Introduction
Temozolomide (TMZ;
3,4-dihydro-3-methyl-4-oxoimidazo-[5,1-d]-1,2,3,5-tetrazin-8-carboxamide),
a 3-methyl analogue of mitozolomide, is an imidazotetrazine class
methylating agent. Similarly to dacarbazine (DTIC), the active
metabolite of TMZ is the linear triazine monomethyl
triazenoimidazole carboxamide (MTIC) (1). DTIC requires initial activation in the
liver, while TMZ is metabolized to MTIC in the peripheral blood at
physiological pH. The subsequent degradation of MTIC to the highly
reactive methyldiazonium ion results in the formation of toxic
methyl adducts with DNA bases. The methyldiazonium ion promotes
cytotoxicity via O6-methylation of guanine,
leading to base-pair mismatch, inhibition of DNA replication and
subsequent cell cycle arrest and apoptosis (2). TMZ resistance has been ascribed to the
removal of methyl groups by
O6-methylguanine-DNA-methyltransferase.
Furthermore, in vitro and human xenograft studies have
demonstrated a broad spectrum of TMZ activity in murine tumors
(2).
Musculoskeletal sarcomas (MSS) are a heterogeneous
group of malignant neoplasms, which are derived from the connective
tissue. Sarcomas represent ~1% of cancers in adults. MSS may occur
anywhere in the body; however, the lower extremities represent the
most common site of appearance, followed by the upper extremities,
trunk, retroperitoneum and the head and neck area (3). MSS treatment has markedly improved in
recent years, owing to a greater understanding of the local growth
characteristics of tumors, novel computed tomography (CT) and
magnetic resonance imaging (MRI) technology for the evaluation and
diagnosis of solid tumors, as well as the development of
multidisciplinary treatments that facilitate local tumor treatment
and functional tissue preservation (4). Surgical resection is the primary
treatment strategy for MSS; however, chemotherapy has become more
common for treatment of the majority of bone sarcomas, and may have
potential benefits for the treatment of patients with soft-tissue
sarcomas (STS). Numerous patients that present with locally
advanced disease may require multimodal therapy, including
chemotherapy and chemo-radiation. Doxorubicin and ifosfamide are
typically recommended as the first-line systemic treatment in
unresectable and/or metastatic STS. However, despite an initial
anti-tumor response, numerous patients gradually develop resistance
to these therapies. Salvage therapy options following the failure
of frontline MSS chemotherapy are limited; thus, novel, more
effective agents are required for the successful treatment of these
diseases.
Administration of TMZ alone or in combination with
other chemotherapeutic agents has demonstrated activity in patients
with pretreated MSS, particularly among patients with certain
histological subtypes, for example leiomyosarcoma, solitary fibrous
tumors and Ewing's sarcoma (5–9). However,
the enhancement in patient survival following treatment with TMZ is
not significant. Thus, elucidating the mechanism of TMZ action
against MSS is essential. Therefore, the present study aimed to
determine the effect of TMZ on cell viability in MSS in order to
identify histological subtypes that may be suitable for TMZ-based
treatment.
Materials and methods
Cell lines and cell culture
Sarcoma cell lines, including the NOS1 osteosarcoma
cell line (10), NMS-2 malignant
peripheral nerve sheath tumor cell line (11) and NEPS epithelioid sarcoma cell line
(12), were established under the
approval of the Institutional Review Board of Niigata University
Hospital (Niigata, Japan). The HS-SY-II synovial sarcoma cell line
was a gift from Dr H. Sonobe (Department of Pathology, Kochi
Medical School, Kochi, Japan) (13),
the SYO-1 synovial sarcoma cell line was gifted by Dr A. Kawai
(Department of Orthopedic Surgery, Okayama University, Okayama,
Japan) (14), the 402–92 liposarcoma
cell line was a gift from Dr P. Åman (Department of Clinical
Genetics, University Hospital, Lund, Sweden) (15), the ASPS-KY alveolar soft part sarcoma
cell line was a gift from Dr S. Yanoma (Kanagawa Cancer Center,
Yokohama, Japan) (16) and the
FU-EPS-1 (12) and SFT-8606 (17) epithelioid sarcoma cell lines were
gifts from Dr H. Iwasaki (Department of Pathology, Fukuoka
University School of Medicine, Fukuoka, Japan). The aforeentioned
cell lines underwent 50–100 passages and screening tests confirmed
no mycoplasma infection. The remaining cell lines used in the
present study were obtained from commercial sources, and all cell
lines used are outlined in Table I.
