Introduction
Colitis-associated colorectal cancer (CAC) is a
subtype of colorectal cancer that is associated with inflammatory
bowel diseases (IBD), including ulcerative colitis (UC) and Crohn's
disease (1). Over 20% of patients
with IBD develop CAC within 30 years of the disease onset and
>50% of these will succumb to CAC (2). The primary risk factors for CAC
progression are the duration and severity of intestinal
inflammation (3). Increasing evidence
supports the importance of pro-inflammatory cytokines in CAC
progression (4–7). Interleukin-6 (IL-6), a pro-inflammatory
cytokine, is a key regulator in the development of CAC (8). A previous study demonstrated that IL-6
is overexpressed in patients with active UC and CAC (9). However, despite evidence implicating the
importance of IL-6 in the progression of CAC, the underlying
molecular mechanisms of this association remain unclear.
Hypoxia is a common feature of solid tumors,
including CAC, to which tumor cells frequentlyadapt through changes
in their gene expression patterns (10). Hypoxia-inducible factor 1α (HIF-1α),
an inducible transcription factor, is typically considered to be
the primary regulator of the hypoxic adaptive response (10) and is regulated by the partial pressure
of oxygen (11). In addition, a
number of growth factors, including human epidermal growth factor
receptor-2 and insulin-like growth factor-2, and cytokines,
including tumor necrosis factor-α, have been demonstrated to
regulate the expression of HIF-1α (12–14).
Furthermore, previous studies identified that the activation of
oncogenes and loss of function of tumor suppressor genes effects
HIF-1α expression (15,16). HIF-1α induces the expression of
hundreds of genes and serves important roles in various aspects of
cancer biology, including angiogenesis, invasion and metastasis
(17–19). In clinical studies, HIF-1α was shown
to be overexpressed in multiple types of cancer, including
colorectal (20,21). In addition, HIF-1α was identified to
be overexpressed in patients with IBD and in the early stages of
colorectal cancer (22). Furthermore,
patients carrying one or both of the common HIF-1α polymorphisms,
C1772T and G1790A, displayed a significantly higher risk for the
development of CAC (23). These
results indicate that HIF-1α serves a significant role in the
progression of CAC. However, the association between IL-6 and
HIF-1α during the progression of CAC remains unclear.
In the present study, the association between IL-6
and HIF-1α in the development of CAC was investigated. The results
of the current study demonstrated that, in a CAC mouse model,
theanti-IL-6 receptor antibody used inhibited the progression of
CAC and decreased the expression of HIF-1α. Further analysis in
human samples revealed that the serum concentration of IL-6 was
positively correlated with HIF-1α mRNA expression in CAC tumor
tissues. In conclusion, the results of the present study indicate
that IL-6 promotes CAC progression through the regulation of
HIF-1α.
Materials and methods
Mouse CAC model
Male BALB/c mice (6–8 weeks old; 18.3±1.4 g),
purchased from the Shanghai Laboratory Animal Center (Shanghai
Institutes for Biological Sciences, Chinese Academy of Sciences,
Shanghai, China), were used to produce the CAC model. Mice were
housed at 22±1°C, with a 12 h light/dark cycle, and ad
libitum access to food and water. Mice were divided into two
groups; the immunoglobulin G (IgG) group (control group, n=8) and
the anti-IL-6 receptor antibody (both eBioscience, Inc., San Diego,
CA, USA) group (treatment group, n=8). The protocol to induce CAC
in a mouse model was performed as previously described by Neufert
et al (24). Briefly, mice
were peritoneally injected with azoxymethane (AOM, 10 mg/kg;
Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) at the
beginning of the first week. Then, mice were given drinking water
supplemented with 2% (w/v) dextran sulfate sodium (DSS;
Sigma-Aldrich; Merck Millipore) for 1 week, followed by water alone
for 2 weeks, this was repeated 3 times (Fig. 1A). Simultaneously, mice in the
treatment and control groups were injected peritoneally every 2
days with 10 µg anti-IL-6 receptor antibody or 10 µg IgG,
respectively, diluted in 200 µl normal saline.
 | Figure 1.Anti-IL-6 receptor antibody inhibits
the development of CAC. (A) Schematic overview of CAC induction
with AOM and DSS. Mice were peritoneally injected with AOM (10
mg/kg) at the beginning of the first week. Subsequently, mice were
treated with 2% DSS supplemented into their drinking water for 1
week, followed by 2 weeks of water alone. This cycle was repeated 3
times. In addition, anti-IL-6 receptor antibody and immunoglobulin
G were peritoneally injected into treatment and control mice,
respectively, every 2 days. (B) Macroscopic observation showed that
the majority of colorectal neoplasms were located in the middle and
distal colon. (C) Fewer and (D) smaller macroscopic neoplasms were
observed in the treatment group compared with the control group.
