Introduction
Resveratrol (molecular formula,
C14H12O3; CAS number, 501–36-0) is
a non-toxic phytoalexin antioxidant and an effective anticancer
compound that can be extracted from grapes, red wines, berries and
peanuts (1), which imparts cancer
chemopreventive and therapeutic responses (2,3). It is
suggested to have potent antitumor properties against numerous
human cancers (4,5). The renin-angiotensin system (RAS) is
classically described as an important endocrine system that
regulates blood pressure and electrolyte balance. Studies have
demonstrated that RAS may be involved in numerous
pathophysiological processes, such as maintaining blood
pressure/blood volume homeostasis and ion-fluid balance,
phylogenetic advancement, growth promotion and angiogenesis,
ontogeny and phylogeny (6). RAS is
also involved in several cancers, including glioblastoma multiform
(7), and bladder (8) and renal (9) cancers. Previous studies have suggested
that resveratrol may play anticancer roles via a RAS-dependent
pathway, such as in renal (10) and
bladder (8) cancer cells. Angiotensin
II (AngII), which is known to be the main effector of the RAS
pathway, has an important role in regulating cancer inflammation
and tumor angiogenesis (11). The
majority of AngII effects are mediated by 2 specific receptors
subtypes, consisting of AngII type 1 receptor (AT1R) and AngII type
2 receptor (AT2R) (12), which are
detected in astrocytomas (13),
esophageal squamous cell carcinoma (14) and renal clear cell carcinoma (15). AT1R-mediated AngII activity plays a
central role in mediating the majority of the actions of the RAS
pathway (16). Vascular endothelial
growth factor (VEGF) and epidermal growth factor receptor (EGFR)
are important molecules in tumor-associated angiogenesis, as they
activate endothelial cell metastasis and increase vascular
permeability (17–19). Inhibition of VEGF suppresses tumor
angiogenesis and tumor growth in vivo (20). As an important downstream regulator in
RAS, VEGF is reported to be induced by AngII, by binding AT1R in
pancreatic cancer cells (21).
Cyclooxygenase-2 (COX-2) is a key enzyme involved in carcinogenesis
and tumor progression, particularly in neoangiogenesis and
lymphovascular invasion (22–24), and has also been detected as
upregulated through mechanisms distinct from the VEGF axis
(25–27). In the present study, the suppression
of proliferation and induction of apoptosis by resveratrol was
investigated in renal carcinoma cell lines. The levels of the 4
important factors in RAS, consisting of AngII, AT1R, VEGF and COX-2
were examined, and the potential mechanisms were analyzed.
Materials and methods
Cell culture and resveratrol
treatment
Human renal carcinoma ACHN and A498 cell lines were
purchased from Guangzhou Jennio Biological Technology Co., Ltd
(Guangzhou, China). ACHN cells were cultured in high glucose
Dulbecco's modified Eagle medium (Gibco, Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% (v/v) fetal bovine
serum (FBS; Gibco, Thermo Fisher Scientific, Inc.). A498 cells were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% (v/v) FBS. The two cell lines were
cultured at 37°C in a 5% humidified CO2 atmosphere.
Resveratrol was purchased from Sigma-Aldrich; Merck Millipore
(Darmstadt, Germany). Resveratrol was dissolved in DMSO to create a
stock solution at a concentration of 100 mM, which was subsequently
diluted in culture medium to the desired concentration for
experiments. DMSO was used as the vehicle control.
Enzyme-linked immunosorbent assay
(ELISA)
Subsequent to ACHN and A498 cells being treated with
resveratrol, the medium was collected by centrifugation at 400 ×
g for 15 min at 4°C, and the liquid supernatant was stored
at −80°C until ELISA. ACHN and A498 cells were lysed with sodium
dodecyl sulfate (SDS) lysis buffer (Beyotime Institute of
Biotechnology, Shanghai, China), and total proteins were extracted
at 4°C. The concentrations of AngII, AT1R, VEGF and COX-2 in the
culture medium and cells were determined using a human AngII ELISA
kit (catalogue no., ml003766; Shanghai Enzyme-linked Biotechnology
Co., Ltd., Shanghai, China), an AT1R ELISA kit (catalogue no.,
ml006788; Shanghai Enzyme-linked Biotechnology Co., Ltd.), a VEGF
ELISA kit (catalogue no., ml009877; Shanghai Enzyme-linked
Biotechnology Co., Ltd.) and a COX-2 ELISA Kit (catalogue no.,
ml006532; Shanghai Enzyme-linked Biotechnology Co., Ltd.) in
accordance with the manufacturers' protocols.
Cell proliferation assay
Effects of resveratrol treatment on the cell
proliferation of renal carcinoma cells were detected by Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Mashikimachi, Kumamoto, Japan) assay. ACHN and A498 cells were
seeded in 96-well plates (Corning Incorporated, Corning, NY, USA)
at a density of 5×103 cells per well with 100 µl culture
medium. Subsequent to allowing 24 h for adhering, resveratrol was
added at varying concentrations for 12, 24 and 48 h. The culture
medium was then removed and replaced with 100 µl medium containing
CCK-8 reagent (10 µl; Dojindo Molecular Technologies, Inc.) in each
well. The plates were incubated at 37°C for 2 h. Absorbance at 450
nm was recorded using a spectrophotometer (EnSpire 2300 Multilabel
Reader; PerkinElmer, Waltham, MA, USA).
Apoptosis detection
Apoptosis cells were quantified using a fluorescein
isothiocyanate Annexin V Apoptosis Detection Kit (BD Pharmingen,
San Diego, CA, USA) according to the manufacturer's protocol. Cells
were cultured in 6-well plates at a density of 1×105
cells per well. Following 24 h growth, cells were treated with
various resveratrol concentrations and harvested for the apoptosis
assay. Untreated cells were used as a negative control.
Colony formation assay
Cells (1×103 cells per well) were seeded
into 6-well plates subsequent to 6 h-treatment with various
concentrations of resveratrol and were cultured for 2 weeks. The
number of colonies formed was counted subsequent to cells being
fixed with 4% paraformaldehyde, and stained with a crystal violet
staining solution (Beyotime Institute of Biotechnology).
Caspase-Glo 3/7 assays
Cells (5×103 cells per well) were seeded
onto 96-well plates and exposed to different concentrations of
resveratrol. Equal volume of Caspase-Glo 3/7 reagent was
subsequently added into each well and incubated for 30 min at room
temperature in the dark. The luminescence was determined using a
luminometer (Berthold Sirius L; Titertek-Berthold, Pforzheim,
Germany).