NOS1, Saos-2, NMS-2, 402-92, ASPS-KY, HT-1080, SFT-8606, FU-EPS-1
and NEPS cells were maintained in RPMI-1640 (Invitrogen Life
Technologies, Carlsbad, CA, USA). U-87 MG, HS-SY-II and SYO-1 cells
were maintained in Dulbecco's modified Eagle's medium (Invitrogen
Life Technologies). The SKNMC cells were maintained in α-minimum
essential medium (Invitrogen Life Technologies). All media were
supplemented with 10% fetal bovine serum (FBS; PAA Laboratories
GmbH, Pasching, Austria) containing 1% antibiotics and antimycotics
(penicillin, streptomycin and amphotericin B; Invitrogen Life
Technologies). All cell cultures were incubated at 37°C in an
atmosphere containing 5% CO2 with 100% humidity.
 | Table I.Summary of human cell lines used in
the present study. |
Table I.
Summary of human cell lines used in
the present study.
| Name | Cell origin | Source |
|---|
| SKNMC | Ewing's sarcoma | ATCC |
| Saos-2 | Osteosarcoma | ATCC |
| NOS1 | Osteosarcoma | Niigata University,
Niigata, Japan |
| NMS-2 | Malignant peripheral
nerve sheath tumor | Niigata
University |
| NEPS | Epithelioid
sarcoma | Niigata
University |
| FU-EPS-1 | Epithelioid
sarcoma | Dr H. Iwasaki |
| SFT-8606 | Epithelioid
sarcoma | Dr H. Iwasaki |
| HS-SY-II | Synovial sarcoma | Dr H. Sonobe |
| SYO-1 | Synovial sarcoma | Dr A. Kawai |
| HT-1080 | Fibrosarcoma | Health Science
Research Resources Bank, Osaka, Japan |
| 402-92 | Liposarcoma | Dr P. Åman |
| ASPS-KY | Alveolar soft part
sarcoma | Dr S. Yanoma |
| U-87 MG | Glioblastoma | Riken Cell Bank,
Tsukuba, Japan |
Effect of TMZ on cell viability and
morphology
All cell lines were plated on 96-well plates
(~3.5×103 cells/well) and incubated for 24 h.
Subsequently, cells were washed with medium and incubated with
fresh medium (control) or medium containing 125, 250, 500 or 1000
µM TMZ. Cell viability was assessed using the XTT (sodium
3′-[1-(phenylaminocarbonyl)-3,4-tetrazolium]-bis
(4-methoxy-6-nitro) benzene sulfonic acid hydrate) assay according
to the manufacturer's instructions, following exposure to the
various concentrations of TMZ for 120 h at 37°C. Subsequently, the
plates were incubated for 4 h at 37°C under the same conditions as
the previous 120-h incubation. Absorbance was measured at 490 nm
against a reference wavelength of 655 nm using a microplate reader
(model 680; Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
cell viability (%) and IC50 value (µM) were calculated
relative to the controls at 100%, and the mean IC50
value was calculated from three experiments experiments using
CurveExpert software (version 1.3; Daniel G. Hyams, Madison, AL,
USA). Subsequent to the 120-h incubation but prior to the 4-h
incubation, cell morphology was observed daily using an Olympus
phase-contrast microscope ULWCD 0.30 (IMT2; Olympus Corp., Tokyo,
Japan) and photomicrographs of the control wells and 1000 µM TMZ
wells were taken at a magnification of ×40.
Western blot analysis of apoptosis and
cell survival/proliferation pathways
Cultured cells were harvested and subjected to
western blot analysis to assess the induction of apoptosis via the
phosphorylation status of phosphoinositide 3-kinase (PI3K)/Akt and
extracellular signal-regulated kinase (ERK)1/2 mitogen activated
protein kinase (MAPK), and the cleavage of caspase-3 and poly
(adenosine diphosphate-ribose)-polymerase (PARP). The cells were
plated on 60-mm culture dishes at a density of
0.3–5.0×105 cells/dish 24 h prior to treatment with 250
µM TMZ. At 24, 48 and 72 h post-exposure, the cells were subjected
to western blot analysis to determine the expression levels of each
of the aforementioned proteins. Briefly, the cells were washed
twice with ice-cold phosphate-buffered saline (PBS) and suspended
in SDS-PAGE running buffer comprised of: 62.5 mM Tris, pH 6.8; 2%
SDS; 5% glycerol and 6 M urea (Invitrogen Life Technologies). Cell
lysates were passed through a 23-gauge needle (Terumo, Tokyo,
Japan) ten times, and the protein concentration of the supernatant
was measured by bicinchoninic acid assay (Pierce Biotechnology,
Inc.; Thermo Fisher Scientific, Rockford, IL, USA). Subsequently,
dithiothreitol (50 mM) and bromophenol blue (0.025%)
(Sigma-Aldrich, St. Louis, MO, USA) were added, and the samples
were boiled for 5 min at 95°C and stored at −80°C. Equal quantities
of protein were subjected to 7% SDS-PAGE and then transferred to a
nitrocellulose membrane (GE Healthcare Life Sciences, Little
Chalfont, UK). The membrane was probed with primary antibodies
against caspase-3 (rabbit polyclonal; cat no. 9662; 1:1,000), PARP
(rabbit polyclonal; cat no. 9542; 1:1,000), pT202/Y204-MAPK (rabbit
polyclonal; cat no. 9101; 1:1,000), total MAPK (rabbit polyclonal;
cat no. 9102; 1:1,000), pS473-AKT (rabbit polyclonal; cat no. 9271;
1:1,000), total AKT (rabbit polyclonal; cat no. 9272; 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA), and actin (mouse
monoclonal; cat no. A2228; 1:3,000; Sigma-Aldrich), and then with
horseradish peroxidase-conjugated IgG donkey anti-rabbit (cat no.
