxorubicin-induced differentiation and apoptosis in MCF-7 breast
cancer cells. Breast Cancer Res Treat. 79:321–328. 2003.
|
|
6
|
Li F, Meng L, Xing H, et al: Essential
role of c-Jun-NH2-terminal kinase on synergy induction of apoptosis
by TRAIL plus ADM in ADM resistant MCF-7/ADM cells. Apoptosis.
11:1239–1246. 2006.
|
|
7
|
Mansouri A, Ridgway LD, Korapati AL, et
al: Sustained activation of JNK/p38 MAPK pathways in response to
cisplatin leads to Fas ligand induction and cell death in ovarian
carcinoma cells. J Biol Chem. 278:19245–19256. 2003.
|
|
8
|
Yamazaki H, Wewengkang DS, Kanno S, et al:
Papuamine and haliclonadiamine, obtained from an Indonesian sponge
Haliclona sp, inhibited cell proliferation of human cancer
cell lines. Nat Prod Res. 27:1012–1015. 2013.
|
|
9
|
Kanno S, Yomogida S, Tomizawa A, et al:
Papuamine causes autophagy following the reduction of cell survival
through mitochondrial damage and JNK activation in MCF-7 human
breast cancer cells. Int J Oncol. 43:1413–1419. 2013.
|
|
10
|
Hanauske AR, Hanauske U and Von Hoff DD:
The human tumor cloning assay in cancer research and therapy: a
review with clinical correlations. Curr Probl Cancer. 9:1–66.
1985.
|
|
11
|
Slee PH, Van Oosterom AT and De Bruijn EA:
Predictive testing in cancer chemotherapy. II. In vitro. Pharm
Weekbl Sci. 7:125–133. 1985.
|
|
12
|
Shoemaker RH, Wolpert-DeFilippes MK, Kern
DH, et al: Application of a human tumor colony-forming assay to new
drug screening. Cancer Res. 45:2145–2153. 1985.
|
|
13
|
Freedman VH and Shin SI: Cellular
tumorigenicity in nude mice: correlation with cell growth in
semi-solid medium. Cell. 3:355–359. 1974.
|
|
14
|
Panaretakis T, Laane E, Pokrovskaja K, et
al: Doxorubicin requires the sequential activation of caspase-2,
protein kinase Cdelta, and c-Jun NH2-terminal kinase to induce
apoptosis. Mol Biol Cell. 16:3821–3831. 2005.
|
|
15
|
Ferry DR: Testing the role of
P-glycoprotein expression in clinical trials: applying
pharmacological principles and best methods for detection together
with good clinical trials methodology. Int J Clin Pharmacol Ther.
36:29–40. 1998.
|
|
16
|
Robert J and Jarry C: Multidrug resistance
reversal agents. J Med Chem. 46:4805–4817. 2003.
|
|
17
|
Sikic BI, Fisher GA, Lum BL, Halsey J,
Beketic-Oreskovic L and Chen G: Modulation and prevention of
multidrug resistance by inhibitors of P-glycoprotein. Cancer
Chemother Pharmaco. 40(Suppl): S13–S19. 1997.
|