Introduction
Hepatocellular carcinoma (HCC) is the third most
common cancer worldwide (1). Each
year, >620,000 patients are diagnosed with HCC, and the 1-year
survival rate remains <50% (2).
Over the past few decades, HCC age-adjusted incidence rates have
doubled (3), and primary liver cancer
mortality rates have increased faster than the mortality rates of
most other cancers (4).
Current therapeutic modalities for HCC include
curative options, such as surgical resection, liver
transplantation, and local ablation; or palliative procedures, such
as catheter-directed therapies and systemic therapy (5). The main risk factors for HCC are viral
hepatitis, alcohol abuse, aflatoxin B1-contaminated food,
nonalcoholic fatty liver disease, and metabolic disorders (6). However, not all individuals exposed to
these factors possess the same risk of developing HCC. It is a
multifactorial disease, with a wide range of genetic and epigenetic
alterations identified as contributory to the deregulation of key
oncogenes and tumor-suppressor genes; however, genetic events in
hepatic carcinogenesis are poorly understood (7). Previous studies undertaken by our group
uncovered potential molecular targets for HCC treatment, by
comparing the gene expression patterns of HCC with those of normal
liver tissues using a cDNA expression microarray containing 1361
unique genes (8). Among the genes
explored, NIMA-related kinase 2 (Nek2), also known as
Serine/threonine-protein kinase 2, was identified as a potential
target gene as it is highly expressed in HCC.
Nek2 is a member of the serine/threonine kinase
family, and is important in regulating the cell cycle, gene
expression, and maintaining centrosomal structure and function
(9–11). Nek2 belongs to the never in mitosis A
(NIMA)-related family of kinases, otherwise known as Neks (9). The Nek family comprises 11 members, all
of which have been identified in mammals (9). Given the role of Nek2 in regulating
centrosomal function during the S- and G2-phases, Nek2 regulation
is strictly controlled throughout the cell cycle, with a hinged
expression in such phases (12).
Functional research has implicated Nek2 in the regulation of
centrosome separation and spindle formation (9,13).
Furthermore, high Nek2 expression levels have been reported in cell
lines derived from breast, cervical, prostate, and colorectal
cancers, as well as those from cholangiocarcinoma and lymphoma
(14–17). A recent study involving a cohort of
normal liver tissues, chronic liver disease induced by the
hepatitis C virus, and HCC, identified Nek2 co-expression with
several closely related genes (18).
The present study aims to investigate Nek2
expression and biological regulation in human HCC, analyze the
association of Nek2 expression with the clinicopathological
characteristics of HCC patients, and examine the Nek2-mediated
control of cell growth and proliferation to explore the potential
molecular mechanisms involved in the progress of HCC.
Materials and methods
Patients and tissue samples
Patients were recruited from the First Municipal
People's Hospital of Guangzhou, and clinical information was
acquired from hospital registries. Primary tumor specimens were
obtained from 52 patients diagnosed with HCC, all of whom provided
informed consent for participation in the study. All 52 patients
underwent routine resection in the First Municipal People's
Hospital of Guangzhou between March 2010 and November 2012. HCC was
diagnosed according to the World Health Organization histological
classification of tumors of the liver and intrahepatic bile ducts
(2000). All surgical specimens were immediately snap-frozen in
liquid nitrogen and stored at −80°C until RNA and protein
extraction. The samples were obtained from tumor tissues and
corresponding adjacent healthy tissues 5–10 cm from each tumor
<30 min following surgery. Clinical histopathological data
including histopathological diagnosis, Edmondson-Steiner stage, and
tumor grade were obtained from patient medical records. The present
study was approved by the Ethics Committee of First Municipal
People's Hospital of Guangzhou, Guangdong, China.
Cell culture and transfection
The human hepatocellular carcinoma cell lines of
HepG2, Hep3B, BEL-7402, SMMC-7721, QCY-7701, PLC/PRF/5, and the
human hepatocyte cell line HL-7702, were obtained from the cell
repository of Sun Yat-sen University (Guangzhou, Guangdong, China).
All cells were cultured at 37°C in a humidified atmosphere with 5%
CO2 in Dulbecco's modified eagle medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) containing 10% fetal
calf serum (Invitrogen; Thermo Fisher Scientific Inc.) and 100
IU/ml penicillin-streptomycin solution (Corning Inc., Corning, NY,
USA). The cells were grown on sterilized culture dishes (Corning
Inc.), harvested, and passaged with 0.25% trypsin-EDTA (Corning,
Inc.) upon reaching 80% confluence. X-treme GENE siRNA Transfection
Reagent (Roche Diagnostics GmbH, Mannheim, Germany) was used for
siRNA transfection according to the manufacturer's instructions.
Protein levels were assessed 72 h later by western blot analysis,
and mRNA levels were quantified 48 h later by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR). The
anti-Nek2 siRNA (Sc43960; Santa Cruz Biotechnology Inc., Dallas,
TX, USA) and control siRNA (Sc37007; Santa Cruz Biotechnology,
Inc.) were used to transfect HepG2 cells.
Immunohistochemistry (IHC)
IHC experiments were conducted on 5 µm
formalin-fixed, paraffin-embedded sections of healthy tissue and
tumor samples. The tissue sections were mounted on slides, treated
with xylene, and rehydrated using alcohol-water mixtures before
heat treatment in 10 mM citric acid (pH 6.0) for antigen retrieval.
Endogenous peroxidase activity was stopped by the use of 0.3%
H2O2 solution. After blocking with 10% bovine
serum albumin (Beyotime Institute of Biotechnology, Haimen, China;
ST023) the sections were incubated at 4°C overnight with primary
human Nek2 antibodies (Abcam, Cambridge, UK; Ab55550; 1:400
dilution), rinsed, and incubated with biotinylated anti-mouse IgG
(Dako North America Inc., Carpinteria, CA, USA; F0232; 1:100
dilution) for 30 min at room temperature. After rinsing, the
samples were treated with streptavidin and biotinylated horseradish
peroxidase (Dako North America Inc.). Counterstaining with
hematoxylin was performed, and the sections were dehydrated in
ethanol before mounting. The sections were analyzed using optical
microscopy (Nikon Corp., Tokyo, Japan; magnification, ×100 or
×400). Staining specificity was confirmed using positive (gastric
mucosa) and negative (heart muscle tissue) controls. Low Nek2
expression was defined as <10% and high expression as
>10%.
