Introduction
Human lung cancer is the most frequently occurring
malignant tumor and the leading cause of cancer-associated
mortality worldwide (1). In 2015, it
was estimated that there would be 221,200 novel cases and 158,040
mortalities due to lung cancer in the United States of America
(2). Epidemiological studies have
demonstrated that a number of closely-associated risk factors
contribute to the carcinogenesis and progression of lung cancer,
including environmental pollution, smoking and occupational
carcinogens (3–6). Lung cancer has become a common
malignancy, particularly in China, mainly due to the increased rate
of cigarette smoking and environmental pollution (7). Non-small cell lung cancer (NSCLC), the
predominant category of lung cancer, accounts for ~85% of all cases
(8). According to its histology,
NSCLC may be divided into three subtypes: Squamous-cell carcinoma,
adenocarcinoma and large-cell carcinoma (9). Despite improvements in traditional
treatments, including surgery, followed by radiotherapy and/or
chemotherapy, the prognosis of NSCLC remains poor, with a 5-year
overall survival rate of 10–15% (10,11).
Therefore, an improved understanding of the molecular mechanisms
underlying the progression of NSCLC would be of benefit to the
investigation of novel therapeutic targets and would improve the
prognosis of patients with NSCLC.
The microRNAs (miRs/miRNAs) are a family of
endogenous, short and non-coding molecules inhibiting the
translation or degradation of target genes by combining with the
complementary sequences in the 3′-untranslated region (3′UTR) of
target genes (12). An miRNA may
regulate numerous target genes and one gene may be regulated by
multiple miRNAs. Therefore, >60% of all human genes have been
predicted to be regulated by miRNAs (13). The genes modulated by miRNAs have been
reported in various types of physiological processes. Thus, miRNAs
serve critical regulatory roles in diverse biological processes,
including proliferation, apoptosis, migration, invasion,
metastasis, cell differentiation and metabolism (12,14,15). A
previous study revealed that miRNAs may function as oncogenes or
tumor suppressors during tumor development and progression,
depending on the roles of their target genes, and that they are
aberrantly expressed in a number of human cancer types, including
bladder cancer (16), colorectal
cancer (17), gastric cancer
(18) and glioma (19). In conclusion, these findings indicated
that miRNAs demonstrate potential as efficient therapeutic targets
for cancer treatment.
In the present study, the expression level of
miR-320a in NSCLC tissues and cell lines was determined, and the
functions of miR-320a in NSCLC cells was evaluated. Furthermore,
the molecular mechanisms underlying its functions in NSCLC cells
were also investigated.
Materials and methods
Ethics statement and tissue
samples
The present study was approved by the Ethical
Committee of the Affiliated Hospital of Hebei University (Baoding,
Hebei, China). All patients provided written consent and were
informed of the purposes of the present study. Between October 2013
and February 2015, NSCLC tissues and the matched adjacent tissues
(normal tissues) were collected from 32 patients (19 male and 13
female; age range, 41–69 years) with NSCLC who had undergone
surgical resection at the Affiliated Hospital of Hebei University
(China). All the tissue samples were immediately snap-frozen in
liquid nitrogen and stored at −80°C.
Cell culture and oligonucleotide
transfection
The HEK293T cell line, human NSCLC H23, H522, SPC-A1
and A549 cell lines, and the 16HBE normal bronchial epithelial cell
line were purchased from the American Type Culture Collection
(Manassas, VA, USA). These cell lines were maintained in RPMI-1640
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) or
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) at 37°C in 5% CO2
in a cell incubator.
miR-320a mimics, negative control mimics (NC) and
luciferase report vectors [PGL3-IGF-1R-3′UTR wild-type (Wt) and
PGL3-IGF-1R-3′UTR mutation (Mut)] were synthesized by Shanghai
GenePharma Co., Ltd. (Shanghai, China). Insulin-like growth factor
1 receptor (IGF-1R) small interfering (si)RNA and control (ctrl)
siRNA were obtained from Guangzhou Ribobio Co., Ltd. (Guangzhou,
China). Cell transfection was performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the tissues and cell
lines using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Quantification of total RNA was determined using a
Nanodrop® ND-1000 spectrophotometer (NanoDrop
Technologies; Thermo Fisher Scientific, Inc., Pittsburgh, PA, USA).
