Introduction
Radiotherapy is an established treatment modality
for non-small cell lung cancer (NSCLC), which can provide an
effective cure for a proportion of patients (1). Although NSCLC is a moderately
radiation-responsive tumor, local control is still not achieved in
numerous patients, primarily due to intrinsic and acquired
resistance of tumor cells to ionizing radiation (2,3). There are
difficulties in delivering sufficiently high radiotherapy doses to
the tumor due to potential toxicity development in the normal
tissue (4). Therefore, it is
particularly beneficial for radiotherapy of NSCLC to increase the
radiosensitivity of resistant NSCLC cells by using natural or
synthetic compounds.
Local failure of radiotherapy is associated with a
series of factors, where the radiosensitivity of irradiated cells
depends on a complex interplay of nuclear and cytoplasmic signaling
cascades (5–8). Ionizing radiation induces DNA
double-strand breakages (DSBs) and production of free radicals
(FRs) and reactive oxygen species (ROS), which can cause DSBs of
DNA and lipid oxidation of the cellular membrane; these events are
widely acknowledged as the principal determinants of
radiation-induced cell death (9,10).
Intracellular antioxidants such as the tripeptide thiol
L-γ-glutamyl-L-cysteinyl-glycine (glutathione, GSH) play a key role
in the protection of cells against the oxidative stress induced by
FR and ROS (11). GSH, which is
present in all mammalian cells, is the most important intracellular
thiol-based scavenger of FR and ROS (12–14). The
level of GSH is increased in various tumor cells, including NSCLC
cells (15,16). Agents that decrease the cellular
content of GSH could effectively inhibit the DNA damage repair to
increase the response of tumor cells to ionizing radiation
(17,18).
Numerous compounds extracted from natural sources
have been observed to elevate the radiosensitivity of tumor cells
(19,20). Ursolic acid
(3-beta-hydroxy-urs-12-en-28-oic acid, UA) is one of the
pentacyclic triterpenoids that exist widely in the plant kingdom
(Fig. 1) (21). UA is a compound of interest in
oncology research due to its cytotoxicity, its anti-invasive and
anti-migration activities, and its ability to induce cell
differentiation (22,23). UA also interferes with damage repair
of cancer cells and induces apoptosis in cancer cells by regulating
different signaling pathways, including inhibition of Wnt/β-catenin
and activation of the c-Jun N-terminal kinase and the
phosphoinositide 3-kinase/Akt/nuclear factor (NF)-κB signaling
pathways (24–26). Previous studies have demonstrated
sensitization caused by UA towards chemotherapy and radiotherapy
(27–29). With regard to the treatment of NSCLC,
chemosensitization to low doses of UA was found in the ASTC-a-1
cell line (human lung adenocarcinoma cells) through suppression of
NF-κB (27). However, it remains
unclear whether the radiosensitivity of NSCLC cells, particularly
that of cells with refractory response to radiation, could be
increased by UA. In present study, a radioresistant NSCLC cell line
was established by transfecting a mutant HIF-1α (M-HIF-1α) plasmid,
and the radiosensitivity of the parental and radioresistant NSCLC
cells was investigated upon UA pretreatment. In order to
preliminarily analyze the mechanism, the alteration of
intracellular GSH level was also analyzed.
Materials and methods
Cell culture and treatment
The human lung cancer H1299 cell line was kindly
provided by Professor Qinghua Shi (College of Biological Science,
University of Science and Technology of China, Hefei, China). Cells
were maintained in Dulbecco's modified Eagle medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.),
penicillin (100 U/ml) and streptomycin (100 µg/ml) (Sigma-Aldrich,
St. Louis, MO, USA) at 37°C in an incubator containing a humid
atmosphere of 95% air and 5% CO2, and propagated
according to the protocol supplied by the American Type Culture
Collection (Manassas, VA, USA). UA was purchased from Nanjing
Zelang Medical Technology Co., Ltd. (Nanjing, China), dissolved in
dimethyl sulfoxide (DMSO; Sigma-Aldrich) at a stock concentration
of 5 mmol/l and stored at −20°C.
Plasmids and transfection assays
The pcDNA3.0 vector with enhanced green fluorescence
protein (EGFP) was kindly provided by Professor Qinghua Shi. HIF-1α
complementary DNA (cDNA) for with three mutant motifs, including
the prolines at the 402 and 564 sites in the oxygen-dependent
degradation domain (ODDD) of HIF-1α, and the aspartic acid at the
803 site in the C-terminal transactivation domain (CTAD) of HIF-1α,
was purchased from Beijing Zhongyuan Ltd. (Beijing, China). The
M-HIF-1α cDNA was cloned into the pcDNA3.0-EGFP vector to construct
the pcDNA3.0-EGFP-HIF-1α recombinant plasmid. The pcDNA3.0-EGFP
empty vector was used as control. H1299 cells (5×105)
were transfected with 4 µg plasmid DNA using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The neomycin-resistant clones were
selected in DMEM (Gibco; Thermo Fisher Scientific, Inc.) containing
500 µg/ml G418 (Promega Corporation, Madison, WI, USA), and
transferred into a 24-well culture plate with cloning discs
(Sigma-Aldrich). The selected clones were expanded in medium
containing 200 µg/ml G418, and identified by detecting the
messenger RNA and protein expression of M-HIF-1α.
Cell viability assay
The influence of UA on cell growth was determined
using the MTT (Sigma-Aldrich) assay. The parental H1299 cells and
H1299 cells expressing the M-HIF-1α fragment were seeded in 96-well
plates at a density of 5×103 cells/well, and then
treated with various concentrations of UA for 24 h. Next, the
medium was replaced with fresh medium to allow cells to
continuously grow for 72 h. MTT dye was then added to a final
concentration of 50 mg/ml, and the cells were subsequently
incubated for additional 4 h at 37°C. The medium containing
residual MTT dye was carefully aspirated from each of the wells,
and 200 µl dimethyl sulfoxide was added to each well to dissolve
the reduced formazan dye. The survival rates of viable cells were
calculated by comparing the optical absorbance of the culture
exposed to UA treatment with that of the untreated control.
Irradiation
Irradiation was emitted using a 6 MV X-ray linear
accelerator (Varian Inc., Palo Alto, CA, USA) at a dose rate of 250
cGy/min.
Micronucleus (MN) assay
MN frequencies were tested with the
cytokinesis-block technique as a biological end point for the
response of mimetic hypoxia to irradiation (30). Briefly, the cells were exposed to 0.83
µg/ml cytochalasin B (Sigma-Aldrich) for 19–20 h, followed by 75 mM
KCl hypotonic treatment for 1–3 min, and then fixed in situ
with methanol:acetic acid (9:1 v/v) for 30 min. Air-dried cells
were stained with 5% Giemsa for 10 min. MN were scored in
binucleated cells, and the formation of binucleated cells was
measured as the percentage of the total number of cells scored. For
each sample, ≥1,000 binucleated cells were counted. The MN yield
was calculated as the ratio of the number of MN to the number of
binucleated cells scored.
Western blot analysis of HIF-1α
expression
Cells subjected to different treatments were scraped
off from culture flasks and lysed in lysis buffer containing 10%
glycerol, 10 mM Tris-HCl (pH 6.8), 1% sodium dodecyl sulfate (SDS),
5 mM dithiothreitol and 1X complete protease inhibitor cocktail
(Sigma-Aldrich). The Bradford method was used to detect
concentrations of protein in diverse samples. Protein concentration
was measured using an automatic multifunctional microplate reader.