RT-qPCR
Total RNA was extracted from HepG2 cells or tissues
using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol. Complementary DNA was synthesized
from 1.5 µg of total RNA at 42°C for 60 min using the SYBR-Green
PCR master mix (Thermo Fisher Scientific, Inc.; 4368577) and
RT-qPCR was performed using the Brilliant II Sybr Green QPCR Master
Mix (Agilient Technologies, Inc., Santa Clara, CA, USA). GAPDH was
used as the reference gene. The primer sequences were as follows:
Forward, 5′-CCAGCCCTGTATTGAGTG-3′ and reverse,
5′-ACTTCCGTTCCTTTAGCA-3′ for Nek2; forward
5′-GACCTGACCTGCCGTCTA-3′, and reverse, 5′-AGGAGTGGGTGTCGCTGT-3′ for
GAPDH. The relative levels of gene expression were represented as
ΔCq, and the fold-change of gene expression was calculated using
the 2−ΔΔCq method.
Western blotting
The cells and tissue samples were lysed with 20
mmol/l Tris-HCl (pH 8.0), 137 mmol/l NaCl, 10% glycerol, 1% Triton
X-100, and 2 mmol/l EDTA containing protease and phosphatase
inhibitors at 4°C. Each lysate sample (20 µg) was separated by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and transferred to nitrocellulose membranes. The
membranes were incubated overnight at 4°C with antibodies against
Nek2 (Santa Cruz Biotechnology, Inc.; Sc55601; 1:1,000 dilution),
β-catenin, C-Myc, CycD1, CycB1, CDK1, CycE, and P27 (Santa Cruz
Biotechnology, Inc., 1:1,000 dilution). The membranes were
developed using enhanced chemiluminescence. The Nek2 expression
levels were normalized to that of GAPDH, and were detected using
mouse anti-GAPDH antibody (Santa Cruz Biotechnology, Inc.;
Sc365062; 1:2,000 dilution).
Cell growth and proliferation
The cultured HepG2 cells were pelleted by
centrifugation at 900 × g and re-suspended in phenol red-free
medium supplemented with 10% charcoal-stripped fetal bovine serum
(HyClone; GE Healthcare Life Sciences, Logan, UT, USA) and 2%
L-glutamine. The HepG2 cells were then plated in triplicate,
(1×103 cells/ml) in 96-well micro plates. This process
was repeated six times. After seeding for 24 h, the HepG2 cells
were transfected with either anti-Nek2 or control siRNAs (10 nM
each) using the X-treme GENE siRNA Transfection Reagent (Roche
Diagnostics GmbH) according to the manufacturer's protocol. The
cells were counted daily over a 5-day period through the Cell Titer
96 Non-Radioactive cell proliferation assay (Promega Corp.,
Madison, WI, USA). Following this, 20 µl of 5 mg/ml MTS solution
was added to each well, and the resulting mixture was incubated at
37°C for 4 h. Each solution was measured by a Bio-Rad micro plate
reader (Bio-Rad Laboratories Inc.) at a test wavelength of 490
nm.
Cell colony formation
In the anchorage-independent colony growth assay,
the HepG2 cells were transfected with siRNA for 48 h. A 12-well
cell culture plate containing 0.5% agarose medium on top of regular
medium containing 1% low-melting agar was seeded with ~500
transfected cells. After 15 days, the resulting colonies were
stained with Giemsa and counted under a microscope.
Cell cycle and apoptosis analysis
HepG2 cells were seeded in 8 cm sterilized culture
dishes for 12 h. The cells were then transfected with indicated
amounts of anti-Nek2 or control siRNAs. At indicated time points,
the cells were harvested, fixed with 1% paraformaldehyde, washed
with phosphate buffer solution (PBS), and stained with 5 mg/ml
propidium iodide (PI) in PBS supplemented with RNase A
(Sigma-Aldrich, St. Loius, MO, USA) for 30 min at room temperature.
Data were collected using a BD FACSCAlibur flow cytometer (BD
Biosciences, San Jose, CA, USA). Apoptosis was measured using an
Annexin V/FITC kit (Sigma-Aldrich) according to the manufacturer's
protocol. Each assay was repeated in triplicate.
Statistical analyses
All statistical calculations were performed using
SPSS 13.0 software (SPSS Inc., Chicago, IL, USA). A chi-square test
was used to examine possible correlations between Nek2 expression
and clinicopathological factors. The differences in the
densitometry data on focus numbers between the control and
Nek2-transfected cells were analyzed using Student's t-test.
P<0.05 was considered to indicate a statistically significant
result in both Student's t-test and the chi-square test.
