Introduction
Human esophageal squamous cell carcinoma (ESCC) is
the 6th leading cause of cancer-associated mortality, and the 8th
most common type of cancer worldwide (1). ESCC is the most common histology of
esophageal cancers, and has a high incidence in China, South
Africa, France and Italy (2).
Epidemiological and etiological studies have previously
demonstrated that certain environmental and genetic factors have
crucial roles in esophageal carcinogenesis (3–7). Although
there have been advances in the fields of surgery and chemotherapy,
the prognosis for ESCC is typically poor as the majority of
patients diagnosed are elderly and present with advanced-stage
disease, rendering them unsuitable for curative resection and
maintaining the five-year survival rate for ESCC at <15%
(1). Therefore, improving the current
understanding of the molecular mechanisms underlying ESCC
pathogenesis and identifying effective therapeutic agents are
required to facilitate the development of novel therapies for this
disease.
Resveratrol is present in red grapes, wine, nuts and
common garden plants, and is one of the most intensively
investigated of all the phytochemicals with putative beneficial
effects on human health (8). The
effects of resveratrol include anti-oxidant and anti-aging
properties, improvement of insulin sensitivity and reduction of
cardiovascular disease risk (9–11).
Previous studies have also indicated that resveratrol is able to
function as a chemo preventive and chemotherapeutic agent in
certain types of human carcinomas, including those of the
colorectum, skin, liver, lung, pancreas, breast and prostate
(8,12–14).
Previous studies suggesting that a dietary pattern rich in
resveratrol may reduce the risk of ESCC (15). However, the molecular mechanisms
underlying resveratrol-induced antitumor effects have yet to be
elucidated. A number of potential mechanisms have been suggested to
be involved in resveratrol-induced cancer preventative effects,
including the inhibition of tumorigenesis-associated transcription
factor activation, the suppression of various protein kinases and
the regulation of the expression of specific genes involved in cell
cycle progression, apoptosis, angiogenesis and metastasis (8,12,16–18).
The epidermal growth factor receptor (EGFR) is a
receptor tyrosine kinase that mediates intracellular signaling in
response to various extracellular stimuli (19). Previous studies have demonstrated that
EGFR is activated and overexpressed in numerous types of human
epithelial cancers, including human ESCC (19–21). The
aberrant regulation of EGFR expression activates downstream
signaling pathways, including extracellular signal-regulated
kinases (ERKs) and protein kinase B (AKT), which increase tumor
cell proliferation, survival and invasiveness (19). Modulation of the EGFR signaling
pathway is considered to have a crucial role in cancer prevention
(19). However, the role of
resveratrol in the regulation of EGFR in order to mediate its
carcinogenic effects has yet to be demonstrated in ESCC. In the
present study, the antitumor efficacy of resveratrol in human ESCC
cells and the role of EGFR in the antitumor activity of
resveratrol, as well as the mechanisms underlying the inhibition of
EGFR activation by resveratrol, are investigated. The results
demonstrated that resveratrol exhibited a marked inhibitory effect
on EGFR tyrosine kinase activity in vitro, in addition to
downregulating EGFR protein levels and the expression levels of the
downstream gene cyclin D1, which was also associated with
resveratrol-mediated cell cycle arrest and cell death.
Materials and methods
Cell lines and culture
All cell lines were purchased from the Cell Bank of
Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). KYSE150, KYSE510 and Eca109 ESCC cells were
cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(Invitrogen; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
and 100 mg/ml streptomycin. All cell lines were incubated at 37°C
in a humidified atmosphere containing 5% CO2.
Reagents and antibodies
Resveratrol was purchased from Sigma-Aldrich (Merck
Millipore, Darmstadt, Germany). Sub-confluent cells were treated
with the compound at various concentrations (0, 20, 40 and 80 µM)
for 24 h. The final concentration of dimethyl sulfoxide (DMSO) in
the culture media was maintained at <0.1%, which had no
significant effect on the cell growth. Anti-EGFR (#4267; 1:2,000),
anti-phosphorylated (p)-EGFR (Tyr1068; #3777; 1:2,000), anti-p-AKT
(Ser473; #4060; 1:1,000), anti-p-ERK1/2 (Thr202/Tyr204; #4370;
1:2,000), anti-cyclinD1 (#2978; 1:1,000), anti-α-tubulin (#2144;
1:5,000), cleaved PARP (#5625; 1:1,000), cleaved caspase-3 (#9664;
1:1,000) and anti-lamin B (#13415; 1:2,000) antibodies were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Anti-β-actin (#sc-1615; 1: 5,000), anti-rabbit immunoglobulin (Ig)
G-horseradish peroxidase (HRP) (#sc-2040; 1:10,000) and anti-mouse
IgG-HRP (#sc-2039; 1:10,000) were purchased from Santa Cruz
Biotechnology (Dallas, TX, USA). Anti-N-cadherin (#610920; 1:3,000)
was purchased from BD Biosciences (San Jose, CA, USA).
Western blotting
Cells were harvested by trypsinization and
centrifuged (108 × g for 5 min at 4°C). The cell pellets
were lysed in Nonidet P-40 cell lysis buffer (50 mM Tris-Cl, pH
8.0, 150 mM NaCl, 0.5% Nonidet P-40 and protease inhibitor mixture;
Thermo Fisher Scientific, Inc.). The extracted protein
concentrations were determined using a Bradford assay (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Proteins were separated by
sodium dodecyl sulfate polyacrylamide gel electrophoresis and
electrically transferred to a polyvinylidene difluoride membrane
(EMD Millipore, Billerica, MA, USA). Following blocking with 5%
nonfat dry milk in Tris-buffered saline (TBS), the membranes were
hybridized to specific primary antibodies overnight at 4°C, washed
three times with TBS-Tween 20 and then incubated with the secondary
antibodies conjugated with HRP for 1 h at room temperature. The
membranes were then washed three times in TBS-Tween 20 at room
temperature. Visualization of proteins was performed using the
ChemiDoc XRS system with Image Lab software (Bio-Rad Laboratories,
Inc.). For EGF treatment, the cells were incubated in serum-free
medium overnight and then treated with resveratrol at various
concentrations (0, 20 and 40 µM) for 2 h. Following stimulation
with 50 ng/ml EGF (Thermo Fisher Scientific, Inc.) for 30 min, the
cells were harvested and the relative protein expression levels
were determined by western blot analysis.
