Introduction
Toll-like receptors (TLRs) comprise an important
family of pattern recognition receptors that result in the
stimulation of innate and adaptive immunity. TLRs form part of the
first line of defense against microbial infection (1). Functional TLRs are expressed not only in
immune cells, but also in certain types of cancer cells (2–5). Activated
TLRs in cancer cells are able to upregulate NF-κB signaling cascade
and induce the expression of anti-apoptotic proteins that lead to
cancer cell proliferation. Furthermore, TLRs can increase the
production of pro-inflammatory cytokines and chemokines that
recruit immune cells to promote an immunosuppressive
microenvironment, induce gene mutation or angiogenesis, and further
contribute to tumor progression (6).
By contrast, functional TLRs can also induce programmed cell death,
strengthening immune surveillance and enhancing chemosensitivity
(7,8).
These results suggest that TLRs may provide dual effects on cancer
cells, and their major roles may depend on tumor origin and TLR
type.
Gastric cancer is the fourth most common type of
cancer in the world and is highly prevalent in Asia; of 988,000
novel cases of gastric cancer, 45% are identified in China
(9,10). The incidence and mortality of gastric
cancer in China are 36.21 and 26.88 per 10 million individuals,
respectively (11). In the past few
decades, the mortality rate of gastric cancer has decreased in
numerous areas of the world, due to the availability of screening
for early detection and improved treatments (12). However, the resistance to chemotherapy
and tumor recurrence continue to make this disease the second
leading cause of cancer-associated mortality worldwide (13). For these reasons, the identification
of novel prognostic factors and development of more effective drugs
has become a new direction in gastric cancer treatment.
Accumulating evidence suggests that the chronic inflammation caused
by microbial infection is an important risk factor correlated with
the aberrant proliferation of gastric cancer cells. TLRs exert a
crucial role in the induction and progression of chronic
inflammation (14). These receptors
recognize conserved molecular patterns expressed in infectious
agents, and mediate the production of pro-inflammatory cytokines
and chemokines. Various TLRs, such as TLR2, TLR4, TLR5 and TLR9,
have been reported to be expressed in gastric cancer cells
(15); however, little is known about
the expression of TLR7 in gastric cancer cells. TLR7 is an
endosomal TLR that is primarily located in intracellular vesicles,
such as endoplasmic reticulum, endosomes, lysosomes and
endolysosomes. Furthermore, the activation of TLR7 has potent
anti-tumor effects by mediating immune activation and programmed
cell death in various types of cancer cell, such as colon cancer
cells (16). However, the role of
TLR7 on gastric cancer cell growth remains unknown. The objective
of the present study was to determine the mRNA and protein
expression levels of TLR7 in gastric cancer tissues, and their
association with the clinicopathological characteristics of gastric
cancer. The effects of TLR7 activation on human gastric cancer
cells were also investigated.
Subjects and methods
Subjects
Patients with gastric cancer (n=30) and healthy
patients without gastric cancer (control; n=14) who underwent
gastroscopy in the Department of Gastroenterology, Second
Affiliated Hospital of Xi'an Jiaotong University (Xi'an, China)
between July 2010 and August 2011 were enrolled in the present
study. The basal characteristics of the two groups are summarized
in Table I. No patient received
pre-operative radiotherapy or chemotherapy. The gastric cancer and
cancer-adjacent tissues were obtained from patients with gastric
cancer, and the normal gastric epithelial tissues were obtained
from the control patients. Ethical approval for tissue use in the
present study was obtained from the Human Subjects Committee of
Xi'an Jiaotong University and written informed consent was obtained
from all subjects. All specimens were immediately frozen in liquid
nitrogen for at least 1 h and then stored at a temperature of −80°C
until use. No significant difference in gender, age, smoking,
drinking or Helicobacter pylori infection was observed
between the gastric cancer and control patients.
 | Table I.Basal characteristics of patients with
gastric cancer (n=30) and control patients (n=14). |
Table I.