Proteins (50 µg) were separated by 8% SDS-polyacrylamide gel
electrophoresis. The separated proteins were electrophoretically
transferred to polyvinylidene difluoride membranes, which were then
blocked with 5% skim milk in Tris-buffered saline containing 0.1%
Tween 20 at room temperature for 1 h, and then incubated with mouse
anti-HIF-1α antibody (catalog no. ab82832; Abcam, Cambridge, MA,
USA) at a 1:500 dilution overnight at 4°C, followed by goat
anti-mouse immunoglobulin G (catalog no. ab8226; Abcam) for 1 h at
room temperature. Signals were detected with enhanced
chemiluminescence (ECL Plus; GE Healthcare Life Sciences, Chalfont,
UK). An antibody against the microtubule protein tubulin
(anti-tubulin; Abcam) at a 1:1,000 dilution was used as an internal
control to observe the changes in the HIF-1α bands.
Intracellular GSH assay
Following the treatment of triplicate samples of
106 cells with different reagents, the intracellular GSH
content was measured with a glutathione
reductase/5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB) recycling
assay kit obtained from Nanjing Jiancheng Bioengineering Research
Institute (Nanjing, China), following the protocol recommended by
the manufacturer. Briefly, GSH was determined using a reaction
mixture containing 50 µl of cell lysates, 50 µl of 2.4 mM DTNB and
50 µl of 10.64 mU/µl glutathione reductase in the assay buffer (153
mM sodium phosphate and 8.4 mM ethylenediaminetetraacetic acid, pH
7.5). After 5 min incubation at 25°C, the reaction was started by
the addition of 50 µl of reduced nicotinamide adenine dinucleotide
phosphate (NADPH) solution (0.16 mg/ml) in the assay buffer. The
standard and the sample cuvettes were placed into a dual-beam
spectrophotometer, and the increases in absorbance at 412 nm were
followed as a function of time.
Measurement of intracellular ROS
Cell suspension from the different treatmentswas
incubated with 10 µM of 2′,7′-dichloro-dihydro-fluorescein
diacetate (DCFH-DA) for 30 min, and then washed three times with
phosphate-buffered saline for removing excess DCFH probe. Upon
counting the viable cells, the fluorescence intensities of the
cells were observed under an inverted fluorescence microscope
(Nikon Corporation, Tokyo, Japan), at excitation and emission
wavelengths of 488 and 525 nm, respectively.
Statistical analysis
Data are reported as the mean ± standard error of
the mean of three separate experiments unless stated otherwise.
Statistical significance was measured by independent samples t-test
and analysis of variance. P<0.05 was considered to indicate a
statistically significant difference.
Results
Identification of radioresistant NSCLC
cells
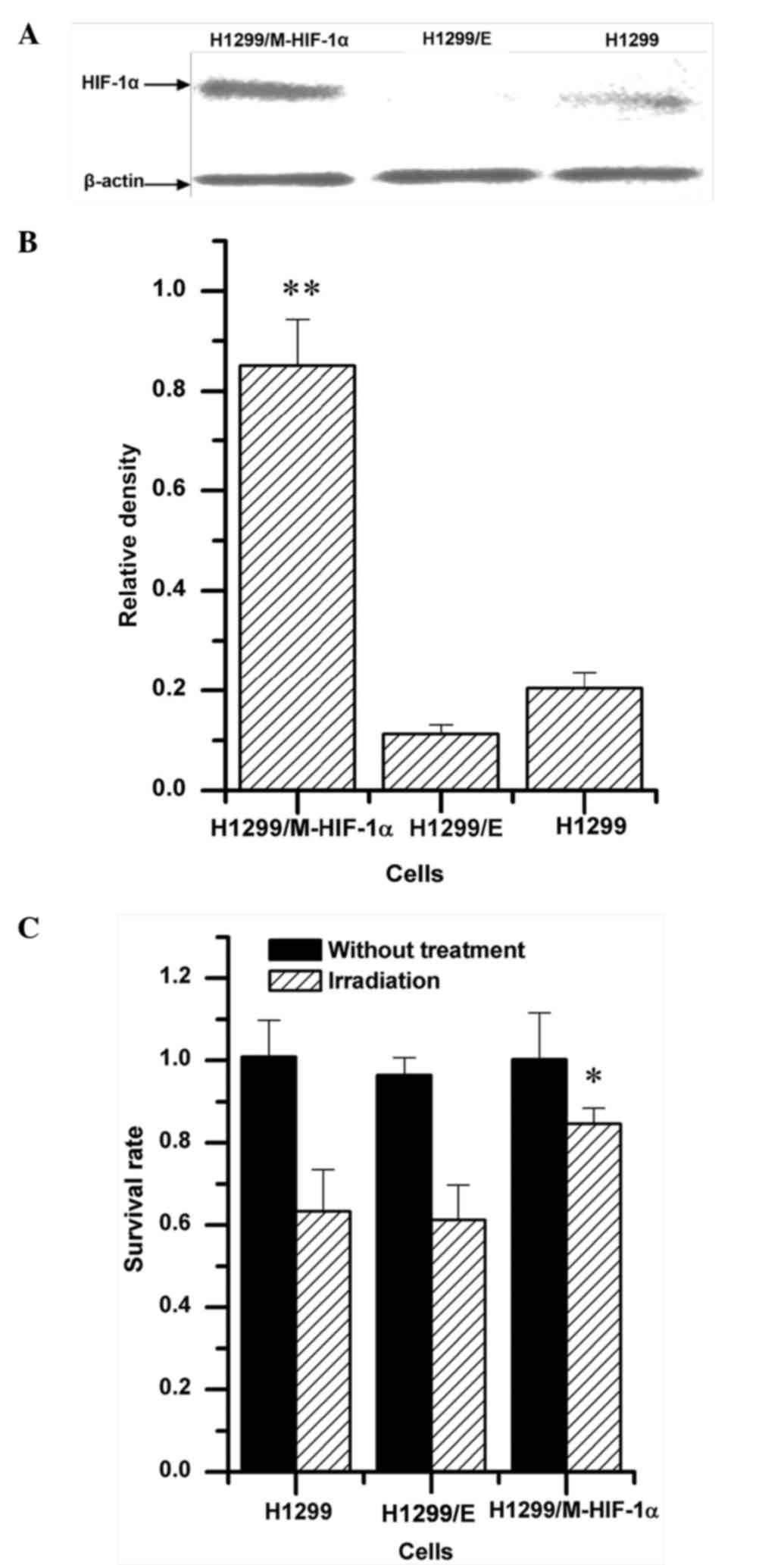
The radiosensitivity of H1299 cells was regulated by
transfecting recombinant pcDNA3.0-EGFP-HIF-1α plasmid. As shown in
Fig. 2A and B, under aerobic
conditions, overexpression of HIF-1α was detected in H1299 cells
transfected with the H1299/M-HIF-1α recombinant plasmid, named
H1299/M-HIF-1α cells. Both the parental H1299 cells and the H1299
cells transfected with empty vector (H1299/E cells) exhibited loss
of HIF-1α expression. Upon exposure to 2 Gy irradiation, cellular
viability was significantly upregulated in H1299/M-HIF-1α cells
compared with that in H1299 and H1299/E cells (Fig. 2C). By contrast, no obvious difference
was observed between H1299 cells and H1299/E cells. It was
therefore demonstrated that H1299/M-HIF-1α cells had lower
radiosensitivity than H1299 cells and H1299/E cells.