Cytotoxicity assay
The cytotoxicity of resveratrol was assessed using a
lactate dehydrogenase (LDH) Cytotoxicity Assay kit (Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. Cells were cultured in 96-well plates (Corning
Incorporated) and then treated with resveratrol for 24 h. The
medium was collected by centrifugation at 400 × g for 5 min.
Supernatant (120 µl/well) was transferred into another 96-well
plate and 60 µl LDH detection reagent was added to each well, and
then incubated for 30 min at room temperature in the dark.
Absorbance was recorded at 490 nm with a spectrophotometer (EnSpire
2300 Multilabel Reader; PerkinElmer).
Western blot analysis
The expression of the apoptosis-associated proteins
caspase 9, B-cell lymphoma 2 (Bcl-2) and Bcl-2-like protein 4 (Bax)
were detected in renal carcinoma cells. Cells were lysed with
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology) and then total proteins were extracted at 4°C.
Proteins were separated by SDS-polyacrylamide gel electrophoresis
and transferred onto a polyvinylidene fluoride membrane (EMD
Millipore, Billerica, MA, USA). Membranes were then blocked using
5% non-fat milk in Tris-buffered saline and Tween 20 (TBS-T) at
room temperature for 1 h, and the membranes were then probed with
rabbit anti-human caspase 9 (catalog no., 9502; dilution, 1:1,000;
Cell Signaling Technology, Inc., Danvers, MA, USA), Bax (catalog
no., 2772; dilution, 1:1,000; Cell Signaling Technology, Inc.) and
Bcl-2 rabbit monoclonal antibodies (catalog no., 2876; dilution,
1:1000; Cell Signaling Technology, Inc.). The membranes were probed
with a rabbit anti-human GAPDH polyclonal antibody (catalog no.,
ab37168; dilution, 1:100,000; Abcam, Cambridge, UK) as a loading
control. The membranes were washed 3 times with TBS-T for 5 min
each time and incubated for 1.5 h with a horseradish
peroxidase-conjugated goat anti-rabbit secondary antibody (catalog
no., E030120; dilution, 1:10,000; EarthOx Life Sciences, Millbrae,
CA, USA).
Statistical analysis
ELISA, colony formation and Caspase-Glo 3/7 assays
were repeated 3 times; CCK-8 and cytotoxicity assays were performed
4 times. One-way analysis of variance (SPSS 18.0; SPSS, Inc.,
Chicago, IL, USA) and Student's t-test (Microsoft Excel;
Redmond, WA, USA) were used to evaluate the differences between 2
groups of data in all the experiments. All data were presented as
the mean ± standard deviation (SD). P<0.05 was considered to
indicate a statistically significant difference and P<0.01 was
considered to indicate an extremely significant difference.
Results
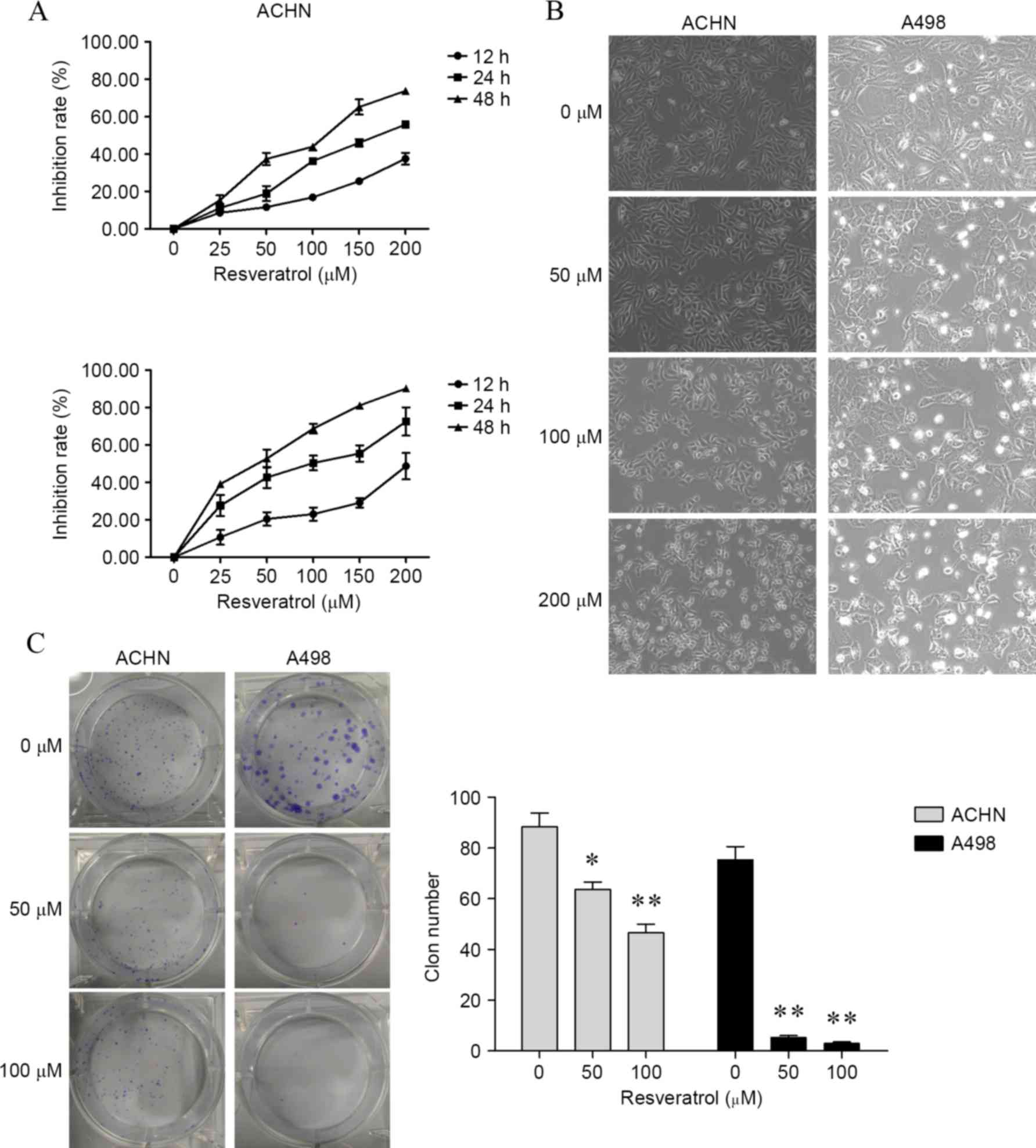
Resveratrol suppressed growth of renal
carcinoma cells
To determine the effect of resveratrol on cell
proliferation in renal carcinoma cells, the present study firstly
determined cell viability by CCK-8 assay. As shown in Fig. 1A, renal carcinoma cells treated with
25, 50, 100, 150 and 200 resveratrol for 12, 24 and 48 h, cell
viability was significantly inhibited in a dose- and time-dependent
manner compared with cells treated with 0 µM resveratrol [ACHN
cells, P<0.001 for all concentrations at 12, 24 and 48 h; A498
cells, P<0.001 for all concentrations at 12, 24 and 48 h, with
the exception of 25 µM for 12 h (P=0.0017)]. For morphology
analysis, untreated renal carcinoma cells grew well, whereas the
cells treated with resveratrol were distorted in shape and became
round and underwent apoptosis (Fig.
