Introduction
Previously, studies have suggested that the
chaperone heat shock protein 90 (Hsp90) is a promising target for
cancer therapy (1–4). According to previous studies, the
constitutive expression of Hsp90 was found to be 2–10 folds higher
in malignant cells compared with normal cells (5). Additionally, several oncogenic proteins
are dependent on Hsp90 to function, including B-RAF, protein kinase
B (AKT) and extracellular signal-related kinase (ERK) (6). Inhibition of Hsp90 results in the
dissociation of corresponding client proteins and induction of
apoptosis in cancer cells, and may be accompanied by a reduction in
chemotherapy resistance (7). Overall,
these results highlight the crucial role of Hsp90 in the survival
and growth of cancer cells and hold considerable potential for the
development of small molecular inhibitors in drug treatment.
As an essential chaperone in vivo, Hsp90
assists a vast variety of client proteins, which may be categorized
into three groups: i) Protein kinases, including B-RAF, AKT and
ERK; ii) transcriptional factors or polymerases; and iii)
miscellaneous groups (8). Numerous
Hsp90-associated proteins are important in the occurrence and
development of cancer. By forming complexes with Hsp90, RAF is
stabilized, consequently inhibiting proteasome-dependent protein
degradation (9). Disruption of Hsp90
function may significantly interfere with the RAF signaling pathway
(10). In addition, AKT pathways
participate in the regulation of cellular proliferation and
apoptosis (11,12). Previous studies have suggested that
the occupancy of an Hsp90 binding site by ansamycins can result in
a reduced half-life of AKT in vivo (8,13).
Similarly, ERK is closely involved in the pathogenesis of cancer
and neurological disease (14,15).
Previous studies indicate that Hsp90 inhibitors may reduce the
phosphorylation of ERK, without affecting the expression at mRNA
and protein levels (16,17). These oncogenic protein kinases
contribute to cell proliferation, which is associated with
apoptosis induced by Hsp90 inhibition.
TNF-related apoptosis-inducing ligand (TRAIL) is a
cytokine produced and secreted by the majority of normal tissue
cells (18). By binding to the death
receptors DR4 and DR5, TRAIL causes apoptosis in cancer cells. For
this reason, TRAIL has been an anti-cancer therapy target since the
mid-1990s; however, no significant survival benefits have been
found. Previous studies have reported that TRAIL can bind to and
therefore activate NF-κB, leading to the transcription of genes
known to antagonize the death signaling pathway (19). Studies have also indicated that
activation of the AKT survival pathway contributes to TRAIL
resistance in cancer cells (20).
These results suggest that the combination of oncogenic kinase
inhibitors, such as AKT and TRAIL demonstrate promise in the
induction of apoptosis.
Gastric cancer is the second most common cause of
cancer-associated mortality. More importantly, gastric cancer is
characterized by extremely low survival rates among patients
(21). Numerous studies have
suggested the importance of Hsp90 in the progression of gastric
cancer (22–24). Certain small molecular inhibitors of
Hsp90, including MPC-3100 (Myriad Pharmaceuticals, Salt Lake City,
UT, USA), have demonstrated potential in the treatment of gastric
cancer (25). Geldanamycin (GA) is a
naturally occurring benzoquinone ansamycin that specifically binds
to and inhibits Hsp90 (26). GA
inhibits proliferation and induces apoptosis in certain types of
cancer, including leukemia, colon cancer and breast cancer
(27). However, the effect of GA on
gastric cancer is unclear. In the present study, a systematic
evaluation of GA in the treatment of gastric cancer was performed
and the molecular mechanism behind Hsp90 inhibition was
investigated. GA was found to significantly decrease the expression
and phosphorylation of numerous oncogenic protein kinases,
including AKT, which holds promise as a drug treatment, when used
in combination with apoptosis-inducing TRAIL.
Materials and methods
Cell line and reagents
The human gastric cancer cell line SGC-7901 was
purchased from Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). Monoclonal mouse anti-human antibodies against
phosphorylated-AKT (p-AKT; 100 µl; cat no. 5106S) and
phosphorylated-ERK (p-ERK; 200 µl; cat no. 9106S) were purchased
from Cell Signaling Technology (Beverly, MA, USA). Annexin
V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit was
purchased from Sigma-Aldrich (St. Louis, MO, USA). In-Situ
Cell Death Detection kit was obtained from Roche (Mannheim,
Germany). Dulbeccos modified Eagles medium (DMEM) and fetal bovine
serum (FBS) were purchased from Thermo Fisher Scientific (Waltham,
MA, USA).