Total RNA was synthesized into cDNA using the TaqMan®
MicroRNA Reverse Transcription kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The procedure for reverse transcription
was as follows: 16°C for 30 min; 42°C for 30 min, and 85°C for 5
min. The expression level of miR-320a was quantified using the
TaqMan® miRNA assay (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The qPCR was performed using 40 cycles of
denaturation at 95°C for 15 sec and annealing/extension at 60°C for
60 sec. U6 served as an endogenous control for miR-320a expression.
The SYBR®-Green RT-PCR kit (Takara Bio, Inc., Otsu,
Japan) was used to determine the IGF-1R mRNA expression level, with
GADPH as an internal control. The cycling conditions were as
follows: 42°C for 5 min; 95°C for 10 sec; and 40 cycles of 95°C for
5 sec, 55°C for 30 sec and 70°C for 30 sec. Each sample was
analyzed in triplicate. The relative expression of miR-320a and
IGF-1R mRNA was analyzed by use of the 2−ΔΔCq method
(20).
Cell proliferation assay
The cell proliferation assay was performed using the
MTT (Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) method.
Following 24 h of transfection, the cells were collected and
re-seeded into 96-well plates at a density of 3,000 cells/well. The
cell proliferation assays were performed every 24 h. In brief, 20
µl MTT solution (5 mg/ml) was added to DMEM supplemented with 10%
FBS and incubated for a further 4 h at 37°C. Subsequently, the
culture medium was removed and the remaining formazan precipitates
were dissolved in 150 µl dimethylsulfoxide (Sigma-Aldrich; Merck
Millipore). The optical density values of each well were detected
at a wavelength of 490 nm. This assay was performed in
triplicate.
Transwell cell invasion assay
The cell invasion ability was evaluated using the
Transwell cell invasion assay with Transwell plates coated with
Matrigel (BD Biosciences, San Jose, CA, USA). At 48 h
post-transfection, the cells were collected and resuspended in
FBS-free DMEM. Transfected cells (1×105) were added to
the upper chamber of the Transwell plates and 500 µl culture medium
supplemented with 20% FBS was added to the lower portion of the
chamber. Following incubation for 48 h at 37°C, the non-invading
cells were carefully removed using cotton wool. The cells that
invaded the lower surface of the Transwell membrane were fixed with
95% ethanol, stained with 0.1% crystal violet (Beyotime Institute
of Biotechnology, Haimen, China) and quantified under a light
microscope.
Western blot analysis
The total protein was isolated from the transfected
cells and the concentration of total protein was determined using
the Pierce bicinchoninic acid assay (Thermo Fisher Scientific,
Inc.). Equal amounts of protein (20 µg/lane) were fractionated by
SDS-PAGE on a 10% gel and transferred to polyvinylidene fluoride
membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
membranes were blocked using 5% skimmed dry milk at room
temperature for 1 h, followed by immunostaining with the following
primary antibodies overnight at 4°C: Mouse anti-human monoclonal
IGF-1R (1:1,000 dilution; cat. no. sc-81464; Santa Cruz
Biotechnology, Inc.) and GAPDH (1:1,000 dilution; cat. no.
sc-59540; both Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
The membranes were washed with Tris-buffered saline solution
containing 0.1% Tween 20 and probed with horseradish
peroxidase-conjugated secondary antibodies (both 1:5,000 dilution;
cat. no. sc-2005 for IGF-1R, cat no. sc-2064 for GAPDH; both Santa
Cruz Biotechnology, Inc.) at room temperature for 1 h. Protein
bands were visualized using a chemiluminescent detection system
(Thermo Fisher Scientific, Inc.).
Luciferase report assay
The HEK293T cells were seeded in 24-well plates at
40–50% confluence and co-transfected with PGL3-IGF-1R-3′UTR Wt or
PGL3-IGF-1R-3′UTR Mut, miR-320a mimics or NC, using Lipofectamine
2000. Following 48 h of transfection, a Dual-Luciferase®
Reporter assay system (Promega Corporation, Madison, WI, USA) was
used to detect firefly and Renilla luciferase activities using the
GLOMAX multimode reader (Promega Corporation). Renilla activity was
evaluated as an internal control for firefly activity.
Statistical analysis
SPSS 13 (SPSS, Inc., Chicago, IL, USA) was used for
the statistical analysis. Data are presented as the mean ± standard
deviation. A two-tailed P-value of <0.05 was considered to
indicate a statistically significant difference.
