Introduction
Cisplatin is a chemotherapy drug that is widely used
to treat various cancers, including bladder (1), ovarian (2), breast (3),
and pancreatic cancers (4), and
osteosarcoma (5). Cisplatin is a
highly reactive compound that interferes with cell division by
mitosis and cross-linking DNA (6).
When the DNA repair mechanism fails to restore the integrity of
genomic DNA, the apoptosis pathway is activated in turn (7). The presence of DNA damage-mediated cell
death elucidates the high toxicity of cisplatin for fast-dividing
cells. Nevertheless, cisplatin also demonstrates multiple side
effects on normal tissues and non-dividing cells, including kidney,
ear and sensory nerves (8–10), which suggests that the mechanism of
cisplatin-induced DNA damage may not explain the entire story and
that the underlying pathways remain poorly defined. Previous
research demonstrates that cisplatin accumulates in the
mitochondria, damaging mitochondrial DNA and proteins (11), and leads to the increase of an
oxidative stress reaction in normal cells (12,13),
suggesting that oxidative stress is involved in the pathogenesis of
cisplatin-induced cytotoxicity. Nuclear factor erythroid 2-related
factor 2 (Nrf2) is pivotal for modulating cellular redox
homeostasis and aberrant Nrf2 expression is commonly observed in
numerous cancer cells; furthermore, Nrf2/antioxidant response
element (ARE) signaling is closely associated with tumor cell
resistance to chemotherapeutic drugs (14).
Numerous endogenous antioxidants, including
superoxide and dismutase catalase, are reported to block oxidative
stress (15,16), with each possessing the capability of
scavenging reaction oxygen species (ROS). Deficiency of these
enzymes may worsen the oxidative stress-induced cell damage.
Worldwide, studies are devoted to identifying exogenous
antioxidants that ameliorate oxidative stress conditions. For
example, curcumin, isolated from the rhizome of Curcuma
longa, demonstrates antioxidant abilities in vivo
(17). Emodin, another natural
anthraquinone, demonstrates protective or ameliorative effects
against inflammation or oxidative stress-associated diseases
(18,19). The aim of the present study is to
assess the ability of emodin to alleviate the oxidative stress
induced by cisplatin, and the results may elucidate the benefits of
combined chemotherapy using emodin and cisplatin.
Materials and methods
Drugs and reagents
Cisplatin was supplied by Qilu Pharmaceutical Co.,
Ltd. (Jinan, Shandong, China) and was prepared in
phosphate-buffered saline (PBS) prior to use. Emodin (catalog no.,
A7687) was obtained from Sigma-Aldrich (St. Louis, MO, USA) and was
dissolved in dimethyl sulfoxide. Unless specified, all reagents for
cell culture were either Gibco or Invitrogen products obtained from
(Thermo Fisher Scientific, Inc., Waltham, MA USA). Mouse anti-human
monoclonal Nrf2 (catalog no., sc-365949) and β-actin (catalog no.,
sc-1616) primary antibodies were from Santa Cruz Biotechnology
(Dallas, TX, USA). ONEGlo reagent (catalog no., E6110) for the
detection of firefly luciferase activity was purchased from Promega
Corporation (Madison, WI, USA).
Cell line and culture conditions
Human osteosarcoma MG-63 cells were obtained from
American Type Culture Collection (Manassas, VA, USA) and cultured
at 37°C in a humidified atmosphere containing 5% CO2 in
Eagle's minimal essential medium complete medium supplemented with
10% fetal bovine serum.
Cell proliferation assay using
MTS
The cell proliferation assay was conducted using a
colorimetric method with an MTS assay kit (CellTiter
96®Aqueous Non-Radioactive Cell Proliferation Assay;
Promega Corporation, Madison, WI, USA). In brief, the MG-63 cells
in the logarithmic phase of growth were seeded into a 96-well plate
at a density of 1×104 cells per well, 24 h prior to
treatment. The cells were then treated with cisplatin (0.1, 0.2,
0.4, 0.8, 1.6, 3.2 µM) alone or in combination with emodin (5, 10,
20, 40, 80 and 100 µM) at the indicated concentration for 24, 36
and 48 h, respectively. The production of formazan was read at 490
nm using Flexstation 3 (Molecular Devices, Sunnyvale, CA, USA). The
intensity of the color produced was proportional to the number of
living cells.