Cell culture of the SGC-7901 cancer
cell line
Human gastric cancer SGC-7901 cells were cultured in
DMEM containing 10% FBS, 1% glutathione (Sigma-Aldrich) and 1%
penicillin/streptomycin (Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA), at 37°C in a humidified atmosphere containing 5%
CO2. For all experiments, the cells were used in the
exponential growth phase.
In vitro migration and invasion
assays
An in vitro cell migration assay was
performed with Millicell chambers (EMD Millipore, Bedford, MA,
USA), as previously described (28).
Briefly, the SGC-7901 cells were treated with 0.4 M of GA for 48 h,
then harvested and suspended with serum-free medium. Then, 200 µl
of cell suspension (2.5×105 cells/ml) was added to the
upper chamber and 500 µl NIH3T3-conditioned medium (Gibco; Thermo
Fisher Scientific, Waltham, MA, USA) was added to the lower
compartment. The chamber was incubated in 5% CO2 at 37°C
for 48 h. The cells were removed from the upper chamber and the
attached cells in the lower section were stained with 0.1% crystal
violet. A similar procedure was performed for the invasion assay,
but Matrigel (Clontech Laboratories, Inc., Mountain View, CA, USA)
was added to the Transwell chambers (EMD Millipore) (28).
Terminal deoxynucleotidyl transferase
mediated digoxigenin-dUTP nick-end labeling (TUNEL) assay
Cells were seeded in 6-well plates
(1.5×105 cells/well) after placing a coverslip in each
well. Apoptotic cells were detected using a TUNEL assay using the
in situ cell death assay kit, according to the
manufacturer's protocol. A BX53 microscope (Olympus, Tokyo, Japan)
was used to count the total cell number and TUNEL-positive cell
number in five fields of each TUNEL-stained section. The apoptotic
index was calculated as the mean percentage of cells with definite
positive TUNEL staining in the five microscopic fields.
Annexin V/PI staining assay
To evaluate the early stage of apoptosis induced by
GA, an Annexin V/PI staining assay was performed. Subsequent to
treatment with different concentrations of inhibitor for 48 h, the
SGC-7901 cells were harvested and washed three times with PBS. The
cells were then stained with Annexin V/PI staining kit, as per the
manufacturer's instructions. Flow cytometric analysis was
immediately performed with a FACSCalibur Instrument (Becton
Dickinson, Franklin Lakes, NJ, USA).
Immunocytochemistry assay
The SGC-7901 cells were treated with various
concentrations of GA for 48 h. The phosphorylation of AKT and ERK
kinases was then evaluated by immunocytochemistry assay, as
previously described (29).
Statistical analysis
The data were presented as the mean ± standard
deviation based on three independent experiments and statistically
analyzed using a t-test or χ2 test. Analyses were
performed using SPSS-13.0 statistical software (SPSS Inc., Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant different.
Results
GA inhibits proliferation, migration
and invasion of SGC-7901 cancer cells
By forming protein complexes, Hsp90 assists in
maintaining the conformation, stability and function of certain
oncogenic client proteins. Therefore, Hsp90 is important in cell
proliferation, cell cycle progression, apoptosis and other features
of malignant phenotypes, such as invasion, angiogenesis and
metastasis (2,4). To evaluate the potency of GA on gastric
cancer, the effect of GA on the proliferation, migration and
invasion of the gastric SGC-7901 cancer cell line was determined.