Results
Nek2 expression in human HCC tissue
and cell lines
Nek2 expression in resected human tissue specimens
from healthy and cancerous livers was analyzed. Fifty-two HCC and
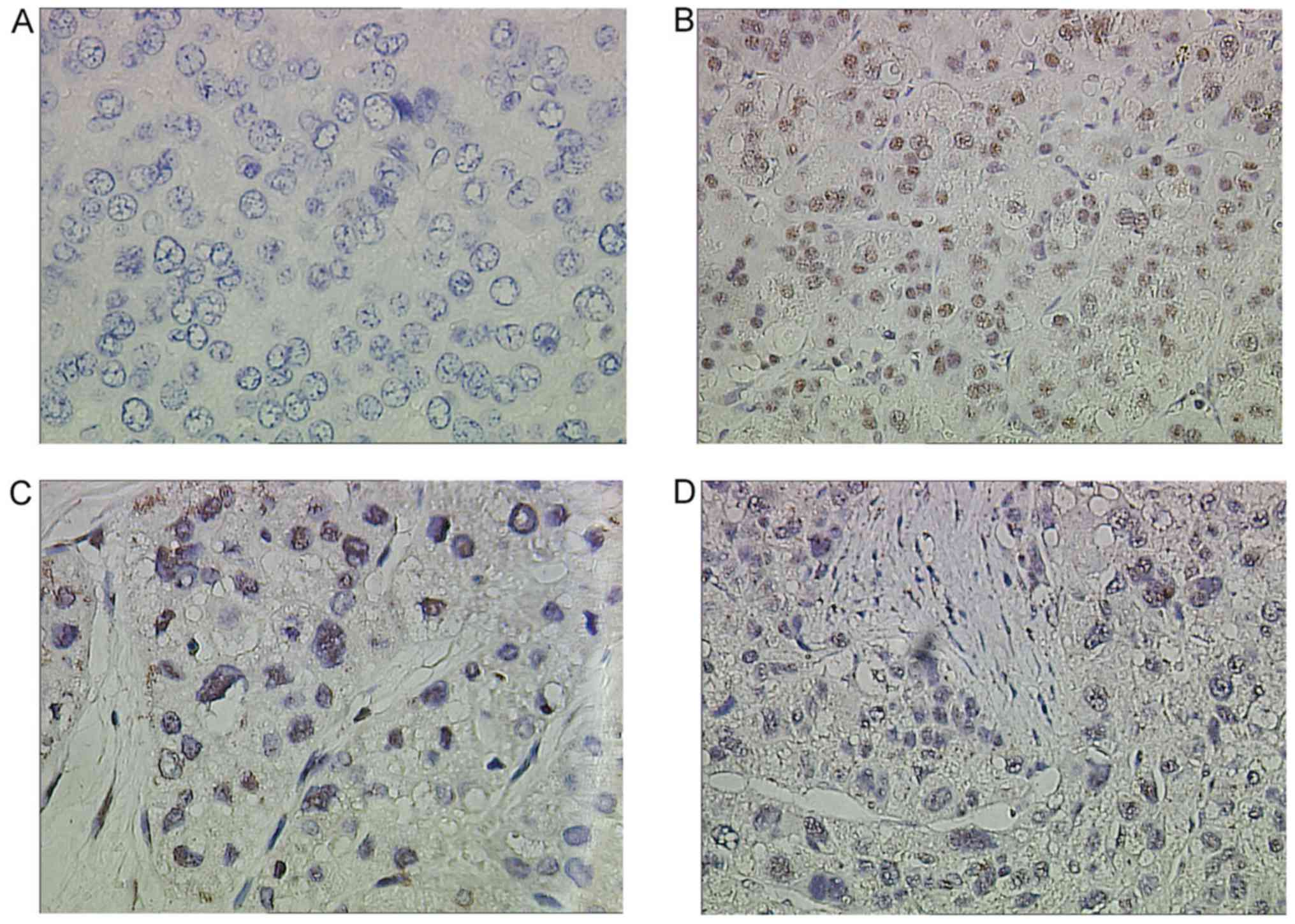
normal samples were examined using Nek2-specific IHC staining
antibodies (Fig. 1). The IHC results
revealed clear differences in Nek2 expression between healthy and
cancerous liver tissues. In the healthy and adjacent liver tissues,
Nek2 was not expressed (Fig. 1A). By
contrast, most HCC cases presented elevated protein levels in the
nuclei (Fig. 1B-D). Expression of
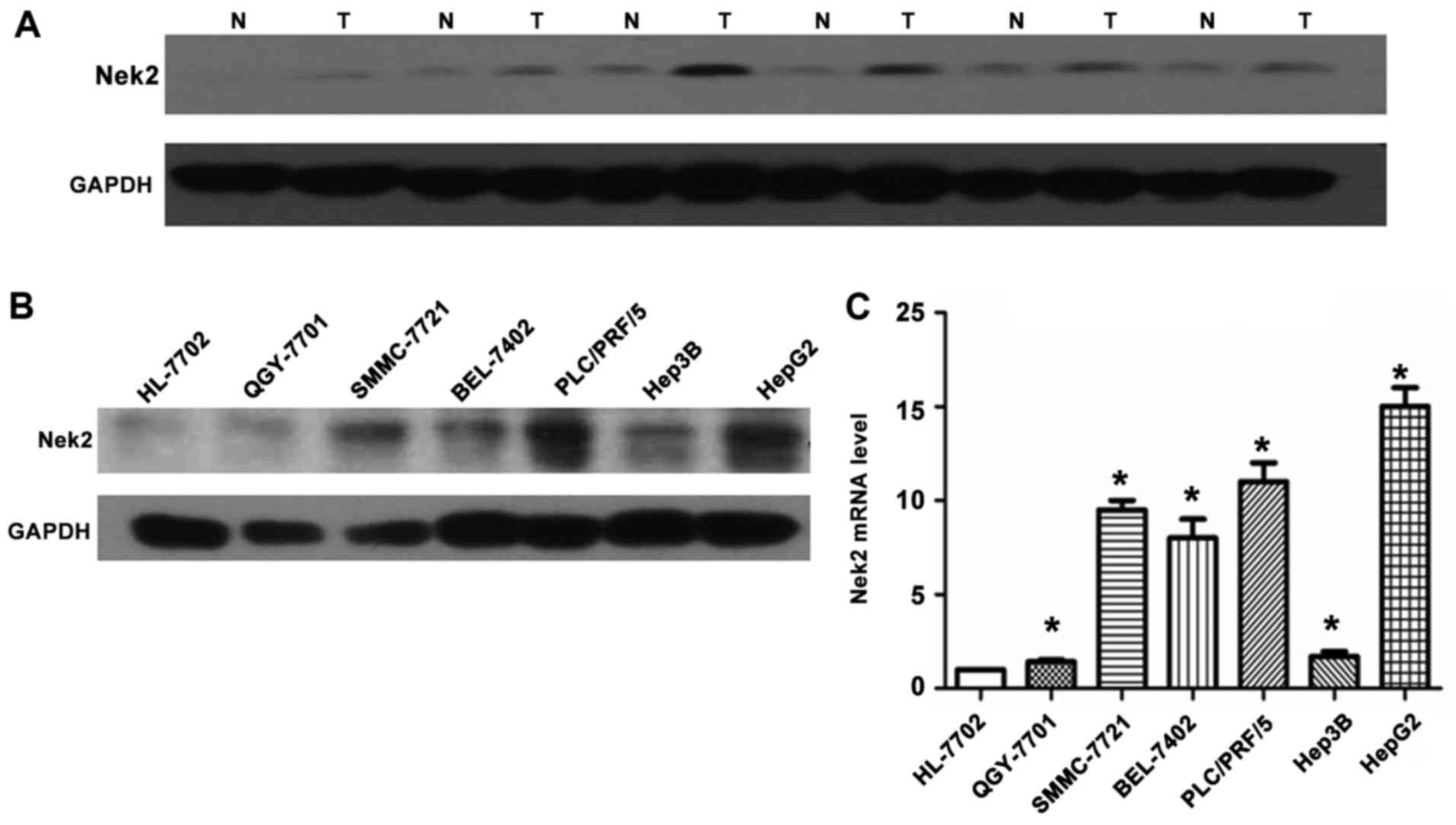
Nek2 was examined in 17 pairs of snap-frozen HCC and adjacent
nonmalignant liver tissues by western blotting. Levels of Nek2
protein were significantly elevated in the HCC specimens (T)
compared with those in adjacent nonmalignant tissues (N; Fig. 2A). Meanwhile, analysis of Nek2
transcription by RT-qPCR and western blot analysis demonstrated
that HepG2 cells expressed significantly higher levels of Nek2 mRNA
and protein compared to the other six cell lines (HL-7702,
PLC/PRF/5, Hep3B, BEL-7402, SMMC-7721, and QCY-7701; Fig. 2B and C).
