Introduction
Cervical cancer is the third most common malignancy
in females globally, accounting for 8.8% of all cases of cancer.
Furthermore, in 2008, ~529,000 new cases and ~274,000 mortalities
due to cervical cancer were reported worldwide (1). Approximately 80% of these cases occurred
in individuals in developing countries (2).
Cervical cancer occurs in a multi-step process that
involves the transformation of healthy cervical epithelium into
preneoplastic cervical intraepithelial neoplasia (CIN) that
eventually evolve into invasive cervical cancer cells (3,4). It is
well recognized that specific oncogenic human papilloma virus (HPV)
is a primary etiological factor in cervical cancer (5,6). However,
only a small fraction of HPV-infected lesions progress to cervical
cancer or its precursor lesion, cervical intraepithelial neoplasia
(CIN), indicating that other genetic factors, including
tumor-suppressor genes, proto-oncogenes and immunological factors,
may be involved in the development of cervical cancer (7–9).
Girdin is a multi-functional protein with the
ability to promote cell proliferation and migration (10–12).
Girdin is located at the crossroad of G protein signaling and
tyrosine kinase receptor signaling and, therefore, promotes the
activation of a series of molecules, including G protein, Akt and
signal transducer and activator of transcription (STAT3) (13,14). An
increasing number of studies have demonstrated that Girdin is
preferentially expressed in various malignant tissues, such as
breast, colon and lung cancer, and glioblastoma (10,13,15–17).
In specific types of tumor, for example glioblastoma, the
expression of Girdin increases with the evolution of tumor
progression and invasion (17).
Furthermore, increased expression of Girdin appears to facilitate
cell migration, and concomitant metastasis in breast and colon
cancers (10,16). Therefore, it has been suggested that
the expression of Girdin may be predictive of outcome in breast and
colorectal carcinoma (16,18). However, the expression and function of
Girdin in cervical carcinoma has yet to be investigated.
In the current study, the association between Girdin
expression, and the various stages of intraepithelial neoplasia and
squamous cell carcinoma was investigated. Furthermore, a
short-hairpin RNA (shRNA) approach was used to selectively knock
down the expression of Girdin mRNA in the cervical HeLa tumor cell
line, and investigate its role in cell growth and apoptosis, two
key characteristics in disease progression.
Materials and methods
Patient specimens
A total of 87 fresh cervical tissues were obtained
from patients prior to undergoing chemo- or radiotherapy at Beijing
Hospital, Ministry of Health (Beijing, China) between January 2010
and December 2012. The mean age of the 87 patients in the present
cohort was 39.4 years (range, 23–76 years). The samples comprised
66 invasive lesions, nine CIN I lesions and 12 CIN III lesions
according to the International Federation of Gynecology and
Obstetrics (19). In addition, five
healthy cervical tissue biopsies were used as controls. The
invasive lesion samples were obtained during radical hysterectomy
and the CIN samples were obtained during cervical cone biopsy. The
diagnostic criteria used were: Invasive lesions were defined as
those that broke through the basement membrane and infiltrated to a
depth of >5 mm; samples with cells representing atypical
hyperplasia confined to the superficial one-third of the cervical
epithelia were diagnosed as CIN I; and lesions involved in the full
epithelia demonstrating no invasiveness were diagnosed as CIN
III.
The present study was approved by the Ethics
Committee of the Beijing Hospital (Beijing, China) and adhered to
the tenets of the Declaration of Helsinki. Written informed consent
was obtained from all the donors.
Immunohistochemistry
Endogenous expression of Girdin protein was
evaluated on 4-µm, paraffin-embedded, serial sections of cervical
tissues using rabbit polyclonal IgG anti-human Girdin antibody
(#sc-133371; Santa Cruz Biotechnology, Inc.) or control IgG, as
previously described (10).
Immunohistochemisty was performed using standard techniques.
Antigen retrieval was performed with microwave treatment in a 0.01
mol/l citrate buffer (pH 6.0) at 95°C for 10 min. Girdin
immunostaining was examined by counting ≥500 cells in five random
high power fields from each specimen using a Eclipse 80i light
microscope (Nikon Corporation, Tokyo, Japan), as previously
described (15). After counting,
Girdin expression was semiquantitatively classified according to
the proportion of immunoreactive cancer cells in the cytoplasm, as
follows: 0, <1% of cells; 1+, ≥1 to <10% of cells; 2+, ≥10 to
<50% of cells; and 3+, ≥50% of cells.