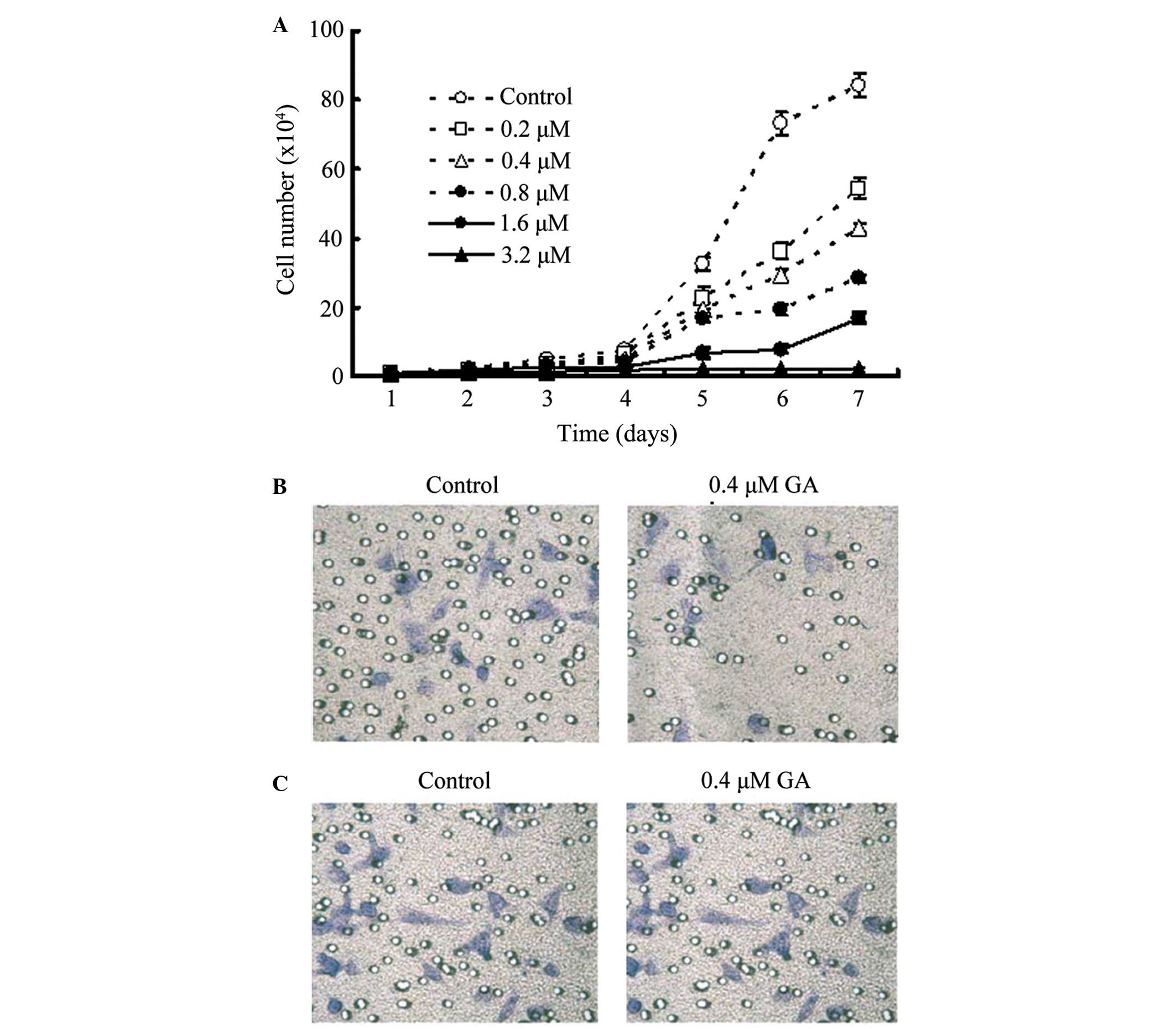
The time taken for GA-induced proliferation inhibition is shown in
Fig. 1A. GA significantly inhibits
the proliferation of SGC-7901 cancer cells even at a low
concentration of GA (0.2 M). The effect of GA on migration was
studied using a Transwell assay. In this method, the cells that
migrated through the pored membrane under the induction of
NIH3T3-conditioned medium in the lower compartment were fixed and
stained with crystal violet. The results revealed that treatment
with 0.4 M GA for 48 h significantly inhibits the migration of
SGC-7901 cells (Fig. 1B). The effect
on invasion was also evaluated using a Matrigel invasion assay,
where cells invade the Matrigel with the induction of
NIH3T3-conditioned medium. Following incubation at 37°C for 48 h,
the cells invading the surface of the filter were fixed and stained
using crystal violet. The treatment with 0.4 M GA was found to
significantly inhibit the invasion of SGC-7901 cells (Fig. 1C).