Associations between Nek2 expression
and the clinicopathological parameters of HCC patients
Thirty (57.6%) out of 52 tumor tissues expressed
Nek2 (Table I), and high levels of
Nek2 were identified predominantly in the nuclei of cancerous liver
cells. The correlation between Nek2 protein expression and the
histological staging of liver cancer was analysed (Table I). Rates of Nek2 expression in tumors
classed as Edmondson-Steiner stages I and II (Fig. 1B and C) were 52.38% (22/42, Table I) and those with Edmondson-Steiner
stages III and IV (Fig. 1D) were 80%
(8/10; Table I). The difference in
Nek2 expression between samples from the two tumor classes was
statistically significant (P<0.05). Among all the samples, those
with concomitant portal venous tumor emboli exhibiting Nek2
expression comprised 90.0% (9/10), however, only 50.0% (21/42)
without portal venous tumor emboli expressed Nek2 (Table I). A significant difference in Nek2
expression was noted in tumor malignancy (P<0.05). Nek2
expression was observed to be independent of episode age, tumor
size, Hepatic Function and liver α-fetoprotein levels (Table I).
 | Table I.Association of Nek2 expression with
clinicopathological characteristics of patients with HCC. |
Table I.
Association of Nek2 expression with
clinicopathological characteristics of patients with HCC.
| Characteristics | Patients, no. | Nek2 positive, no.
(%) | χ2 | P-value |
|---|
| Age, years |
|
|
|
|
|
<60 | 17 | 11 (64.7) | 0.632 | 0.426 |
| ≥60 | 35 | 19 (54.3) |
|
|
| Tumor size, cm |
|
|
|
|
|
<5 | 28 | 17 (60.9) | 1.213 | 0.271 |
| ≥5 | 24 | 13 (54.2) |
|
|
| Hepatic
function |
|
|
|
|
| Child
A | 15 | 9 (60) | 0.001 | 0.971 |
| Child
B | 37 | 21 (56.7) |
|
|
| Edmondson
stage |
|
|
|
|
| Stage I
and II | 42 | 22 (52.4) | 4.434 | 0.037a |
| Stage
III and IV | 10 | 8 (80) |
|
|
| Portal vein tumor
thrombus |
|
|
|
|