Influence of UA on the sensitivity of
resistant NSCLC cells
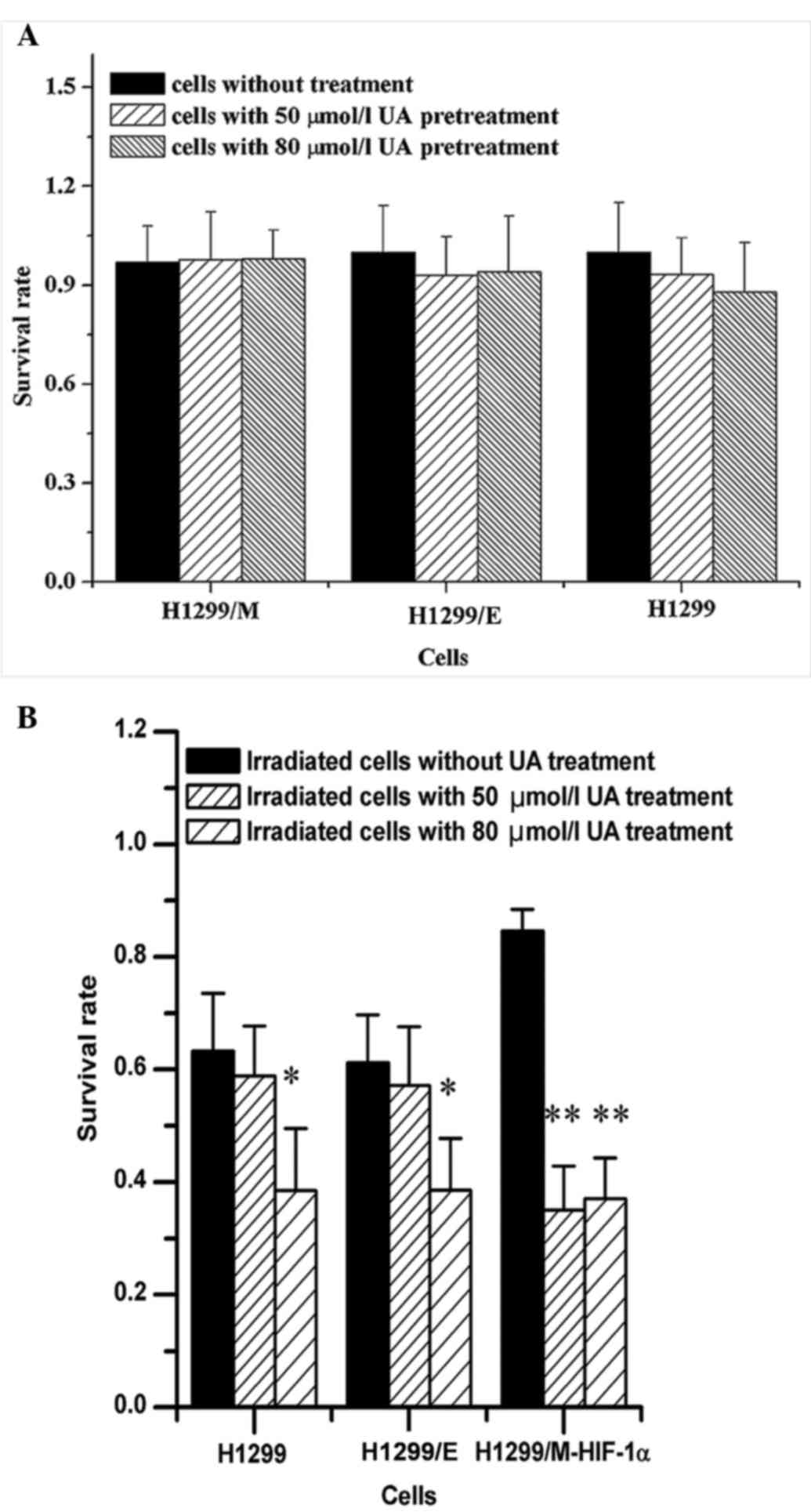
To evaluate the radiosensitizing effect of UA on
NSCLC cells, the experimental concentration of UA was first
selected. The result from the cytotoxicity test indicated that
there were not significant changes in the survival rates of the
three types of NSCLC cells pretreated with UA at 50 and 80 µmol/l,
as shown in Fig. 3A. Alterations in
the survival rates of the cells subjected to treatment with two
different concentrations of UA for 24 h were detected upon
irradiation at 2 Gy. It was observed that UA at two different
concentrations could significantly increase the radiosensitivity of
H1299/M-HIF-1α cells, leading to the reduction of survival rates of
the irradiated cells. However, the radiosensitizing effects of UA
on H1299 and H1299/E cells were only observed at high concentration
(80 µmol/l) (Fig. 3B). Consequently,
it was demonstrated that radioresistant NSCLC cells were the most
sensitive to the UA treatment combined with irradiation.
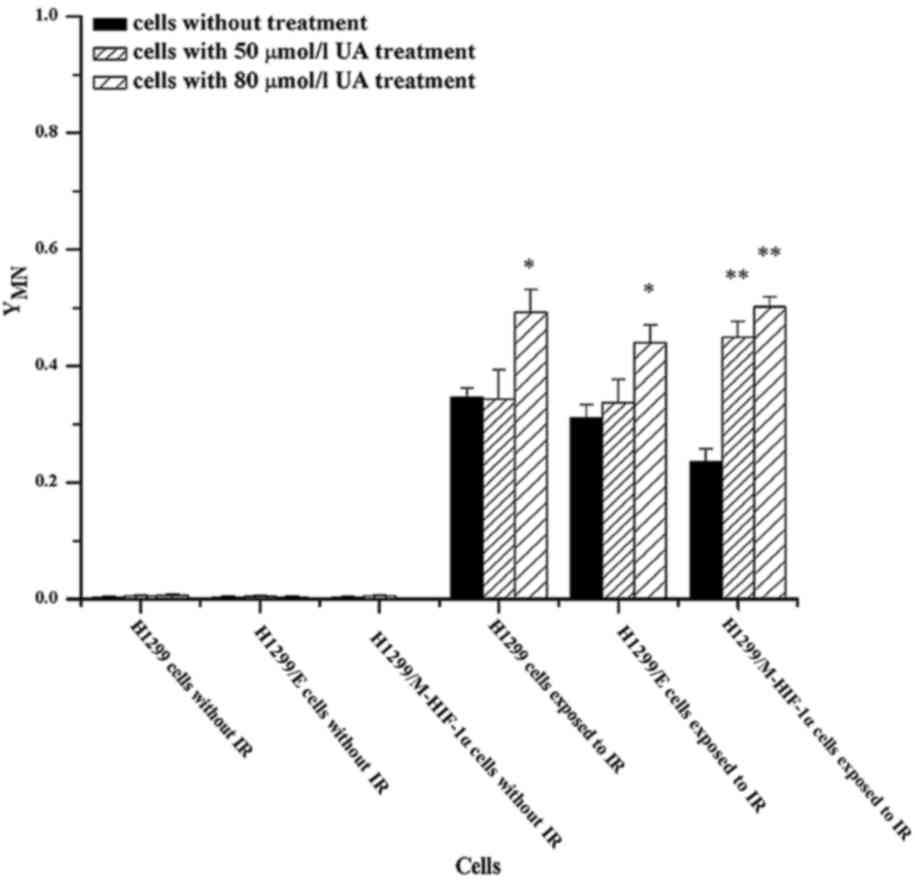
Alteration of MN formation in
irradiated NSCLC cells by UA treatment
The MN frequencies in irradiated cells were further
measured for the assessment of DNA damage. As shown in Fig. 4, when NSCLC cells were not exposed to
ionizing radiation, the intracellular MN frequencies were very low,
even with UA pretreatment. Conversely, the MN ratio increased in
irradiated cells. Subsequently, the addition of UA at 50 and 80
µmol/l could further increase the formation of MN in irradiated
H1299/M-HIF-1α cells. Further elevation of MN frequencies, however,
was not observed in H1299 or H1299/E cells subjected to the
combination treatment of UA at 50 µmol/l concentration and
radiation. It was thus obvious that UA effectively promoted the
formation of MN in irradiated NSCLC cells, particularly in
irradiated H1299/M-HIF-1α cells.