*P<0.05. (E) Colorectal neoplasms were divided into adenomas and
adenocarcinomas following histopathological examination
(magnification, ×200). IL-6, interleukin-6; CAC, colitis-associated
colorectal cancer; AOM, azoxymethane; DSS, dextran sulfate
sodium. |
The mice were sacrificed via cervical dislocation at
the end of week 9. Subsequently, large intestines (from the
ileocecal junction to the anal verge) were collected,
longitudinally cut open along the main axis and washed with normal
saline. Visible neoplasms were counted and collected, the length
and width of each neoplasm was measured using a digital micro
ruler, and the neoplasm area was calculated by multiplying the
length and width. Subsequently, neoplasms were cut into halves,
where 1 half was prepared for histopathological analysis and
immunohistochemistry, and the other half was prepared for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis and western blotting. All procedures were carried out in
accordance with the guidelines and regulations for the Use of
Experimental Animals by the Chinese Academy of Sciences (Beijing,
China). The current study was approved by the Animal Ethics
Committee of Zhongshan Hospital of Fudan University (Shanghai,
China).
Collection of patient samples
A total of 13 patients (males n=8, females n=5),
aged between 41 and 76 years old (58.6±10.9 years), diagnosed with
CAC and receiving surgical treatment at Zhongshan Hospital of Fudan
University between June 2012 and December 2014 were included in the
present study. Tumor tissue and adjacent non-malignant tissue was
collected during surgery and stored at −80°C until required for
further analysis. A total of 5 ml blood was collected from each
patient prior to surgery and immediately centrifuged at 2,400 ×
g for 10 min at room temperature. The serum obtained was
preserved at −80°C until required. The patients included in the
present study had not received chemotherapy or radiotherapy prior
to surgery. Informed consent was obtained from all participants
included in the current study and ethical permission for the study
was obtained from the Zhongshan Hospital of Fudan University.
Histopathological analysis
Biopsy specimens were fixed with 10% formalin,
embedded in paraffin and cut into 4-µm-thick sections. Hematoxylin
and eosin staining was performed on the sections to distinguish
between colorectal adenomas and adenocarcinomas. All results were
confirmed by 2 pathologists.
Measurement of serum IL-6
concentration
The serum concentration of IL-6 was assessed using
Canine IL-6 DuoSet ELISA (R&D Systems, Inc., Minneapolis, MN,
USA; cat. no. DY1609), according to the manufacturer's protocol.
The detection range of this kit was between 0.156 and 10.0 pg/ml,
with a sensitivity of 0.11 pg/ml.
Immunohistochemistry
Immunohistochemical staining of colorectal tissue
sections was performed as previously described by Yuan et al
(25). Briefly, tissue sections were
treated with xylene and a series of graded alcohols. Antigen
retrieval was performed with 0.01 M citrate buffer (pH 6.0) at 95°C
for 20 min. Following antigen retrieval, the sections were
incubated with primary mouse monoclonal anti-HIF-1α antibody
(1:100; Abcam, Cambridge, UK; cat. no. ab113642) at 4°C for 12 h.
Following three washes with 0.01 mmol/l PBS, sections were
incubated with goat anti-mouse IgG H&L (FITC) biotinylated
secondary antibody (1:50; Abcam; cat. no. ab6785) at 37°C for 1 h
and horseradish peroxidase-conjugated streptavidin (1:10,000;
Abcam; cat no. ab7403) for at room temperature 15 min. Then,
sections were treated with diaminobenzidine and hydrogen peroxide
chromogen substrate plus diaminobenzidine enhancer for 10 min,
followed by counter staining with Mayer's hematoxylin. The
percentage and intensity of stained immunoassayed cells was
evaluated over 5 visual fields, selected at random, at a
magnification of ×400 using a light microscope. The degree of
staining was evaluated independently by 2 pathologists. Expression
of HIF-1α was indicated by the presence of cytoplasmic and/or
membranous staining.