|
Positive | 10 | 9
(90) | 5.295 | 0.032a |
|
Negative | 42 | 21 (50) |
|
|
| AFP |
|
Positive | 44 | 24 (54.5) | 1.160 | 0.281 |
|
Negative | 8 | 6
(75.0) |
|
|
Inhibition of Nek2 suppresses HepG2
proliferation
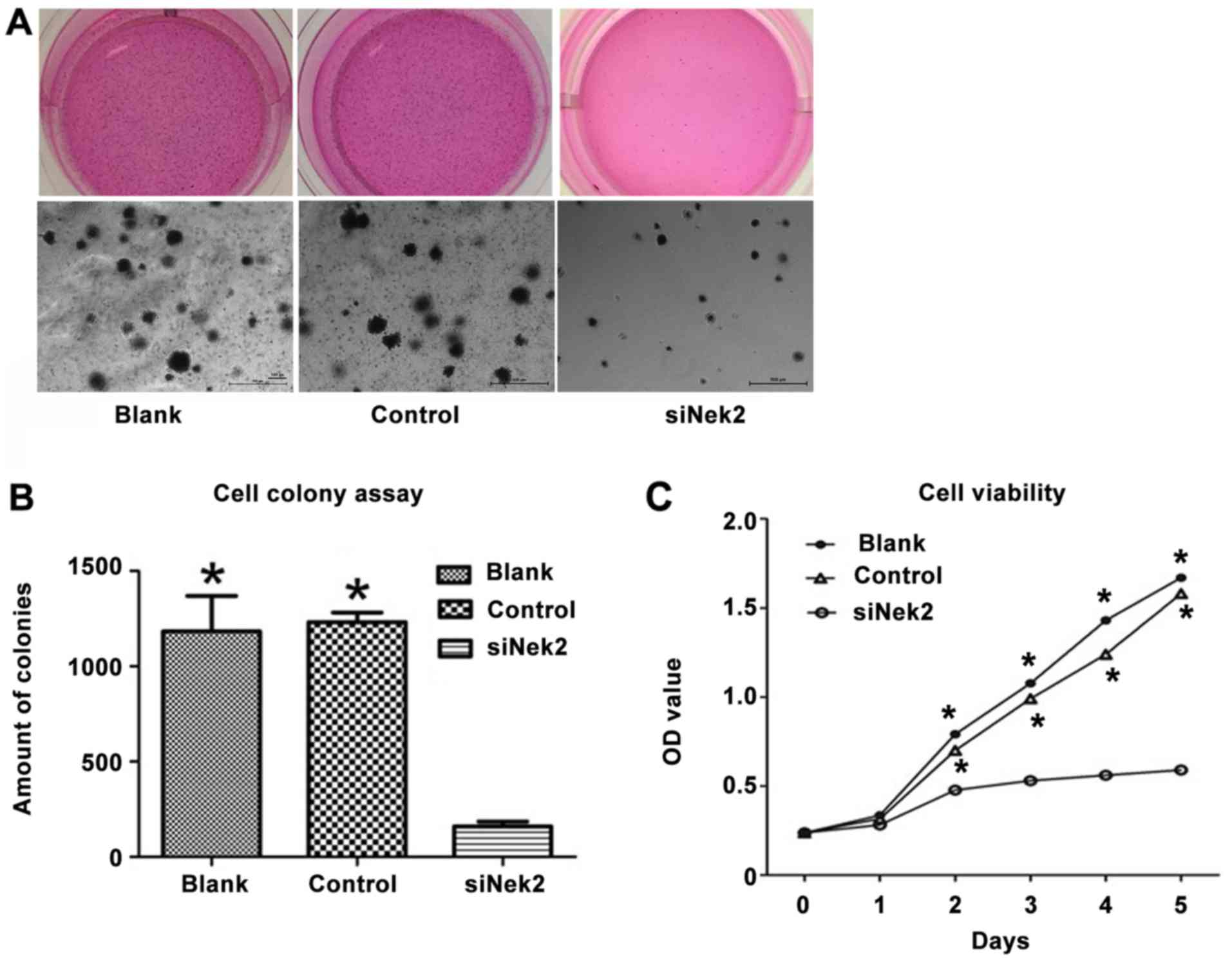
Using siRNA as an efficient tool for Nek2 gene
silencing in the HepG2 cell line, the role of Nek2 in HCC cell
proliferation was analyzed. The proliferation rate was determined
by MTS assay, assessing cell metabolic activity. A decrease in
proliferation was observed in the HepG2 cells treated with
anti-Nek2 siRNA compared with the cells transfected with the
control siRNA and blank group (Fig.
3A). Consistent with the MTS result, the soft agar colony
formation assays showed that Nek2 knockdown in HepG2 cells led to a
significant decrease in focus number [1183±97 (Blank), 1232±28
(Control) vs. 160±14 (siNek2), P<0.05; Fig. 3B]. The cell viability of HepG2 cells
treated with Nek siRNA decreased significantly compared with the
control and blank groups (P<0.05; Fig.
3C).
Effect of Nek2 on cell cycle and
apoptosis of HepG2
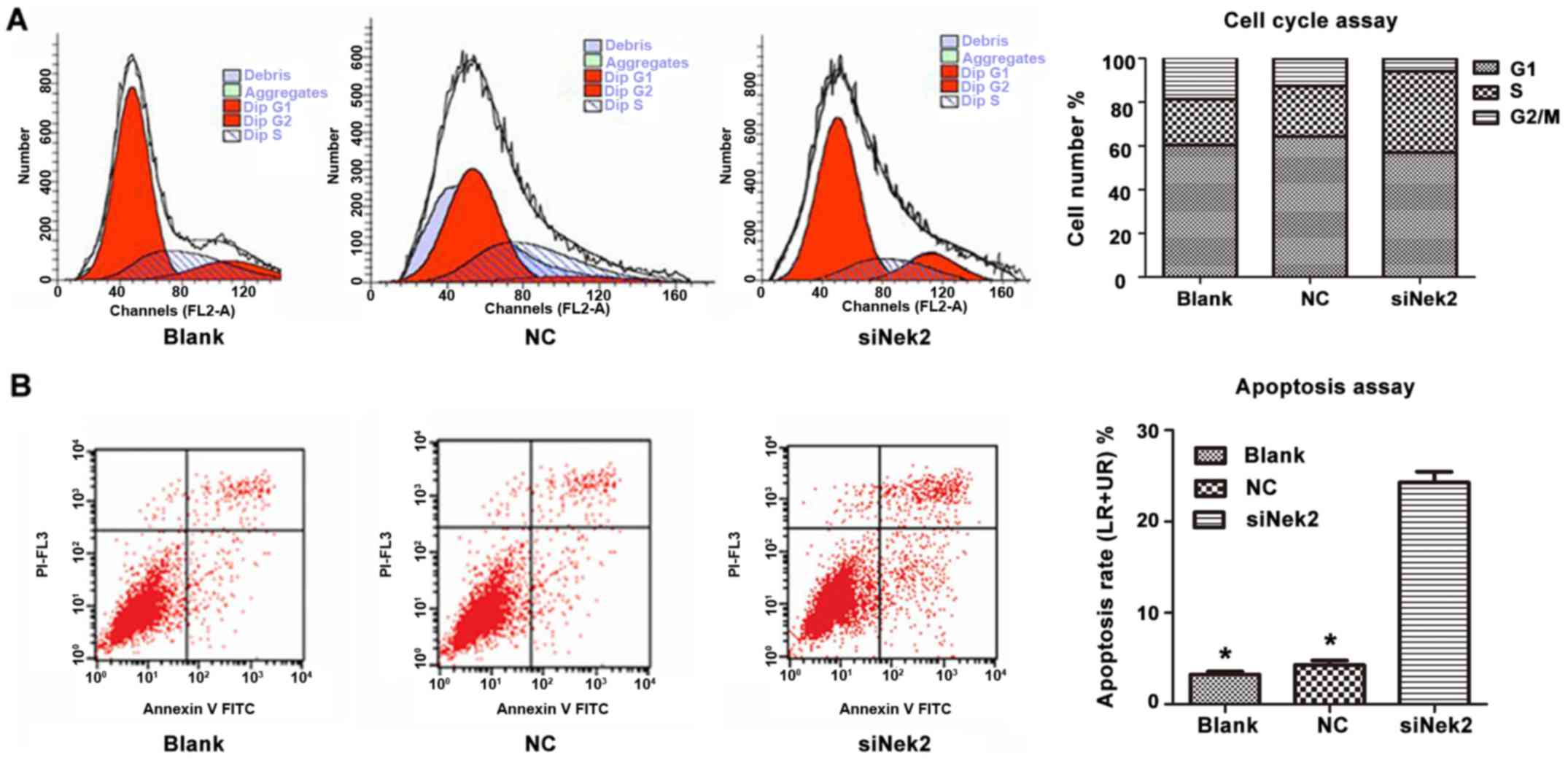
The cell cycle of HepG2 cells transfected with
either control siRNA or anti-Nek2 siRNA was analysed using PI
staining followed by fluorescence-activated cell sorting. This
revealed that the proportion of cells transfected with anti-Nek2
siRNA in the G2/M phase decreased. Furthermore, the proportion of
cells treated with anti-Nek2 siRNA in the S-phase increased,
whereas the proportion progressing through the G1/S-phase decreased
(Fig. 4A). These observations suggest
that Nek2 knockdown by specific siRNA in HepG2 cells induces cell
cycle arrest in the G2/M phase by retarding the S-phase. Moreover,
apoptosis analysis revealed that HepG2 cells treated with anti-Nek2
siRNA exhibited a significantly higher apoptotic index than
controls (Fig. 4B).