Cell culture and proliferation
assay
The human cervical squamous cell carcinoma line HeLa
was maintained in culture medium (DMEM supplemented with 10% fetal
bovine serum). HeLa cells (ScienCell Research Laboratories,
Carlsbad, CA, USA) were transfected with Girdin shRNA (Invitrogen
Life Technologies, Carlsbad, CA, USA) or control shRNA, as
previously described by Jiang et al (10), and then seeded (2×104
cells) in 35-mm Petri dishes. After 24, 48 and 72 h incubation in
culture medium, the cells were trypsinized, stained with 0.4%
trypan blue (Yi San Biotechnology Co., Ltd., Shanghai, China), and
counted using a hemocytometer (Qiujing, Inc, Zhejiang, China).
Apoptosis detection
Cell apoptosis was determined using Hoechst 33342
staining. All the steps were performed according to the
manufacturer's instructions.
Western blot analysis
Samples were separated by 8% SDS-PAGE. Proteins were
transferred to nitrocellulose membranes, blocked in 3% skim milk in
phosphate buffered saline and 0.05% Tween 20, incubated with
primary antibody (polyclonal rabbit IgG anti-human Girdin antibody;
Santa Cruz Biotechnology, Inc.; #sc-133371; dilution, 1:100), and
detected with horseradish peroxidase-conjugated polyclonal goat
anti-rabbit secondary antibody (#P-0448; Dako, Glostrup, Denmark;
dilution, 1:5,000). Immunoreactive bands were visualized using
enhanced chemiluminescence detection reagents (Applygen
Technologies Inc., Beijing, China).
Statistical analysis
Data were analyzed using SPSS statistical software
(version 13.0; SPSS, Inc., Chicago, IL, USA). Differences between
various groups were compared using a χ2 test, Fisher's
exact test or one-way analysis of variance (ANOVA), where
appropriate. The correlation between Girdin protein expression
levels and the grade of lesion was determined by performing a
Spearman's rank correlation test. The intensity of western blotting