Results
miR-320a is downregulated in NSCLC
tissues and cell lines
In order to investigate the functions of miR-320a in
NSCLC cells, the present study first determined the expression
level of miR-320a in NSCLC tissues and matched adjacent tissues
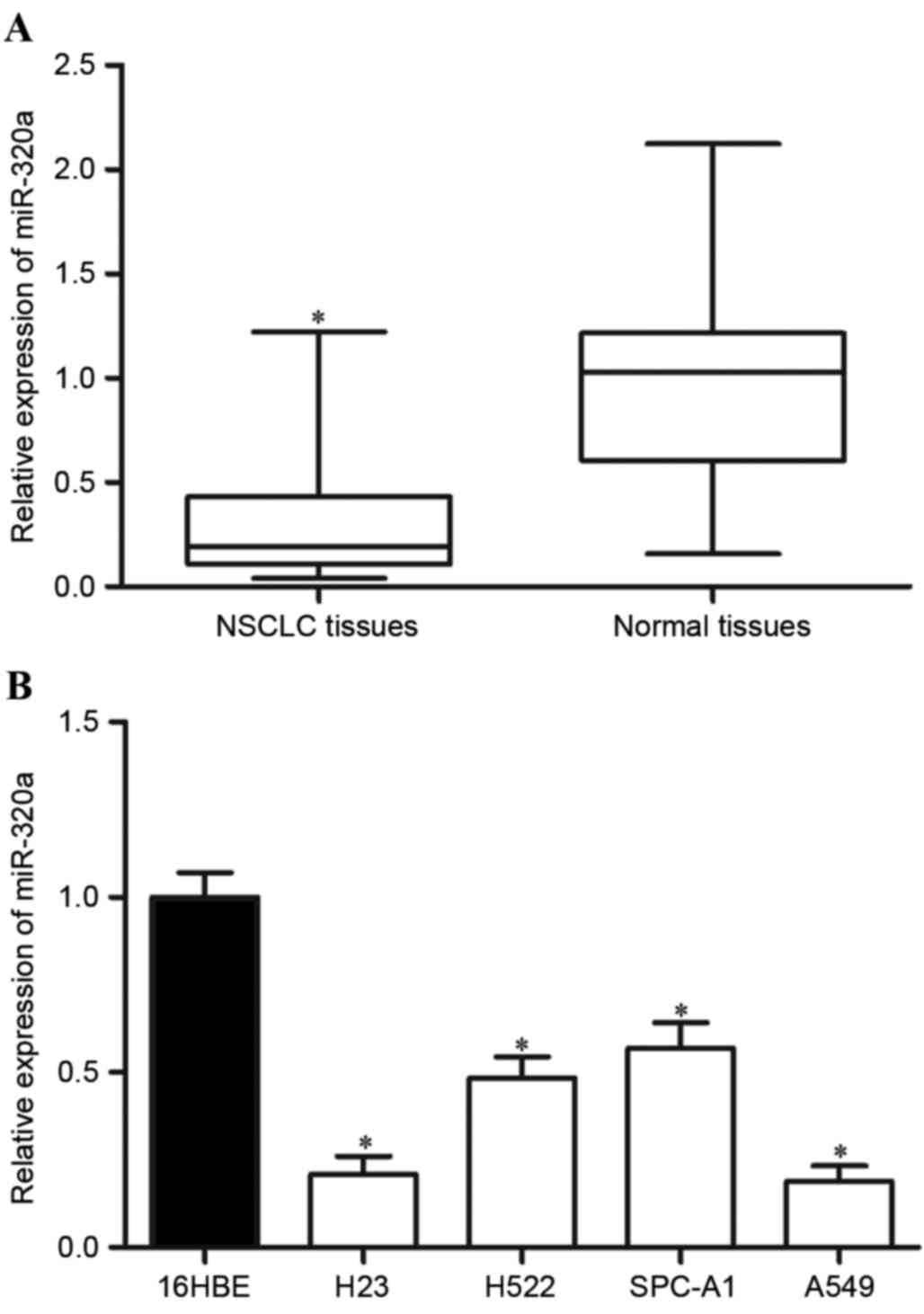
using RT-qPCR. As demonstrated in Fig.
1A, miR-320a was significantly downregulated in NSCLC tissues
compared with that in matched adjacent tissues (P=0.006).