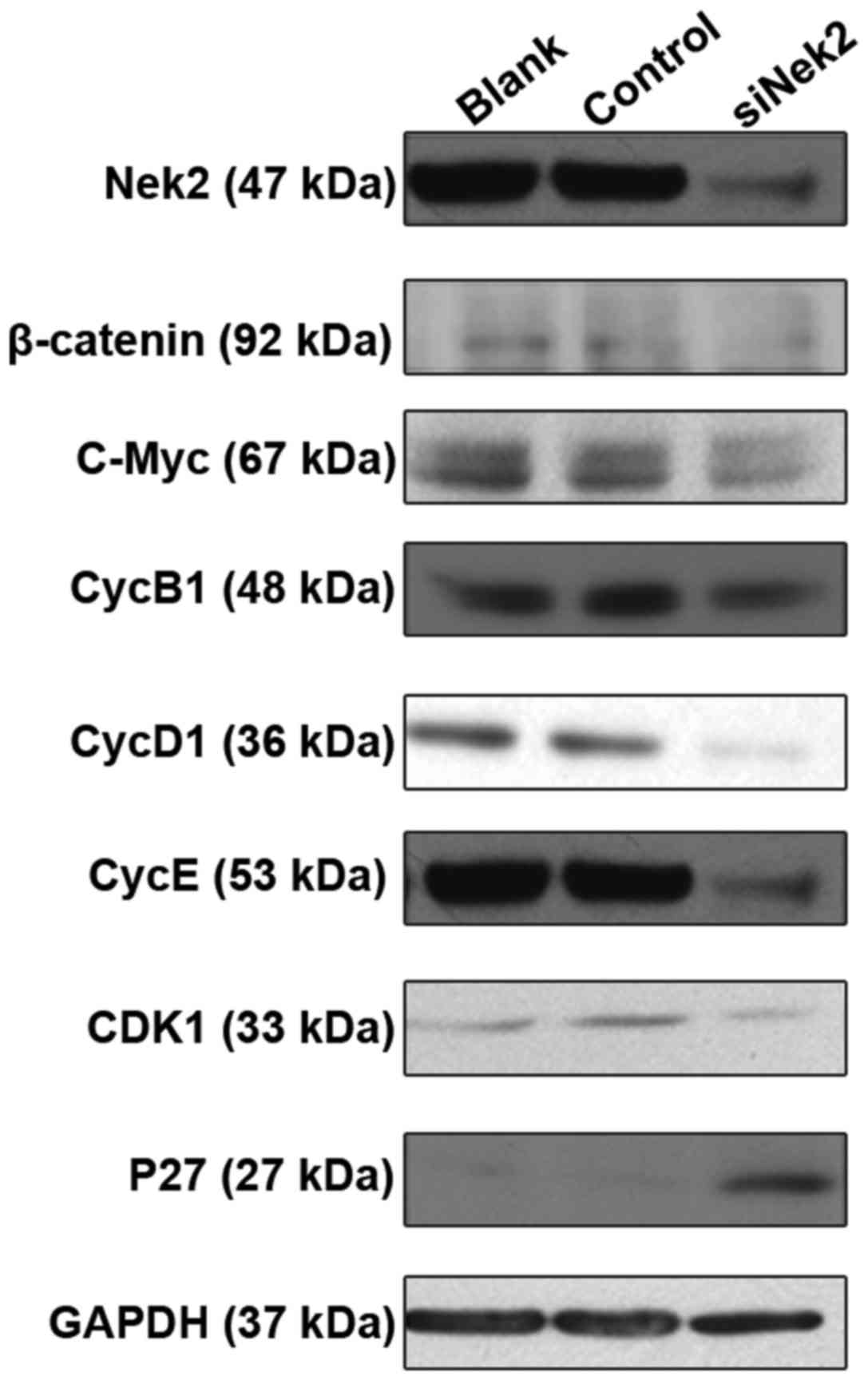
Nek2 regulates HCC growth through the
β-catenin/Wnt pathway
The possible mechanisms of Nek2 involvement in HCC
were detected by evaluating the protein levels of its established
targets, including β-catenin, C-Myc, CycD1, CycB1, CDK1, CycE, and
P27 genes. These are all involved in the β-catenin/Wnt pathway and
are essential for cell growth and proliferation. Western blot
analysis indicated that β-catenin protein expression was
compromised in HepG2 cells treated with Nek2-specific siRNA
(Fig. 5). Furthermore, Nek2
inhibition resulted in the diminished expression of C-Myc, CycD1,
CycB1, CDK1, and CycE proteins. Treatment of HepG2 cells with
control siRNA did not influence the protein expression of any of
the tested targets. However, western blot analysis also revealed
that Nek2 knock down in the HepG2 cells led to increased P27
protein expression. Overall, these results suggest that Nek2
expression induces cell cycle progression and proliferation by
regulating the β-catenin/Wnt pathway.