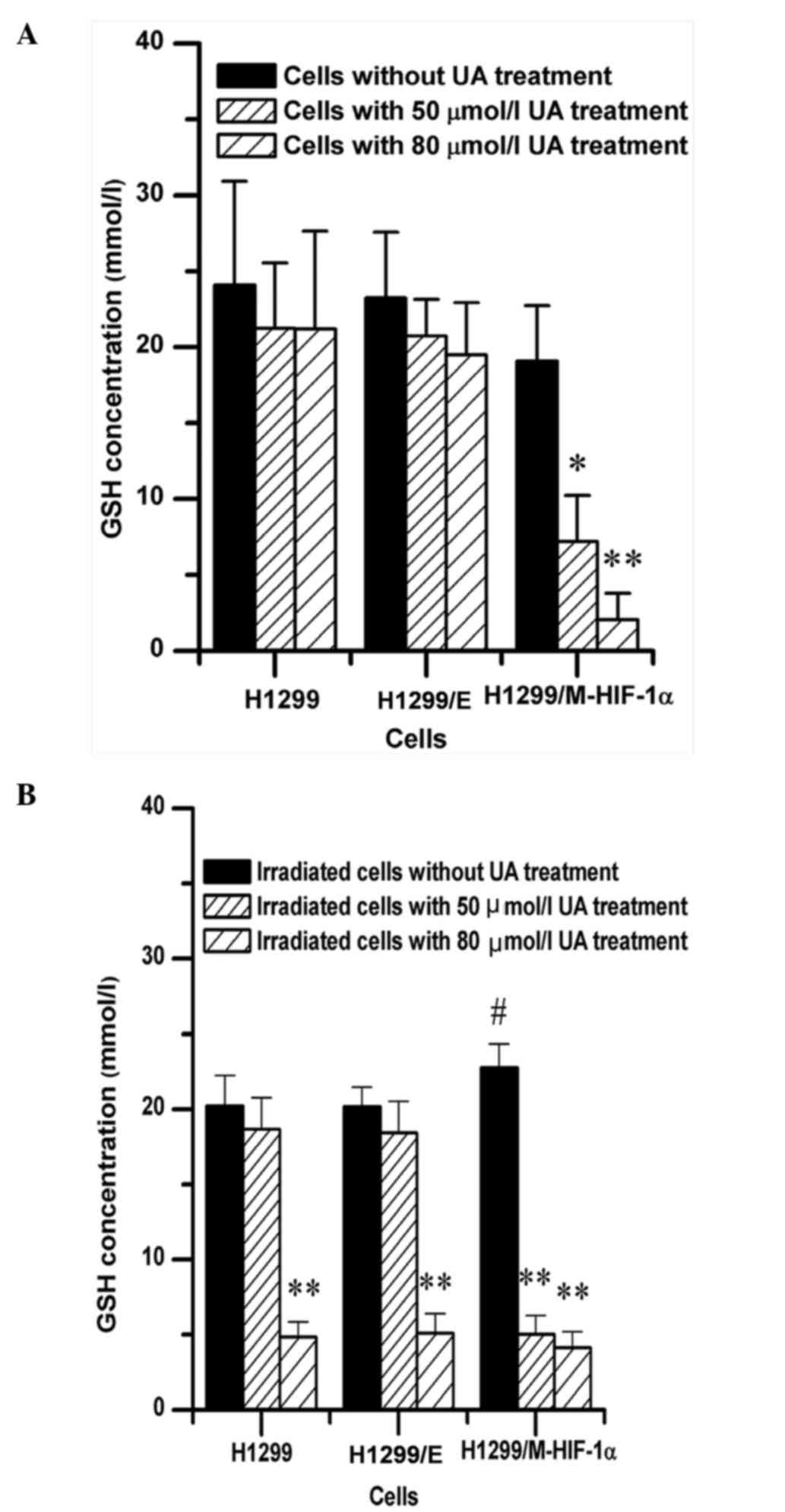
Changes in intracellular GSH content
by UA treatment
Due to the strong radioprotection of endogenous GSH,
the intracellular GSH content was analyzed in NSCLC cells upon
non-exposure or exposure to irradiation following UA pretreatment.
The results revealed that UA could remarkably decrease the
endogenous GSH content in H1299/M-HIF-1α cells not exposed to
radiation but not in H1299 or H1299/E cells, as shown in Fig. 5A. Additionally, following NSCLC cells
exposure to irradiation, the level of cellular GSH in
H1299/M-HIF-1α cells was higher than that in H1299 cell and H1299/E
cells, as shown in Fig. 5B.
Furthermore, UA at high concentration (80 µmol/l) could effectively
attenuate the intracellular GSH content of H1299 and H1299/E cells.
The combination treatment of UA with radiation could decrease the
GSH intracellular contents in H1299/M-HIF-1α cells, both at 50 and