Furthermore, the miR-320a expression level was evaluated in NSCLC
cell lines and a normal bronchial epithelial cell line (16HBE). As
revealed in Fig. 1B, reduced miR-320a
expression levels were also observed in the four NSCLC cell lines
compared with that in the 16HBE cell line. These results indicated
that miR-320a was frequently downregulated in NSCLC tissues in
addition to NSCLC cell lines, and the low expression level may have
contributed to the initiation and progression of NSCLC.
Overexpression of miR-320a inhibits
the proliferation and invasion of NSCLC cells
In order to investigate the functions of miR-320a in
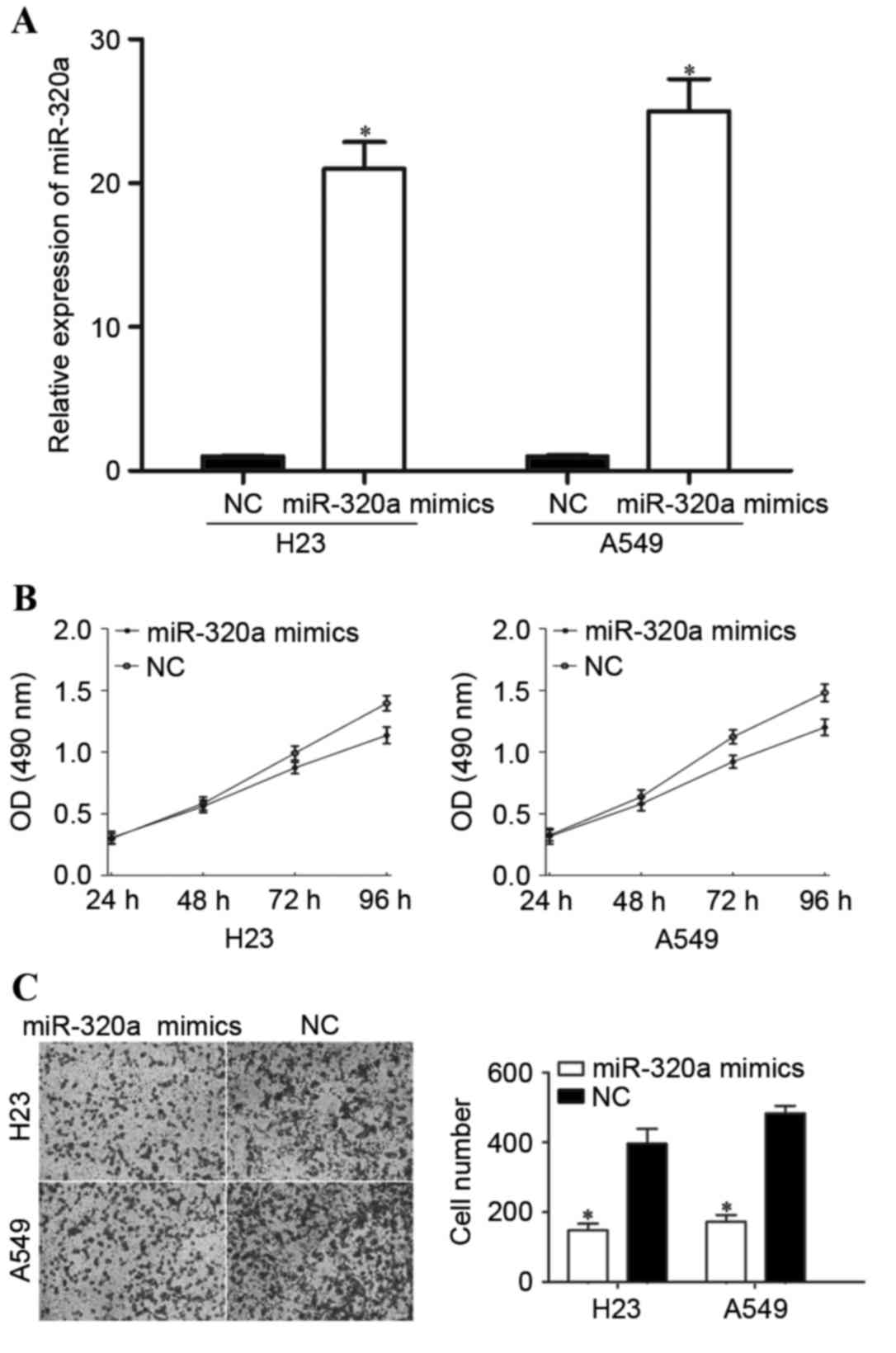
NSCLC, H23 and A549 cells were transfected with miR-320a mimics or
NC. The efficiency of the transfection was determined by RT-qPCR.