NA934; 1:3,000) or sheep anti-mouse (cat no. NA931; 1:3,000)
secondary antibodies (GE Healthcare Life Sciences). The bands were
visualized by enhanced chemiluminescence (ECL Western Blotting
Detection kit; GE Healthcare Life Sciences).
Statistical analysis
All data are expressed as the mean ± standard
deviation. The differences between groups were evaluated using a
two-tailed Student's t-test and all statistical analysis was
performed using Microsoft Excel software (Microsoft Corporation,
Redmond, WA, USA). P<0.01 was considered to indicate a
statistically significant difference.
Results
Cell growth is inhibited in
TMZ-treated MSS cells
As shown in Fig. 1,
TMZ reduced the viability of all tumor cells in a dose-dependent
manner following incubation for 120 h (P<0.01). However, the
IC50 value of TMZ varied between cell lines. In U87-MG
cells, the IC50 was 348 µM, whereas the IC50
value of TMZ in SKNMC, NOS1, HS-SY-II, SYO-1 and 402-92 cells was
<300 µM. By contrast, the IC50 value of TMZ in
HT1080, NMS-2, Saos-2, ASPS-KY, NEPS, FU-EPS-1 and SFT-8606 cells
was >366 µM. Based on these results, TMZ inhibited the
proliferation of SKNMC, NOS1, HS-SY-II, SYO-1 and 402-92 cells more
potently than that of U87-MG cells. By contrast, HT1080, NMS-2,
Saos-2, ASPS-KY, NEPS, FU-EPS-1 and SFT-8606 cells were more
resistant to TMZ than U87-MG cells.
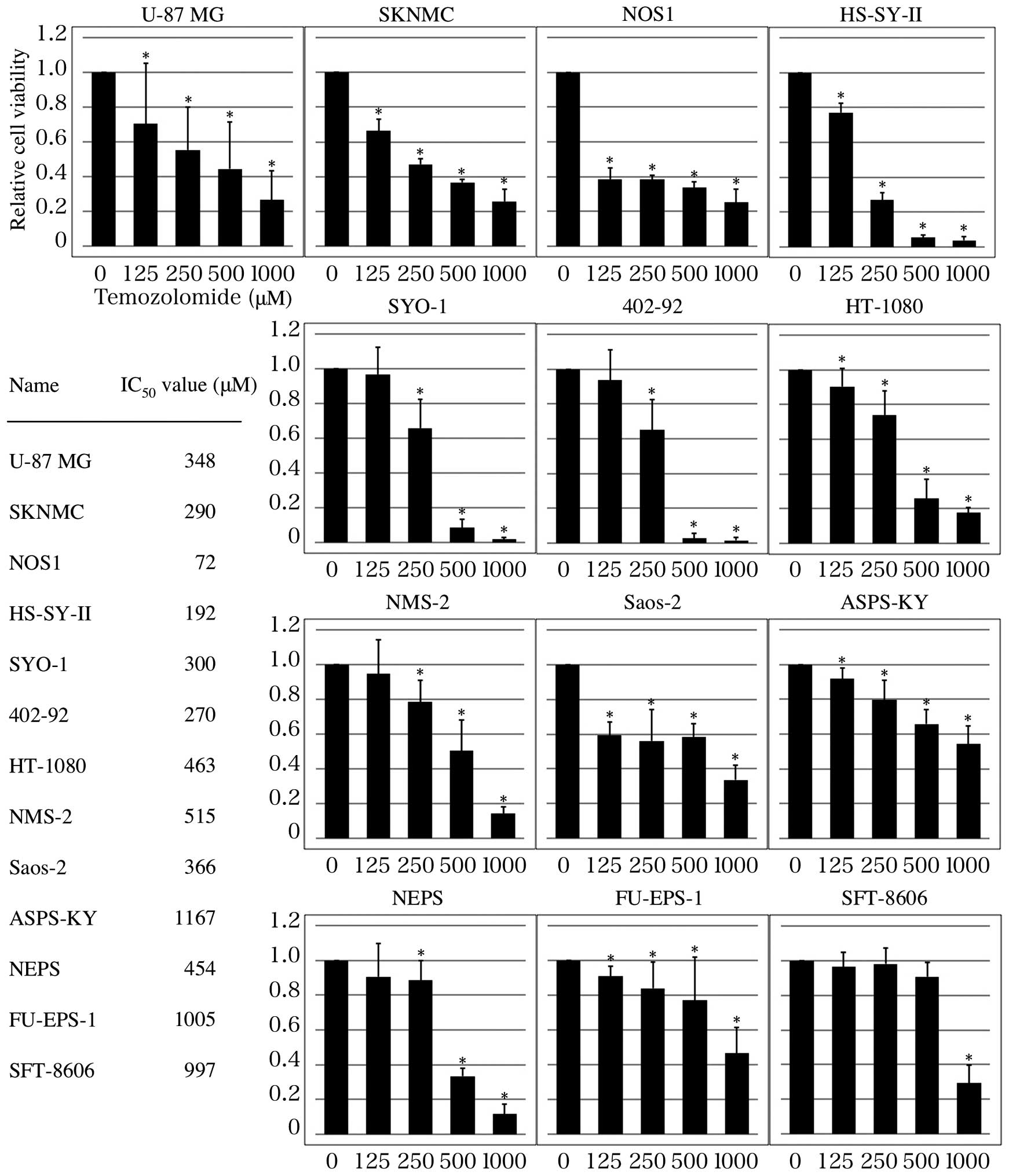 | Figure 1.TMZ inhibits MSS cell growth.
Inhibitory effects of TMZ on MSS growth were determined using an
XTT assay. Cell viability (%) and IC50 values (µM) were
calculated relative to the controls at 100%, and the mean
IC50 value was calculated from three replicates. The