80 µmol/l concentration of UA.
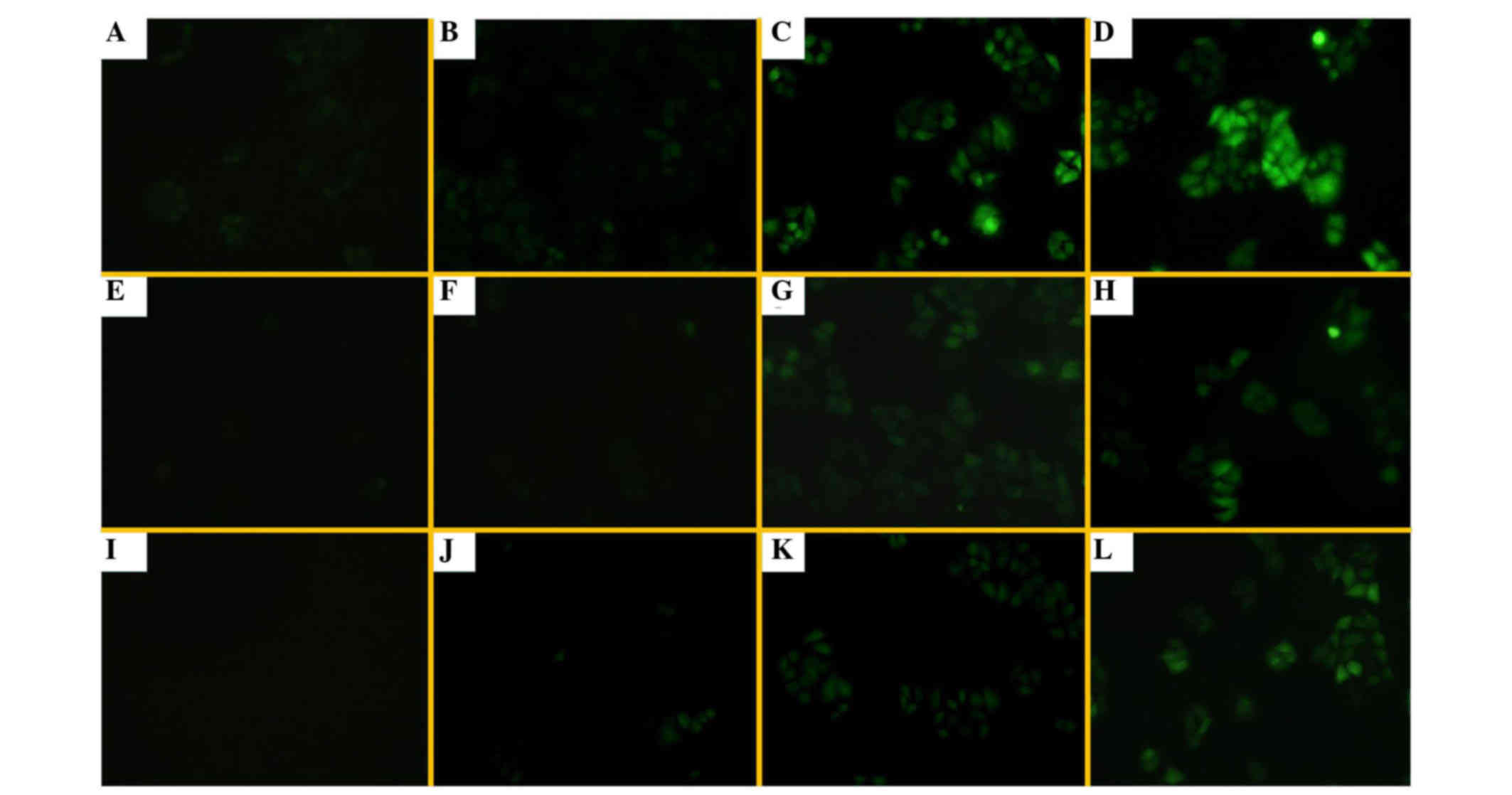
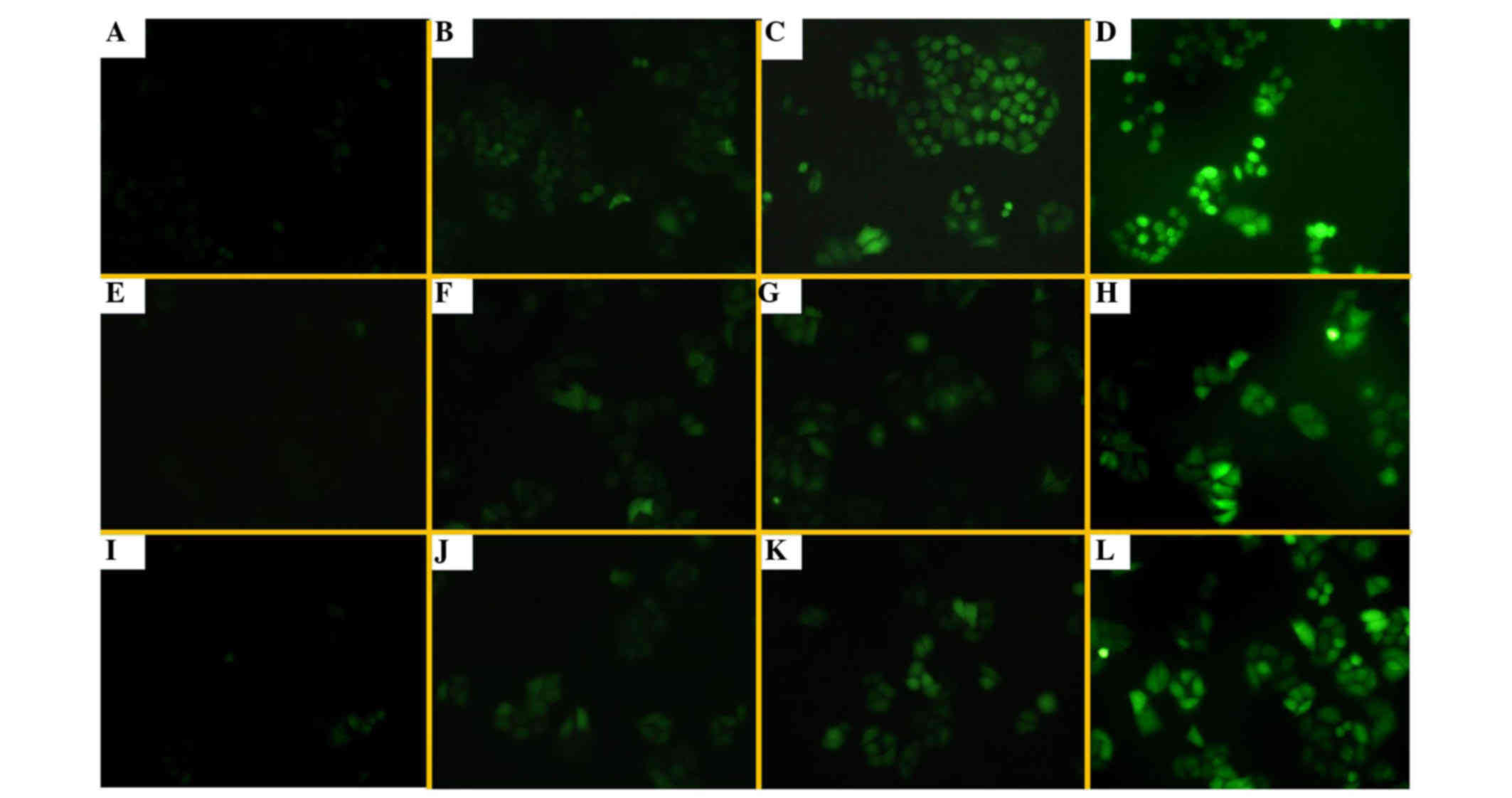
Alteration of intracellular ROS levels
by UA treatment
Under an inverted fluorescence microscope, there was
fluorescence in H1299/E and H1299/M-HIF-1α cells without DCFH-DA
probe treatment, due to the presence of EGFP in the transfected
plasmid (Fig. 6A and E). The
fluorescence intensities in the three groups of cells evaluated
were weakly increased following DCFH-DA treatment. UA at different
concentrations could enhance the levels of intracellular ROS and FR
in these cells, particularly in H1299/M-HIF-1α cells (Fig. 6). The results shown in Fig. 7 further demonstrate that the
combination of UA with radiation treatment significantly enhanced
the generation of ROS and FR. It was revealed that UA with or
without irradiation could promote an increase in cellular ROS and
FR, particularly in H1299/M-HIF-1α cells.