bands were analyzed by using Image-Pro Plus software, version 6.0.
The level of Girdin expression and apoptotic cells were analyzed in
triplicate. Furthermore, a t-test was used to detect the difference
in cell migration and invasion. P<0.05 was considered to
indicate a statistically significant difference.
Results
Girdin protein expression in cervical
cancer lesions
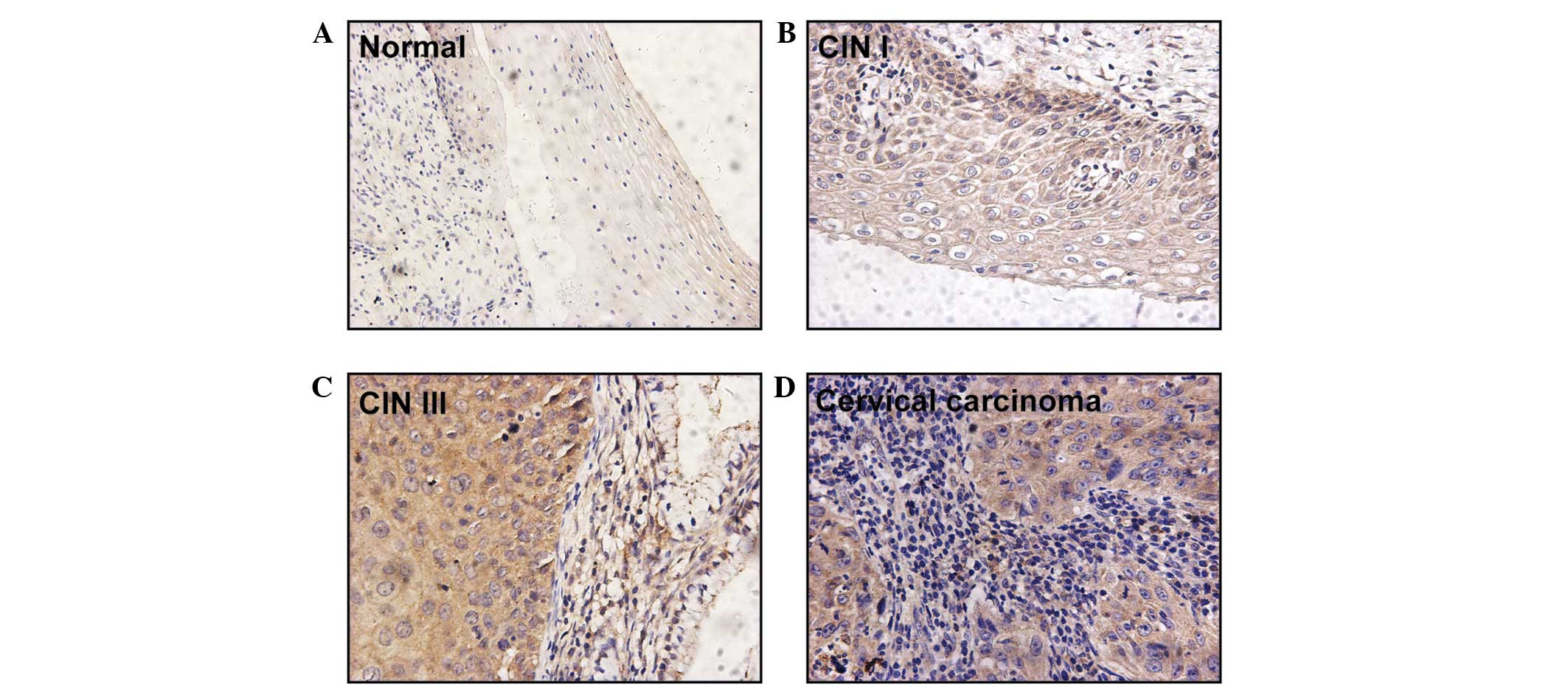
Girdin protein expression was observed in the
cytoplasm of 79/87 (90.8%) cervical cancer lesion specimens.
However, no positive signal was identified in the five healthy
cervical squamous epithelium samples (Table I; Fig.
1). In comparison to the negative staining observed in healthy
cervical epithelial cells (Fig. 1A),
all cases of invasive carcinoma expressed Girdin protein and
positive Girdin protein signals were observed in 13/21 identifiable
pre-malignant lesions (Table I).
Specifically, Girdin was expressed in the basal and parabasal
layers of CIN I samples (n=4; Fig.
1B) and in the superficial layers of CIN III samples (n=9;
Fig. 1C). A staining degree of 3+ was
not observed in any of the CIN cases but was observed in eight
cases of invasive carcinoma. Furthermore, moderate (2+) and intense
(3+) staining for Girdin was detected in one (11.1%) case of CIN I,
three (25.0%) cases of CIN III, and 50 (75.7%) cases of invasive
carcinoma (Table I). Statistical
analysis of the expression of Girdin protein indicated a
significant difference between the three grades of cervical cancer
lesion (one-way ANOVA, F-value=22.976; Spearman's correlation
coefficient, 0.566; P<0.001; data not shown).
 | Table I.Expression of Girdin in 87 cases of
cervical squamous cell carcinoma using immunohistochemistry. |
Table I.
Expression of Girdin in 87 cases of
cervical squamous cell carcinoma using immunohistochemistry.