Basal characteristics of patients with
gastric cancer (n=30) and control patients (n=14).
|
| Gastric cancer
patients | Control patients |
|
|---|
|
|
|
|
|
|---|
| Characteristic | n | % | n | % | P-value |
|---|
| Gender |
|
|
|
| 0.583 |
| Male | 22 | 73.3 | 10 | 71.4 |
|
|
Female | 8 | 26.7 | 4 | 28.6 |
|
| Age |
|
|
|
| 0.315 |
| ≥60
years | 13 | 43.3 | 7 | 50.0 |
|
| <60
years | 17 | 56.7 | 7 | 50.0 |
|
| Smoking |
|
|
|
| 0.537 |
| Yes | 14 | 46.7 | 6 | 42.8 |
|
| No | 16 | 53.3 | 8 | 57.1 |
|
| Drinking |
|
|
|
| 0.556 |
| Yes | 18 | 60.0 | 8 | 57.2 |
|
| No | 12 | 40.0 | 6 | 42.8 |
|
| Helicobacter
pylori |
|
|
|
| 0.603 |
|
Positive | 24 | 80.0 | 11 | 78.6 |
|
|
Negative | 6 | 20.0 | 3 | 21.4 |
|
Immunohistochemistry
The gastric cancer and cancer-adjacent tissues were
embedded in paraffin (Sigma-Aldrich, St. Louis, MO, USA). The
paraffin-embedded tissues were cut into 4-µm thick sections,
deparaffinized with xylene (Sigma-Aldrich) and rehydrated in ethyl
alcohol (Sigma-Aldrich). For blocking endogenous peroxidase
activity and antigen retrieval, the sections were treated with 3%
hydrogen peroxide (Sigma-Aldrich), followed by microwaving at 95°C
for 15 min. Sections were incubated with rabbit anti-human TLR7
polyclonal antibody (1:50 dilution; catalog no., ab45371; Abcam,
Cambridge, MA, USA) at 4°C overnight then washed with
phosphate-buffered saline (PBS; Fuzhou Maixin Biotechnology
Development Co., Ltd., Fuzhou, China). Controls were incubated with
PBS in place of primary antibody. Then the sections were treated
with goat anti-rabbit secondary antibody (1:100 dilution; catalog
no., MX10043; Fuzhou Maixin Biotechnology Development Co., Ltd.)
using SuperPicture Polymer Detection kit (Zymed; Thermo Fisher
Scientific, Inc., Waltham, CA, USA) and 3,3′-diaminobenzidine
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The sections were observed using a Q550CW
image acquisition and analysis system (Leica Microsystems GmbH,
Wetzlar, Germany). In total, 4–5 fields of view in each slice were
selected and measured, and the integral optical density (IOD) of
TLR7 protein expression was calculated.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from each biopsies sample
using an RNA Fast 200 kit (Shanghai Fastagen Biotech Co., Ltd.,
Shanghai, China) in accordance with manufacturer's protocol. mRNA
was reverse-transcribed with RevertAid First Strand cDNA Synthesis
kit (Thermo Fisher Scientific, Inc.) at 37°C for 15 min and stopped
at 85°C for 5 sec. RT-qPCR was performed in a final volume of 20 µl
SYBR Green PCR mixture (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The PCR conditions were as follows: Preliminary
denaturation at 94°C for 3 min, followed by 40 cycles of 95°C for
20 sec, 60°C for 20 sec and 72°C for 20 sec. All cDNA samples were
analyzed in duplicate. β-actin was used as a normalization control
in RT-qPCR. The TLR7 mRNA expression ratio was calculated as the
fold-expression of TLR7 relative to β-actin using 2−ΔΔCq
(17). The primer sequences used for
amplification were as follows: Upstream, 5′-AAACTCTGTGATGTC-3′ and
downstream, 5′-GATGTCTGGTATGTGGTTAATGG-3′ for TLR7; upstream,
5′-ATCGTGCGTGACATTAAGGAGAAG-3′ and downstream,
5′-AGGAAGGAAGGCTGGAAGAGTG-3′ for β-actin.