GA induces apoptosis of SGC-7901
cancer cells
The effect of GA on SGC-7901 cells was evaluated
using TUNEL assay and Annexin V-FITC/PI staining assays. Cells
undergoing apoptosis were indicated by brown granules in the
nucleus. Treatment with 0.4 M GA induced apoptosis in SGC-7901
cancer cells compared with control cells (Fig. 2A). Similar results were observed by
Annexin V-FITC/PI staining. GA significantly increased the
percentage of apoptosis in SGC-7901 cancer cells at various
concentrations (0.2 and 0.4 M) compared with control cells
(P<0.001) (Fig. 2B).
GA treatment decreases the expression
of B-RAF
Most frequently found in melanoma and other types of
cancer, mutation in B-RAF may cause B-RAF activation, thus
facilitating its binding to the Hsp90-cdc37 complex that is
essential for stability as well as function. Hsp90 inhibitors, such
as 17-N-allylamino-17-demethoxygeldanamycin (17-AAG), stimulate
B-RAF degradation in cancer cells (30,31). When
the B-RAF mRNA levels were measured using RT-PCR, it was found that
treatment with GA (0.4 M) for 48 h induced a significant reduction
of B-RAF mRNA in SGC-7901 cells (Fig.
3).
GA treatment decreases the
phosphorylation of AKT and ERK
AKT and ERK can each promote cell survival and
growth, as well as suppress cell apoptosis. The two kinases are
considered to be involved in apoptosis induced by the inhibition of
Hsp90. In the present study, the phosphorylation levels of two
protein targets in GA-treated SGC-7901 cancer cells were determined
with immunocytochemistry assays. Treatment with GA (0.2 and 0.4 M)
significantly decreases the phosphorylation of AKT and ERK
(Fig. 4). This is a notable result
regarding the well-established role of activated AKT in the
resistance of TRAIL in cancer cells, and holds great promise for
the combination therapy of GA and TRAIL in gastric cancer
chemotherapy.
Discussion
The molecular chaperone Hsp90 has been
well-established as an important target of anticancer therapies.
Hsp90 may stabilize and regulate the function of numerous client
proteins, including kinases involved in cancer development
(2,32). The anticancer potential of chemical
inhibitors of Hsp90, including GA and 17-AAG, has been extensively
and comprehensively validated with cell models, xenografts and
clinical studies (6,27,30). The
results of the present study revealed that GA strongly inhibits the
proliferation of gastric cancer SGC-7901 cells. In addition, GA
significantly increases the apoptosis of SGC-7901 cells, which
supports the anticancer potential of GA.
As protein clients of Hsp90, B-RAF, AKT and ERK
kinases are regarded to be involved in apoptosis induced by Hsp90
inhibitors (33,34). The expression of B-RAF and the
phosphorylation of AKT and ERK were determined. As a result, mRNA
levels of B-RAF were found to be largely reduced in the GA-treated
SGC-7901 cancer cells compared with control cells. By contrast, the
phosphorylation of AKT and ERK significantly decreased in
GA-treated SGC-7901 cells at various concentrations. These results
suggest that the inhibition of oncogenic signaling pathways, such
as the PI3K-AKT and Ras-RAF-MEK-ERK pathways, may contribute to
apoptosis induced by GA in SGC-7901 cells. The present study
provides a potential mechanism for the drug treatment of gastric
cancer with GA. As previously mentioned, numerous studies have
highlighted the inhibitory role of AKT in TRAIL signaling (20,35).
Down-regulation of constitutively active AKT may lead to the
resistance of cancer cells to recombinant human (rh)TRAIL. During
this process, Hsp90 protects cancer cells from apoptosis by
stabilizing AKT and NF-κB. Therefore, inhibition of Hsp90 may
result in cell cycle arrest and more importantly, apoptosis
(36,37). The use of 17-AAG as an Hsp90 inhibitor
is currently being clinically evaluated in phase I trials (38,39).
In conclusion, combinations of Hsp90 inhibitors with
TRAIL inducers exhibited synergistic effects on apoptosis induction
in gastric cancer. Thus, the present study provides a rationale for
the combined use of GA and TRAIL in the clinic.
Acknowledgements
The present study was supported by the National Key
Clinical Specialist Construction Project of China.