As demonstrated in Fig. 2A, miR-320a
was significantly upregulated in miR-320a mimic-transfected H23 and
A549 cells compared with cells transfected with NC.
A cell proliferation assay was performed in order to
investigate the effect of miR-320a overexpression on the
proliferation of the NSCLC cells. As demonstrated in Fig. 2B, the overexpression of miR-320a
significantly inhibited cell proliferation in the H23 (P=0.016) and
A549 (P=0.010) cells compared with cells transfected with NC.
Furthermore, Transwell cell invasion assays were performed in order
to investigate the role of miR-320a in NSCLC cell invasion ability.
The results revealed that the invasion ability was significantly
decreased in H23 (P=0.020) and A549 (P=0.026) cells transfected
with miR-320a mimics compared with cells transfected with NC
(Fig. 2C). These results suggested
that miR-320a acted as a tumor suppressor in NSCLC.
miR-320a negatively regulates IGF-1R
expression by directly binding to its 3′-untranslated region
(3′UTR)
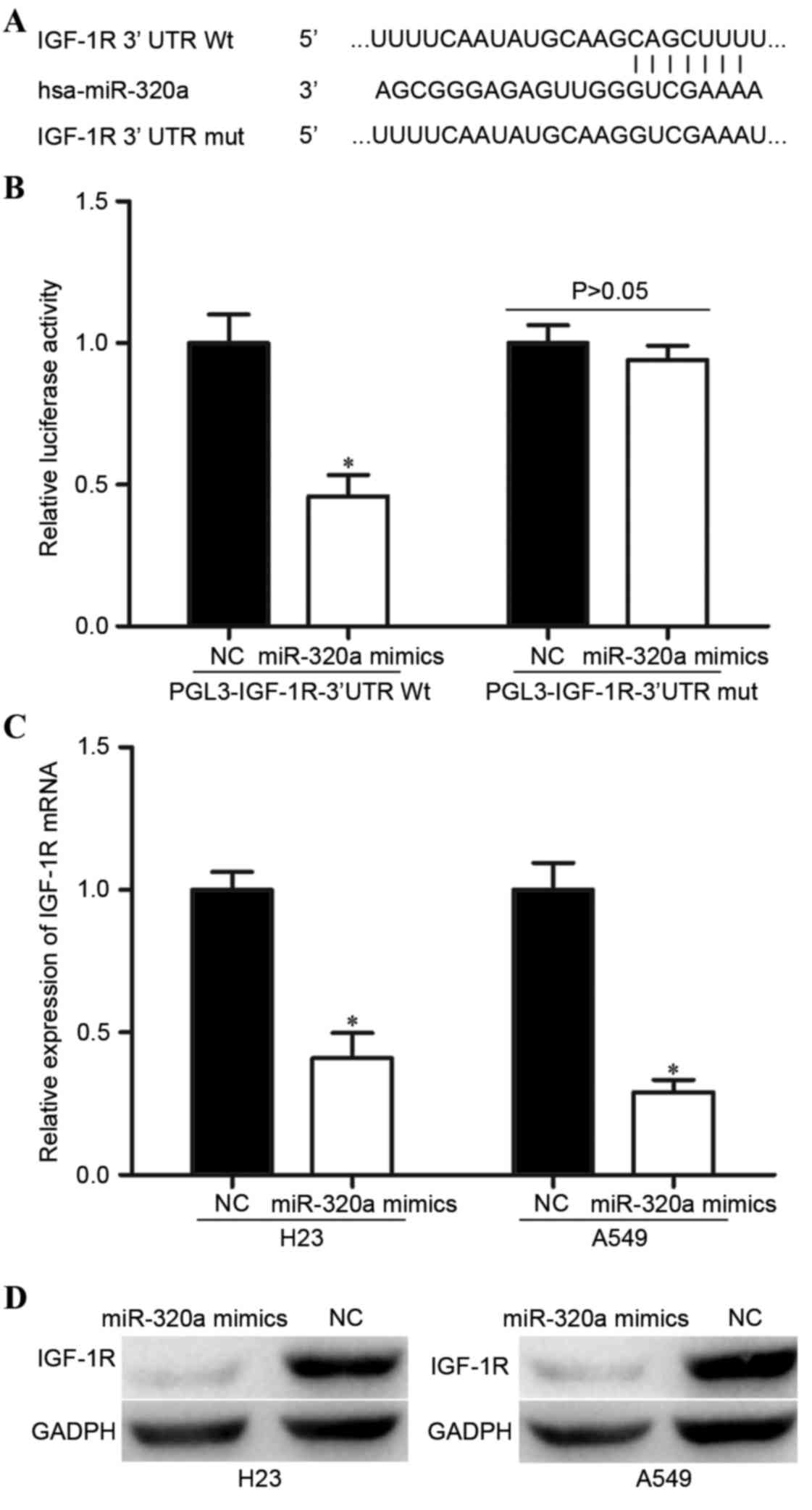
In order to find a potential target gene of miR-320a
that may contribute to its suppressive functions in NSCLC cells,
bioinformatic analysis was performed. As presented in Fig. 3A, there was complementarity between
miR-320a and the 3′UTR region of IGF-1R. The direct regulation of
IGF-1R by miR-320a in the NSCLC cells was confirmed by luciferase
reporter assay. The luciferase reporter assay demonstrated markedly
decreased activity with miR-320a mimics and PGL3-IGF-1R-3′UTR Wt
compared with the NC and PGL3-IGF-1R-3′UTR Mut (Fig. 3B).
In addition, RT-qPCR and western blot analysis was
performed in order to confirm the regulatory effect of miR-320a on