Total RNA extraction and RT-qPCR
analysis
Total RNA was isolated from human and mouse
colorectal tissue samples using the RNAprep Pure Tissue Kit
(Tiangen Biotech Co., Ltd., Beijing, China; cat. no. DP431) and
cDNA was synthesized from 1 µg total RNA using the cDNA Synthesis
Kit (Takara Bio, Inc., Otsu, Japan; cat. no. 6130), following the
manufacturer's protocol. RT-qPCR reactions were performed using the
SYBR Premix Ex Taq kit (Takara Bio, Inc.; cat no. DRR820A) on the
StepOnePlus Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Thermocycling conditions were
as follows: 95°C for 2 min; 40 cycles of 30 sec at 95°C; 10 sec at
60°C; 30 sec at 72°C; and 10 min at 72°C. The expression level of
the HIF-1α gene relative to β-actin was determined using the SYBR
Green-based comparative CT method (2−ΔΔCq) (26). The primer sequences used were as
follows: Human HIF-1α sense, 5′-ACTTCTGGATGCTGGTGATT-3′ and
antisense, 5′-GTCGTGCTGAATAATACCACT–3′; mouse HIF-1α sense,
5′-AGCCCTAGATGGCTTTGTGA-3′ and antisense,
5′-TATCGAGGCTGTGTCGACTG-3′; and human and mouse β-actin sense,
5′-CACGATGGAGGGGCCGGACTCATC-3′ and antisense,
5′-TAAAGACCTCTATGCCAACACAGT-3′.
Western blotting
Preparation of total protein lysates and western
blot analysis was performed as previously described by Yuan et
al (25). To detect HIF-1α,
monoclonal anti-HIF-1α antibody (1:1,000) was used. Tubulin
expression, detected using an anti-tubulin antibody (1:2,000;
Sigma-Aldrich; Merck Millipore) was used as an endogenous
control.
Statistical analysis
Results are presented as the mean ± standard error
of the mean. Data from the present study was analyzed with GraphPad
Prism (version 5.0; GraphPad Software, Inc., La Jolla, CA, USA)
using a Student's t-test. The correlation between serum IL-6
concentration and HIF-1α mRNA expression was analyzed using the
Pearson product-momentum correlation coefficient. P<0.05 was
considered to indicate a statistically significant difference.
Results
Anti-IL-6 receptor antibody inhibits
the development of CAC
CAC was successfully induced in the mice using AOM
and DSS (Fig. 1A). Macroscopically,
the majority of colorectal neoplasms were located in the middle and
distal colon (Fig. 1B). Mice in the
treatment group developed significantly fewer and smaller neoplasms
compared with mice in the control group (P<0.05; Fig. 1C and D). Following histopathological
examination, colorectal neoplasms were divided into adenomas and
adenocarcinomas (Fig. 1E). The
incidence of colorectal adenocarcinomas was 87.5% (7/8 mice) in the
treatment group and 100% (8/8 mice) in the control group, while the
incidence of colorectal adenomas was 100% (8/8 mice) in the
treatment and control groups. These results suggest that the
anti-IL-6 receptor antibody used inhibits the development of
CAC.