1B). Furthermore, following 6 h of treatment with resveratrol,
colony formation assay was performed and revealed a marked decrease
in colony formation compared to the control group (ACHN cells,
P=0.0158 by 50 µM and P=0.0026 by 100 µM; A498 cells, P=0.0002 by
50 µM and P=0.0002 by 100 µM; Fig.
1C).
Resveratrol induced apoptosis in renal
carcinoma cells
To illustrate the functions of resveratrol on
apoptosis in renal carcinoma cells, cytotoxicity, caspase 3/7
activity, cell apoptosis and expression of caspase 9, Bcl-2 and Bax
were determined. It was found that resveratrol significantly
increased cytotoxicity (ACHN cells: 25 µM, P<0.001; 50 µM,
P=0.0036; 100 µM, P=0.0314; 150 µM, P=0.0034; and 200 µM, P=0.0007;
A498 cells: 25 µM, P<0.001; 50 µM, P=0.0041; 100 µM, P=0.0327;
150 µM, P=0.0032; and 200 µM P=0.0005; Fig. 2A) and caspase 3/7 activity (ACHN
cells, P<0.001 for 50, 100 and 200 µM; A498 cells, P<0.001
for 50, 100 and 200 µM; Fig. 2B) in
renal cancer cells compared with cells treated with 0 µM
resveratrol. Results of flow cytometry assay revealed that
subsequent to 24 h of treatment with resveratrol, apoptosis was
induced in renal carcinoma cells (Fig.
2C). In addition, resveratrol was shown to upregulate
cleaved-caspase 9 and Bax levels and downregulate Bcl-2 level in
the two cell lines (Fig. 2D).
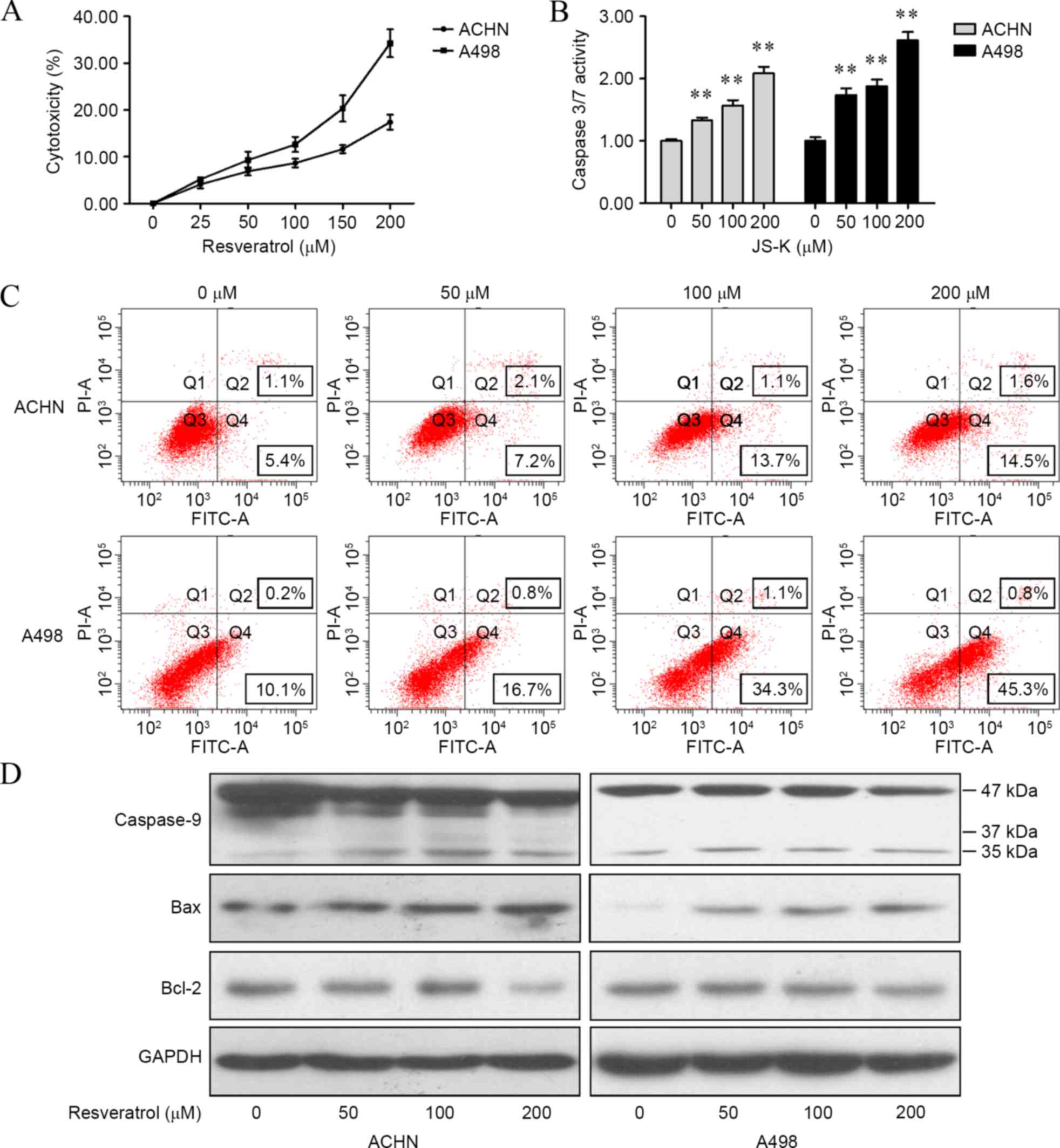 | Figure 2.Resveratrol induced apoptosis in
renal carcinoma cells. (A) Cytotoxicity of resveratrol was
determined by a lactate dehydrogenase assay. (B) Caspase 3/7
activity subsequent to resveratrol treatment was determined by a
Caspase-Glo 3/7 assay. (C) Resveratrol-induced apoptosis in renal
carcinoma cells with different concentrations (0, 50, 100 and 200
µM) for 24 h, which was analyzed by flow cytometry. (D) Cells were
treated with resveratrol (0, 50, 100 and 200 µM) for 24 h, and the
levels of caspase 9, Bax and Bcl-2 were detected by western
blotting. The data showed that resveratrol regulated apoptotic
proteins in renal carcinoma cells in a dose-dependent manner.