IGF-1R expression level in the NSCLC cells. As demonstrated in
Fig. 3C and D, miR-320a mimics
decreased the IGF-1R expression level in the H23 and A549 cells at
the mRNA and protein levels. Taken together, these results
indicated that miR-320a negatively regulated IGF-1R expression by
directly binding to its 3′UTR region in the NSCLC cells.
IGF-1R serves a role in
miR-320a-induced functions in NSCLC cells
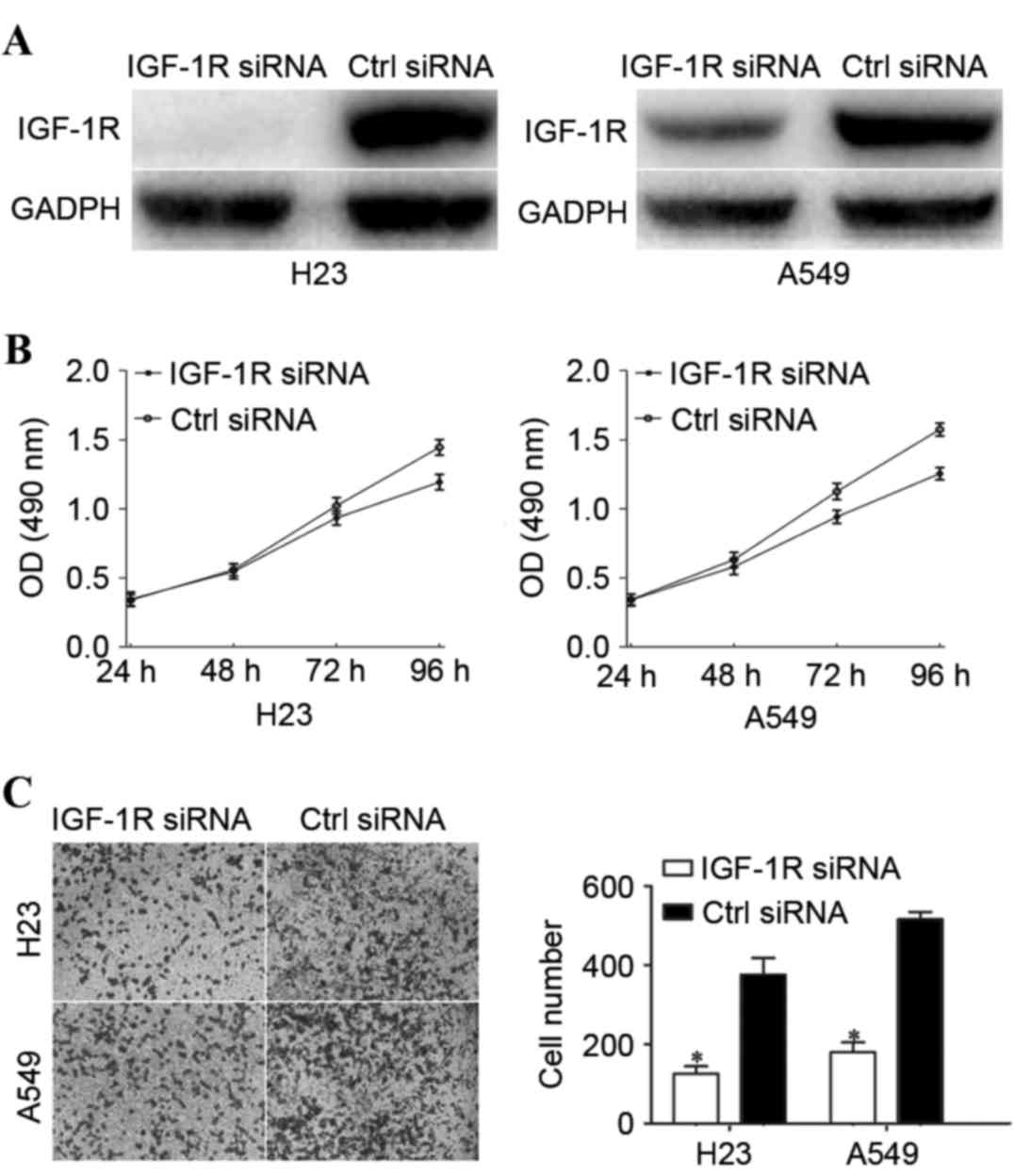
In order to investigate whether the functions of
miR-320a on NSCLC cells were dependent on IGF-1R, the H23 and A549
cells were transfected with IGF-1R siRNA or ctrl siRNA, followed by
evaluation by cell proliferation and Transwell cell invasion
assays. Firstly, western blot analysis was performed to determine
the transfection efficiency. As presented in Fig. 4A, the IGF-1R expression level was
decreased in the H23 and A549 cells following transfection with
IGF-1R siRNA. Furthermore, cell proliferation and Transwell cell
invasion assays revealed that IGF-1R-knockdown significantly
suppressed cell proliferation (Fig.
4B) and invasion (Fig. 4C),
compared with the ctrl siRNA groups. These results indicated that
the functions of IGF-1R downregulation were similar to the
functions induced by miR-320a overexpression in NSCLC cells,
rendering IGF-1R as a direct functional target of miR-320a in NSCLC
cells.
Discussion
NSCLC is the leading cause of cancer-associated
mortality worldwide and in China (21). There has been progression towards the
development of therapeutic treatments for NSCLC patients (22). However, the overall five-year survival
rate of NSCLC has remained unfavorable since the 1970s (23). A number of previous studies have