Detection of intracellular reactive
oxygen species
The ROS in live cells were determined using a
fluorogenic probe (CellROX® Oxidative Stress Reagents;
catalog no., C10422; Molecular Probes Life Technologies, Carlsbad,
CA, USA). The assay procedure was performed as follows. Briefly,
the MG-63 cells were plated in a 6-cm plate and treated with
cisplatin (0.1, 0.2, 0.4, 0.8, 1.6, 3.2 µM) alone or in combination
with emodin (5, 10, 20, 40, 80 and 100 µM) at the indicated
concentration. The cells were then stained with 5 µM
CellROX® Deep Red Reagent, by adding the complete probe
to the complete medium and incubating the cells at 37°C for 30 min.
The cells were then washed using PBS and analyzed using BD
FACSCalibur (BD Biosciences, Franklin Lakes, NJ, USA).
Western blot analysis
For immunoblot analysis, the cells treated with
cisplatin or emodin plus cisplatin were washed with PBS and then
lysed using radioimmunoprecipitation assay buffer. Subsequent to
centrifugation at 12,000 × g for 15 min at 4°C, the supernatant was
separated on a 10% sodium dodecyl sulfate polyacrylamide gel
electrophoresis gel and transblotted onto a polyvinylidene fluoride
membrane. Subsequent to blocking with 5% bovine serum albumin in
Tris-buffered saline with Tween-20, the membrane was incubated with
aforementioned Nrf2 and β-actin (catalog no., sc-1616; Santa Cruz
Biotechnology) antibodies at dilutions of 1:3,000, and then with
the fluorescein-labeled rabbit anti-mouse polyclonal secondary
antibody (catalog no., ab6728; Santa Cruz Biotechnology) at a
dilution of 1:2,000. Specific bands were visualized using Li-COR
Odyssey (LI-COR Biosciences, Lincoln, NE, USA).
Generation of the stable reporter cell
line and ARE-luciferase activity assay
The ARE-luciferase reporter plasmid was constructed
using a pGL3 basic vector containing the full-length firefly
luciferase gene. The sequence of the minimal thymidine kinase (TK)
promoter, which was coupled to eight ARE (8xARE) repeats, was
inserted upstream of the open reading frame of luciferase. The
plasmid was sequenced to verify the accuracy of the inserts, with
the sequence of the minimal TK promoter coupled to 8xARE repeats as
follows: GTGACAAAGCACCCGTGACAAAGCACCCGTGACAAA
GCACCCGTGACAAAGCACCCGGACAAAGCACCCGTGACAAAGCACCCGTGACAAAGCACCCGTGACAAGCATTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCG
AGCGACCCTGCAGCGACCCGCTTAA.
The pGL-6xARE plasmid containing the neomycin
selectable marker was stably transfected into the MG-63 cells using
electrotransformation (Cell Line Nucleofector® kit C;
catalog no., VCA-1004; Lonza Group, Ltd., Basel, Switzerland). The
stable clones were screened using G418-containing media for 3–4
weeks.
To investigate the luciferase activity, the stable
MG-63 cells were seeded into a 96-well plate at a density of
2×104 cells per well in the conditional growth medium
(50 µl Dulbecco's modified Eagle medium; catalog no., 11965118;
Gibco, Thermo Fisher Scientific, Inc.) and incubated overnight at
37°C. The culture medium was then replaced with fresh medium
containing cisplatin (0.1, 0.2, 0.4, 0.8, 1.6, 3.2 µM) alone or
cisplatin plus emodin (5, 10, 20, 40, 80 and 100 µM) at the
indicated concentration for 24 h. Firefly luciferase activity was
determined by ONEGlo reagent (Promega Corporation, Madison, WI,
USA).
Statistical analysis
The statistical analysis was performed using
GraphPad Prism (version 6.0; GraphPad Software Inc., San Diego, CA,
USA). The results were expressed as the mean ± standard error of
the mean. Comparisons between groups were performed using one-way
analysis of variance. Student's unpaired t-test was used and
P<0.05 was considered to indicate a statistically significant
difference.