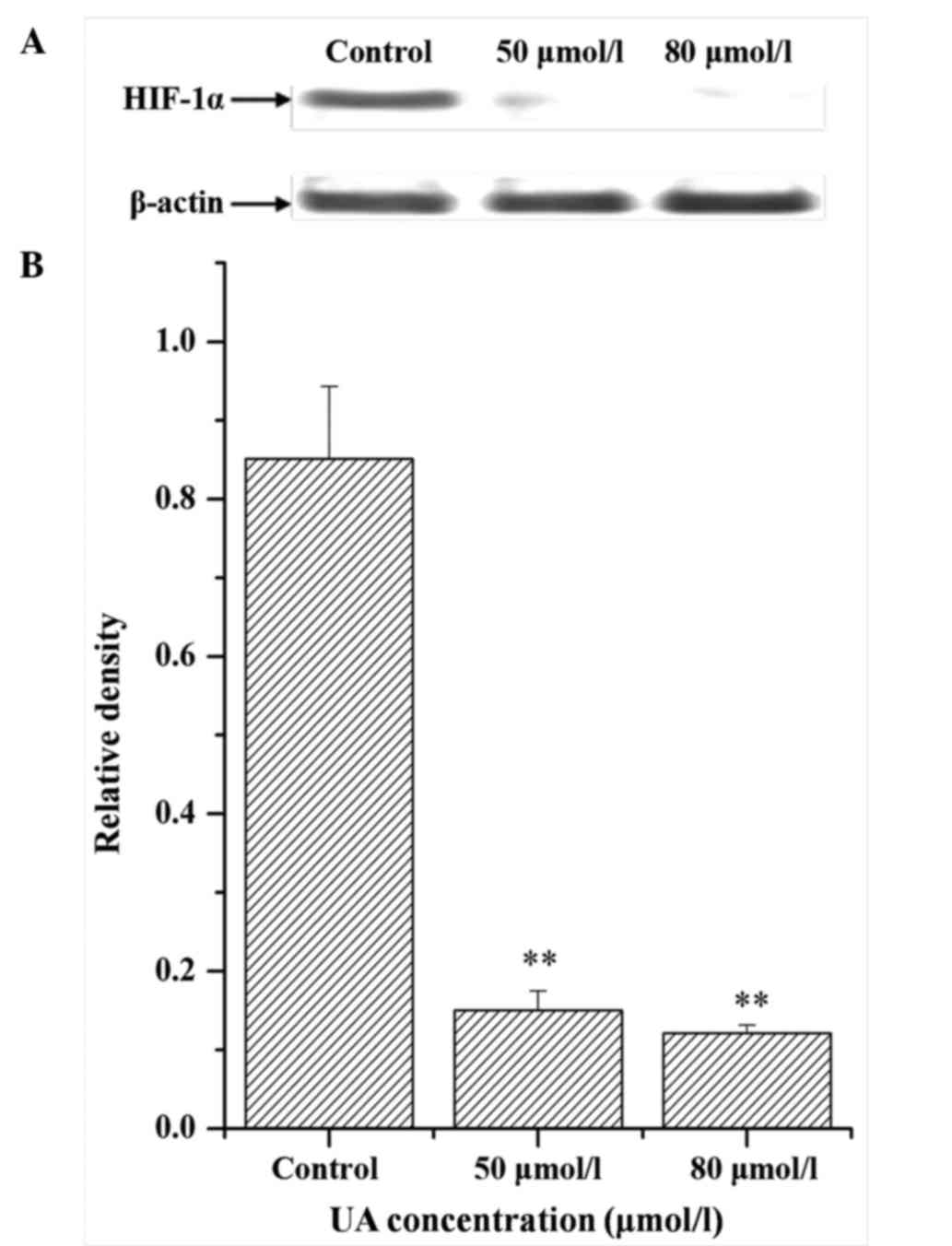
Influence of UA on the expression of
HIF-1α
Besides detection of intracellular GSH, the levels
of HIF-1α in radioresistant cells were investigated upon UA
pretreatment. It was observed that UA at different concentrations
markedly reduced the protein expression levels of HIF-1α, as
detected by western blotting (Fig.
8). The results also revealed that UA could suppress the
expression of M-HIF-1α.
Discussion
In our previous study, it was observed that
oleanolic acid (OA), an isomer of UA, could effectively increase
the radiosensitivity of aerobic and hypoxic A549 cells, a NSCLC
cell line, by inhibition of intracellular GSH synthesis and HIF-1α
expression (31,32). The radiosensitizing efficiency of UA
on NSCLC cells, particularly on radioresistant cells, was also
observed in the present study, due to the higher anti-tumor
activity of UA compared with that of OA (33). In the current study, a radioresistant
cell line was firstly established by transfection with an M-HIF-1α
plasmid. It is known that HIF-1α is closely associated with the
radiosensitivity of tumor cells, including NSCLC cells (34,35).
Numerous studies have shown that, following inhibition of the
master transcription factor triggered in response to hypoxia, the
radiotherapeutic effect on NSCLC may be significantly increased
(36,37). Under aerobic conditions, HIF-1α is
hydroxylated by prolyl hydroxylases at the proline residues 402 and
564 in the ODDD (38). Subsequently,
it is targeted for proteasome-mediated degradation via a protein
ubiquitin ligase complex containing the product of von
Hippel-Lindau tumor suppressor (39).
Another blockage pathway of HIF-1α activity is hydroxylation of
asparagine 803 by factor inhibiting HIF-1 within the CTAD, followed
by interruption of the binding of HIF-1α to the p300/CREB-binding
protein coactivator, thus preventing the transactivation
capabilities of HIF-1 (40). In the
present study, the cDNA of HIF-1α within the pcDNA3.0-EGFP-HIF-1α
recombinant plasmid contained three mutant sites, namely proline
residues 402 and 564, and asparagine 803, which enable HIF-1α to
evade hydroxylation by maintaining HIF-1α expression and
transcriptional activity under aerobic conditions. Thus,
H1299/M-HIF-1α cells exhibited high level of HIF-1α under aerobic
conditions, concomitant with a refractory response to radiation
(41). This in vitro model is
convenient to observe alteration of the sensitivity of resistant
cells to radiation, as HIF-1α need not be induced by physical or
chemical hypoxia.
As a natural antitumor drug, the activity of UA has
been reported in numerous studies (22,23,42).
Previous data reported the sensitization of cancer cells to
ionizing radiation-induced apoptosis by UA, including human
prostate cancer DU145 cells, mouse colon cancer CT26 cells and
mouse melanoma B16F10 cells (28).
The present study demonstrated that UA exerted a similar
radiosensitizing effect on NSCLC cells. Notably, a stronger
sensitizing effect of UA was observed on radioresistant H1299 cells
with high level of HIF-1α expression compared with that observed in
H1299 cells without HIF-1α expression. Similarly, sensitization to
ionizing radiation of radioresistant tumor cells was previously
observed to be effected by other natural or synthetic compounds
(43,44). For instance, Biddlestone-Thorpe et
al (43) reported that glioma
cells with a mutant p53 gene that induced tolerance to radiation
were markedly more sensitive to radiosensitization induced by
KU-60019 (an ataxia telangiectasia mutated kinase inhibitor) than
genetically matched wild-type glioma cells. The mechanism could be
associated with changes in the expression of various genes and
modification of intracellular homeostasis.
There are numerous studies demonstrating that the
numbers and frequencies of intracellular MN represent DNA damage
induced by radiation; thus MN is considered as an appropriate
biological tool to evaluate in vitro radiosensitivity due to
its high reliability and reproducibility (45–47). Based
on the results of the present cell viability assay, intracellular
MN frequencies were detected as a biomarker of DNA damage caused by
ionizing radiation. The results revealed that the intracellular MN
formation was in agreement with the change in tendency observed in
the cell viability assay. Both 50 and 80 µmol/l of UA could
increase the MN frequencies in binucleated H1299/M-HIF-1α cells
following irradiation. By contrast, elevation of MN formation in
irradiated H1299 cells and H1299/E cells was only observed with a
UA concentration of 80 µmol/l. These findings indicate the marked
sensitization caused by UA on radioresistant NSCLC cells.
According to previous studies, the depletion of GSH
content strongly resulted in the production of cellular MN and the
death of irradiated NSCLC cells by increasing the levels of
cellular ROS and FR (31,32,48). Thus,
the present study measured the intracellular GSH content and the
levels of ROS and FR through either UA treatment alone or
combination treatment with UA and irradiation. The results
indicated that treatment of UA alone decreased the GSH content and
increased the levels of ROS and FR in H1299/M-HIF-1α cells, but not
in H1299 cells or H1299/E cells, whereas the combination of UA at
high concentration (80 µmol/l) and irradiation could diminish the
GSH content and increase the levels of ROS and FR in H1299 cells
and H1299/E cells. For H1299/M-HIF-1α cells, either exposed to
irradiation or not, there was a reduction in cellular GSH content,
concomitant with an enhancement in ROS and FR, following treatment
with UA at various concentrations. To explore the mechanism behind
these observations, the expression of HIF-1α in H1299/M-HIF-1α
cells was detected upon UA treatment. It was observed that
increasing HIF-1α expression was inhibited by UA, which is in
agreement with previously reported data (49,50).