**P<0.01, experimental vs. control groups. JS-K,
O2-(2,4-dinitrophenyl)1-[(4-ethoxyxarbonyl)
piperazin-1-yl]diazen-1-ium-1,2-diolate; FITC, fluorescein
isothiocyanate; Bcl-2, B-cell lymphoma 2; Bax, bcl-2-like protein
4; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. |
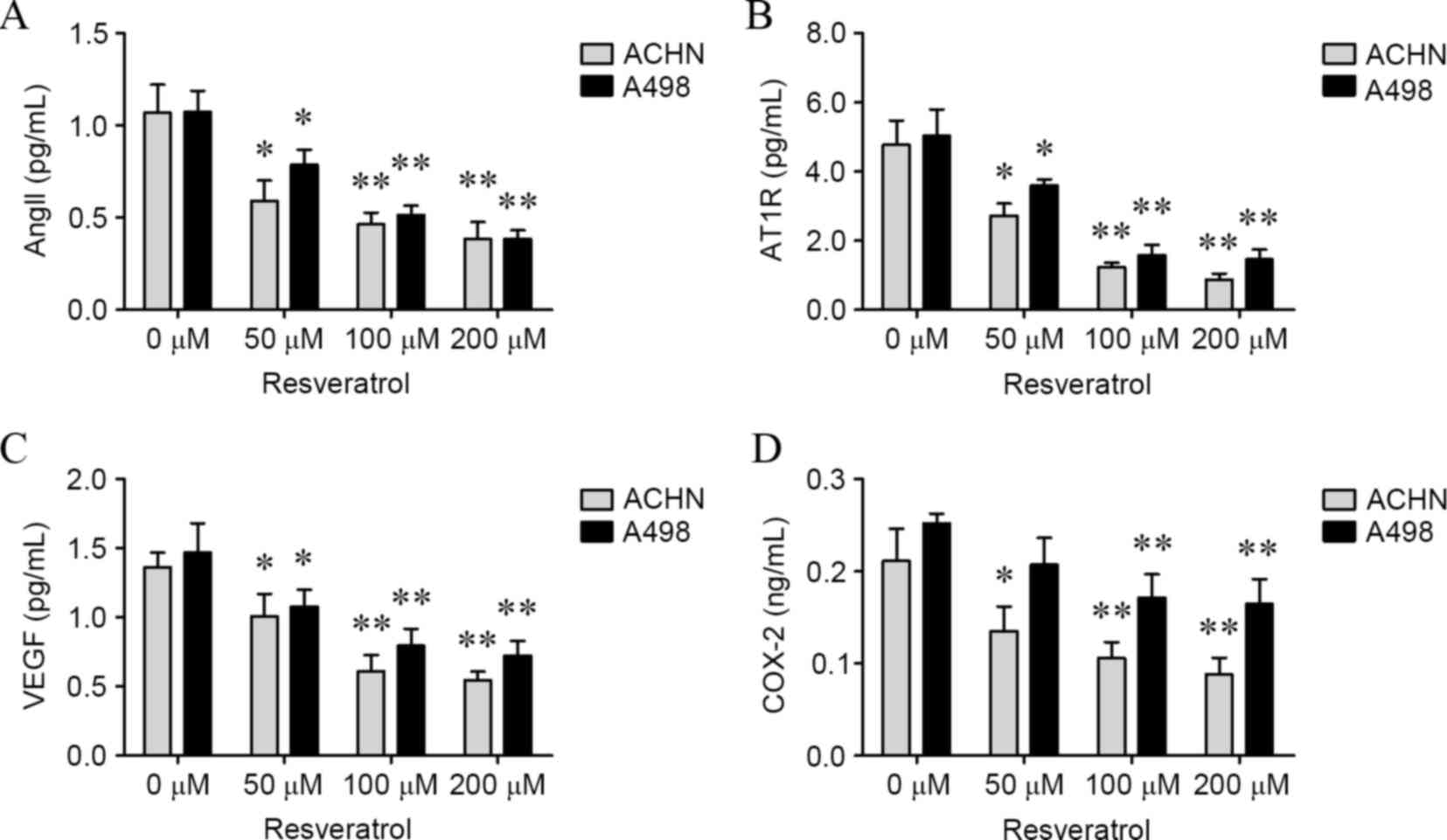
Resveratrol suppressed AngII, AT1R,
VEGF and COX-2 in a dose-dependent manner
To confirm whether AngII, AT1R, VEGF and COX-2
levels would be affected by resveratrol, ACHN and A498 cells were
treated with various concentrations of resveratrol for 24 h, and
the levels of AngII, AT1R, VEGF and COX-2 in cells and culture
medium were determined. As shown in Figs.