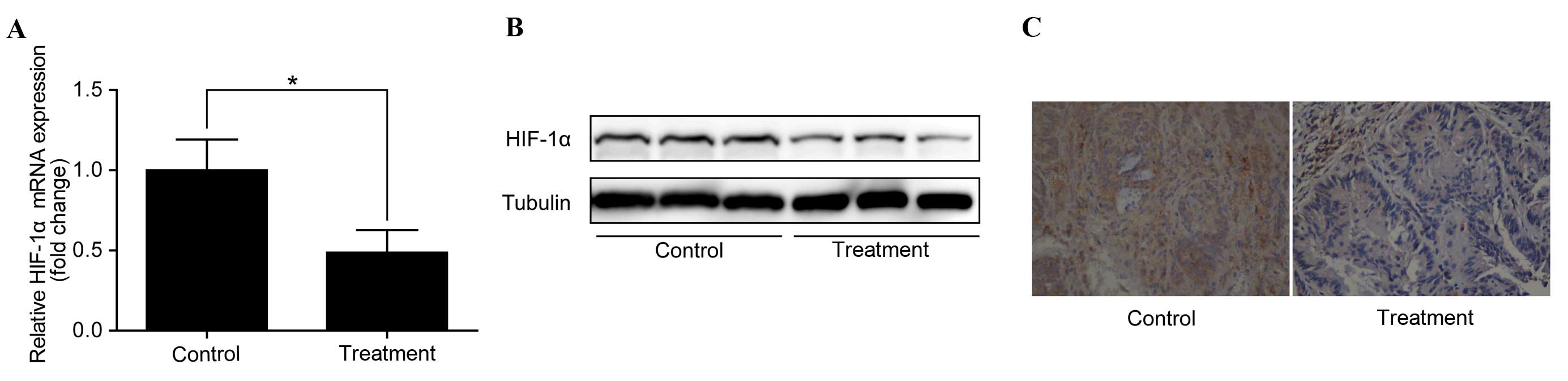
Anti-IL-6 receptor antibody
downregulates HIF-1α in colorectal adenocarcinomas
To identify the potential mechanisms by which the
anti-IL-6 receptor antibody inhibits the development of CAC and to
investigate whether this effect is through the regulation of
HIF-1α, the expression of HIF-1α in colorectal adenocarcinomas was
examined. RT-qPCR analysis demonstrated that HIF-1α mRNA expression
was significantly downregulated in colorectal adenocarcinomas
following treatment with the anti-IL-6 receptor antibody (P<0.05
vs. control; Fig. 2A). In addition,
western blotting identified a corresponding downregulation of
HIF-1α protein levels (Fig. 2B).
Furthermore, immunohistochemistry revealed a notable decrease in
HIF-1α protein levels in the treatment group compared with the
control group (Fig. 2C).
Anti-IL-6 receptor antibody
downregulates HIF-1α in colorectal adenomas
To investigate whether the anti-IL-6 receptor
antibody used downregulates HIF-1α in early stage CAC, the
expression of HIF-1α in colorectal adenomas was examined. Similarly
to the results observed in colorectal adenocarcinomas, HIF-1α was
found to be downregulated in colorectal adenomas following
treatment with the anti-IL-6 receptor antibody (Fig. 3A-C). This indicates that IL-6
regulates HIF-1α expression in early stage CAC.
HIF-1α mRNA expression positively
correlates with serum IL-6 concentration in patients with CAC
As HIF-1α expression was significantly downregulated
by the anti-IL-6 receptor antibody used in the mouse CAC model,
HIF-1α expression was analyzed in patients with CAC to determine
whether it positively correlated with serum IL-6 concentration.
RT-qPCR analysis revealed that HIF-1α mRNA levels were
significantly increased in CAC tumor tissues compared with adjacent
non-malignant tissues (P<0.05; Fig.
4A). The fold change in HIF-1α mRNA expression in tumor tissues
compared with adjacent non-malignant tissues was between 0.8 and
5.2 (median, 2.1). In addition, western blotting confirmed that
there was an overexpression of HIF-1α protein in the malignant
tissues (Fig. 4B). Then, serum IL-6
concentration was examined in the participants, which identified
that serum IL-6 concentrations were between 1.2 and 9.7 pg/ml
(median, 6.4 pg/ml). Furthermore, the Pearson product-momentum
correlation coefficient showed that serum IL-6 concentration
positively correlated with HIF-1α mRNA expression in the tumor
tissue of these patients with CAC (r=0.58; P=0.04; Fig. 4C).
Discussion
IL-6 overexpression is a well-known risk factor for
CAC and the role of HIF-1α during CAC progression is well
documented (22,27). However, whether IL-6 contributes to
CAC progression through the regulation of HIF-1α remains unclear.
The results of the present study indicate that inhibition of IL-6
using an anti-IL-6 receptor antibody hindered the development of
CAC and decreased HIF-1α expression, even in early stage CAC. In
addition, HIF-1α was demonstrated to be overexpressed in human CAC
tumor tissue and positively correlated with serum IL-6
concentration. These results indicate that IL-6 promotes CAC
progression by regulating HIF-1α expression during the early stages
of CAC development.
Following the combination of exposure to AOM and 3
cycles of DSS treatment, CAC was successfully induced in BALB/c
mice. Similarly to a previous study (28), the tumor incidence rate was 100% in
the mice. In addition, following the inhibition of IL-6 using the
anti-IL-6 receptor antibody, tumor size and number was
significantly reduced, which was consistent with the results of a
previous study (29).