References
|
1
|
Solit DB and Rosen N: Hsp90: A novel
target for cancer therapy. Curr Top Med Chem. 6:1205–1214. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maloney A and Workman P: HSP90 as a new
therapeutic target for cancer therapy: The story unfolds. Expert
Opin Biol Ther. 2:3–24. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mahalingam D, Swords R, Carew JS, Nawrocki
ST, Bhalla K and Giles FJ: Targeting HSP90 for cancer therapy. Br J
Cancer. 100:1523–1529. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sidera K and Patsavoudi E: HSP90
inhibitors: Current development and potential in cancer therapy.
Recent Pat Anticancer Drug Discov. 9:1–20. 2014.PubMed/NCBI
|
|
5
|
Ferrarini M, Heltai S, Zocchi MR and
Rugarli C: Unusual expression and localization of heat-shock
proteins in human tumor cells. Int J Cancer. 51:613–619. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Neckers L and Neckers K: Heat-shock
protein 90 inhibitors as novel cancer chemotherapeutic agents.
Expert Opin Emerg Drugs. 7:277–288. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Connell P, Ballinger CA, Jiang J, Wu Y,
Thompson LJ, Höhfeld J and Patterson C: The co-chaperone CHIP
regulates protein triage decisions mediated by heat-shock proteins.
Nat Cell Biol. 3:93–96. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goetz MP, Toft DO, Ames MM and Erlichman
C: The Hsp90 chaperone complex as a novel target for cancer
therapy. Ann Oncol. 14:1169–1176. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schulte TW, Blagosklonny MV, Ingui C and
Neckers L: Disruption of the Raf-1-Hsp90 molecular complex results
in destabilization of Raf-1 and loss of Raf-1-Ras association. J
Biol Chem. 270:24585–24588. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schulte TW, Blagosklonny MV, Romanova L,
Mushinski JF, Monia BP, Johnston JF, Nguyen P, Trepel J and Neckers
LM: Destabilization of Raf-1 by geldanamycin leads to disruption of
the Raf-1-MEK-mitogen-activated protein kinase signalling pathway.
Mol Cell Biol. 16:5839–5845. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Neri LM, Borgatti P, Capitani S and
Martelli AM: The nuclear phosphoinositide 3-kinase/AKT pathway: A
new second messenger system. Biochim Biophys Acta. 1584:73–80.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kumar A, Rajendran V, Sethumadhavan R and
Purohit R: AKT kinase pathway: A leading target in cancer research.
ScientificWorldJournal. 2013:7561342013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Basso AD, Solit DB, Chiosis G, Giri B,
Tsichlis P and Rosen N: Akt forms an intracellular complex with
heat shock protein 90 (Hsp90) and Cdc37 and is destabilized by
inhibitors of Hsp90 function. J Biol Chem. 277:39858–39866. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma W and Quirion R: The ERK/MAPK pathway,
as a target for the treatment of neuropathic pain. Expert Opin Ther
Targets. 9:699–713. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee JT Jr and McCubrey JA: The Raf/MEK/ERK
signal transduction cascade as a target for chemotherapeutic
intervention in leukemia. Leukemia. 16:486–507. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dou F, Yuan LD and Zhu JJ: Heat shock
protein 90 indirectly regulates ERK activity by affecting Raf
protein metabolism. Acta Biochim Biophys Sin (Shanghai).
37:501–505. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Johnstone RW, Frew AJ and Smyth MJ: The
TRAIL apoptotic pathway in cancer onset, progression and therapy.
Nat Rev Cancer. 8:782–798. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wiley SR, Schooley K, Smolak PJ, Din WS,
Huang CP, Nicholl JK, Sutherland GR, Smith TD, Rauch C, Smith CA,
et al: Identification and characterization of a new member of the
TNF family that induces apoptosis. Immunity. 3:673–682. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Song JJ and Lee YJ: Differential cleavage
of Mst1 by caspase-7/-3 is responsible for TRAIL-induced activation
of the MAPK superfamily. Cell Signal. 20:892–906. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu J, Zhou JY, Wei WZ and Wu GS:
Activation of the Akt survival pathway contributes to TRAIL
resistance in cancer cells. PloS One. 5:e102262010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Roder DM: The epidemiology of gastric
cancer. Gastric Cancer. 5(Suppl): 5–11. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen M, Xu J and Zhao J: Effects of HSP90
inhibitor 17-AAG on cell cycle and apoptosis of human gastric
cancer cell lines SGC-7901. Nan Fang Yi Ke Da Xue Xue Bao.