3 and 4, levels of AngII (ACHN
cells: 50 µM, P=0.0086; 100 µM, P<0.001; and 200 µM, P<0.001;
A498 cells: 50 µM, P=0.0015; 100 µM, P=0.0002; and 200 µM,
P=0.0003; culture medium of ACHN cells: 50 µM, P=0.0113; 100 µM,
P=0.0031; and 200 µM, P=0.0026; culture medium for A498 cells: 50
µM, P=0.0246; 100 µM, P=0.0015 and 200 µM, P=0.0007; Figs. 3A and 4A), AT1R (ACHN cells: 50 µM, P=0.0317; 100
µM, P=0.0033; and 200 µM, P=0.0013; A498 cells: 50 µM, P=0.0221;
100 µM, P=0.0031; and 200 µM, P=0.0023; culture medium for ACHN
cells: 50 µM, 0.0110; 100 µM, 0.0011; and 200 µM, 0.0008; culture
medium for A498 cells: 50 µM, P=0.0340; 100 µM, P=0.0019; and 200
µM, P=0.0016; Figs. 3B and 4B), VEGF (ACHN cells: 50 µM, P=0.0062; 100
µM, P=0.0068; and 200 µM, P=0.0024; A498 cells: 50 µM, P=0.0221;
100 µM, P=0.0259; and 200 µM, 0.0045; culture medium for ACHN
cells: 50 µM, P=0.0341; 100 µM, P=0.0012; and 200 µM, P=0.0003;
culture medium for A498 cells: 100 µM, 0.0090; and 200 µM, 0.0056;
Figs. 3C and 4C) and COX-2 (ACHN cells: 100 µM, 0.0246;
and 200 µM, 0.0138; A498 cells: 100 µM, 0.0398; and 200 µM, 0.0259;
culture medium for ACHN cells: 50 µM, 0.0406; 100 µM, 0.0097; and
200 µM, 0.0056; culture medium for A498 cells: 100 µM, 0.0065; and
200 µM, 0.0061; Figs. 3D and 4D) were significantly decreased in the two
cell lines and culture mediums in a dose-dependent manner compared
with cells treated with 0 µM resveratrol.
 | Figure 3.Resveratrol suppressed intracellular
AngII, AT1R, VEGF and COX-2 levels in a dose-dependent manner.
Subsequent to renal carcinoma cells being treated with resveratrol
(0, 50, 100 and 200 µM) for 24 h, the intracellular levels of (A)
AngII, (B) AT1R, (C) VEGF and (D) COX-2 were determined. *P<0.05
and **P<0.01, experimental vs. control groups. AngII,
angiotensin II; AT1R, AngII type 1 receptor; VEGF, vascular
endothelial growth factor; COX-2, cyclooxygenase-2. |
AT1R and VEGF may reverse
resveratrol-induced renal carcinoma cell growth suppression and
apoptosis
To determine whether AngII, AT1R, VEGF and COX-2 may
reverse resveratrol-induced renal carcinoma cell regression and
apoptosis, cells were incubated with AngII (2 pg/ml), AT1R (5
pg/ml), VEGF (2 pg/ml) or COX-2 (0.5 ng/ml) when exposed to
resveratrol (100 µM). AT1R and VEGF were shown to reverse cell
growth suppression by resveratrol (AT1R-treatment: RES vs.
RES+AT1P, P=0.0442 in ACHN cells and P=0.0235 in A498A cells;
VEGF-treatment: RES vs. RES+VEGF, P=0.0429 in ACHN cells and
P=0.0257 in A498A cells; Fig. 5A),
and cell apoptosis assay indicated a similar result with the cell
growth results (AT1R-treatment: RES vs. RES+AT1P, P=0.0312 in ACHN
cells and P=0.01245 in A498A cells; VEGF-treatment: RES vs.
RES+VEGF, P=0.0351 in ACHN cells and P=0.0153 in A498A cells;
Fig. 5B). By contrast, AngII and
COX-2 played undetected roles of resveratrol-induced cell growth
suppression and apoptosis.
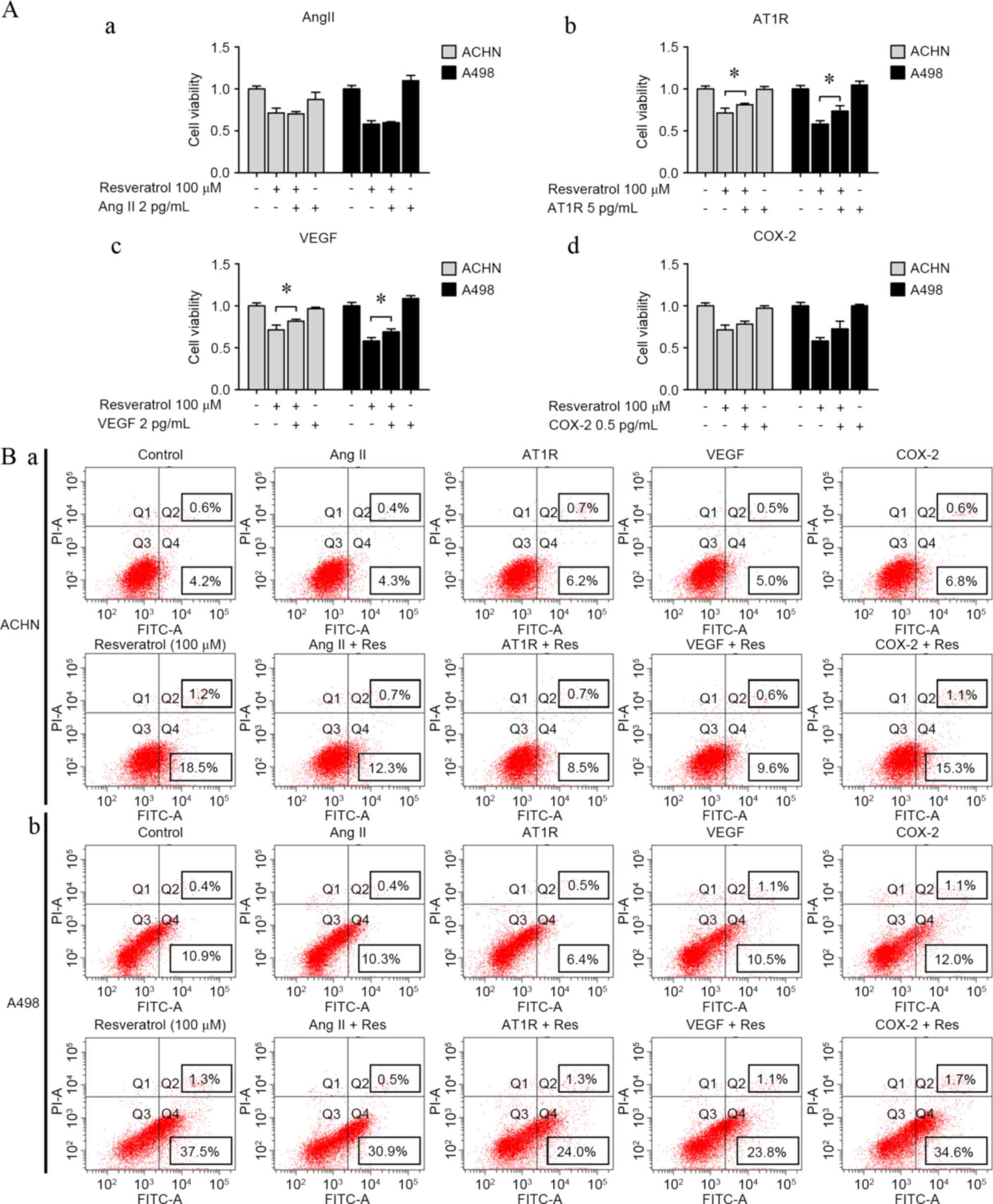 | Figure 5.AT1R and VEGF reversed
resveratrol-induced cell proliferation inhibition and apoptosis.