Results
Combination of cisplatin with emodin
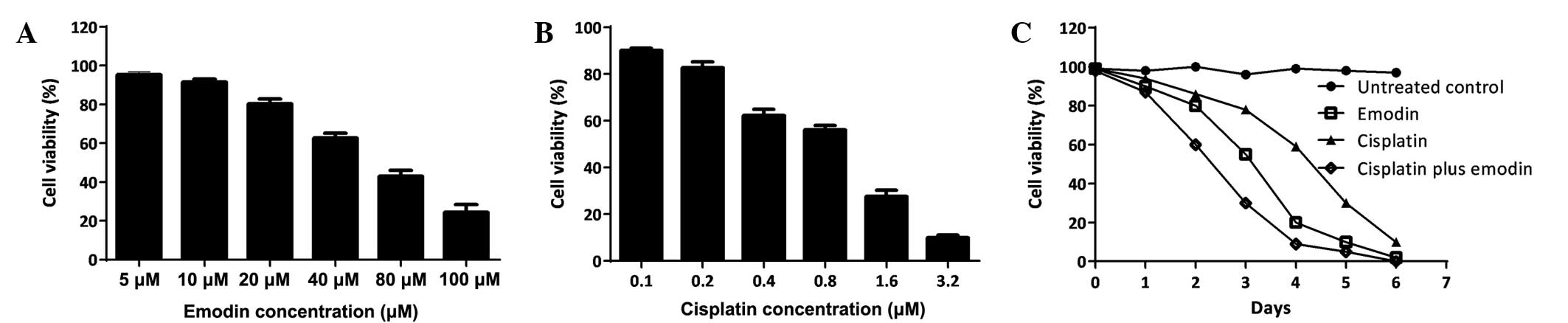
enhanced the inhibitory effects on MG-63 cell proliferation
The effects of cisplatin or emodin on human
osteosarcoma MG-63 cell growth were studied. First, the
IC50 of a single treatment with cisplatin or emodin was
determined by incubating the cells with a series of drug
concentrations and then measuring the cell survival rate at 6 days
post-treatment (Fig. 1A and B). Then,
the time course of single cisplatin or emodin, as well as the
combination of these two drugs, was assessed using a single dose
(Fig. 1C). The data demonstrated that
cisplatin and emodin could efficiently suppress the proliferation
of MG-63 cells in a dose-dependent manner. The administration of a
single dose of 0.2 µM cisplatin or 10 µM emodin inhibited MG-63
cell growth time-dependently; and using a combination of cisplatin
with emodin strengthened the inhibitory effects on MG-63 growth,
demonstrating synergistic function.
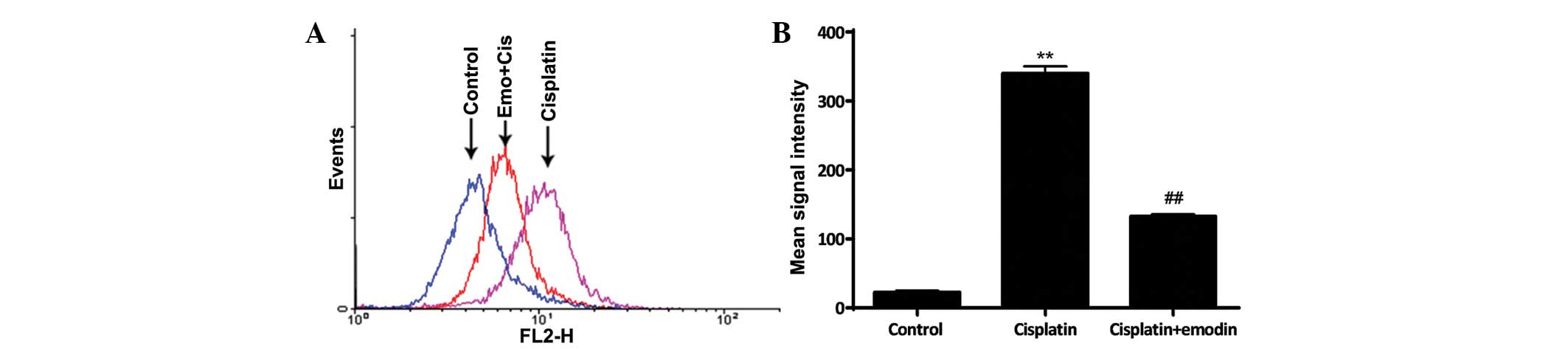
Emodin attenuated cisplatin-induced
oxidative stress in MG-63 cells
ROS was important in the pathogenesis of
cisplatin-induced cell damage. The data demonstrated that the
intracellular ROS level was significantly increased subsequent to
the treatment of the cells with 0.1 µM cisplatin for 24 h
(P<0.01; Fig. 2). However, ROS
accumulation was suppressed when the cells were simultaneously
treated with 0.1 µM cisplatin and 5 µM emodin, when compared with
the single treatment of cisplatin (P<0.01; Fig. 2). These results suggested that emodin
prevents cisplatin-induced ROS accumulation in MG-63 cells.