data represents the mean of three experiments. Based on these
results, TMZ inhibited the growth of SKNMC, NOS1, HS-SY-II, SYO-1
and 402-92 cells more potently than U-87 MG cells. By contrast,
HT1080, NMS-2, Saos-2, ASPS-KY, NEPS, FU-EPS-1 and SFT-8606 cells
were more resistant to TMZ than U-87 MG cells. *P<0.01 vs.
non-treated control group. TMZ, temozolomide; MSS, musculoskeletal
sarcoma. |
TMZ induces morphological changes in
MSS cells
Photomicrographs of TMZ-treated cells were obtained
using phase-contrast microscopy. Cell death was observed 120 h
post-exposure to 1000 µM TMZ, as indicated by a mass of rounded,
floating, highly refractive cells. Such cell death was observed in
11 of the 13 cell lines evaluated (Fig.
2). Notably, no cell lysis was observed in the FU-EPS-1 or
ASPS-KY cells (IC50 >1000 µM).
TMZ induces apoptosis in all cell
lines
Following treatment with TMZ, all 13 cell lines were
examined for evidence of apoptosis by western blot analysis, using
caspase-3 and PARP antibodies (Fig.
3). Activation of caspase-3 via protein cleavage has a critical
role in apoptosis (18). Furthermore,
the proteolytic cleavage of PARP by activated caspase-3 serves as a
more prominent marker of apoptosis (19). The results of the present study
indicated that pro-caspase-3 and PARP levels were reduced, while
cleaved caspase-3 and PARP were upregulated. These data indicated
that TMZ induced caspase activation and proteolysis of caspase-3
and PARP in MSS cells. In particular, a marked enhancement in
caspase-3 cleavage was observed at 48 and 72 h following TMZ
exposure in the SKNMC, Saos-2, NOS1, HS-SY-II, HT1080, 402-92 and
NEPS cells.