In recent years IL-6 has emerged as a potential
therapeutic target in patients diagnosed with malignant disease
(30). For example, anti-IL-6
antibodies have been used to treat patients with multiple myeloma
or acquired immune deficiency syndrome-associated Kaposi's sarcoma
since the 1990s (31,32). In 2014 siltuximab, a chimeric anti-IL6
antibody, was approved by the U.S. Food and Drug Administration
(FDA; Silver Spring, MD, USA) for the treatment of patients
diagnosed with Castleman disease (33). In addition, siltuximab was used in
clinical trials for a number of malignant tumors, including
metastatic renal cell cancer, prostate cancer and colorectal cancer
(34–36). Although tocilizumab, an anti-IL-6
receptor antibody, has not yet been approved for the treatment of
malignant tumors, it has been approved by the FDA for the treatment
of chronic inflammatory diseases, including rheumatoid arthritis
and juvenile idiopathic arthritis (37). In the present study, the anti-IL-6
receptor antibody used was observed to have a protective effect in
a mouse CAC model. However, further investigations are required to
validate the effectiveness of anti-IL-6 receptor antibody treatment
of CAC.
Signal transducer and activator of transcription-3,
suppressor of cytokine signaling-3 and vascular endothelial growth
factor receptor-2 were reported to be the downstream proteins
through which IL-6 exerts its tumor promoting effect in CAC
(38–41). However, there is little literature on
the association between IL-6 and HIF-1α in the development of CAC.
In the present study, HIF-1α was downregulated in colorectal
adenocarcinomas and adenomas in the mouse CAC model by the
anti-IL-6 receptor antibody, which indicates that IL-6 stimulates
the development of CAC through the regulation of HIF-1α. However,
whether IL-6 exerts its tumor promoting effect directly through the
regulation of HIF-1α remains unclear.
In patients with CAC, HIF-1α was overexpressed in
tumor tissues compared with adjacent non-malignant tissues, which
is consistent with the results of a previous study (18). In addition, a positive correlation
between HIF-1α mRNA expression and serum IL-6 concentration in
patients with CAC was observed. However, these findings should be
interpreted with caution due to the relatively small number of
patients included in the present study. Further research with a
larger sample size is required to confirm these results.
In conclusion, the present study provides evidence
that IL-6 promotes the development of CAC through the regulation of
HIF-1α during the early stages of CAC progression. In addition, the
results of the current study demonstrate that anti-IL-6 receptor
antibodies are a potential therapeutic agent for CAC that warrant
further investigation.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81372197).
References
|
1
|
Grivennikov SI: Inflammation and
colorectal cancer: Colitis-associated neoplasia. Semin
Immunopathol. 35:229–244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lakatos PL and Lakatos L: Risk for
colorectal cancer in ulcerative colitis: Changes, causes and
management strategies. World J Gastroenterol. 14:3937–3947. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ullman TA and Itzkowitz SH: Intestinal
inflammation and cancer. Gastroenterology. 140:1807–1816. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Waldner MJ and Neurath MF: Cytokines in
colitis associated cancer: Potential drug targets? Inflamm Allergy
Drug Targets. 7:187–194. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dubois RN: Role of inflammation and
inflammatory mediators in colorectal cancer. Trans Am Clin Climatol
Assoc. 125:358–373. 2014.PubMed/NCBI
|
|
6
|
Li Y, Deuring J, Peppelenbosch MP, Kuipers
EJ, de Haar C and van der Woude CJ: IL-6-induced DNMT1 activity
mediates SOCS3 promoter hypermethylation in ulcerative
colitis-related colorectal cancer. Carcinogenesis. 33:1889–1896.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qi H, Yang H, Xu G, Ren J, Hua W, Shi Y,
Torsvik M, Florholmen J and Cui G: Therapeutic efficacy of IL-17A
antibody injection in preventing the development of colitis
associated carcinogenesis in mice. Immunobiology. 220:54–59. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Waldner MJ, Foersch S and Neurath MF:
Interleukin-6-a key regulator of colorectal cancer development. Int
J Biol Sci. 8:1248–1253. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Y, de Haar C, Chen M, Deuring J,
Gerrits MM, Smits R, Xia B, Kuipers EJ and van der Woude CJ:
Disease-related expression of the IL6/STAT3/SOCS3 signalling
pathway in ulcerative colitis and ulcerative colitis-related
carcinogenesis. Gut. 59:227–235. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Monti E and Gariboldi MB: HIF-1 as a
target for cancer chemotherapy, chemosensitization and
chemoprevention. Curr Mol Pharmacol. 4:62–77. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci
USA. 92:5510–5514. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feldser D, Agani F, Iyer NV, Pak B,
Ferreira G and Semenza GL: Reciprocal positive regulation of
hypoxia-inducible factor 1alpha and insulin-like growth factor 2.