TLR7 activation and cell viability
assay
SGC-7901 human gastric cancer cells were obtained
from the Cell Bank of Chinese Academy of Sciences (Shanghai,
China), and maintained in RPMI-1640 medium (Invitrogen; Thermo
Fisher Scientific, Inc.) containing 10% of fetal bovine serum
(Invitrogen; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
(Fuzhou Maixin Biotechnology Development Co., Ltd.) and 100 µg/ml
streptomycin (Fuzhou Maixin Biotechnology Development Co., Ltd.) at
37°C in a humidified incubator containing 5% CO2. Cells
were seeded in 96-well plates at 5×103 cells/well, and
then exposed to 0, 1.5, 3.125, 6.25, 12.5, 25, 50, 100 or 200 µg/ml
of the TLR7 agonist imiquimod (Enzo Life Sciences International,
Inc., Plymouth Meeting, PA, USA) for various time periods (12, 24,
48, 72 h). Following treatment, 20 µl MTT (5 mg/ml; Sigma-Aldrich)
was added to each well and the plates were re-incubated at 37°C for
4 h. Subsequent to centrifugation at 800 × g for 5 min, the
supernatant was carefully removed and the product was dissolved in
150 µl dimethyl sulfoxide (Sigma-Aldrich), followed by shaking for
5 min. The absorbance of the solution was measured at 490 nm using
a microtiter plate reader (BD176863; BD Biosciences, Franklin
Lakes, NJ, USA). All experiments were performed in triplicate.
Measurement of cytokines
SGC-7901 cells were seeded in 6-well plates and
incubated overnight. The cells were treated with 100 µg/ml
imiquimod for 24 h. Subsequent to centrifugation at 1,000 × g for 5
min, the culture supernatant was collected for additional analysis.
The control cells were not treated with imiquimod; they were
instead treated with dimethyl sulfoxide (Sigma-Aldrich). The
interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α)
concentrations in culture supernatant were determined using
commercial enzyme-linked immunosorbent assay (ELISA) kits
(MQ2-39C3; R&D Systems Inc., Minneapolis, MN, USA), according
to the manufacturer's protocol.
Western blot analysis
SGC-7901 cells were seeded in 6-well plates at
1×105 cells/well. Cells were lysed with
radioimmunoprecipitation buffer on ice for 1 h. The lysate was
centrifuged (15,000 × g, 4°C) for 30 min and then the supernatant
was collected. The protein concentration was measured using a
bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. Equal amounts of
protein (20–30 µg) were applied to the SDS-PAGE (10% gel; 120V; 90
min). Glyceraldehyde 3-phosphate dehydrogenase was used as the
loading control. Following electrophoresis, the resolved protein
was transferred to a polyvinylidene difluoride membrane. The
membrane was blocked in Tris-buffered saline (Fuzhou Maixin Biotech
Development Co., Ltd.) supplemented with 5% skim milk for 2 h, and
then incubated with primary antibody (rabbit anti-human TLR7
antibody; 1:200 dilution; catalog no., ab45371; Abcam) at 4°C
overnight following by secondary antibody (goat anti-rabbit;
1:5,000 dilution; catalog no., MX20032; Fuzhou Maixin Biotechnology
Development Co., Ltd.) for 2 h at 37°C. The specific protein
expression level was detected using an electrochemiluminescence kit
(Santa Cruz Biotechnology, Inc., Dallas, TX, USA) in accordance
with the manufacturer's protocol. The images were captured using a
CAS-400SM chemiluminescence imaging system (Core Bio System, Seoul,
Republic of Korea).
Statistical analysis
Data are expressed as mean ± standard deviation.