(A) Resveratrol-induced cell proliferation inhibition in renal
carcinoma cells with or without (a) AngII (2 pg/ml), (b) AT1R (5
pg/ml), (c) VEGF (2 pg/ml) and (d) COX-2 (0.5 ng/ml). (B) Apoptosis
of (a) ACHN and (b) A498 cells was analyzed according to flow
cytometry distributions with or without AngII (2 pg/ml), AT1R (5
pg/ml), VEGF (2 pg/ml) and COX-2 (0.5 ng/ml). *P<0.05,
experimental vs. control groups. AngII, angiotensin II; AT1R, AngII
type 1 receptor; VEGF, vascular endothelial growth factor; COX-2,
cyclooxygenase-2; FITC, fluorescein isothiocyanate. |
Discussion
As a type of free radical scavenger and antioxidant,
resveratrol is recognized as an effective anticancer compound in
several types of cancer, such as leukemia (28), breast (29–32),
prostate (33) and ovarian cancers
(34), melanoma (35) and primary brain tumors (36). In the present study, it was found that
resveratrol may suppress renal carcinoma cell growth in a time- and
dose-dependent manner. The present study also demonstrated that
resveratrol induced renal carcinoma cell apoptosis by regulating
apoptosis-associated proteins caspase 3/7/9, Bcl-2 and Bax. Reduced
AngII, AT1R, VEGF and COX-2 levels were detected subsequent to
resveratrol treatment and highlighted the importance of AT1R and
VEGF in resveratrol-induced cell apoptosis.
Previously, the renin-angiotensin system (RAS) has
been considered to be an endocrine system, and the function of
which is limited to regulate blood pressure and electrolyte
balance. However, recent studies have detected RAS in several
tissues and indicated that local RAS may be involved in regulating
a variety of physiological and pathological processes (7–9,37). In the RAS, AngII is recognized as a
key biological peptide (38), with 2
major specific receptors subtypes, consisting of AT1R and AT2R
(39). AngII acting on AT1R has a
central role in mediating the majority of the actions of the RAS
(16). Studies indicated that RAS was
involved in biological activities through the AngII/AT1R pathway
(39–41). In the present study, the levels of
AngII and AT1R were determined subsequent to resveratrol treatment.
The results demonstrated that AngII and AT1R were downregulated in
the cells and the culture medium. As VEGF and COX-2 were important
downstream regulators in RAS (42–45), it
was found that VEGF and COX-2 were inhibited in resveratrol-treated
cells and this result is similar to a previous study (29). The present data suggested that RAS
suppression would be an important event during resveratrol-induced
renal carcinoma cell apoptosis.
RAS was demonstrated to be involved in tumor
occurrence and development (46–48). For
example, AngII was able to promote epithelial-to-mesenchymal
transition in intrahepatic cholangiocarcinoma (38), and angiotensin-converting enzyme
inhibitor suppresses growth of colorectal cancer cells (49). AngII was demonstrated to induce VEGF
in pancreatic cancer cells through binding AT1R and extracellular
signal-regulated kinase 1/2 signaling; in addition, AngII/AT1R may
increase expression of VEGF (21).
AT1R was identified as upregulated in renal carcinoma and in
urogenital cancer in a previous study (40). COX-2 was detected as upregulated in
hypoxic conditions and induced angiogenesis through mechanisms
distinct from the VEGF axis (25–27). As a
type of anti-cancer drug, resveratrol was reported to act via a
RAS-dependent pathway (10). In the
present study, the effects of AngII, AT1R, VEGF and COX-2 on
resveratrol-induced cell growth suppression and apoptosis in renal
carcinoma were investigated. It was found that AT1R and VEGF
reverse resveratrol-induced renal carcinoma cell growth suppression
and apoptosis; however, AngII and COX-2 had no significant effect
on resveratrol-induced cell growth suppression and apoptosis. The
present results suggest that AT1R and VEGF are the critical factors
during resveratrol-induced renal carcinoma cell proliferation
suppression and apoptosis.
In summary, the present study examined the effect of
resveratrol on renal carcinoma cell growth, suppression and
apoptosis and investigated the potential mechanism through RAS. The
results indicated that resveratrol may inhibit cell growth, induce
apoptosis, and decrease AngII, AT1R, VEGF and COX-2 production in
renal carcinoma cells. However, AT1R and VEGF may reverse the
functions of resveratrol on renal carcinoma cells. Additional
studies should be conducted to demonstrate the exact mechanism
underlying the resveratrol-induced cell growth, suppression and
apoptosis, which may be, at least partially, through a
RAS-associated pathway in renal carcinoma cells.
Acknowledgements
This study was supported in part by the following
grants: Scientific Research Fund of Guangdong Medical College
(grant no. M2014019); Science and Technology Planning Project of
Guangdong Province (grant no. 2012B031800221); and The National
Natural Science Funds (grant no. 81272833) of China.
References
|
1
|
Aggarwal BB, Bhardwaj A, Aggarwal RS,
Seeram NP, Shishodia S and Takada Y: Role of resveratrol in
prevention and therapy of cancer: Preclinical and clinical studies.