Cancer Res. 59:3915–3918. 1999.PubMed/NCBI
|
|
13
|
Laughner E, Taghavi P, Chiles K, Mahon PC
and Semenza GL: HER2 (neu) signaling increases the rate of
hypoxia-inducible factor 1alpha (HIF-1alpha) synthesis: Novel
mechanism for HIF-1-mediated vascular endothelial growth factor
expression. Mol Cell Biol. 21:3995–4004. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hellwig-Bürgel T, Rutkowski K, Metzen E,
Fandrey J and Jelkmann W: Interleukin-1beta and tumor necrosis
factor-alpha stimulate DNA binding of hypoxia-inducible factor-1.
Blood. 94:1561–1567. 1999.PubMed/NCBI
|
|
15
|
Chen C, Pore N, Behrooz A, Ismail-Beigi F
and Maity A: Regulation of glut1 mRNA by hypoxia-inducible
factor-1. Interaction between H-ras and hypoxia. J Biol Chem.
276:9519–9525. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zundel W, Schindler C, Haas-Kogan D, Koong
A, Kaper F, Chen E, Gottschalk AR, Ryan HE, Johnson RS, Jefferson
AB, et al: Loss of PTEN facilitates HIF-1-mediated gene expression.
Genes Dev. 14:391–396. 2000.PubMed/NCBI
|
|
17
|
Liao D and Johnson RS: Hypoxia: A key
regulator of angiogenesis in cancer. Cancer Metastasis Rev.
26:281–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Semenza GL: Defining the role of
hypoxia-inducible factor 1 in cancer biology and therapeutics.
Oncogene. 29:625–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pugh CW and Ratcliffe PJ: Regulation of
angiogenesis by hypoxia: Role of the HIF system. Nat Med.
9:677–684. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Krishnamachary B, Berg-Dixon S, Kelly B,
Agani F, Feldser D, Ferreira G, Iyer N, LaRusch J, Pak B, Taghavi P
and Semenza GL: Regulation of colon carcinoma cell invasion by
hypoxia-inducible factor 1. Cancer Res. 63:1138–1143.
2003.PubMed/NCBI
|
|
21
|
Zhong H, De Marzo AM, Laughner E, Lim M,
Hilton DA, Zagzag D, Buechler P, Isaacs WB, Semenza GL and Simons
JW: Overexpression of hypoxia-inducible factor 1alpha in common
human cancers and their metastases. Cancer Res. 59:5830–5835.
1999.PubMed/NCBI
|
|
22
|
Mariani F, Sena P, Marzona L, Riccio M,
Fano R, Manni P, Gregorio CD, Pezzi A, Leon MP, Monni S, et al:
Cyclooxygenase-2 and Hypoxia-Inducible Factor-1alpha protein
expression is related to inflammation, and up-regulated since the
early steps of colorectal carcinogenesis. Cancer Lett. 279:221–229.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fransén K, Fenech M, Fredrikson M,
Dabrosin C and Söderkvist P: Association between ulcerative growth
and hypoxia inducible factor-1alpha polymorphisms in colorectal
cancer patients. Mol Carcinog. 45:833–840. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Neufert C, Becker C and Neurath MF: An
inducible mouse model of colon carcinogenesis for the analysis of
sporadic and inflammation-driven tumor progression. Nat Protoc.
2:1998–2004. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yuan L, Han J, Meng Q, Xi Q, Zhuang Q,
Jiang Y, Han Y, Zhang B, Fang J and Wu G: Muscle-specific E3
ubiquitin ligases are involved in muscle atrophy of cancer
cachexia: An in vitro and in vivo study. Oncol Rep. 33:2261–2268.
2015.PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Grivennikov SI and Karin M: Inflammation
and oncogenesis: A vicious connection. Curr Opin Genet Dev.