Statistical analysis was performed using SPSS statistical software
(version 16.0; SPSS Inc., Chicago, IL, USA). The differences
between the measured data were compared using t-test and
enumeration data were compared using the χ2 test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
TLR7 mRNA expression in different
tissues
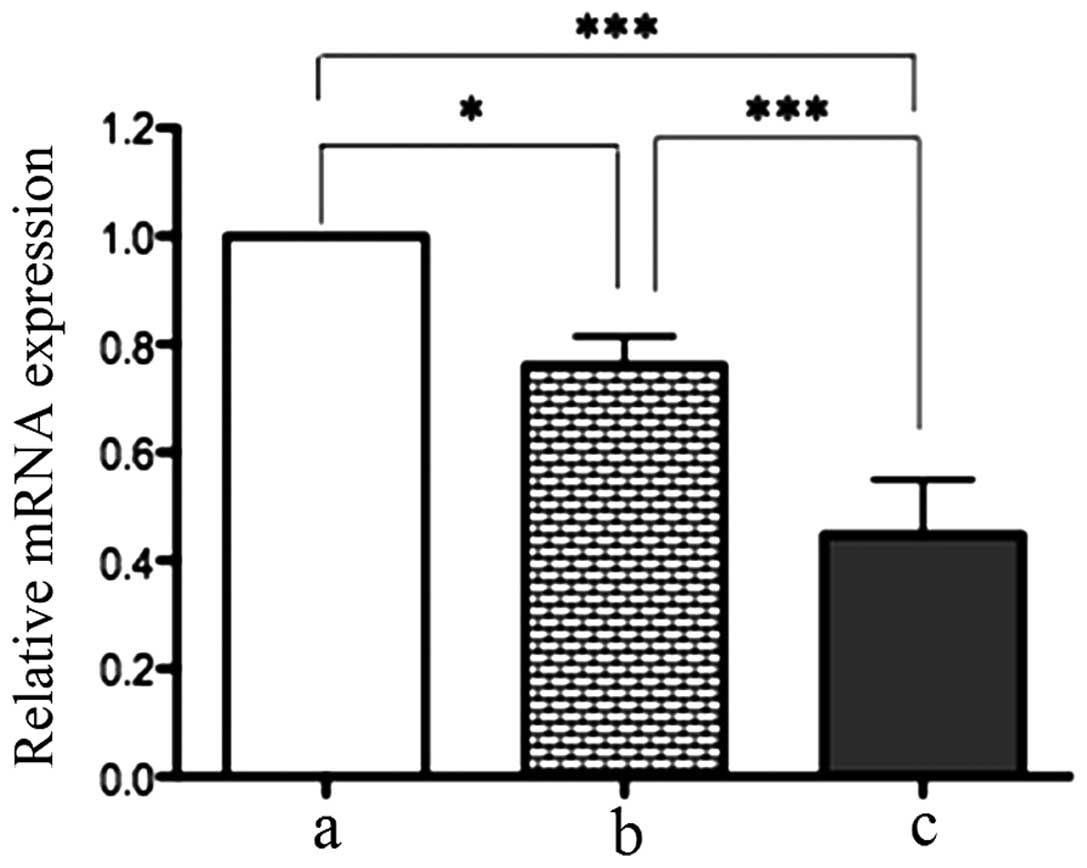
RT-qPCR demonstrated that there were significant
differences in TLR7 mRNA expression among gastric cancer,
cancer-adjacent tissues and normal gastric epithelial tissues. The
TLR7 mRNA level in gastric cancer tissues was 0.24-fold lower than
that in cancer-adjacent tissues (P<0.05) and 0.76-fold lower
compared with normal gastric epithelial tissues (P<0.01). This
indicates that TLR7 mRNA expression is downregulated in gastric
cancer tissues (Fig. 1).
TLR7 protein expression in different
tissues
Immunohistochemistry results of TLR7 in the biopsies
are presented in Fig. 2. TLR7
predominantly localized in the nucleus of glandular epithelial
cells, as well as in inflammatory cells, such as lymphomonocytes.
The expression level of TLR7 protein was markedly reduced in
gastric cancer tissues compared with cancer-adjacent tissues.
Certain gastric cancer biopsies even appeared to have a complete
absence of TLR7 expression (Fig. 2C).
As indicated in Table II, the IOD in
gastric cancer tissues was significantly lower than that in
cancer-adjacent and normal gastric epithelial tissues (P<0.01).
Although TLR7 mRNA expression levels were higher in cancer-adjacent
tissues compared with normal gastric epithelial tissues
(P<0.05), no significant difference in TLR7 protein expression
was observed between the two groups, as evaluated by IOD
(P>0.05; Table II). Furthermore,
there was no significant difference in IOD between the diffuse type
and intestinal type gastric cancer tissues (P>0.05; Table II).
| Table II.Mean integrated optical density of
toll-like receptor 7 expression in different tissues. |
Table II.