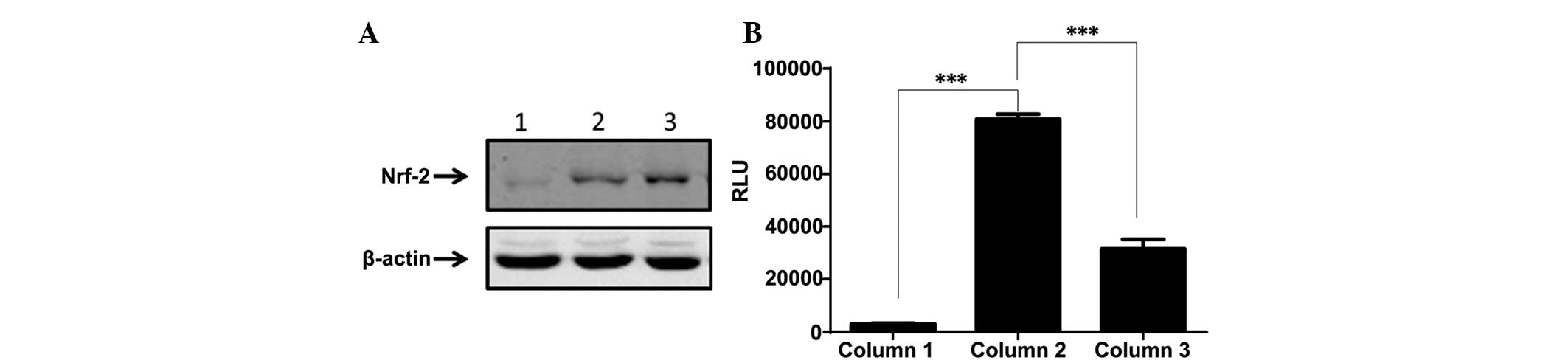
Emodin and cisplatin changed oxidative
stress conditions through the Nrf2/ARE signaling pathway
Nrf2 regulated cytoprotective genes that contained
ARE in the promoter region. An ARE-driven firefly luciferase
reporter MG-63 stable subline was developed, as previously reported
(20). Nrf2 may be retained in
cytoplasm under normal conditions, but may be translocated into the
nucleus when the cells are switched to oxidative stress conditions.
The nuclear faction of the cells was isolated and analyzed using
western blot analysis (Fig. 3A). The
data demonstrated that Nrf2 expression in the nucleus was
dramatically improved by treatment with cisplatin, while emodin
mildly blocked the cisplatin-induced translocation of Nrf2
(P<0.001; Fig. 3B). To investigate
whether the Nrf2/ARE pathway was fully activated, the
ARE/luciferase reporter MG-63 cells were tested with single
cisplatin treatment or in combination with emodin. The results
demonstrated that cisplatin triggered the cascade of Nrf2-ARE,
resulting in the robust elevation of luciferase activity. Emodin
counteracted the function of cisplatin to a certain extent. These
data indicated that emodin mitigated cisplatin-induced cell damage
by blocking the translocation of Nrf2 into the nucleus.
Discussion
The present study investigated the function of
emodin in attenuating cisplatin-induced oxidative stress in human
osteosarcoma MG-63 cells. The findings demonstrated that the
combined application of emodin and cisplatin inhibited the
malignant properties of MG-63 cells, further suppressing cellular
proliferation when compared with a single treatment with cisplatin.
In addition, the oxidative stress pathway, which was activated by
cisplatin treatment, was reversed by the addition of emodin,
demonstrating the clinical relevance of using emodin and cisplatin
simultaneously for novel therapeutic strategies for the treatment
of osteosarcoma. Emodin plus cisplatin inhibits tumor cell growth
and ameliorates cisplatin-induced oxidative stress.
As a natural bioactive anthraquinone, emodin has
previously been used therapeutically to treat gastroenteric and
liver diseases (21). Emodin is
widely used for multiple effects, such as antiviral and
hepatoprotective activities (22,23);
however, current studies focus more on the anti-tumor activity of
emodin. Previously, emodin was demonstrated to inhibit breast
cancer cell growth (24). Therefore,
it was possible that emodin may arrest human osteosarcoma
proliferation. The present study identified that emodin could
suppress MG-63 cell growth in a time- and dose-dependent manner.