Subcellular proteome
fractionation
The ESCC cells were treated with resveratrol (0, 20,
40 and 80 µM) for 48 h, the cells were harvested by trypsinization
and centrifuged (108 × g for 5 min at 4°C). The subcellular
proteome fractions were prepared using a ProteoExtract Subcellular
Proteome Extraction kit (EMD Millipore) according to the
manufacturer's instructions. The subcellular proteome fractions
were subjected to western blot analysis.
MTS assay
To determine the cytotoxicity of resveratrol, the
cells were seeded (3×103 cells/well) in 96-well plates
and cultured at 37°C in a humidified atmosphere containing 5%
CO2 overnight. The cells were then cultured with fresh
medium, treated with various doses of resveratrol or DMSO and
cultured for various time periods (0, 24, 48 and 72 h). The
cytotoxicity of resveratrol was evaluated using an MTS assay kit
(Promega Corporation, Madison, WI, USA) according to the
manufacturer's instructions. The absorbance was read at 490 nm.
Soft agar colony assay
In order to examine anchorage-independent growth,
KYSE150, Eca109 and KYSE510 cells (8,000 cells/ml) were suspended
in 1 ml 0.3% agar with Eagle's basal medium (Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS, 1% antibiotics (100
U/ml penicillin and 100 mg/ml streptomycin) and various
concentrations of resveratrol (0, 20, 40 and 80 µM) and seeded onto
6-well plates containing a 0.6% agar base. The cultures were
maintained in a 5% CO2 incubator at 37°C for 1–2 weeks,
and then the colonies were imaged under a light microscope and
counted using the Image-Pro Plus software program version 6 (Media
Cybernetics, Inc., Rockville, MD, USA).
Flow cytometry
Flow cytometry was used to quantify the number of
cells in each phase of the cell cycle. The cells (2×105)
were seeded into 6-well plates and treated with various
concentrations (0, 20, 40, 80 µM) of resveratrol for 24 h. The
cells were then harvested by trypsinization and centrifuged (108 ×
g for 5 min at 4°C), then washed with phosphate-buffered
saline (PBS) twice and fixed in 70% ethanol overnight at 4°C.
Subsequently, the cells were stained in the dark with 50 µg/ml
propidium iodide containing 0.1% ribonuclease A (BD Biosciences) in
400 µl PBS at 25°C for 30 min. Stained cells were assayed and
quantified using a FACSort Flow Cytometer (BD Biosciences). For
apoptosis analysis, KYSE150 cells were suspended (1×106
cells/ml) in staining buffer and 5 µl Annexin V and propidium
iodide (PI) staining solution were added to 300 µl cell suspension.
Following incubation of the cells for 10–15 min at room temperature
in the dark, the stained cells were assayed and quantified using a
FACSort Flow Cytometer.
Statistical analysis
All statistical analyses were performed using SPSS
software version 13.0 (SPSS, Inc., Chicago, IL, USA). The
experiments were all performed in triplicate and the data are
expressed as the mean ± standard deviation. The significant
differences between the groups were assessed using a two-tailed
Student's t-test. P<0.05 was considered to indicate a
statistically significant result.