Mean integrated optical density of
toll-like receptor 7 expression in different tissues.
| Group | n | Integrated optical
density |
|---|
| Normal gastric
epithelial tissues | 14 | 0.6059±0.0609 |
| Gastric cancer
tissues | 30 |
0.4987±0.0607a,b |
|
Intestinal type | 19 |
0.4966±0.0634c |
| Diffuse
type | 11 | 0.4955±0.0443 |
| Cancer-adjacent
tissues | 30 |
0.6191±0.0737d |
Effects of TLR7 agonist on TLR7
protein expression and proinflammatory cytokine secretion in
SGC-7901 cells
Following 24 h of treatment with 100 µg/ml of TLR7
agonist imiquimod, the protein expression level of TLR7 was
markedly enhanced (Fig. 3A). This
indicates that TLR7 expression is activated by its agonist. To
evaluate whether the upregulated TLR7 could exert a biological
function, proinflammatory cytokines IL-6 and TNF-α were detected by
ELISA assays in SGC-7901 cells treated with 100 µg/ml imiquimod for
24 h As indicated in Fig. 3B,
imiquimod significantly increased the secretion of IL-6 and TNF-α
in SGC-7901 cells compared with the control (no imiquimod
treatment; P<0.01). This indicates that the activation of TLR7
may induce a biological effect, such as an immune response.
Effects of TLR7 agonist on the
viability of SGC-7901 cells
SGC-7901 cells were treated with various doses of
imiquimod for 12, 24, 48 and 72 h. Cell viability was assessed by
performing an MTT assay. As indicated in Fig. 4, imiquimod significantly reduced the
viability of SGC-7901 cells in a dose- and time-dependent manner.
Treatment with imiquimod (100 µg/ml) for 24 h led to an ~72%
inhibition in proliferation of SGC-7901 cells. This indicates that
the activation of TLR7 may inhibit the proliferation of gastric
cancer cells.
Discussion
The majority of TLRs are expressed in gastric cancer
tissues. For example, Schmausser et al (15) reported that TLR4, TLR5 and TLR9 can be
detected in lesions of intestinal metaplasia and dysplasia, as well
as gastric cancer. Notably, TLR7 expression has not yet been
identified in any gastric cell lines or tissues. The present study,
to the best of our knowledge, is the first prospective research
that evaluates the expression levels of TLR7 in patients with
gastric cancer.
TLR7 expression and function in tumorigenesis have
been examined in several types of cancer, including lung,
esophageal, colon and liver cancer. TLR7 expression is
significantly increased in non-small cell lung cancer compared with
normal bronchoscopic controls (18).
In one study of esophageal squamous cell carcinoma, only 9.2% of
normal controls were TLR7-positive, in contrast to 50%
TLR7-positive cases of esophageal squamous cell carcinoma (19). Conversely, TLR7 expression was lower
in hyperplastic and tubulovillous adenoma polyps from patients with
colorectal carcinoma, indicating a possible protective role of TLR7
against malignant transformation in colorectal mucosa (20). Furthermore, TLR7 mRNA is markedly
expressed in RWPE-1 non-cancerous prostate epithelial cells, but
not in PC3 and DU145 prostate cancer cells (21). Furthermore, a recent study revealed
that TLR7 is significantly downregulated in hepatocellular
carcinoma; in particular, patients with hepatitis B viral infection
may induce interferon-γ (IFN-γ) release to inhibit TLR7 expression,
resulting in immune escape and even immunological tolerance
(22). Thus, TLR7 may differently act
on tumorigenesis depending on the tumor origin. The present study
demonstrated that gastric cancer tissues exhibit significantly
lower mRNA and protein levels of TLR7 compared with cancer-adjacent
or normal gastric epithelial tissues. This indicates that
downregulated TLR7 expression may contribute to the promotion of
gastric carcinogenesis; however, the mechanism is unknown.