Furthermore, concomitant incubation of MG-63 cells with cisplatin
and emodin demonstrated higher cellular susceptibility when
compared with single treatment with cisplatin or emodin, indicating
synergistic effects.
ROS have been demonstrated to suppress the
proliferation of malignant cells and even induce cellular apoptosis
(6). Cisplatin may induce the ROS
response and alter the redox status in malignant and normal cells,
which may limit the clinical applications of cisplatin (25). The present study also demonstrated the
augmentation of ROS in cisplatin-treated MG-63 cells, as previously
reported in another type of cell model (19). Waly et al demonstrated that
emodin protected normal cells from cisplatin-induced oxidative
stress (26). The findings of the
present study were in accordance with the outcomes of the study by
Waly et al. Furthermore, in the present study, the
concomitant application of emodin with cisplatin exerted
synergistic effects on osteosarcoma cell proliferation, although
emodin counteracted the oxidative stress conditions induced by
cisplatin.
In addition to the experiments involving the
blocking of cell proliferation, the present study also investigated
how emodin manipulated the oxidative stress signaling pathway.
Previously, Nrf2 regulated the expression of multiple oxidative
stress-associated genes (27), and
upon stimulation with certain inducers, Nrf2 was translocated into
the nucleus, where Nrf2 activated the ARE-driven antioxidant and
detoxification genes. This classical oxidative stress signaling
pathway was hypothesized to be modulated by certain anticancer
drugs. In the present study, cisplatin treatment was found to carry
Nrf2 from the cytoplasm into the nucleus and trigger the ARE-driven
luciferase expression, while concomitant treatment with emodin
diminished the translocation of Nrf2.
The molecular mechanism behind the present findings
remains undefined in detail, but the present study indicates that
the combined application of emodin with cisplatin may be a novel
therapeutic strategy for the treatment of osteosarcoma.
References
|
1
|
Shirato A, Kikugawa T, Miura N, Tanji N,
Takemori N, Higashiyama S and Yokoyama M: Cisplatin resistance by
induction of aldo-keto reductase family 1 member C2 in human
bladder cancer cells. Oncol Lett. 7:674–678. 2014.PubMed/NCBI
|
|
2
|
Muggia F: Platinum compounds 30 years
after the introduction of cisplatin: Implications for the treatment
of ovarian cancer. Gynecol Oncol. 112:275–281. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Heinemann V: Gemcitabine plus cisplatin
for the treatment of metastatic breast cancer. Clin Breast Cancer.
3(Suppl 1): 24–29. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Elligers KT, Davies M, Sanchis D, Ferencz
T and Saif MW: Rechallenge with cisplatin in a patient with
pancreatic cancer who developed a hypersensitivity reaction to
oxaliplatin. Is skin test useful in this setting? JOP. 9:197–202.
2008.PubMed/NCBI
|
|
5
|
Zhao J, Wang M, Li Z, Chen J, Yin Z, Chang
J, Gao D and Wang S: Interferon-α suppresses invasion and enhances
cisplatin-mediated apoptosis and autophagy in human osteosarcoma
cells. Oncol Lett. 7:827–833. 2014.PubMed/NCBI
|
|
6
|
Davis W Jr, Ronai Z and Tew KD: Cellular
thiols and reactive oxygen species in drug-induced apoptosis. J
Pharmacol Exp Ther. 296:1–6. 2001.PubMed/NCBI
|
|
7
|
Casares C, Ramirez-Camacho R, Trinidad A,
Roldán A, Jorge E and Garcia-Berrocal JR: Reactive oxygen species
in apoptosis induced by cisplatin: Review of physiopathological
mechanisms in animal models. Eur Arch Otorhinolaryngol.
269:2455–2459. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kursunluoglu G, Kayali HA and Taskiran D:
The effect of cisplatin toxicity and capsaicin on electron
transport chain in liver and kidney of sprague dawley rats. Cell
Biochem Biophys. 69:707–16. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leinung M, Cuny C, Diensthuber M, Stöver T
and Wagenblast J: Small molecules in combination with conventional
chemotherapeutic drugs: Light at the end of the tunnel? Oncol Lett.