Results
Resveratrol inhibits ESCC cell
proliferation and anchorage-independent growth
The antitumor efficacy of resveratrol on ESCC cell
lines was investigated using MTS and soft agar assays. At a low
concentration (20 µM), resveratrol exhibited little effect on the
growth inhibition, but at higher concentrations (40–80 µM),
long-term (48–72 h) treatment with resveratrol significantly
inhibited cell proliferation (Fig.
1A; P<0.05). Furthermore, as presented in Fig. 1B, resveratrol may potently inhibit
anchorage-independent cell growth, even at 20 µM as the number of
colonies that formed on the soft agar were significantly decreased;
at high concentrations (80 µM), almost no colonies were observed to
form (P<0.05, P<0.01 and P<0.001). These results indicate
that resveratrol has a marked antitumor efficacy in ESCC cells.
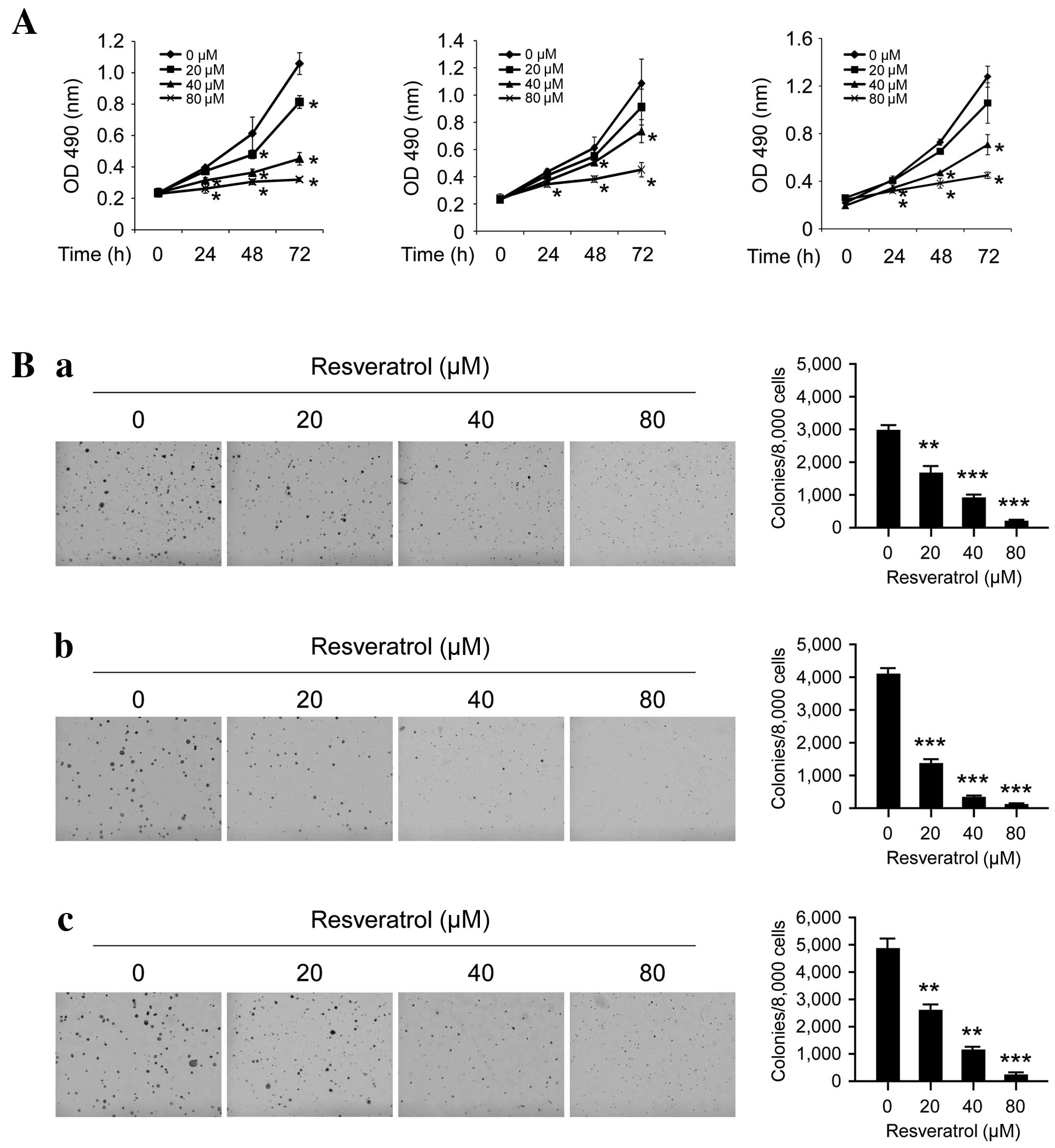 | Figure 1.Inhibitory effects of resveratrol on
the anchorage-dependent and -independent growth of human ESCC
cells. (A) Resveratrol inhibits anchorage-dependent growth in human
ESCC cells, including KYSE510 (left), Eca109 (middle) and KYSE150
(right) cells. The MTS assay was used to assess the inhibitory
effect of resveratrol on ESCC cells. (B) Resveratrol suppresses
anchorage-independent growth in human ESCC cells, including (a)
KYSE510, (b) Eca109 and (c) KYSE150 cells. MTS and colony formation
assays were conducted to investigate the viability and colony
formation ability of human ESCC cells treated with various
concentrations of resveratrol, compared with ESCC cells treated
with DMSO. Each experiment was conducted in triplicate and
independently. The data are presented as the mean ± standard
deviation. *P<0.05, **P<0.01, ***P<0.001 suppression of
viability or colony formation by resveratrol vs. DMSO (control).
ESCC, esophageal squamous cell carcinoma; DMSO, dimethyl sulfoxide;
OD, optical density. |
Short-term exposure to resveratrol
downregulates the EGF-induced EGFR signaling pathway
The EGFR signaling pathway is frequently
dysregulated in human ESCC cells or tissues and is considered to
have an important role in cell proliferation and survival (22). The results of the present study
revealed that treatment with resveratrol markedly inhibits EGFR
activation in a dose-dependent manner (Fig. 2A). The EGF-induced phosphorylation of
EGFR was markedly inhibited in KYSE150 cells by 40 µM resveratrol;
the phosphorylation of ERK1/2 and AKT, two downstream kinases of
EGFR, were also significantly inhibited. In order to further
investigate the effect of resveratrol on EGFR activation,
EGF-induced EGFR phosphorylation was evaluated at various time
points. Resveratrol suppressed EGFR activation, as well as its
downstream kinases ERK1/2 and AKT at all selected time points
(Fig. 2B). These results suggest that
resveratrol exerts its antitumor effects via inhibiting EGFR
activation and its downstream signaling pathways.
 | Figure 2.Resveratrol affects EGF-induced EGFR
signaling in KYSE150 ESCC cells. (A) Resveratrol inhibits the
EGF-induced EGFR signaling pathway in a dose-dependent manner.