|
| Degree of Girdin
staining |
|
|---|
|
|
|
|
|---|
| Lesion grade | 0, n (%) | 1+, n (%) | 2+, n (%) | 3+, n (%) | Total patients, n
(%) |
|---|
| Cervical
intraepithelial neoplasia I | 5
(55.6) | 3
(33.3) | 1
(11.1) | 0 (0.0) | 9
(100.0) |
| Cervical
intraepithelial neoplasia III | 3
(25.0) | 6
(50.0) | 3
(25.0) | 0 (0.0) | 12 (100.0) |
| Invasive squamous
cell carcinoma | 0 (0.0) | 16 (24.2) | 42 (63.6) | 8
(12.1) | 66 (100.0) |
Girdin is essential for proliferation
and serum-deprived induced apoptosis of HeLa cells
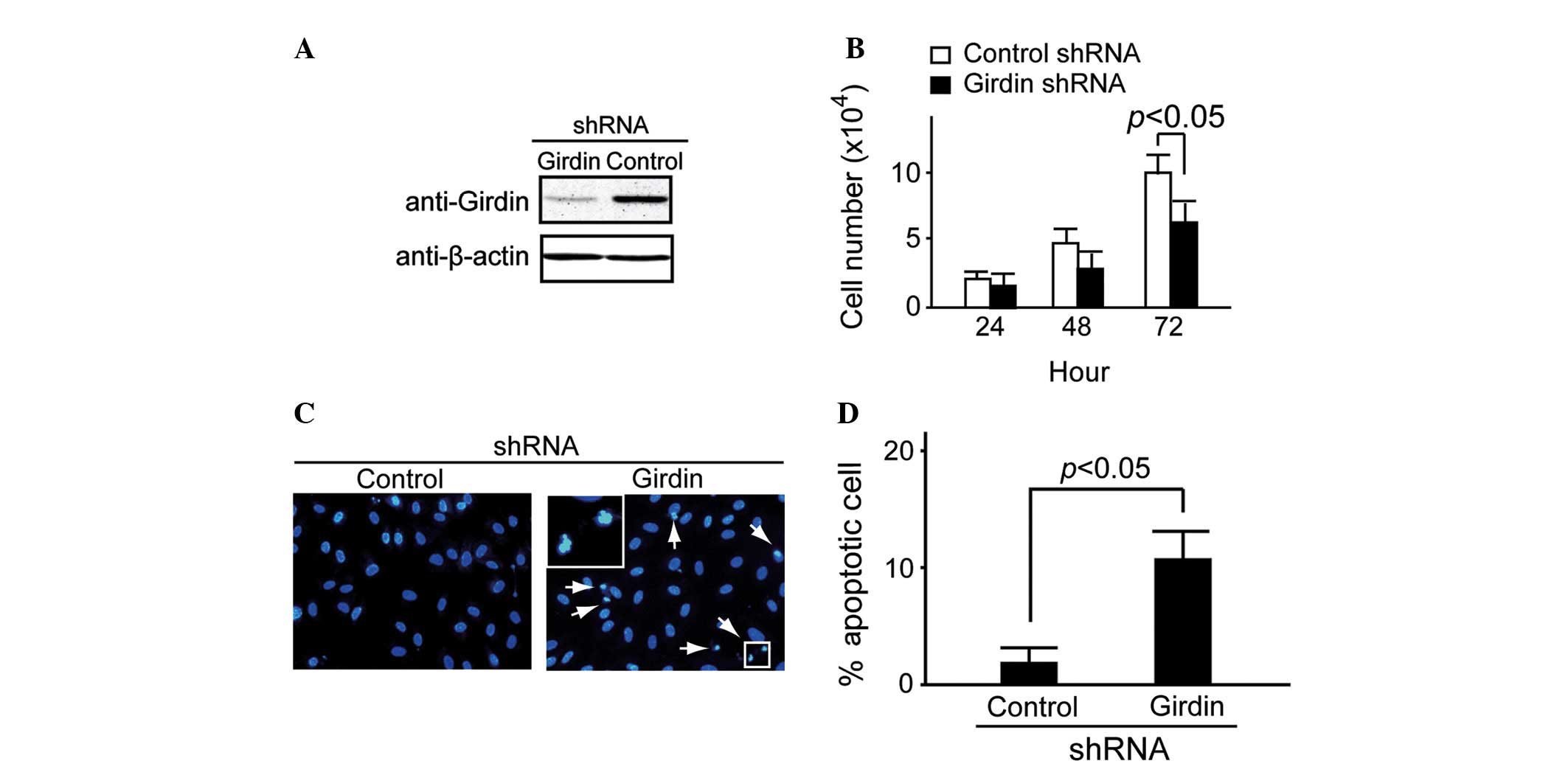
shRNA methodology was used to examine the potential
role of endogenous Girdin expression in HeLa cells, as previously
described by Jiang et al (10). The Girdin-specific shRNA targets and
control shRNA targets were transfected into HeLa cells. After 48 h
of transfection, western blot analysis demonstrated that Girdin
protein expression was suppressed in the Girdin shRNA group
compared with the control (Fig. 2A).
Cell proliferation was detected at 24, 48 and 72 h using trypan
blue staining. It was identified that Girdin shRNA caused
significant attenuation of HeLa cell proliferation at 72 h
(P<0.05; Fig. 2B).
Under normal conditions, almost no apoptosis was
observed in Girdin-suppressed HeLa cells (data not shown). However,
a large number of apoptotic cells was observed in the
Girdin-suppressed HeLa cells when they were cultured in
serum-deprived medium for 48 h. By contrast, the control HeLa cells
exhibited little apoptosis (Fig. 2C).