20:65–71. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Suzuki R, Kohno H, Sugie S, Nakagama H and
Tanaka T: Strain differences in the susceptibility to azoxymethane
and dextran sodium sulfate-induced colon carcinogenesis in mice.
Carcinogenesis. 27:162–169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Becker C, Fantini MC, Schramm C, Lehr HA,
Wirtz S, Nikolaev A, Burg J, Strand S, Kiesslich R, Huber S, et al:
TGF-beta suppresses tumor progression in colon cancer by inhibition
of IL-6 trans-signaling. Immunity. 21:491–501. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Middleton K, Jones J, Lwin Z and Coward
JI: Interleukin-6: An angiogenic target in solid tumours. Crit Rev
Oncol Hematol. 89:129–139. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bataille R, Barlogie B, Lu ZY, Rossi JF,
Lavabre-Bertrand T, Beck T, Wijdenes J, Brochier J and Klein B:
Biologic effects of anti-interleukin-6 murine monoclonal antibody
in advanced multiple myeloma. Blood. 86:685–691. 1995.PubMed/NCBI
|
|
32
|
Racadot E, Audhuy B, Duvernoy H, Thyss A,
Lang JM, Wijdenes J and Hervé P: Clinical and immunological
follow-up of patients with AIDS-associated Kaposi's sarcoma treated
with an anti-IL-6 monoclonal antibody. Cytokines Mol Ther.
1:133–138. 1995.PubMed/NCBI
|
|
33
|
van Rhee F, Wong RS, Munshi N, Rossi JF,
Ke XY, Fosså A, Simpson D, Capra M, Liu T, Hsieh RK, et al:
Siltuximab for multicentric Castleman's disease: A randomised,
double-blind, placebo-controlled trial. Lancet Oncol. 15:966–974.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rossi JF, Négrier S, James ND, Kocak I,
Hawkins R, Davis H, Prabhakar U, Qin X, Mulders P and Berns B: A
phase I/II study of siltuximab (CNTO 328), an anti-interleukin-6
monoclonal antibody, in metastatic renal cell cancer. Br J Cancer.
103:1154–1162. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fizazi K, De Bono JS, Flechon A,
Heidenreich A, Voog E, Davis NB, Qi M, Bandekar R, Vermeulen JT,
Cornfeld M and Hudes GR: Randomised phase II study of siltuximab
(CNTO 328), an anti-IL-6 monoclonal antibody, in combination with
mitoxantrone/prednisone versus mitoxantrone/prednisone alone in
metastatic castration-resistant prostate cancer. Eur J Cancer.
48:85–93. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Angevin E, Tabernero J, Elez E, Cohen SJ,
Bahleda R, van Laethem JL, Ottensmeier C, Lopez-Martin JA, Clive S,
Joly F, et al: A phase I/II, multiple-dose, dose-escalation study
of siltuximab, an anti-interleukin-6 monoclonal antibody, in
patients with advanced solid tumors. Clin Cancer Res. 20:2192–2204.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tanaka T, Narazaki M and Kishimoto T:
Therapeutic targeting of the interleukin-6 receptor. Annu Rev
Pharmacol Toxicol. 52:199–219. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Grivennikov S, Karin E, Terzic J, Mucida
D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H,
Eckmann L and Karin M: IL-6 and Stat3 are required for survival of
intestinal epithelial cells and development of colitis-associated
cancer. Cancer Cell. 15:103–113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bollrath J, Phesse TJ, von Burstin VA,
Putoczki T, Bennecke M, Bateman T, Nebelsiek T, Lundgren-May T,
Canli O, Schwitalla S, et al: gp130-mediated Stat3 activation in
enterocytes regulates cell survival and cell-cycle progression
during colitis-associated tumorigenesis. Cancer Cell. 15:91–102.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rigby RJ, Simmons JG, Greenhalgh CJ,
Alexander WS and Lund PK: Suppressor of cytokine signaling 3
(SOCS3) limits damage-induced crypt hyper-proliferation and
inflammation-associated tumorigenesis in the colon. Oncogene.
26:4833–4841. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Waldner MJ, Wirtz S, Jefremow A, Warntjen
M, Neufert C, Atreya R, Becker C, Weigmann B, Vieth M, Rose-John S
and Neurath MF: VEGF receptor signaling links inflammation and
tumorigenesis in colitis-associated cancer. J Exp Med.
207:2855–2868. 2010. View Article : Google Scholar : PubMed/NCBI
|