KYSE150 cells were treated with resveratrol, and the expression of
p-EGFR, p-AKT, p-ERK1/2 and β-actin was determined by western
blotting. (B) Resveratrol inhibits the EGF-induced EGFR signaling
pathway in a time-dependent manner. KYSE150 cells were incubated in
serum-free medium overnight. Following treatment with resveratrol
for 2 h, the cells were stimulated with EGF (50 ng/ml) for various
time periods (0, 15, 30 and 60 min), p-EGFR, p-AKT, p-ERK1/2 and
β-actin expression was determined by western blotting. The
histograms represent the means ± standard deviations from three
independent experiments; **P<0.01 and ***P<0.001 for
suppression of protein expression by resveratrol vs. DMSO (control)
group. EGF, epidermal growth factor; EGFR, epidermal growth factor
receptor; ESCC, esophageal squamous cell carcinoma; p,
phosphorylated; AKT, protein kinase B; ERK1/2, extracellular
signal-regulated kinases 1/2. |
Long-term exposure to resveratrol
downregulates EGFR protein expression in KYSE150 cells
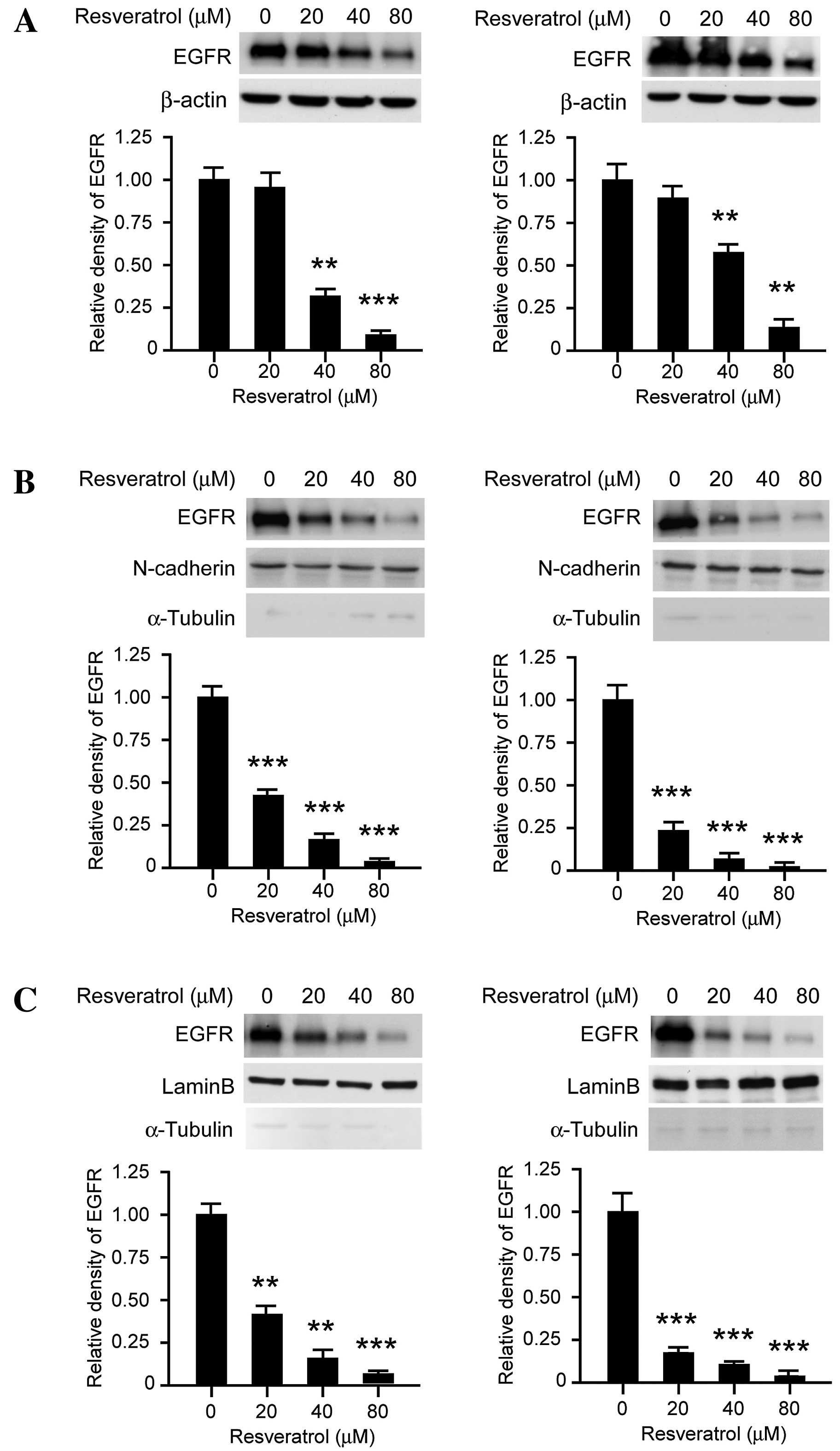
The results of the current study demonstrated that
the EGFR signaling pathway is inhibited following transient
resveratrol treatment, which indicated that resveratrol may
suppress the tyrosine kinase activity of EGFR in a dose-dependent
manner. As presented in Fig. 3A,
long-term exposure (>48 h) to resveratrol decreased the
expression levels of EGFR in dose-dependent manner. Following
treatment with 80 µM resveratrol for 48 h, EGFR protein expression
levels in KYSE150 cells were significantly downregulated. As
previous studies have reported that EGFR is located in the nucleus
in addition to the cellular membrane, subcellular separation was
performed in order to investigate the effect of resveratrol on
subcellular EGFR expression patterns. As presented in Fig. 3B, membrane-associated EGFR expression
levels were markedly decreased following treatment with resveratrol
treated for 48 h, in KYSE150 (left) and Eca109 (right) cells.
Furthermore, the expression levels of EGFR localized to the nucleus
were also downregulated following resveratrol exposure for 48 h
(Fig. 3C). These results suggest
that, following long-term treatment with resveratrol, the
expression levels of EGFR at the cell membrane and in the nucleus
were suppressed.