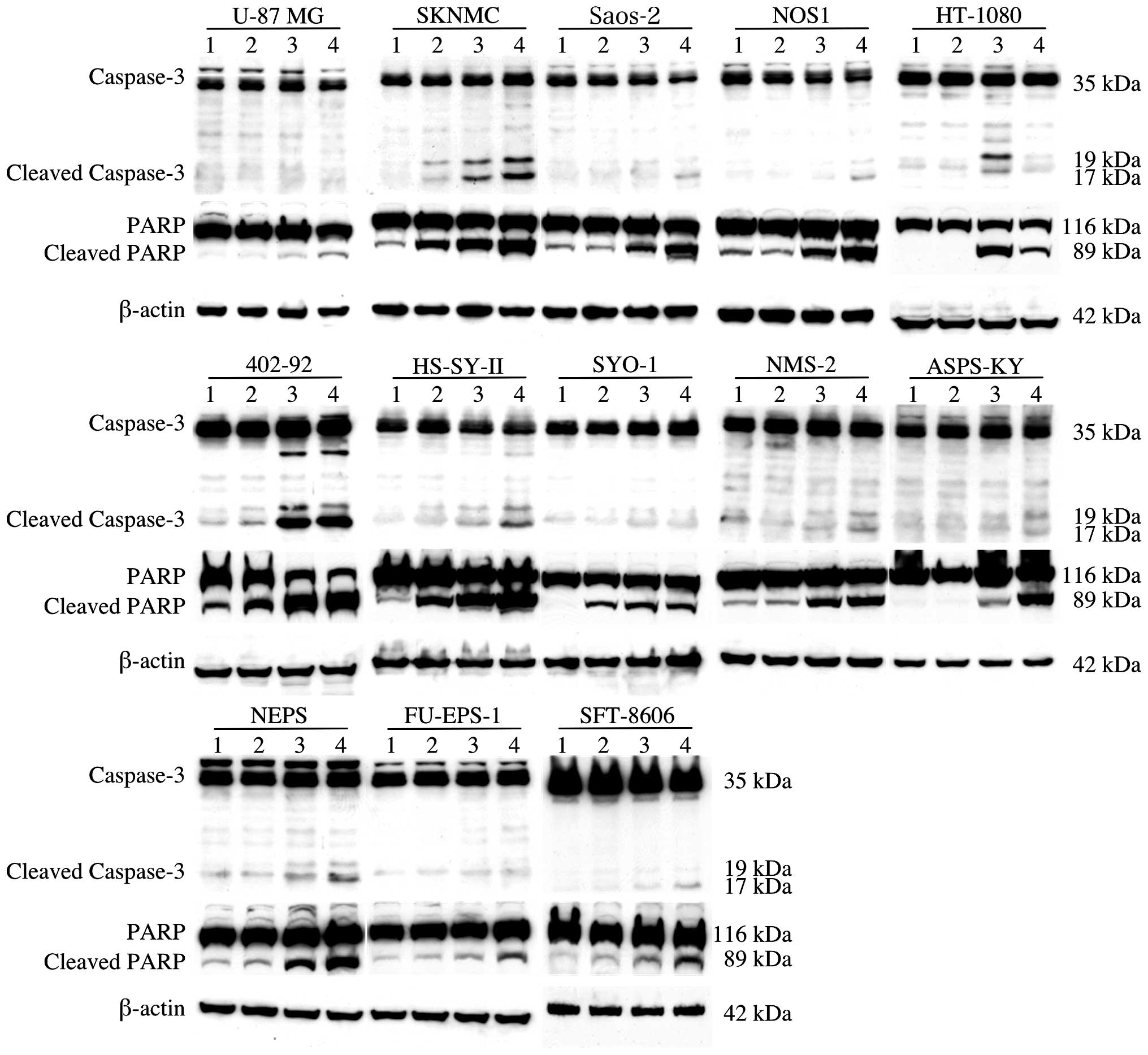 | Figure 3.Western blot analysis of apoptotic
proteins. Cleavage of caspase-3 has a critical role in apoptosis,
and the proteolytic cleavage of PARP by activated caspase-3 serves
as a marker of apoptosis. Apoptosis was induced by TMZ in all
tested cell lines. Marked activation of caspase-3 protein was
detected at 48 or 72 h following exposure to TMZ in SKNMC, Saos-2,
NOS1, HS-SY-II, HT-1080, 402-92 and NEPS cells. Lanes: 1, control;
2, treated with TMZ for 24 h; 3, treated with TMZ for 48 h; 4,
treated with TMZ for 72 h. TMZ, temozolomide; PARP, poly adenosine
diphosphate ribose polymerase. |
Effects of PI3K/Akt and ERK1/2 MAPK on
MSS cell survival and proliferation
The initiation of apoptosis is dependent on the DNA
repair system, as well as on multiple signaling pathways, including
the phosphorylation status of PI3K/Akt and ERK1/2 MAPK. These
signaling pathways were evaluated via western blot analysis. The
results indicated that endogenous Akt was characterized by the
phosphorylation of Ser473 (Fig. 4).