According to the study by Guo et al (51), the inhibition of HIF-1α could decrease
the cellular GSH content and increase the generation of ROS by
regulating the level of NADPH, which led to a more oxidizing
environment for the cells. In present study, the GSH content of
H1299/M-HIF-1α cells could further be reduced due to UA-mediated
suppression of HIF-1α expression. Therefore, the increased
sensitization of H1299/M-HIF-1α cells by UA was associated with
reduced HIF-1α expression.
In summary, the present results demonstrated that UA
significantly exerted a radiosensitizing effect on NSCLC cells,
particularly on radioresistant cells overexpressing HIF-1α. The
primary mechanism is explained as follows: i) Radiosensitivity of
NSCLC cells without HIF-1α expression was upregulated through UA,
which decreased the intracellular GSH content; and ii) two
pathways, including the attenuation of GSH and the suppression of
HIF-1α by UA, enhanced the sensitization towards ionizing
radiation-induced cell death of NSCLC cells overexpressing HIF-1α.
UA should therefore be deeply studied as a potential
radiosensitizing reagent for NSCLC radiotherapy.
Acknowledgements
The present study was funded by the Anhui Provincial
Natural Science Foundation of China (Hefei, China; grant no.
11040606M210).
References
|
1
|
Senthi S, Haasbeek CJ, Slotman BJ and
Senan S: Outcomes of stereotactic ablative radiotherapy for central
lung tumours: A systematic review. Radiother Oncol. 106:276–282.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shen Z, Wu X, Wang Z, Li B and Zhu X:
Effect of miR-18a overexpression on the radiosensitivity of
non-small cell lung cancer. Int J Clin Exp Pathol. 8:643–648.
2015.PubMed/NCBI
|
|
3
|
Subtil FS, Wilhelm J, Bill V, Westholt N,
Rudolph S, Fischer J, Scheel S, Seay U, Fournier C, Taucher-Scholz
G, et al: Carbon ion radiotherapy of human lung cancer attenuates
HIF-1 signaling and acts with considerably enhanced therape-utic
efficiency. FASEB J. 28:1412–1421. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cannon DM, Mehta MP, Adkison JB, Khuntia
D, Traynor AM, Tomé WA, Chappell RJ, Tolakanahalli R, Mohindra P,
Bentzen SM and Cannon GM: Dose-limiting toxicity after
hypofractionated dose-escalated radiotherapy in non-small-cell lung
cancer. J Clin Oncol. 31:4343–4348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim E, Youn H, Kwon T, Son B, Kang J, Yang
HJ, Seong KM, Kim W and Youn B: PAK1 tyrosine phosphorylation is
required to induce epithelial-mesenchymal transition and
radioresistance in lung cancer cells. Cancer Res. 74:5520–5531.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sekhar KR, Benamar M, Venkateswaran A,
Sasi S, Penthala NR, Crooks PA, Crooks PA, Hann SR, Geng L, Balusu
R, et al: Targeting nucleophosmin 1 represents a rational strategy
for radiationsensitization. Int J Radiat Oncol Biol Phys.
89:1106–1114. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Theys J, Yahyanejad S, Habets R, Span P,
Dubois L, Paesmans K, Kattenbeld B, Cleutjens J, Groot AJ,
Schuurbiers OC, et al: High NOTCH activity induces radiation
resistance in non small cell lung cancer. Radiother Oncol.
108:440–445. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okaichi K, Nose K, Kotake T, Izumi N and
Kudo T: Phosphorylation of p53 modifies sensitivity to ionizing
radiation. Anticancer Res. 31:2255–2258. 2011.PubMed/NCBI
|
|
9
|
Jamal M, Rath BH, Williams ES, Camphausen
K and Tofilon PJ: Microenvironmental regulation of glioblastoma
radioresponse. Clin Cancer Res. 16:6049–6059. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ayouaz A, Raynaud C, Heride C, Revaud D
and Sabatier L: Telomeres: Hallmarks of radiosensitivity.
Biochimie. 90:60–72. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Quintana-Cabrera R and Bolaños JP:
Glutathione and γ-glutamylcysteine in the antioxidant and survival
functions of mitochondria. Biochem Soc Trans. 41:106–110. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pereira CV, Nadanaciva S, Oliveira PJ and
Will Y: The contribution of oxidative stress to drug-induced organ
toxicity and its detection in vitro and in vivo. Expert Opin Drug
Metab Toxicol. 8:219–237. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Slemmer JE, Shacka JJ, Sweeney MI and
Weber JT: Antioxidants and free radical scavengers for the
treatment of stroke, traumatic brain injury and aging. Curr Med
Chem. 15:404–414. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wells PG, Bhuller Y, Chen CS, Jeng W,
Kasapinovic S, Kennedy JC, Kim PM, Laposa RR, McCallum GP, Nicol
CJ, et al: Molecular and biochemical mechanisms in teratogenesis
involving reactive oxygen species. Toxicol Appl Pharmacol 207 (2
Suppl). 354–366. 2005. View Article : Google Scholar
|
|
15
|
Ogunrinu TA and Sontheimer H: Hypoxia
increases the dependence of glioma cells on glutathione. J Biol
Chem. 285:37716–37724. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Honda T, Coppola S, Ghibelli L, Cho SH,
Kagawa S, Spurgers KB, Brisbay SM, Roth JA, Meyn RE, Fang B and
McDonnell TJ: GSH depletion enhances adenoviral bax-induced
apoptosis in lung cancer cells. Cancer Gene Ther. 11:249–255. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee JH, Tak JK, Park KM and Park JW:
N-t-Butyl hydroxylamine regulates ionizing radiation-induced
apoptosis in U937 cells. Biochimie. 89:1509–1516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Karthikeyan S, Kanimozhi G, Prasad NR and
Mahalakshmi R: Radiosensitizing effect of ferulic acid on human
cervical carcinoma cells in vitro. Toxicol In Vitro. 25:1366–1375.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Keshmiri-Neghab H, Goliaei B and Nikoofar
A: Gossypol enhances radiation induced autophagy in glioblastoma
multiforme. Gen Physiol Biophys. 33:433–442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Elbaz HA, Lee I, Antwih DA, Liu J,
Hüttemann M and Zielske SP: Epicatechin stimulates mitochondrial
activity and selectively sensitizes cancer cells to radiation. PLoS
One. 9:e883222014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zang LL, Wu BN, Lin Y, Wang J, Fu L and
Tang ZY: Research progress of ursolic acid's anti-tumor actions.
Chin J Integr Med. 20:72–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang W, Zhao C, Jou D, Lü J, Zhang C, Lin
L and Lin J: Ursolic acid inhibits the growth of colon
cancer-initiating cells by targeting STAT3. Anticancer Res.
33:4279–4284. 2013.PubMed/NCBI
|
|
23
|
Huang CY, Lin CY, Tsai CW and Yin MC:
Inhibition of cell proliferation, invasion and migration by ursolic
acid in human lung cancer cell lines. Toxicol In Vitro.
25:1274–1280. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park JH, Kwon HY, Sohn EJ, Kim KA, Kim B,
Jeong SJ, Song JH, Koo JS and Kim SH: Inhibition of Wnt/β-catenin
signaling mediates ursolic acid-induced apoptosis in PC-3 prostate
cancer cells. Pharmacol Rep. 65:1366–1374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xavier CP, Lima CF, Pedro DF, Wilson JM,
Kristiansen K and Pereira-Wilson C: Ursolic acid induces cell death
and modulates autophagy through JNK pathway in apoptosis-resistant
colorectal cancer cells. J Nutr Biochem. 24:706–712. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li J, Liang X and Yang X: Ursolic acid
inhibits growth and induces apoptosis in gemcitabine-resistant
human pancreatic cancer via the JNK and PI3K/Akt/NF-κB pathways.