4:1043–1046. 2012.PubMed/NCBI
|
|
10
|
Slattery EL and Warchol ME: Cisplatin
ototoxicity blocks sensory regeneration in the avian inner ear. J
Neurosci. 30:3473–3481. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rodrigues MA, Rodrigues JL, Martins NM,
Barbosa F, Curti C, Santos NA and Santos AC: Carvedilol protects
against cisplatin-induced oxidative stress, redox state unbalance
and apoptosis in rat kidney mitochondria. Chem Biol Interact.
189:45–51. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dehne N, Lautermann J, Petrat F, Rauen U
and de Groot H: Cisplatin ototoxicity: Involvement of iron and
enhanced formation of superoxide anion radicals. Toxicol Appl
Pharmacol. 174:27–34. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang Y, Guo C, Vasko MR and Kelley MR:
Implications of apurinic/apyrimidinic endonuclease in reactive
oxygen signaling response after cisplatin treatment of dorsal root
ganglion neurons. Cancer Res. 68:6425–6434. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lau A, Villeneuve NF, Sun Z, Wong PK and
Zhang DD: Dual roles of Nrf2 in cancer. Pharmacol Res. 58:262–270.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kessova IG, Ho YS, Thung S and Cederbaum
AI: Alcohol-induced liver injury in mice lacking Cu, Zn-superoxide
dismutase. Hepatology. 38:1136–1145. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim SJ, Lee JW, Jung YS, Kwon do Y, Park
HK, Ryu CS, Kim SK, Oh GT and Kim YC: Ethanol-induced liver injury
and changes in sulfur amino acid metabolomics in glutathione
peroxidase and catalase double knockout mice. J Hepatol.
50:1184–1191. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Khopde MS, Priyadarsini KI, Venkatesan P
and Rao MN: Free radical scavenging ability and antioxidant
efficiency of curcumin and its substituted analogue. Biophys Chem.
80:85–91. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin KY and Uen YH: Aloe-emodin, an
anthraquinone, in vitro inhibits proliferation and induces
apoptosis in human colon carcinoma cells. Oncol Lett. 1:541–547.
2010.PubMed/NCBI
|
|
19
|
Yon JM, Baek IJ, Lee BJ, Yun YW and Nam
SY: Emodin and [6]-gingerol lessen hypoxia-induced embryotoxicities
in cultured mouse whole embryos via upregulation of
hypoxia-inducible factor 1α and intracellular superoxide
dismutases. Reprod Toxicol. 31:513–518. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang XJ, Hayes JD and Wolf CR: Generation
of a stable antioxidant response element-driven reporter gene cell
line and its use to show redox-dependent activation of nrf2 by
cancer chemotherapeutic agents. Cancer Res. 66:10983–10994. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Peigen X, Liyi H and Liwei W:
Ethnopharmacologic study of Chinese rhubarb. J Ethnopharmacol.
10:275–293. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Andersen DO, Weber ND, Wood SG, Hughes BG,
Murray BK and North JA: In vitro virucidal activity of selected
anthraquinones and anthraquinone derivatives. Antiviral Res.
16:185–196. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Arosio B, Gagliano N, Fusaro LM,
Parmeggiani L, Tagliabue J, Galetti P, De Castri D, Moscheni C and
Annoni G: Aloe-Emodin quinone pretreatment reduces acute liver
injury induced by carbon tetrachloride. Pharmacol Toxicol.
87:229–233. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang PH, Huang CY, Chen MC, Lee YT, Yue
CH, Wang HY and Lin H: Emodin and aloe-emodin suppress breast
cancer cell proliferation through ER α inhibition. Evid Based
Complement Alternat Med. 2013:3761232013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Stewart JH IV, Tran TL, Levi N, Tsai WS,
Schrump DS and Nguyen DM: The essential role of the mitochondria
and reactive oxygen species in cisplatin-mediated enhancement of
rats fas ligand-induced apoptosis in malignant pleural
mesothelioma. J Surg Res. 141:120–131. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Waly MI, Ali BH, Al-Lawati I and Nemmar A:
Protective effects of emodin against cisplatin-induced oxidative
stress in cultured human kidney (HEK 293) cells. J Appl Toxicol.
33:626–630. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ge M, Chi X, Zhang A, Luo G, Sun G, Xie H
and Hei Z: Intestinal NF-E2-related factor-2 expression and
antioxidant activity changes in rats undergoing orthotopic liver
autotransplantation. Oncol Lett. 6:1307–1312. 2013.PubMed/NCBI
|