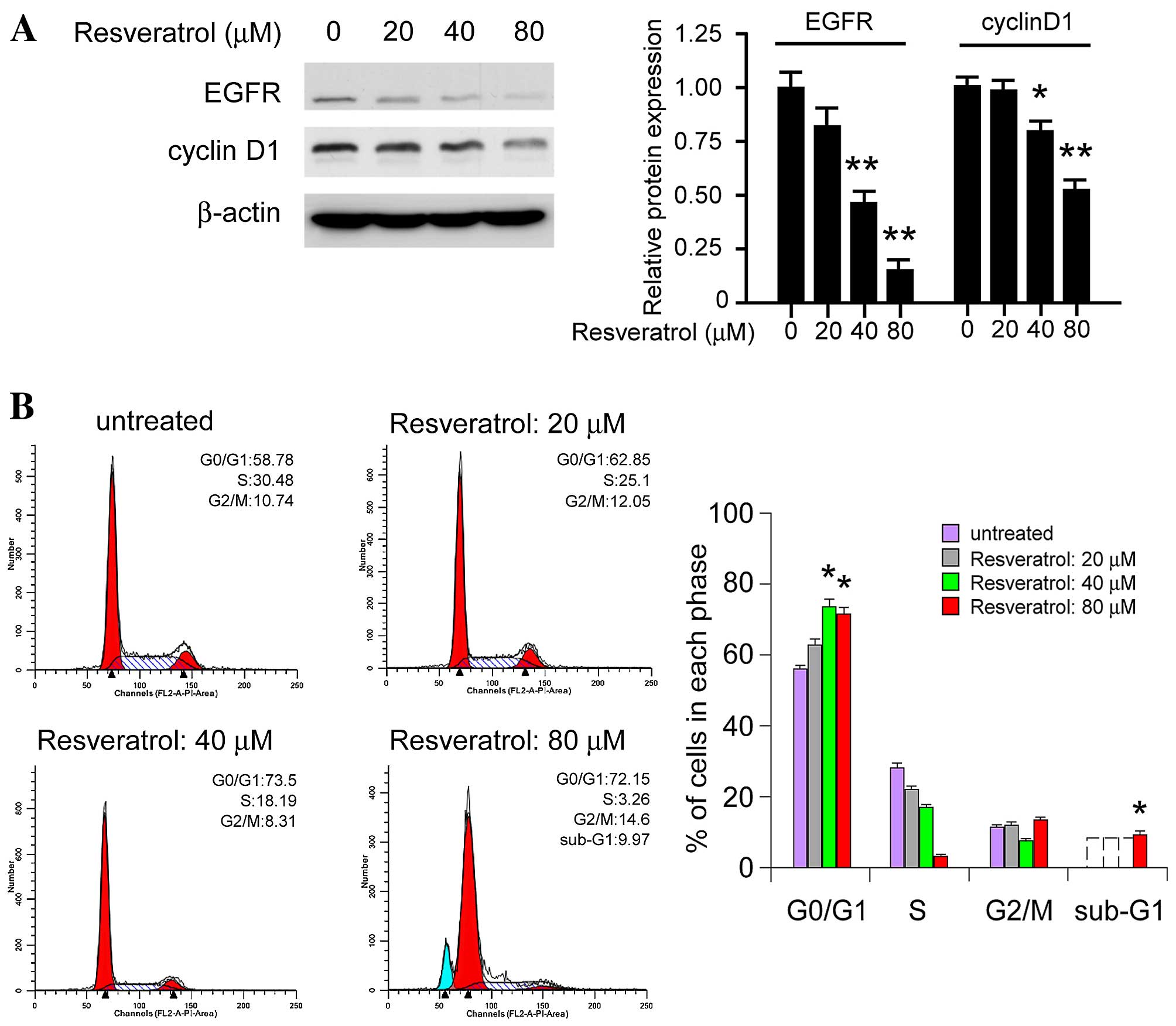
Resveratrol suppresses cyclin D1
expression and induces cell cycle arrest at
G0/G1 phase
As an important downstream target of EGFR, cyclin D1
has a crucial role in G1/S progression in the cell
cycle. It was demonstrated that treatment with resveratrol for 24 h
markedly decreased the expression levels of EGFR, as well as cyclin
D1 expression levels in KYSE150 cells (Fig. 4A). Additionally, resveratrol treatment
had an effect on the cell cycle, inducing cell cycle arrest at the
G0/G1 phase in dose-dependent manner. At a
low concentration (20 µM) cell cycle arrest was not observed;
however, higher concentrations (40–80 µM) induced
G0/G1 phase arrest and the proportion of
cells in the G0/G1 phase was significantly
increased (P<0.05). As shown in Fig.
4B, 80 µM resveratrol is able to induce the sub-G1
peak, which indicated that high doses of resveratrol may directly
induce cell death. The results suggest that resveratrol inhibits
cyclin D1 expression levels and induces cell cycle arrest at the
G0/G1 phase.
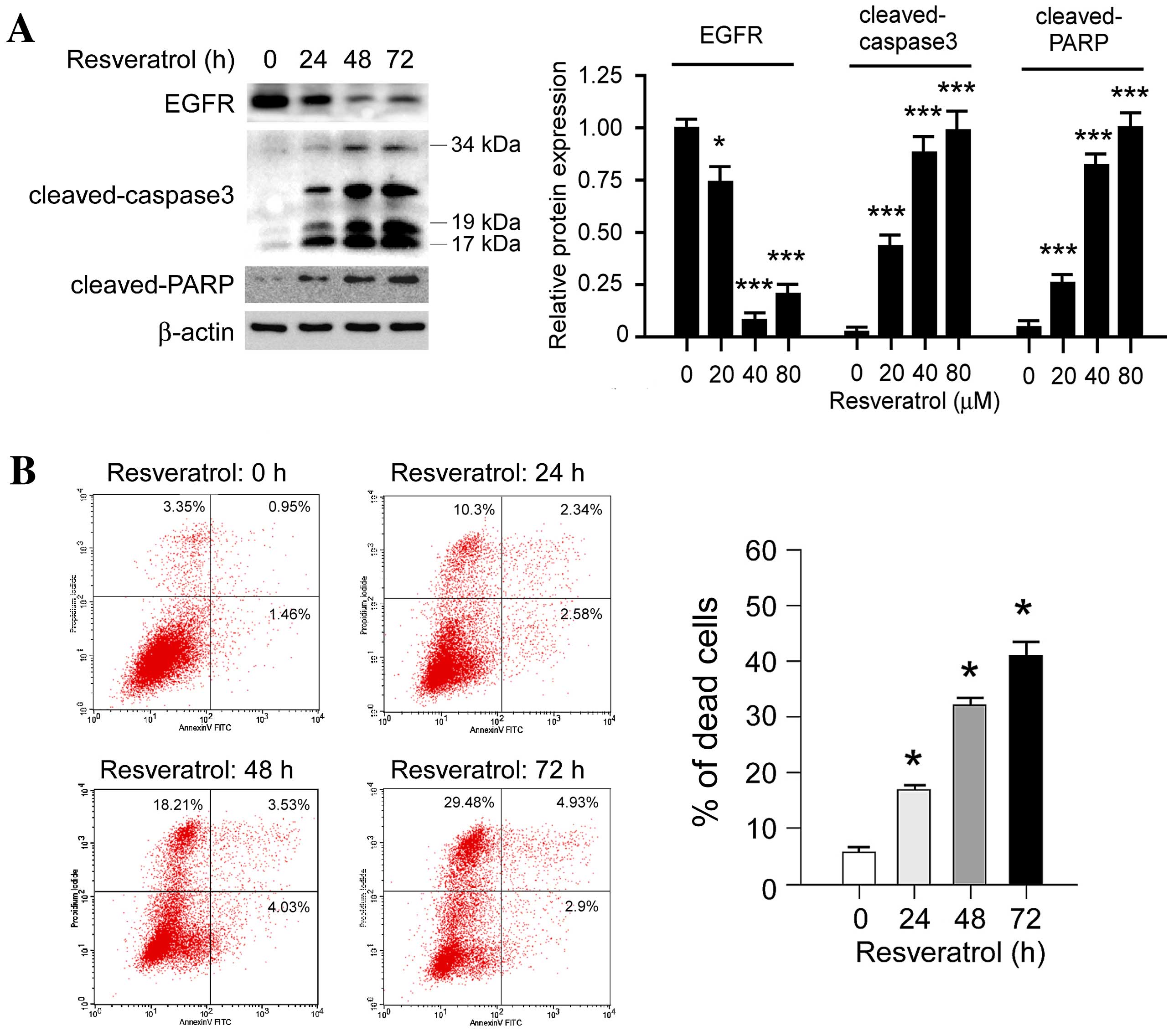
Resveratrol promotes cell death in
KYSE150 cells
In order to further investigate the effect of
resveratrol on cell death, the expression levels of
apoptosis-associated molecules, including cleaved poly (ADP-ribose)
polymerase (PARP) and cleaved caspase-3, were evaluated following
treatment with resveratrol. Western blot analysis revealed that the
expression levels of cleaved PARP and cleaved caspase-3 were
markedly increased in a time-dependent manner following treatment
with 80 µM resveratrol (Fig. 5A). In
addition, PI/Annexin V staining and flow cytometric analysis
indicated that resveratrol induced KYSE150 cell death in a
time-dependent manner (Fig. 5B),
concordant with the results from the western blotting. Notably, it
was identified that high-dose resveratrol treatment induced other
types of cell death as well as apoptosis; The PI-stained cells were
significantly upregulated (P<0.05) following resveratrol
treatment. The results suggest that resveratrol treatment induces
cell death in human ESCC cells.
Discussion
Numerous studies previously reported that
resveratrol exhibited a marked efficacy in various types of tumor
tissues, including hepatocellular carcinoma, ovarian cancer and
breast cancer (16,17). However, its activity and underlying
mechanisms in ESCC have yet to be elucidated. In the present study,
it was demonstrated that resveratrol dose-dependently inhibits ESCC
cell proliferation in vitro and that colony formation is
markedly decreased following resveratrol treatment (Fig. 1). Furthermore, exposure of ESCC cells
to resveratrol effectively induced G0/G1 cell
cycle arrest. In addition, resveratrol treatment stimulated ESCC
cells to undergo apoptosis, as revealed by flow cytometry and
western blot analysis. Further investigation revealed that
resveratrol exerts its antitumor activity by regulating the EGFR
signaling pathway in ESCC.
EGFR has an important role in tumor development,