The function of TLR7 agonists in tumorigenesis has
been examined in several types of cancer. Imiquimod is a synthetic
agonist of TLR7 that has been used as a topical therapy for certain
skin neoplasms, such as basal cell carcinoma (23). The present study identified that TLR7
expression is promoted following stimulation with its agonist
imiquimod, and the secretion of proinflammatory cytokines IL-6 and
TNF-α in gastric cancer cells increases in response to imiquimod.
Thus, it appears that the activation of TLR7 can induce biological
effects and may exert immune function in gastric cancer cells.
Evidence from previous studies indicates that stimulation of TLR
expression in tumor cells can promote inflammation and cell
survival in the tumor microenvironment, and further lead to tumor
progression (24–26). However, in the present study, the
viability of SGC-7901 cells decreased when the supernatant was
supplemented with imiquimod, suggesting that a direct cytotoxic
effect was induced by imiquimod. There may be two reasons for this.
First, imiquimod may induce programmed death of gastric cancer
cells, such as autophagy and apoptosis. Accumulating evidence
indicates that TLR7 activity is not restricted to the elicitation
of innate and adaptive immune responses, but directly triggers
programmed cell death in various types of cancer, such as melanoma
and colon cancer (16,27). Second, TLR7 is selectively expressed
in plasmacytoid dendritic cells (pDCs). When TLR7 is activated,
pDCs are capable of secreting very high levels of type I IFNs in
response to pathogenic agents or danger signals. TLR7 agonists can
effectively induce pDC maturation, resulting in the induction of an
inflammatory response that contributes to the elimination of tumor
cells (28). Further studies are
required to better define the distinct role of TLR7 in gastric
tumorigenesis.
In conclusion, the expression of TLR7 is decreased
in gastric cancer tissues, and TLR7 activation appears to enhance
TLR7 expression, promote the production of proinflammatory
cytokines and reduce the viability of gastric cancer cells. The
present study has provided a basis for investigation into the
molecular mechanism of gastric cancer, and the application of the
TLR7 agonist imiquimod as a treatment modality. However, the
mechanisms of cell apoptosis are complex, and numerous pathways are
involved. Therefore, the mechanism of imiquimod-induced gastric
cancer cell apoptosis requires further study.
References
|
1
|
Akira S and Takeda K: Toll-like receptor
signalling. Nat Rev Immunol. 4:499–511. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Curtin JF, Liu N, Candolfi M, Xiong W,
Assi H, Yagiz K, Edwards MR, Michelsen KS, Kroeger KM, Liu C, et
al: HMGB1 mediates endogenous TLR2 activation and brain tumor
regression. PLoS Med. 6:e102009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Goto Y, Arigami T, Kitago M, Nguyen SL,
Narita N, Ferrone S, Morton DL, Irie RF and Hoon DS: Activation of
Toll-like receptors 2, 3 and 4 on human melanoma cells induces
inflammatory factors. Mol Cancer Ther. 7:3642–3653. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ilvesaro JM, Merrell MA, Swain TM,
Davidson J, Zayzafoon M, Harris KW and Selander KS: Toll like
receptor-9 agonists stimulate prostate cancer invasion in vitro.
Prostate. 67:774–781. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xie W, Wang Y, Huang Y, Yang H, Wang J and
Hu Z: Toll-like receptor 2 mediates invasion via activating
NF-kappaB in MDA-MB-231 breast cancer cells. Biochem Biophys Res
Commun. 379:1027–1032. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sato Y, Goto Y, Narita N and Hoon DS:
Cancer cells expressing toll-like receptors and the tumor
microenvironment. Cancer Microenviron. 2(Suppl 1): S205–S214. 2009.