Anticancer Res. 24:2783–2840. 2004.PubMed/NCBI
|
|
2
|
Carter LG, D'Orazio JA and Pearson KJ:
Resveratrol and cancer: Focus on in vivo evidence. Endocr-Relat
Cancer. 21:R209–R225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ndiaye M, Kumar R and Ahmad N: Resveratrol
in cancer management: Where are we and where we go from here? Ann N
Y Acad Sci. 1215:144–149. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Reagan-Shaw S, Mukhtar H and Ahmad N:
Resveratrol imparts photoprotection of normal cells and enhances
the efficacy of radiation therapy in cancer cells. Photochem
Photobiol. 84:415–421. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu ML, Li H, Yu LJ, Chen XY, Kong QY, Song
X, Shu XH and Liu J: Short-term resveratrol exposure causes in
vitro and in vivo growth inhibition and apoptosis of bladder cancer
cells. PLoS One. 9:e898062014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nishimura H: Renin-angiotensin system in
vertebrates: Phylogenetic view of structure and function. Anat Sci
Int. Oct 7–2016.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bradshaw AR, Wickremesekera AC, Brasch HD,
Chibnall AM, Davis PF, Tan ST and Itinteang T: Glioblastoma
multiforme cancer stem cells express components of the
renin-angiotensin system. Front Surg. 3:512016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yoshida T, Kinoshita H, Fukui K, Matsuzaki
T, Yoshida K, Mishima T, Yanishi M, Komai Y, Sugi M, Inoue T, et
al: Prognostic impact of renin-angiotensin inhibitors in patients
with bladder cancer undergoing radical cystectomy. Ann Surg Oncol.
Oct 11–2016.(Epub ahead of print). View Article : Google Scholar
|
|
9
|
Miyajima A, Yazawa S, Kosaka T, Tanaka N,
Shirotake S, Mizuno R, Kikuchi E and Oya M: Prognostic impact of
renin-angiotensin system blockade on renal cell carcinoma after
surgery. Ann Surg Oncol. 22:3751–3759. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang R, Zhang H and Zhu L: Inhibitory
effect of resveratrol on the expression of the VEGF gene and
proliferation in renal cancer cells. Mol Med Rep. 4:981–983.
2011.PubMed/NCBI
|
|
11
|
Shirotake S, Miyajima A, Kosaka T, Tanaka
N, Kikuchi E, Mikami S, Okada Y and Oya M: Regulation of monocyte
chemoattractant protein-1 through angiotensin II type 1 receptor in
prostate cancer. Am J Pathol. 180:1008–1016. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Crowley SD and Coffman TM: Recent advances
involving the renin-angiotensin system. Exp Cell Res.
318:1049–1056. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arrieta O, PinedaOlvera B, GuevaraSalazar
P, Hernández-Pedro N, Morales-Espinosa D, Cerón-Lizarraga TL,
González-De la Rosa CH, Rembao D, Segura-Pacheco B and Sotelo J:
Expression of AT1 and AT2 angiotensin receptors in astrocytomas is
associated with poor prognosis. Br J Cancer. 99:160–166. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li SH, Lu HI, Chang AY, Huang WT, Lin WC,
Lee CC, Tien WY, Lan YC, Tsai HT and Chen CH: Angiotensin II type I
receptor (AT1R) is an independent prognosticator of esophageal
squamous cell carcinoma and promotes cells proliferation via mTOR
activation. Oncotarget. Aug 24–2016.(Epub ahead of print).
|
|
15
|
Dolley-Hitze T, Jouan F, Martin B, Mottier
S, Edeline J, Moranne O, Le Pogamp P, Belaud-Rotureau MA, Patard
JJ, Rioux-Leclercq N and Vigneau C: Angiotensin-2 receptors (AT1-R
and AT2-R), new prognostic factors for renal clear-cell carcinoma?
Br J Cancer. 103:1698–1705. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Passos-Silva DG, VeranoBraga T and Santos
RA: Angiotensin-(1–7): Beyond the cardio-renal actions. Clin Sci
(Lond). 124:443–456. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ferrara N and Kerbel RS: Angiogenesis as a
therapeutic target. Nature. 438:967–974. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng YD, Yang H, Chen GQ and Zhang ZC:
Molecularly targeted drugs for metastatic colorectal cancer. Drug
Des Devel Ther. 7:1315–1322. 2013.PubMed/NCBI
|
|
20
|
Ding C, Li L, Yang T, Fan X and Wu G:
Combined application of anti-VEGF and anti-EGFR attenuates the
growth and angiogenesis of colorectal cancer mainly through
suppressing AKT and ERK signaling in mice model. BMC Cancer.
16:7912016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Anandanadesan R, Gong Q, Chipitsyna G,
Witkiewicz A, Yeo CJ and Arafat HA: Angiotensin II induces vascular
endothelial growth factor in pancreatic cancer cells through an
angiotensin II type 1 receptor and ERK1/2 signaling. J Gastrointest
Surg. 12:57–66. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hoellen F, Waldmann A, BanzJansen C, Rody
A, Heide M, Köster F, Ribbat-Idel J, Thorns C, Gebhard M,
Oberländer M, et al: Expression of cyclooxygenase-2 in cervical
cancer is associated with lymphovascular invasion. Oncol Lett.
12:2351–2356. 2016.PubMed/NCBI
|
|
23
|
Ali-Fehmi R, Morris RT, Bandyopadhyay S,
Che M, Schimp V, Malone JM Jr and Munkarah AR: Expression of
cyclooxygenase-2 in advanced stage ovarian serous carcinoma:
Correlation with tumor cell proliferation, apoptosis, angiogenesis,
and survival. Am J Obstet Gynecol. 192:819–825. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Subbaramaiah K and Dannenberg AJ:
Cyclooxygenase 2: A molecular target for cancer prevention and
treatment. Trends Pharmacol Sci. 24:96–102. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schmedtje JF Jr, Ji YS, Liu WL, DuBois RN
and Runge MS: Hypoxia induces cyclooxygenase-2 via the NF-kappaB
p65 transcription factor in human vascular endothelial cells. J
Biol Chem. 272:601–608. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee JJ, Natsuizaka M, Ohashi S, Wong GS,
Takaoka M, Michaylira CZ, Budo D, Tobias JW, Kanai M, Shirakawa Y,
et al: Hypoxia activates the cyclooxygenase-2-prostaglandin E
synthase axis. Carcinogenesis. 31:427–434. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rüegg C, Dormond O and Mariotti A:
Endothelial cell integrins and COX-2: Mediators and therapeutic
targets of tumor angiogenesis. Biochim Biophys Acta. 1654:51–67.