Furthermore, the basal levels of p-Akt were dose-dependently
reduced in SYO-1, HT-1080, NMS-2 and ASPS-KY cells following
treatment with 250 µM TMZ for 24 h. Typically, p-Akt is
constitutively expressed in U-87 MG cells (20,21).
Although treatment with TMZ initially reduced p-Akt expression,
p-Akt markedly increased at 72 h. We hypothesze that whilst
constitutive activation of this pathway is emerging as a marker of
sensitivity to TMZ, the activity of TMZ may be limited by the
presence of feedback loops, leading to the reactivation of p-AKT.
The activation status of ERK1/2 MAPK was examined by evaluating the
expression of ERK1/2 by western blotting. The results indicated
that there was basal activation of ERK1 and 2 in the MSS cells. In
the presence of 250 µM TMZ, the levels of p-ERK1/2 were marginally
decreased in ASPS-KY and NEPS cells following 48 h treatment.
 | Figure 4.Western blot analysis of cell
survival/proliferation pathways. PI3K/Akt and ERK1/2 MAPK are
associated with cell survival and proliferation. The
phosphorylation status of PI3K/Akt and ERK1/2 MAPK was evaluated by
western blotting. Basal levels of p-Akt were reduced in a
time-dependent manner in SYO-1, HT-1080, NMS-2, and ASPS-KY cells
following treatment with 250 µM TMZ. By contrast, the expression of
p-ERK1/2 was 2 was marginally decreased in ASPS-KY and NEPS cells.
Lanes: 1, control; 2, treated with TMZ for 24 h; 3, treated with
TMZ for 48 h; 4, treated with TMZ for 72 h. TMZ, temozolomide;
PI3K, phosphoinositide 3-kinase; ERK1/2, extracellular
signal-regulated kinase 1/2; MAPK, mitogen activated protein
kinase. |
Discussion
The activity of TMZ in patients with glioblastoma is
dependent on silencing of the
O6-methylguanine-DNA methyltransferase (MGMT) DNA
repair gene (22). However, MGMT gene
silencing may also occur via promoter methylation in soft tissue
sarcomas, although this is rare (23). The ability of alkylating agents to
induce apoptosis is dependent not only on MGMT activity, but also
on the activity of various survival pathways (24,25), and
the mechanism to induce the apoptosis in MSS may differ from in
glioblastoma. Therefore, analysis of the pro-apoptotic effects of
TMZ, as well as the study of interactions between TMZ and
anti-apoptotic/proliferation pathways (PI3K/Akt and ERK1/2 MAPK),
may contribute to improving MSS patient survival.
The results of the present study demonstrated that
TMZ dose-dependently reduced the viability of MSS cells by inducing
apoptosis, with a parallel increase in caspase-3 and PARP activity.
An XTT assay was used to compare the sensitivity of TMZ of treated
MSS cells and U-87 MG cells to TMZ (26,27). The
IC50 values of the MSS cells following 120 h of
treatment ranged from 72 to 1167 µM, whereas the IC50
value in U-87 MG glioblastoma cells was 348 µM. In 5 of the 12 MSS
cell lines tested, the IC50 value was lower than that of
U-87 MG cells. TMZ treatment was also associated with marked
morphological changes, including cellular floating and cell debris
accumulation following 120 h treatment with 1000 µM TMZ. Although
TMZ induced apoptosis in all cell lines evaluated, the sensitivity
of these sarcomas to TMZ varied. These findings regarding cell
viability suggest that TMZ treatment may be effective in certain
types of MSS.
The mechanism of TMZ action remains controversial.
Hirose et al (28) reported
that, in glioma cells, TMZ treatment induced low levels of
apoptosis compared with those observed in lymphoid cells. In
addition, TMZ was shown to induce cell cycle arrest in the G2/M
phase. By contrast, Kanzawa et al (29) demonstrated that TMZ induced autophagy,
but not apoptosis, in malignant glioma cells. Furthermore, Roos
et al (27) indicated that
cell death induced by TMZ in gliomas was due to apoptosis. In the
present study, TMZ was demonstrated to significantly increase the
activation of apotosis-associated proteins caspase-3 and PARP. This
induction of apoptosis may explain the reduction in MSS cell
proliferation observed in the presence of TMZ, and may constitute a
novel apoptotic mechanism in these cells.
The PI3K/Akt and ERK1/2 MAPK signaling pathways are
significant in protein synthesis, cell proliferation, angiogenesis
and metabolism (30). Therefore, the
phosphorylation of these signaling pathway components was also
analyzed in the present study. The results indicated that PI3K/Akt
was constitutively active in MSS cells, and that this activation
was suppressed in certain cells and maintained in others following
treatment with TMZ. Notably, p-Akt markedly increased at 72 h,
indicating possible re-activation of p-Akt. The data also revealed
that ERK1/2 MAPK activation was marginally decreased in certain
cell types. Combined, these results indicated that MSS cells are
characterized by aberrant activation of these signaling pathways,
an effect which is maintained or suppressed in the presence of TMZ.