Discussion
The role of Nek2 in several human cancers has been
explored previously. Nek2 expression is increased in breast,
cervical, and prostate carcinoma cell lines, as well as in numerous
cell lines established from lymphomas (19). An assessment of ~1,700 genes revealed
marked up-regulation of Nek2 in cell lines derived from Ewing's
tumor and apediatric osteosarcoma (20); and an increase in Nek2 mRNA levels was
subsequently reported in non-Hodgkin lymphoma (21). Furthermore, in breast cancer models,
Nek2 plays a role in tumorigenesis and drug resistance (22). However, though Nek2 is well
established as an oncogene, its expression pattern and clinical
significance remains poorly defined in human HCC.
The present study demonstrated that the cellular
location of Nek2 was notably variable, with increased cytoplasmic
expression with or without nuclear Nek2 accumulation. These
differential results may be explained by Nek2's varying isoform
expression, as its bipartite nuclear localization sequence spans
the splice site (23).
Isoform-specific antibodies may enable the expression and
localization of specific Nek2 splice variants for further
investigation in HCC tissues.
In the present study, Nek2 expression was increased
in human HCC tissues, which has not been reported previously. HCC
cells expressed elevated levels of Nek2, compared to the negligible
levels detected in normal tissue cells. This may be due to the low
Nek2 mRNA copy numbers in healthy tissue. High Nek2 expression was
detected in 22 (52.4%) Edmondson stages I and II cases, and in
eight (80%) of the Edmondson stages III and IV cases, suggesting
that Nek2 is positively associated with more advanced HCC.
Furthermore, Nek2 expression in low-stage HCCs was lower than that
in high-stage HCCs. Meanwhile, other clinical association analyses
demonstrated that expression of Nek2 was positively correlated not
only with tumor histological grade and tumor metastasis, but also
with hepatitis C viral infection, as has been reported previously
(18). Thus, Nek2 may play an
important role in HCC progression and serve as a potential
biomarker for the biological behavior of HCC cells.
To elucidate the function of Nek2 in HCC cells,
anti-Nek2 siRNA was transfected into the HepG2 cell line, which
exhibited the highest Nek2 expression among all the seven HCC cell
lines tested. Nek2 knockdown decreased the proliferation rate and
colony formation ability in HepG2 cells, suggesting that decreased
Nek2 activity in HCC cells may decrease cancer cells' aggressive
growth and increased proliferative properties. The latter
hypothesis is supported by the established role of Nek2 in cell
proliferation. In A549 lung adenocarcinoma cells and human
embryonic kidney HEK293T cells, Nek2 overexpression led to the
activation of the Akt pathway and an increase in β-catenin protein
levels (24). β-Catenin binds to and
is phosphorylated by Nek2, and exists in a complex with Rootletin
(25). In addition, the expression of
CycD1, CycE, and C-Myc genes, which are required for cell
proliferation and regulated by Wnt/β-catenin signaling, is
controlled by Nek2 (24,26). Consistent with these data, blocking
Nek2 expression decreased levels of C-Myc, CycD1, and CycE1. The
analysis of the underlying mechanisms of Nek2-mediated control of
cell proliferation revealed that blocking Nek2 affected the cell
cycle by retarding G2/M transition. This result was congruent with
the expanded role of Nek2 as a regulator of CycB1, CDK1 and P27,
all of which promote cell cycle progression and stimulate the G2/M
transition.
Nek2 regulates the capacity for microtubule
organization of the centrosome at the time of mitotic entry
(26) and is involved in the
regulation of chromatin condensation, mitotic checkpoint signaling,
and cytokine activity (27). It may
trigger centrosome separation at the onset of mitosis through the
phosphorylation of multiple linker components.
Furthermore, Nek2 expression is positively
correlated with human proliferative disease. Nek2 regulates
β-catenin during centrosome separation by phosphorylation. In the
cell cycle, cytoplasmic and nuclear β-catenin levels increase in
the S-phase, peak at late G2/M, and abruptly decrease as the cells
re-enter G1 after mitosis (28).
Manipulating β-catenin levels, particularly in G2, affects
centrosome maturation and function, which highlights the role of
β-catenin in the centrosome (29).
When cytoplasmic β-catenin is stabilized, it can translocate into
the nucleus, where it interacts with a family of T cell
factor/lymphocyte enhancer factor transcription factors to drive
cell proliferation by directly inducting cell cycle regulators,
such as c-Myc, CycB1, and CycD1 (30,31).
However, high β-catenin levels may act via the Wnt signaling
pathway to stimulate cellular proliferation by dysregulating the
cell cycle (high C-Myc, CycD1, and CycE expression) and inhibit
tumor suppresion p16, p27, and wild-type p53; (32,33).
In conclusion, Nek2 is frequently upregulated in HCC
compared to healthy tissues, and is associated with HCC stages. The
present study demonstrates that blocking Nek2 by siRNA inhibits HCC
cell proliferation in vitro. This inhibition can be traced
back to the attenuation of the expression of proteins controlling
cell growth and proliferation. These results are a promising
starting point for the elucidation of the functional importance of
Nek2 in HCC progression. Further studies may allow Nek2 to serve as
a molecular target closely associated with cell proliferation for
the future development of HCC therapeutics.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81270037).
References
|
1
|
Meacham CE and Morrison SJ: Tumour
heterogeneity and cancer cell plasticity. Nature. 501:328–337.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bodzin AS and Busuttil RW: Hepatocellular
carcinoma: Advances in diagnosis, management, and long term
outcome. World J Hepatol. 7:1157–1167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Slotta JE, Kollmar O, Ellenrieder V,
Ghadimi BM and Homayounfar K: Hepatocellular carcinoma: Surgeon's
view on latest findings and future perspectives. World J Hepatol.