2004.PubMed/NCBI
|
|
28
|
Yaseen A, Chen S, Hock S, Rosato R, Dent
P, Dai Y and Grant S: Resveratrol sensitizes acute myelogenous
leukemia cells to histone deacetylase inhibitors through reactive
oxygen species-mediated activation of the extrinsic apoptotic
pathway. Mol Pharmacol. 82:1030–1041. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Singh N, Nigam M, Ranjan V, Zaidi D, Garg
VK, Sharma S, Chaturvedi R, Shankar R, Kumar S, Sharma R, et al:
Resveratrol as an adjunct therapy in cyclophosphamide-treated MCF-7
cells and breast tumor explants. Cancer Sci. 102:1059–1067. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shi Y, Yang S, Troup S, Lu X, Callaghan S,
Park DS, Xing Y and Yang X: Resveratrol induces apoptosis in breast
cancer cells by E2F1-mediated up-regulation of ASPP1. Oncol Rep.
25:1713–1719. 2011.PubMed/NCBI
|
|
31
|
Chottanapund S, Van Duursen MB, Navasumrit
P, Hunsonti P, Timtavorn S, Ruchirawat M and Van den Berg M:
Anti-aromatase effect of resveratrol and melatonin on hormonal
positive breast cancer cells co-cultured with breast adipose
fibroblasts. Toxicol In Vitro. 28:1215–1221. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Leon-Galicia I, DiazChavez J, GarciaVilla
E, UribeFigueroa L, HidalgoMiranda A, Herrera LA, AlvarezRios E,
GarciaMena J and Gariglio P: Resveratrol induces downregulation of
DNA repair genes in MCF-7 human breast cancer cells. Eur J Cancer
Prev. 22:11–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fraser SP, Peters A, FlemingJones S,
Mukhey D and Djamgoz MB: Resveratrol: Inhibitory effects on
metastatic cell behaviors and voltage-gated Na+ channel
activity in rat prostate cancer in vitro. Nutr Cancer.
66:1047–1058. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mikula-Pietrasik J, Sosińska P and Książek
K: Resveratrol inhibits ovarian cancer cell adhesion to peritoneal
mesothelium in vitro by modulating the production of α5β1 integrins
and hyaluronic acid. Gynecol Oncol. 134:624–630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen YJ, Chen YY, Lin YF, Hu HY and Liao
HF: Resveratrol inhibits alpha-melanocyte-stimulating hormone
signaling, viability, and invasiveness in melanoma cells. Evid
Based Complement Alternat Med. 2013:6321212013.PubMed/NCBI
|
|
36
|
Wen S, Li H, Wu ML, Fan SH, Wang Q, Shu
XH, Kong QY, Chen XY and Liu J: Inhibition of NF-κB signaling
commits resveratrol-treated medulloblastoma cells to apoptosis
without neuronal differentiation. J Neurooncol. 104:169–177. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Magliano DC, Penna-de-Carvalho A,
Vazquez-Carrera M, Mandarim-de-Lacerda CA and Aguila MB: Short-term
administration of GW501516 improves inflammatory state in white
adipose tissue and liver damage in high-fructose-fed mice through
modulation of the renin-angiotensin system. Endocrine. 50:355–367.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Okamoto K, Tajima H, Nakanuma S, Sakai S,
Makino I, Kinoshita J, Hayashi H, Nakamura K, Oyama K, Nakagawara
H, et al: Angiotensin II enhances epithelial-to-mesenchymal
transition through the interaction between activated hepatic
stellate cells and the stromal cell-derived factor-1/CXCR4 axis in
intrahepatic cholangiocarcinoma. Int J Oncol. 41:573–582.
2012.PubMed/NCBI
|
|
39
|
Kosaka T, Miyajima A, Takayama E, Kikuchi
E, Nakashima J, Ohigashi T, Asano T, Sakamoto M, Okita H, Murai M
and Hayakawa M: Angiotensin II type 1 receptor antagonist as an
angiogenic inhibitor in prostate cancer. Prostate. 67:41–49. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Miyajima A, Kikuchi E, Kosaka T and Oya M:
Angiotensin II type 1 receptor antagonist as an angiogenic
inhibitor in urogenital cancer. Rev Recent Clin Trials. 4:75–78.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Miyajima A, Kosaka T, Asano T, Seta K,
Kawai T and Hayakawa M: Angiotensin II type I antagonist prevents
pulmonary metastasis of murine renal cancer by inhibiting tumor
angiogenesis. Cancer Res. 62:4176–4179. 2002.PubMed/NCBI
|
|
42
|
de Haas S, Delmar P, Bansal AT, Moisse M,
Miles DW, Leighl N, Escudier B, Van Cutsem E, Carmeliet P, Scherer
SJ, et al: Genetic variability of VEGF pathway genes in six
randomized phase III trials assessing the addition of bevacizumab
to standard therapy. Angiogenesis. 17:909–920. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mittal K, Koon H, Elson P, Triozzi P,
Dowlati A, Chen H, Borden EC and Rini BI: Dual VEGF/VEGFR
inhibition in advanced solid malignancies: Clinical effects and
pharmacodynamic biomarkers. Cancer Biol Ther. 15:975–981. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chang WS, Liao CH, Miao CE, Wu HC, Hou LL,
Hsiao CL, Ji HX, Tsai CW and Bau DT: The role of functional
polymorphisms of cyclooxygenase 2 in renal cell carcinoma.
Anticancer Res. 34:5481–5486. 2014.PubMed/NCBI
|
|
45
|
He W, Zhang M, Zhao M, Davis LS, Blackwell
TS, Yull F, Breyer MD and Hao CM: Increased dietary sodium induces
COX2 expression by activating NFkB in renal medullary interstitial
cells. Pflugers Arch. 466:357–367. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fan L, Feng Y, Wan HY, Ni L, Qian YR, Guo
Y, Xiang Y and Li QY: Hypoxia induces dysregulation of local
renin-angiotensin system in mouse Lewis lung carcinoma cells. Genet
Mol Res. 13:10562–10573. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ino K, Shibata K, Kajiyama H, Nawa A,
Nomura S and Kikkawa F: Manipulating the angiotensin system-new
approaches to the treatment of solid tumours. Expert Opin Biol
Ther. 6:243–255. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Araújo WF, Naves MA, Ravanini JN, Schor N
and Teixeira VP: Renin-angiotensin system (RAS) blockade attenuates
growth and metastatic potential of renal cell carcinoma in mice.
Urol Oncol. 33:389.e1–e7. 2015. View Article : Google Scholar
|
|
49
|
Childers WK: Interactions of the
renin-angiotensin system in colorectal cancer and metastasis. Int J
Colorectal Dis. 30:749–752. 2015. View Article : Google Scholar : PubMed/NCBI
|