TMZ decreased the viability of certain cell lines. The viability of
all cell lines only decreased in very high concentrations of TMZ.
Therefore, it is considered that certain MSS cells were resistant
while some were relatively sensitive to TMZ. Furthermore, these
observations emphasize the plasticity of MSS cells and their
ability to resist chemotherapy by blocking the induction of
apoptosis. This plasticity may explain the resistance of some MSS
cells to TMZ, as well as the differences in MSS cell sensitivity to
alkylating agents. These results also suggested that combinations
of traditional chemotherapy with molecular-based therapy, in
particular therapy associated with signaling pathways, may be used
to effectively treat MSS. The methylation status of the MGMT
promoter in MSS cells was not determined in the present study, and
additional experiments may be required to clarify whether there is
an association between MGMT status and MSS cell sensitivity to
TMZ.
Though the apoptotic signaling pathways induced
during TMZ treatment remain elusive, understanding the mechanisms
involved in the induction of apoptosis may contribute to improving
the prognosis of patients with MSS. More detailed investigations
are needed to clarify whether TMZ presents a promising strategy for
MSS therapy or not.
References
|
1
|
Gottlieb JA, Benjamin RS, Baker LH, et al:
Role of DTIC (NSC-45388) in the chemotherapy of sarcomas. Cancer
Treat Rep. 60:199–203. 1976.PubMed/NCBI
|
|
2
|
Stevens MF, Hickman JA, Langdon SP, et al:
Antitumor activity and pharmacokinetics in mice of
8-carbamoyl-3-methyl-imidazo [5,1-d]-1,2,3,5-tetrazin-4(3H)-one
(CCRG 81045; M & B 39831), a novel drug with potential as an
alternative to dacarbazine. Cancer Res. 47:5846–5852.
1987.PubMed/NCBI
|
|
3
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
MoreiraGonzalez A, Djohan R and Lohman R:
Considerations surrounding reconstruction after resection of
musculoskeletal sarcomas. Cleve Clin J Med. 77 (Suppl 1):S18–S22.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Trent JC, Beach J, Burgess MA, et al: A
two-arm phase II study of temozolomide in patients with advanced
gastrointestinal stromal tumors and other soft tissue sarcomas.
Cancer. 98:2693–2699. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Garcia del Muro X, Lopez-Pousa A, Martin
J, Buesa JM, Martinez-Trufero J, Casado A, Poveda A, Cruz J, Bover
I and Maurel J: Spanish Group for Research on Sarcomas: A phase II
trial of temozolomide as a 6-week, continuous, oral schedule in
patients with advanced soft tissue sarcoma: A study by the Spanish
Group for Research on Sarcomas. Cancer. 104:1706–1712. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stacchiotti S, Tortoreto M, Bozzi F, et
al: Dacarbazine in solitary fibrous tumor: A case series analysis
and preclinical evidence vis-a-vis temozolomide and
antiangiogenics. Clin Cancer Res. 19:5192–5201. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Casey DA, Wexler LH, Merchant MS, Chou AJ,
Merola PR, Price AP and Meyers PA: Irinotecan and temozolomide for
Ewing sarcoma: The Memorial Sloan-Kettering experience. Pediatr
Blood Cancer. 53:1029–1034. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Raciborska A, Bilska K, Drabko K, Chaber
R, Pogorzala M, Wyrobek E, Polczyńska K, Rogowska E,
RodriguezGalindo C and Wozniak W: Vincristine, irinotecan and
temozolomide in patients with relapsed and refractory Ewing
sarcoma. Pediatr Blood Cancer. 60:1621–1625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hotta T, Motoyama T and Watanabe H: Three
human osteosarcoma cell lines exhibiting different phenotypic
expressions. Acta Pathol Jpn. 42:595–603. 1992.PubMed/NCBI
|
|
11
|
Imaizumi S, Motoyama T, Ogose A, Hotta T
and Takahashi HE: Characterization and chemosensitivity of two
human malignant peripheral nerve sheath tumour cell lines derived
from a patient with neurofibromatosis type 1. Virchows Arch.