including in cell proliferation, anti-apoptosis, angiogenesis,
tumor metastasis and invasion (23,24).
Dysregulation of the EGFR signaling pathway through overexpression
or hyperactivation is associated with tumor development (23,25,26). A
meta-analysis demonstrated that the rate of EGFR overexpression in
ESCC is high and EGFR expression levels are a potential predictor
of the T stage (tumor-node-metastasis classification system)
(27), vascular invasion and overall
survival (27). By contrast, Aichler
et al (28) reported that EGFR
is an independent adverse prognostic factor in esophageal
adenocarcinoma patients treated with cisplatin-based neoadjuvant
chemotherapy; therefore, targeting EGFR may be a novel therapeutic
approach in the management of ESCC. EGFR is typically expressed at
the cellular membrane, and is activated via autophosphorylation on
binding with its ligands, epidermal growth factor (EGF) or
transforming growth factor-α. EGFR phosphorylation activates
numerous downstream signaling pathways, including the rat
sarcoma/rapidly accelerated fibrosarcoma/mitogen activated protein
kinase and phosphoinositide-3-kinase/AKT signaling pathways, which
regulate various cellular functions (29,30). In
the present study, the results demonstrated that short-term
exposure to resveratrol significantly inhibits EGF-induced EGFR
phosphorylation. Following the inhibition of EGFR activation, the
downstream signaling pathways and phosphorylation of EGFR, ERK and
AKT were suppressed. Similarly to a number of natural compounds,
including epigallocatechin gallate (31) and quercetin (32), resveratrol has also been demonstrated
to inhibit various kinases (33). The
current study only investigated the effect of resveratrol on the
EGFR signaling pathway in human ESCC cells, which is one limitation
as resveratrol may also target other kinases in ESCC cells. It was
also identified that long-term (48 h) exposure to resveratrol in
ESCC cells induces EGFR degradation. Previous studies have
demonstrated that the phosphorylation of EGFR at Tyr1045 creates a
docking site for casitas B-lineage lymphoma, an adaptor protein
that leads to receptor ubiquitination and degradation following
EGFR activation (33,34). It is possible that resveratrol may
induce EGFR degradation via the ubiquitination or lysosome
signaling pathways in human ESCC cells. Further investigation
revealed that the expression levels of EGFR at the cell membrane
were reduced and EGFR located in the nucleus was also significantly
decreased following resveratrol treatment. It was hypothesized that
reduction of EGFR expression levels at the cell membrane may reduce
the sensitivity of the cell to growth factors, and cell
proliferation may be markedly decreased following stimulation with
growth factors.
Previous studies have demonstrated that the nuclear
import of EGFR and the signal cascade induced by nuclear EGFR had
pivotal roles in the malignant proliferation and survival of
certain types of tumor tissues (35–37).