View Article : Google Scholar
|
|
7
|
Wolska A, Lech-Marańda E and Robak T:
Toll-like receptors and their role in carcinogenesis and anti-tumor
treatment. Cell Mol Biol Lett. 14:248–272. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rakoff-Nahoum S and Medzhitov R: Toll-like
receptors and cancer. Nat Rev Cancer. 9:57–63. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brenner H, Rothenbacher D and Arndt V:
Epidemiology of stomach cancer. Methods Mol Biol. 472:467–477.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen W, Zheng R, Zhang S, Zhao P, Li G, Wu
L and He J: Report of incidence and mortality in China cancer
registries, 2009. Chin J Cancer Res. 25:10–21. 2013.PubMed/NCBI
|
|
12
|
Crew KD and Neugut AI: Epidemiology of
gastric cancer. World J Gastroenterol. 12:354–362. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Killeen SD, Wang JH, Andrews EJ and
Redmond HP: Exploitation of the Toll-like receptor system in
cancer: A doubled-edged sword? Br J Cancer. 95:247–252. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schmausser B, Andrulis M, Endrich S,
Müller-Hermelink HK and Eck M: Toll-like receptors TLR4, TLR5 and
TLR9 on gastric carcinoma cells: An implication for interaction
with helicobacter pylori. Int J Med Microbiol. 295:179–185. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yi JY, Jung YJ, Choi SS, Hwang J and Chung
E: Autophagy-mediated anti-tumoral activity of imiquimod in Caco-2
cells. Biochem Biophys Res Commun. 386:455–458. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cherfils-Vicini J, Platonova S, Gillard M,
Laurans L, Validire P, Caliandro R, Magdeleinat P, Mami-Chouaib F,
Dieu-Nosjean MC, Fridman WH, et al: Triggering of TLR7 and TLR8
expressed by human lung cancer cells induces cell survival and
chemoresistance. J Clin Invest. 120:1285–1297. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sheyhidin I, Nabi G, Hasim A, Zhang RP,
Ainiwaer J, Ma H and Wang H: Overexpression of TLR3, TLR4, TLR7 and
TLR9 in esophageal squamous cell carcinoma. World J Gastroenterol.
17:3745–3751. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eiró N, González L, González LO,
Andicoechea A, Fernández-Díaz M, Altadill A and Vizoso FJ: Study of
the expression of toll-like receptors in different histological
types of colorectal polyps and their relationship with colorectal
cancer. J Clin Immunol. 32:848–854. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han JH, Park SY, Kim JB, Cho SD, Kim B,
Kim BY, Kang MJ, Kim DJ and Park JH and Park JH: TLR7 expression is
decreased during tumour progression in transgenic adenocarcinoma of
mouse prostate mice and its activation inhibits growth of prostate
cancer cells. Am J Reprod Immunol. 70:317–326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin KJ, Lin TM, Wang CH, Liu HC, Lin YL
and Eng HL: Down-regulation of Toll-like receptor 7 expression in
hepatitis-virus-related human hepatocellular carcinoma. Hum Pathol.
44:534–541. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sligh JE Jr: New therapeutic options for
actinic keratosis and basal cell carcinoma. Semin Cutan Med Surg.
33(Suppl 4): S76–S80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu L and Chen S: Toll-like receptors
expressed in tumor cells: Targets for therapy. Cancer Immunol
Immunother. 57:1271–1278. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kelly MG, Alvero AB, Chen R, Silasi DA,
Abrahams VM, Chan S, Visintin I, Rutherford T and Mor G: TLR-4
signaling promotes tumor growth and paclitaxel chemoresistance in
cancer. Cancer Res. 66:3859–3868. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Smith MF Jr, Mitchell A, Li G, Ding S,
Fitzmaurice AM, Ryan K, Crowe S and Goldberg JB: Toll-like receptor
(TLR) 2 and TLR5, but not TLR4, are required for Helicobacter
pylori induced NF-kappa B activation and chemokine expression by
epithelial cells. J Biol Chem. 278:32552–32560. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang SW, Liu KT, Chang CC, Chen YJ, Wu
CY, Tsai JJ, Lu WC, Wang YT, Liu CM and Shieh JJ: Imiquimod
simultaneously induces autophagy and apoptosis in human basal cell
carcinoma cells. Br J Dermatol. 163:310–320. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aspord C, Tramcourt L, Leloup C, Molens
JP, Leccia MT, Charles J and Plumas J: Imiquimod inhibits melanoma
development by promoting pDC cytotoxic functions and impeding tumor
vascularization. J Invest Dermatol. 134:2551–2561. 2014. View Article : Google Scholar : PubMed/NCBI
|