433:435–441. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hoshino M, Kawashima H, Ogose A, et al:
Serum CA 125 expression as a tumor marker for diagnosis and
monitoring the clinical course of epithelioid sarcoma. J Cancer Res
Clin Oncol. 136:457–464. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sonobe H, Manabe Y, Furihata M, Iwata J,
Oka T, Ohtsuki Y, Mizobuchi H, Yamamoto H, Kumano O and Abe S:
Establishment and characterization of a new human synovial sarcoma
cell line, HS-SY-II. Lab Invest. 67:498–505. 1992.PubMed/NCBI
|
|
14
|
Kawai A, Naito N, Yoshida A, et al:
Establishment and characterization of a biphasic synovial sarcoma
cell line, SYO-1. Cancer Lett. 204:105–113. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aman P, Ron D, Mandahl N, et al:
Rearrangement of the transcription factor gene CHOP in myxoid
liposarcomas with t(12;16)(q13;p11). Genes Chromosomes Cancer.
5:278–285. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hoshino M, Ogose A, Kawashima H, et al:
Molecular analyses of cell origin and detection of circulating
tumor cells in the peripheral blood in alveolar soft part sarcoma.
Cancer Genet Cytogenet. 190:75–80. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kusakabe H, Iwasaki H, Sano K and Kiyokane
K: Expression of lung resistance protein in epithelioid sarcoma in
vitro and in vivo. Arch Dermatol Res. 292:292–300. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
FernandesAlnemri T, Litwack G and Alnemri
ES: CPP32, a novel human apoptotic protein with homology to
Caenorhabditis elegans cell death protein Ced-3 and mammalian
interleukin-1 beta-converting enzyme. J Biol Chem. 269:30761–30764.
1994.PubMed/NCBI
|
|
19
|
Oliver FJ, de la Rubia G, Rolli V, et al:
Importance of poly (ADP-ribose) polymerase and its cleavage in
apoptosis. Lesson from an uncleavable mutant. J Biol Chem.
273:33533–33539. 1998.PubMed/NCBI
|
|
20
|
Liu H, Zhou L, Shi S, et al:
Oligosaccharide G19 inhibits U-87 MG human glioma cells growth in
vitro and in vivo by targeting epidermal growth factor (EGF) and
activating p53/p21 signaling. Glycobiology. 24:748–765. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu L, Yang L, Xiong Y, et al: Annexin A5
promotes invasion and chemoresistance to temozolomide in
glioblastoma multiforme cells. Tumour Biol. 35:12327–12337. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hegi ME, Diserens AC, Gorlia T, Hamou MF,
de Tribolet N, Weller M, Kros JM, Hainfellner JA, Mason W, Mariani
L, et al: MGMT gene silencing and benefit from temozolomide in
glioblastoma. N Engl J Med. 352:997–1003. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jakob J, Hille M, Sauer C, Ströbel P, Wenz
F and Hohenberger P: O6-methylguanine-DNA methyltransferase (MGMT)
promoter methylation is a rare event in soft tissue sarcoma. Radiat
Oncol. 7:1802012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
DeSalvo M, Maresca G, D'agnano I, et al:
Temozolomide induced c-Myc-mediated apoptosis via Akt signalling in
MGMT expressing glioblastoma cells. Int J Radiat Biol. 87:518–533.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang H, Cai S, Ernstberger A, Bailey BJ,
Wang MZ, Cai W, Goebel WS, Czader MB, Crean C, Suvannasankha A, et
al: Temozolomide-mediated DNA methylation in human myeloid
precursor cells: Differential involvement of intrinsic and
extrinsic apoptotic pathways. Clin Cancer Res. 19:2699–2709. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kanzawa T, Germano IM, Kondo Y, Ito H, Kyo
S and Kondo S: Inhibition of telomerase activity in malignant
glioma cells correlates with their sensitivity to temozolomide. Br
J Cancer. 89:922–929. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Roos WP, Batista LF, Naumann SC, Wick W,
Weller M, Menck CF and Kaina B: Apoptosis in malignant glioma cells
triggered by the temozolomide-induced DNA lesion
O6-methylguanine. Oncogene. 26:186–197. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hirose Y, Berger MS and Pieper RO: p53
effects both the duration of G2/M arrest and the fate of
temozolomide-treated human glioblastoma cells. Cancer Res.
61:1957–1963. 2001.PubMed/NCBI
|
|
29
|
Kanzawa T, Germano IM, Komata T, Ito H,
Kondo Y and Kondo S: Role of autophagy in temozolomide-induced
cytotoxicity for malignant glioma cells. Cell Death Differ.
11:448–457. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mahalingam D, Mita A, Sankhala K, Swords
R, Kelly K, Giles F and Mita MM: Targeting sarcomas: Novel
biological agents and future perspectives. Curr Drug Targets.
10:937–949. 2009. View Article : Google Scholar : PubMed/NCBI
|