Immunohistochemical analysis of breast tumor tissues revealed that
EGFR expression levels in the nucleus are positively correlated
with the expression of Ki-67, a biomarker of cell proliferation,
and negatively correlated with the survival rate (38,39). In
oral squamous cell carcinoma (OSCC), elevated nuclear import of
EGFR may predict a poor outcome for OSCC patients (40). It has also been identified that
nuclear EGFR is involved in acquired chemotherapy resistance
(41). In pancreatic cancer, nucleus
EGFR combines with sarcoma and signal transducer and activator of
transcription 5 to form a complex that regulates the transcription
of the c-myc gene and to mediates the sensitivity of tumor
cells to chemotherapy (42). Previous
studies also identified that EGFR nuclear import upregulated the
activity of DNA-dependent protein kinases (DNA-PK) and promoted the
repair of DNA damage (43). The EGFR
monoclonal antibody Cetuximab may block EGFR nuclear import,
downregulate the activity of DNA-PK, inhibit DNA repair and induce
tumor cell apoptosis (43–46). The present study demonstrated that
long-term treatment with resveratrol markedly decreases the
expression levels of EGFR in the nucleus, suggesting that
resveratrol may assist the prevention of chemotherapy resistance
when combined with cytotoxic drugs.
In conclusion, the results demonstrated that
resveratrol is effective in inhibiting the proliferation of ESCC
cells, suggesting that a dietary pattern rich in resveratrol may
reduce the risk of ESCC, as initially hypothesized by Lin et
al (15). The results also
revealed that resveratrol influences EGFR activity in ESCC cells,
short-term exposure of ESCC cells to resveratrol had an inhibitory
effect on EGFR phosphorylation and its signaling pathway, but
long-term treatment induced EGFR degradation and had an effect on
the activity of EGFR in the nucleus and at the cell membrane. The
results suggested that resveratrol, or analog, may have a role in
the prevention and management of ESCC.
Acknowledgements
This study was supported by the National Nature
Science Foundation of China (grant no. 81472774).
Glossary
Abbreviations
Abbreviations:
|
EGFR
|
epidermal growth factor receptor
|
|
ESCC
|
esophageal squamous cell carcinoma
|
References
|
1
|
van Hagen P, Hulshof MC, van Lanschot JJ,
Steyerberg EW, van Berge Henegouwen MI, Wijnhoven BP, Richel DJ,
Nieuwenhuijzen GA, Hospers GA, Bonenkamp JJ, et al: Preoperative
chemoradiotherapy for esophageal or junctional cancer. N Engl J
Med. 366:2074–2084. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hongo M, Nagasaki Y and Shoji T:
Epidemiology of esophageal cancer: Orient to Occident. Effects of
chronology, geography and ethnicity. J Gastroenterol Hepatol.
24:729–735. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cheung WY and Liu G: Genetic variations in
esophageal cancer risk and prognosis. Gastroenterol Clin North Am.
3875–91. (viii)2009.PubMed/NCBI
|
|
5
|
Wu C, Hu Z, He Z, Jia W, Wang F, Zhou Y,
Liu Z, Zhan Q, Liu Y, Yu D, et al: Genome-wide association study
identifies three new susceptibility loci for esophageal
squamous-cell carcinoma in Chinese populations. Nat Genet.
43:679–684. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu C, Li D, Jia W, Hu Z, Zhou Y, Yu D,
Tong T, Wang M, Lin D, Qiao Y, et al: Genome-wide association study
identifies common variants in SLC39A6 associated with length of
survival in esophageal squamous-cell carcinoma. Nat Genet.
45:632–638. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gao YB, Chen ZL, Li JG, Hu XD, Shi XJ, Sun
ZM, Zhang F, Zhao ZR, Li ZT, Liu ZY, et al: Genetic landscape of
esophageal squamous cell carcinoma. Nat Genet. 46:1097–1102. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gescher A, Steward WP and Brown K:
Resveratrol in the management of human cancer: How strong is the
clinical evidence? Ann N Y Acad Sci. 1290:12–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Frazzi R and Tigano M: The multiple
mechanisms of cell death triggered by resveratrol in lymphoma and
leukemia. Int J Mol Sci. 15:4977–4993. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park EJ and Pezzuto JM: The pharmacology
of resveratrol in animals and humans. Biochim Biophys Acta.
1852:1071–1113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Novelle MG, Wahl D, Diéguez C, Bernier M
and de Cabo R: Resveratrol supplementation: Where are we now and
where should we go? Ageing Res Rev. 21:1–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Singh CK, George J and Ahmad N:
Resveratrol-based combinatorial strategies for cancer management.
Ann N Y Acad Sci. 1290:113–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xue YQ, Di JM, Luo Y, Cheng KJ, Wei X and
Shi Z: Resveratrol oligomers for the prevention and treatment of
cancers. Oxid Med Cell Longev. 2014:7658322014.PubMed/NCBI
|
|
14
|
Yang X, Li X and Ren J: From French
Paradox to cancer treatment: Anti-cancer activities and mechanisms
of resveratrol. Anticancer Agents Med Chem. 14:806–825. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin Y, Yngve A, Lagergren J and Lu Y: A
dietary pattern rich in lignans, quercetin and resveratrol
decreases the risk of oesophageal cancer. Br J Nutr. 112:2002–2009.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kulkarni SS and Cantó C: The molecular
targets of resveratrol. Biochim Biophys Acta. 1852:1114–1123. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singh CK, Ndiaye MA and Ahmad N:
Resveratrol and cancer: Challenges for clinical translation.
Biochim Biophys Acta. 1852:1178–1185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Britton RG, Kovoor C and Brown K: Direct
molecular targets of resveratrol: Identifying key interactions to
unlock complex mechanisms. Ann N Y Acad Sci. 1348:124–133. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tebbutt N, Pedersen MW and Johns TG:
Targeting the ERBB family in cancer: Couples therapy. Nat Rev
Cancer. 13:663–673. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hanawa M, Suzuki S, Dobashi Y, Yamane T,
Kono K, Enomoto N and Ooi A: EGFR protein overexpression and gene
amplification in squamous cell carcinomas of the esophagus. Int J
Cancer. 118:1173–1180. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fichter CD, Gudernatsch V, Przypadlo CM,
Follo M, Schmidt G, Werner M and Lassmann S: ErbB targeting
inhibitors repress cell migration of esophageal squamous cell
carcinoma and adenocarcinoma cells by distinct signaling pathways.