Oncol Rep. 28:501–510. 2012.PubMed/NCBI
|
|
27
|
Li Y, Xing D, Chen Q and Chen WR:
Enhancement of chemotherapeutic agent-induced apoptosis by
inhibition of NF-kappaB using ursolic acid. Int J Cancer.
127:462–473. 2010.PubMed/NCBI
|
|
28
|
Koh SJ, Tak JK, Kim ST, Nam WS, Kim SY,
Park KM and Park JW: Sensitization of ionizing radiation-induced
apoptosis by ursolic acid. Free Radic Res. 46:339–345. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huq F, Yu JQ, Beale P, Chan C, Arzuman L,
Nessa MU and Mazumder ME: Combinations of platinums and selected
phytochemicals as a means of overcoming resistance in ovarian
cancer. Anticancer Res. 34:541–545. 2014.PubMed/NCBI
|
|
30
|
Romm H, Barnard S, Boulay-Greene H, De
Amicis A, De Sanctis S, Franco M, Herodin F, Jones A, Kulka U,
Lista F, et al: Laboratory intercomparison of the cytokinesis-block
micronucleus assay. Radiat Res. 180:120–128. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qi R and Jin W, Wang J, Yi Q, Yu M, Xu S
and Jin W: Oleanolic acid enhances the radiosensitivity of tumor
cells under mimetic hypoxia through the reduction in intracellular
GSH content and HIF-1α expression. Oncol Rep. 31:2399–2406.
2014.PubMed/NCBI
|
|
32
|
Wang J, Yu M, Xiao L, Xu S, Yi Q and Jin
W: Radiosensitizing effect of oleanolic acid on tumor cells through
the inhibition of GSH synthesis in vitro. Oncol Rep. 30:917–924.
2013.PubMed/NCBI
|
|
33
|
Shan JZ, Xuan YY, Ruan SQ and Sun M:
Proliferation-inhibiting and apoptosis-inducing effects of ursolic
acid and oleanolic acid on multi-drug resistance cancer cells in
vitro. Chin J Integr Med. 17:607–611. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Berghoff AS, Ilhan-Mutlu A, Wöhrer A,
Hackl M, Widhalm G, Hainfellner JA, Dieckmann K, Melchardt T, Dome
B, Heinzl H, et al: Prognostic significance of Ki67 proliferation
index, HIF1 alpha index and microvascular density in patients with
non-small cell lung cancer brain metastases. Strahlenther Onkol.
190:676–685. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yoshimura M, Itasaka S, Harada H and
Hiraoka M: Microenvironment and radiation therapy. Biomed Res Int.
2013:6853082013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
van Baardwijk A, Dooms C, van Suylen RJ,
Verbeken E, Hochstenbag M, Dehing-Oberije C, Rupa D, Pastorekova S,
Stroobants S, Buell U, et al: The maximum uptake of
(18)F-deoxyglucose on positron emission tomography scan correlates
with survival, hypoxia inducible factor-1alpha and GLUT-1 in
non-small cell lung cancer. Eur J Cancer. 43:1392–1398. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zeng L, Ou G, Itasaka S, Harada H, Xie X,
Shibuya K, Kizaka-Kondoh S, Morinibu A, Shinomiya K and Hiraoka M:
TS-1 enhances the effect of radiotherapy by suppressing
radiation-induced hypoxia-inducible factor-1 activation and
inducing endothelial cell apoptosis. Cancer Sci. 99:2327–2335.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rolfo A, Many A, Racano A, Tal R,
Tagliaferro A, Ietta F, Wang J, Post M and Caniggia I:
Abnormalities in oxygen sensing define early and late onset
preeclampsia as distinct pathologies. PLoS One. 5:e132882010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Semenza GL: HIF-1 mediates metabolic
responses to intratumoral hypoxia and oncogenic mutations. J Clin
Invest. 123:3664–3671. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jin WS, Kong ZL, Shen ZF, Jin YZ, Zhang WK
and Chen GF: Regulation of hypoxia inducible factor-1α expression
by the alteration of redox status in HepG2 cells. J Exp Clin Cancer
Res. 30:612011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wei L, Lu J, Feng L, Li S, Shan J and Li
Y: Construction of recombinant adenovirus vector containing a
modified gene that codes for human hypoxia-inducible factor-1alpha
without oxygen-dependent degradation. Plasmid. 63:20–26. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kassi E, Sourlingas TG, Spiliotaki M,
Papoutsi Z, Pratsinis H, Aligiannis N and Moutsatsou P: Ursolic
acid triggers apoptosis and Bcl-2 downregulation in MCF-7 breast
cancer cells. Cancer Invest. 27:723–733. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Biddlestone-Thorpe L, Sajjad M, Rosenberg
E, Beckta JM, Valerie NC, Tokarz M, Adams BR, Wagner AF, Khalil A,
Gilfor D, et al: ATM kinase inhibition preferentially sensitizes
p53-mutant glioma to ionizing radiation. Clin Cancer Res.
19:3189–3200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim JC, Ali MA, Nandi A, Mukhopadhyay P,
Choy H, Cao C and Saha D: Correlation of HER1/EGFR expression and
degree of radiosensitizing effect of the HER1/EGFR-tyrosine kinase
inhibitor erlotinib. Indian J Biochem Biophys. 42:358–365.
2005.PubMed/NCBI
|
|
45
|
Kashino G, Prise KM, Suzuki K, Matsuda N,
Kodama S, Suzuki M, Nagata K, Kinashi Y, Masunaga S, Ono K and
Watanabe M: Effective suppression of bystander effects by DMSO
treatment of irradiated CHO cells. J Radiat Res. 48:327–333. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Thierens H and Vral A: The micronucleus
assay in radiation accidents. Ann Ist Super Sanita. 45:260–264.
2009.PubMed/NCBI
|
|
47
|
Romm H, Barnard S, Boulay-Greene H, De
Amicis A, De Sanctis S, Franco M, Herodin F, Jones A, Kulka U,
Lista F, et al: Laboratory intercomparison of the cytokinesis-block
micronucleus assay. Radiat Res. 180:120–128. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Singh A, Bodas M, Wakabayashi N, Bunz F
and Biswal S: Gain of Nrf2 function in non-small-cell lung cancer
cells confers radioresistance. Antioxid Redox Signal. 13:1627–1637.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Das B, Yeger H, Tsuchida R, Torkin R, Gee
MF, Thorner PS, Shibuya M, Malkin D and Baruchel S: A
hypoxia-driven vascular endothelial growth factor/Flt1 autocrine
loop interacts with hypoxia-inducible factor-1alpha through
mitogen-activated protein kinase/extracellular signal-regulated
kinase 1/2 pathway in neuroblastoma. Cancer Res. 65:7267–7275.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lin CC, Huang CY, Mong MC, Chan CY and Yin
MC: Antiangiogenic potential of three triterpenic acids in human
liver cancer cells. J Agric Food Chem. 59:755–762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Guo S, Miyake M, Liu KJ and Shi H:
Specific inhibition of hypoxia inducible factor 1 exaggerates cell
injury induced by in vitro ischemia through deteriorating cellular
redox environment. J Neurochem. 108:1309–1321. 2009. View Article : Google Scholar : PubMed/NCBI
|