7:1168–1183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oda K, Uto H, Mawatari S and Ido A:
Clinical features of hepatocellular carcinoma associated with
nonalcoholic fatty liver disease: A review of human studies. Clin J
Gastroenterol. 8:1–9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Choi KJ, Baik IH, Ye SK and Lee YH:
Molecular targeted therapy for hepatocellular carcinoma: Present
status and future directions. Biol Pharm Bull. 38:986–991. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Y: Identification of differentially
expressed genes in hepatocellular carcinoma by laser capture
microdissection and microarrayGuangzhou Medical University;
2009
|
|
9
|
Fry AM: The Nek2 protein kinase: A novel
regulator of centrosome structure. Oncogene. 21:6184–6194. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schultz SJ, Fry AM, Sütterlin C, Ried T
and Nigg EA: Cell cycle-dependent expression of Nek2, a novel human
protein kinase related to the NIMA mitotic regulator of Aspergillus
nidulans. Cell Growth Differ. 5:625–635. 1994.PubMed/NCBI
|
|
11
|
Uto K and Sagata N: Nek2B, a novel
maternal form of Nek2 kinase, is essential for the assembly or
maintenance of centrosomes in early Xenopus embryos. Embo J.
19:1816–1826. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fry AM, Schultz SJ, Bartek J and Nigg EA:
Substrate specificity and cell cycle regulation of the Nek2 protein
kinase, a potential human homolog of the mitotic regulator NIMA of
Aspergillus nidulans. J Biol Chem. 270:12899–12905. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fletcher L, Cerniglia GJ, Nigg EA, Yend TJ
and Muschel RJ: Inhibition of centrosome separation after DNA
damage: A role for Nek2. Radiat Res. 162:128–135. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bowers AJ and Boylan JF: Nek8, a NIMA
family kinase member, is overexpressed in primary human breast
tumors. Gene. 328:135–142. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kokuryo T, Senga T, Yokoyama Y, Nagino M,
Nimura Y and Hamaguchi M: Nek2 as an effective target for
inhibition of tumorigenic growth and peritoneal dissemination of
cholangiocarcinoma. Cancer Res. 67:9637–9642. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suzuki K, Kokuryo T, Senga T, Yokoyama Y,
Nagino M and Hamaguchi M: Novel combination treatment for
colorectal cancer using Nek2 siRNA and cisplatin. Cancer Sci.
101:1163–1169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsunoda N, Kokuryo T, Oda K, Senga T,
Yokoyama Y, Nagino M, Nimura Y and Hamaguchi M: Nek2 as a novel
molecular target for the treatment of breast carcinoma. Cancer Sci.
100:111–116. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Drozdov I, Bornschein J, Wex T, Valeyev
NV, Tsoka S and Malfertheiner P: Functional and topological
properties in hepatocellular carcinoma transcriptome. Plos One.
7:e355102012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gu Z, Zhou W, Huang J, Yang Y, Wendlandt
E, Xu H, He X, Tricot G and Zhan F: Nek2 is a novel regulator of B
cell development and immunological response. Biomed Res Int.
2014:6210822014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wai DH, Schaefer KL, Schramm A, Korsching
E, Van Valen F, Ozaki T, Boecker W, Schweigerer L,
Dockhorn-Dworniczak B and Poremba C: Expression analysis of
pediatric solid tumor cell lines using oligonucleotide microarrays.
Int J Oncol. 20:441–451. 2002.PubMed/NCBI
|
|
21
|
Andreasson U, Dictor M, Jerkeman M,
Berglund M, Sundström C, Linderoth J, Rosenquist R, Borrebaeck CA
and Ek S: Identification of molecular targets associated with
transformed diffuse large B cell lymphoma using highly purified
tumor cells. Am J Hematol. 84:803–888. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Marina M and Saavedra HI: Nek2 and Plk4:
Prognostic markers, drivers of breast tumorigenesis and drug
resistance. Front Biosci (Landmark Ed). 19:352–365. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu W, Baxter JE, Wattam SL, Hayward DG,
Fardilha M, Knebel A, Ford EM, da Cruz e Silva EF and Fry AM:
Alternative splicing controls nuclear translocation of the cell
cycle-regulated Nek2 kinase. J Biol Chem. 282:26431–26440. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Das TK, Dana D, Paroly SS, Perumal SK,
Singh S, Jhun H, Pendse J, Cagan RL, Talele TT and Kumar S:
Centrosomal kinase Nek2 cooperates with oncogenic pathways to
promote metastasis. Oncogenesis. 2:e692013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mbom BC, Nelson WJ and Barth A: β-catenin
at the centrosome: Discrete pools of β-catenin communicate during
mitosis and may co-ordinate centrosome functions and cell cycle
progression. Bioessays. 35:804–969. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rapley J, Baxter JE, Blot J, Wattam SL,
Casenghi M, Meraldi P, Nigg EA and Fry AM: Coordinate regulation of
the mother centriole component nlp by nek2 and plk1 protein
kinases. Mol Cell Biol. 25:1309–1324. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hayward DG and Fry AM: Nek2 kinase in
chromosome instability and cancer. Cancer Lett. 237:155–166. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mbom BC, Siemers KA, Ostrowski MA, Nelson
WJ and Barth AI: Nek2 phosphorylates and stabilizes β-catenin at
mitotic centrosomes downstream of Plk1. Mol Biol Cell. 25:977–991.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Olmeda D, Castel S, Vilaro S and Cano A:
β-catenin regulation during the cell cycle: Implications in G2/M
and apoptosis. Mol Biol Cell. 14:2844–2860. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gehrke I, Gandhirajan RK and Kreuzer KA:
Targeting the WNT/β-catenin/TCF/LEF1 axis in solid and
haematological cancers: Multiplicity of therapeutic options. Eur J
Cancer. 45:2759–2767. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sekiya T, Nakamura T, Kazuki Y, Oshimura
M, Kohu K, Tago K, Ohwada S and Akiyama T: Overexpression of Icat
induces G(2) arrest and cell death in tumor cell mutants for
adenomatous polyposis coli, β-catenin, or Axin. Cancer Res.
62:3322–3326. 2002.PubMed/NCBI
|
|
32
|
Chou J, Lin YC, Kim J, You L, Xu Z, He B
and Jablons DM: Nasopharyngeal carcinoma-review of the molecular
mechanisms of tumorigenesis. Head Neck. 30:946–963. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Salomon A, Keramidas M, Maisin C and
Thomas M: Loss of β-catenin in adrenocortical cancer cells causes
growth inhibition and reversal of epithelial-to-mesenchymal
transition. Oncotarget. 6:11421–11433. 2015. View Article : Google Scholar : PubMed/NCBI
|