A significant difference was observed between the proportion of
apoptotic cells in the control and Girdin shRNA groups (P<0.05;
Fig. 2D).
Discussion
Increasing evidence indicates that Girdin promotes
cell motility in epithelial and endothelial cells (10,20). As a
type of actin-binding protein, Girdin is required for lamellipodia
formation as it recruits actin to the leading edge for cell
motility (10). Furthermore, it was
reported that Girdin is essential for cell proliferation (11–13). A
number of reports have demonstrated that Girdin promotes metastasis
in breast carcinoma and colorectal carcinoma (16,18).
Natsume et al (17) recently
identified that Girdin is essential for glioblastoma invasion. The
aforementioned studies indicate that Girdin is pivotal for the
development of epithelium-derived cancer cells. In the present
study, it was identified that Girdin protein is highly expressed in
pre-malignant and malignant cervical carcinoma. Notably, a strong
expression of Girdin was exclusively identified in the invasive
lesions, while a negative or weak expression was detected in the
pre-malignant lesions. In consideration of the current data, it is
suggested that Girdin may promote the progression of cervical
cancer lesions and is important for cancer invasion. This
conclusion is additionally supported by the data from the present
study, which demonstrated that Girdin mRNA is essential for HeLa
cell proliferation.
Immunoexpression of Girdin has been recognized as a
predictor for poor survival in specific malignancies. However, to
the best of our knowledge, its predictive role has yet to be
reported in cervical carcinomas. In the present study,
immunohistochemical analysis was performed to identify Girdin
protein expression in 90.8% of cervical squamous cell lesions. In
addition, the expression of Girdin significantly correlated with
the stage of the lesion. Although the current results indicate that
the expression of Girdin contributes to a poor prognosis, the
mechanism has yet to be sufficiently clarified in cervical
cancer.
Thus far, the role of Girdin in apoptosis remains
unclear. For example, Anai et al (21) reported that COS-7 cells expressing Akt
and Girdin exhibit increased levels of apoptosis and that cells
expressing Girdin alone exhibited little apoptosis. However, in the
present study, it was observed that shRNA-induced depletion of
Girdin induces apoptosis under starvation conditions. Considering
that the Akt signal was inactive under the condition of serum
deprivation, it is proposed that Girdin depletion and Akt
inactivation cooperate to contribute to HeLa cell apoptosis.
However, the detailed mechanisms and physiological significance
require additional investigation.
Aberrant activation of STAT3 has been demonstrated a
dominant association with carcinogenesis, and appears to promote
cell cycle progression, cell proliferation and oncogenic
transformation (22). A recent study
identified that positive staining for phosphorylated STAT3 was
observed in greater than half of the cervical carcinoma cases
(56.8%) investigated, and was significantly correlated with lymph
node metastasis, lymph vascular space invasion and a large tumor
diameter in cervical carcinoma (23).
Notably, Girdin is a direct target of STAT3. Immunohistochemical
analysis of breast carcinoma samples identified a significant
correlation between STAT3 activation and elevated Girdin
expression. Furthermore, Girdin positively autoregulates its own
transcription by enhancing STAT3 activation (14). However, whether a similar interaction
exists between STAT3 and Girdin in cervical lesions merits further
investigation.
In conclusion, the findings of the present study
indicate that Girdin protein is highly expressed in cervical
cancer. Furthermore, Girdin mRNA appears to be essential for cell
proliferation and apoptosis in HeLa cells. Therefore, Girdin may
act as a prospective diagnostic biomarker in cervical cancer
associated with disease stage and tumor grade.
Acknowledgements
The present study was supported by National Natural
Science Foundation of China (grant no. 81072172); the China
Postdoctoral Science Foundation special funding (grant no.
201104043); and the Scientific Research Foundation for the Returned
Overseas Chinese Scholars, State Education Ministry (grant no.
jws1433).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pisani P, Bray F and Parkin DM: Estimates
of the world-wide prevalence of cancer for 25 sites in the adult
population. Int J Cancer. 97:72–81. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Richart RM: A modified terminology for
cervical intraepithelial neoplasia. Obstet Gynecol. 75:131–133.