33:271–275. 2013.(In Chinese). PubMed/NCBI
|
|
23
|
Wainberg ZA, Anghel A, Rogers AM, Desai
AJ, Kalous O, Conklin D, Ayala R, O'Brien NA, Quadt C, Akimov M, et
al: Inhibition of HSP90 with AUY922 induces synergy in
HER2-amplified trastuzumab-resistant breast and gastric cancer. Mol
Cancer Ther. 12:509–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Redlak MJ and Miller TA: Targeting
PI3K/Akt/HSP90 signaling sensitizes gastric cancer cells to
deoxycholate-induced apoptosis. Dig Dis Sci. 56:323–329. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hede K: Gastric cancer: Trastuzumab trial
results spur search for other targets. J Natl Cancer Inst.
101:1306–1307. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Stebbins CE, Russo AA, Schneider C, Rosen
N, Hartl FU and Pavletich NP: Crystal structure of an
Hsp90-geldanamycin complex: Targeting of a protein chaperone by an
antitumor agent. Cell. 89:239–250. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Supko JG, Hickman RL, Grever MR and
Malspeis L: Preclinical pharmacologic evaluation of geldanamycin as
an antitumor agent. Cancer Chemother Pharmacol. 36:305–315. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma Q, Jiang Q, Pu Q, Zhang X, Yang W, Wang
Y, Ye S, Wu S, Zhong G, Ren J, et al: MicroRNA-143 inhibits
migration and invasion of human non-small-cell lung cancer and its
relative mechanism. Int J Biol Sci. 9:680–692. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Köhrmann A, Kammerer U, Kapp M, Dietl J
and Anacker J: Expression of matrix metalloproteinases (MMPs) in
primary human breast cancer and breast cancer cell lines: New
findings and review of the literature. BMC Cancer. 16:1882009.
View Article : Google Scholar
|
|
30
|
da Rocha Dias S, Friedlos F, Light Y,
Springer C, Workman P and Marais R: Activated B-RAF is an Hsp90
client protein that is targeted by the anticancer drug
17-allylamino-17-demethoxygeldanamycin. Cancer Res. 65:10686–10691.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Grbovic OM, Basso AD, Sawai A, Ye Q,
Friedlander P, Solit D and Rosen N: V600E B-Raf requires the Hsp90
chaperone for stability and is degraded in response to Hsp90
inhibitors. Proc Natl Acad Sci USA. 103:57–62. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Neckers L and Neckers K: Heat-shock
protein 90 inhibitors as novel cancer chemotherapeutics - an
update. Expert Opin Emerg Drugs. 10:137–149. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang H and Burrows F: Targeting multiple
signal transduction pathways through inhibition of Hsp90. J Mol Med
(Berl). 82:488–499. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koga F, Xu WP, Karpova TS, McNally JG,
Baron R and Neckers L: Hsp90 inhibition transiently activates Src
kinase and promotes Src-dependent Akt and Erk activation. Proc Natl
Acad Sci USA. 103:11318–11322. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Matsuoka T and Yashiro M: The role of
PI3K/Akt/mTOR signaling in gastric carcinoma. Cancers (Basel).
6:1441–1463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hostein I, Robertson D, DiStefano F,
Workman P and Clarke PA: Inhibition of signal transduction by the
Hsp90 inhibitor 17-Allylamino-17- demethoxygeldanamycin results in
cytostasis and apoptosis. Cancer Res. 61:4003–4009. 2001.PubMed/NCBI
|
|
37
|
Powers MV, Clarke PA and Workman P: Dual
targeting of HSC70 and HSP72 inhibits HSP90 function and induces
tumor-specific apoptosis. Cancer Cell. 14:250–262. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim YS, Alarcon SV, Lee S, Lee MJ,
Giaccone G, Neckers L and Trepel JB: Update on Hsp90 inhibitors in
clinical trial. Curr Top Med Chem. 9:1479–1492. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jhaveri K, Taldone T, Modi S and Chiosis
G: Advances in the clinical development of heat shock protein 90
(Hsp90) inhibitors in cancers. Biochim Biophys Acta. 1823:742–755.
2012. View Article : Google Scholar : PubMed/NCBI
|