J Mol Med (Berl). 92:1209–1223. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hoshino M, Fukui H, Ono Y, Sekikawa A,
Ichikawa K, Tomita S, Imai Y, Imura J, Hiraishi H and Fujimori T:
Nuclear expression of phosphorylated EGFR is associated with poor
prognosis of patients with esophageal squamous cell carcinoma.
Pathobiology. 74:15–21. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mitsudomi T and Yatabe Y: Epidermal growth
factor receptor in relation to tumor development: EGFR gene and
cancer. FEBS J. 277:301–308. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Olayioye MA, Neve RM, Lane HA and Hynes
NE: The ErbB signaling network: Receptor heterodimerization in
development and cancer. EMBO J. 19:3159–3167. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han W and Lo HW: Landscape of EGFR
signaling network in human cancers: Biology and therapeutic
response in relation to receptor subcellular locations. Cancer
Lett. 318:124–134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kolch W and Pitt A: Functional proteomics
to dissect tyrosine kinase signalling pathways in cancer. Nat Rev
Cancer. 10:618–629. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang J, Yu JM, Jing SW, Guo Y, Wu YJ, Li
N, Jiao WP, Wang L and Zhang YJ: Relationship between EGFR
over-expression and clinicopathologic characteristics in squamous
cell carcinoma of the esophagus: A meta-analysis. Asian Pac J
Cancer Prev. 15:5889–5893. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aichler M, Motschmann M, Jutting U, Luber
B, Becker K, Ott K, Lordick F, Langer R, Feith M, Siewert JR and
Walch A: Epidermal growth factor receptor (EGFR) is an independent
adverse prognostic factor in esophageal adenocarcinoma patients
treated with cisplatin-based neoadjuvant chemotherapy. Oncotarget.
5:6620–6632. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gan HK, Burgess AW, Clayton AH and Scott
AM: Targeting of a conformationally exposed, tumor-specific epitope
of EGFR as a strategy for cancer therapy. Cancer Res. 72:2924–2930.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lacouture ME: Mechanisms of cutaneous
toxicities to EGFR inhibitors. Nat Rev Cancer. 6:803–812. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schramm L: Going Green: The role of the
green tea component EGCG in chemoprevention. J Carcinog Mutagen.
4:10001422013.PubMed/NCBI
|
|
32
|
Russo GL, Russo M, Spagnuolo C, Tedesco I,
Bilotto S, Iannitti R and Palumbo R: Quercetin: A pleiotropic
kinase inhibitor against cancer. Cancer Treat Res. 159:185–205.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Levkowitz G, Waterman H, Ettenberg SA,
Katz M, Tsygankov AY, Alroy I, Lavi S, Iwai K, Reiss Y, Ciechanover
A, et al: Ubiquitin ligase activity and tyrosine phosphorylation
underlie suppression of growth factor signaling by c-Cbl/Sli-1. Mol
Cell. 4:1029–1040. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ettenberg SA, Keane MM, Nau MM, Frankel M,
Wang LM, Pierce JH and Lipkowitz S: cbl-b inhibits epidermal growth
factor receptor signaling. Oncogene. 18:1855–1866. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin SY, Makino K, Xia W, Matin A, Wen Y,
Kwong KY, Bourguignon L and Hung MC: Nuclear localization of EGF
receptor and its potential new role as a transcription factor. Nat
Cell Biol. 3:802–808. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Brand TM, Iida M, Luthar N, Starr MM,
Huppert EJ and Wheeler DL: Nuclear EGFR as a molecular target in
cancer. Radiother Oncol. 108:370–377. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li A, Zhang C, Gao S, Chen F, Yang C, Luo
R and Xiao H: TIP30 loss enhances cytoplasmic and nuclear EGFR
signaling and promotes lung adenocarcinogenesis in mice. Oncogene.
32:2273–2281, 2281e.1–12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Umemura S, Takekoshi S, Suzuki Y, Saitoh
Y, Tokuda Y and Osamura RY: Estrogen receptor-negative and human
epidermal growth factor receptor 2-negative breast cancer tissue
have the highest Ki-67 labeling index and EGFR expression: Gene
amplification does not contribute to EGFR expression. Oncol Rep.
14:337–343. 2005.PubMed/NCBI
|
|
39
|
Lo HW, Xia W, Wei Y, Ali-Seyed M, Huang SF
and Hung MC: Novel prognostic value of nuclear epidermal growth
factor receptor in breast cancer. Cancer Res. 65:338–348.
2005.PubMed/NCBI
|
|
40
|
Taguchi T: Nuclear translocation of
epidermal growth factor receptor and its relation to
clinicopathological factors in oral squamous cell carcinomas.
Kokubyo Gakkai Zasshi. 81:45–52. 2014.(In Japanese). PubMed/NCBI
|
|
41
|
Lo HW: Nuclear mode of the EGFR signaling
network: Biology, prognostic value, and therapeutic implications.
Discov Med. 10:44–51. 2010.PubMed/NCBI
|
|
42
|
Jaganathan S, Yue P, Paladino DC,
Bogdanovic J, Huo Q and Turkson J: A functional nuclear epidermal
growth factor receptor, SRC and Stat3 heteromeric complex in
pancreatic cancer cells. PLoS One. 6:e196052011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dittmann K, Mayer C and Rodemann HP:
Inhibition of radiation-induced EGFR nuclear import by C225
(Cetuximab) suppresses DNA-PK activity. Radiother Oncol.
76:157–161. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dittmann K, Mayer C, Wanner G, Kehlbach R
and Rodemann HP: The radioprotector O-phospho-tyrosine stimulates
DNA-repair via epidermal growth factor receptor- and DNA-dependent
kinase phosphorylation. Radiother Oncol. 84:328–334. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liccardi G, Hartley JA and Hochhauser D:
Importance of EGFR/ERCC1 interaction following radiation-induced
DNA damage. Clin Cancer Res. 20:3496–3506. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dittmann K, Mayer C, Kehlbach R and
Rodemann HP: Radiation-induced caveolin-1 associated EGFR
internalization is linked with nuclear EGFR transport and
activation of DNA-PK. Mol Cancer. 7:692008. View Article : Google Scholar : PubMed/NCBI
|