1990.PubMed/NCBI
|
|
4
|
Creasman WJ: Preinvasive disease of the
cervix. Clinical Gynecologic Oncology. DiSaia PJ and Creasman WJ:
(6th). (CV Mosby, St. Louis, MO). 1–34. 2002. View Article : Google Scholar
|
|
5
|
Basu P and Chowdhury D: Cervical cancer
screening & HPV vaccination: a comprehensive approach to
cervical cancer control. Indian J Med Res. 130:241–246.
2009.PubMed/NCBI
|
|
6
|
Saleem A, Tristram A, Fiander A and
Hibbitts S: Prophylactic HPV vaccination: a major breakthrough in
the fight against cervical cancer? Minerva Med. 100:503–523.
2009.PubMed/NCBI
|
|
7
|
Magnusson PK, Sparén P and Gyllensten UB:
Genetic link to cervical tumours. Nature. 400:29–30. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
zur Hausen H: Papillomaviruses and cancer:
from basic studies to clinical application. Nat Rev Cancer.
2:342–350. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Georgieva S, Iordanov V and Sergieva S:
Nature of cervical cancer and other HPV-associated cancers. J BUON.
14:391–398. 2009.PubMed/NCBI
|
|
10
|
Jiang P, Enomoto A, Jijiwa M, et al: An
actin-binding protein Girdin regulates the motility of breast
cancer cells. Cancer Res. 68:1310–1318. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ghosh P, Beas AO, Bornheimer SJ, et al: A
Gαi-GIV molecular complex binds epidermal growth factor receptor
and determines whether cells migrate or proliferate. Mol Biol Cell.
21:2338–2354. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mao JZ, Jiang P, Cui SP, et al: Girdin
locates in centrosome and midbody and plays an important role in
cell division. Cancer Sci. 103:1780–1787. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Weng L, Enomoto A, Ishida-Takagishi M,
Asai N and Takahashi M: Girding for migratory cues: roles of the
Akt substrate Girdin in cancer progression and angiogenesis. Cancer
Sci. 101:836–842. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dunkel Y, Ong A, Notani D, Mittal Y, Lam
M, Mi X and Ghosh P: STAT3 protein up-regulates Gα-interacting
vesicle-associated protein (GIV)/Girdin expression, and GIV
enhances STAT3 activation in a positive feedback loop during wound
healing and tumor invasion/metastasis. J Biol Chem.
287:41667–41683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ling Y, Jiang P, Cui SP, et al: Clinical
implications for girdin protein expression in breast cancer. Cancer
Invest. 29:405–410. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Garcia-Marcos M, Jung BH, Ear J, Cabrera
B, Carethers JM and Ghosh P: Expression of GIV/girdin, a
metastasis-related protein, predicts patient survival in colon
cancer. FASEB J. 25:590–599. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Natsume A, Kato T, Kinjo S, et al: Girdin
maintains the stemness of glioblastoma stem cells. Oncogene.
31:2715–2724. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu C, Zhang Y, Xu H, Zhang R, Li H, Lu P
and Jin F: Girdin protein: a new potential distant metastasis
predictor of breast cancer. Med Oncol. 29:1554–1560. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Benedet JL, Bender H, Jones H 3rd, Ngan HY
and Pecorelli S: FIGO staging classifications and clinical practice
guidelines in the management of gynecologic cancers. FIGO Committee
on Gynecologic Oncology. Int J Gynaecol Obstet. 70:209–262. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kitamura T, Asai N, Enomoto A, Maeda K,
Kato T, Ishida M, Jiang P, Watanabe T, Usukura J, Kondo T, et al:
Regulation of VEGF-mediated angiogenesis by the Akt/PKB substrate
Girdin. Nat Cell Biol. 10:329–337. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Anai M, Shojima N, Katagiri H, et al: A
novel protein kinase B (PKB)/AKT-binding protein enhances PKB
kinase activity and regulates DNA synthesis. J Biol Chem.
280:18525–18535. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim DJ, Chan KS, Sano S and Digiovanni J:
Signal transducer and activator of transcription 3 (stat3) in
epithelial carcinogenesis. Mol Carcinog. 46:725–731. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takemoto S, Ushijima K, Kawano K, et al:
Expression of activated signal transducer and activator of
transcription-3 predicts poor prognosis in cervical squamous-cell
carcinoma. Br J Cancer. 101:967–972. 2009. View Article : Google Scholar : PubMed/NCBI
|