demonstrated that miRNAs may be critical regulators in
cancer-associated processes (24,25).
Therefore, understanding the association between NSCLC and abnormal
expression levels of miRNAs may be beneficial for the
identification of novel efficient therapeutic targets, in order to
improve the prognosis and survival rates of patients with NSCLC.
The present study demonstrated that miR-320a was downregulated in
NSCLC tissues and cell lines. The suppressive functions of miR-320a
on the proliferation and invasion of NSCLC cells was demonstrated
in the present study, and indicated that miR-320a may be involved
in the carcinogenesis and progression of NSCLC. Furthermore, a
molecular association between miR-320a and IGF-1R was validated in
the NSCLC cells.
The expression levels of miR-320a have been revealed
to be reduced in breast cancer (26,27), colon
cancer (28), bladder cancer
(29), glioblastoma (30) and salivary adenoid cystic carcinoma
(31). In breast cancer, miR-320a was
downregulated in tumor tissues, and miR-320a expression levels were
significantly associated with tumor size, clinical stage, lymph
node metastasis and distant metastasis (27). Patients with breast cancer and low
expression levels of miR-320a demonstrated shorter overall survival
times compared with patients with higher expression levels of
miR-320a. Univariate and multivariate analyses revealed that
miR-320a was an independent prognostic biomarker for invasive
breast cancer (26,27). Sun et al (31) demonstrated that miR-320a expression
levels were lower in salivary adenoid cystic carcinoma tissues with
metastasis compared with tissues without metastasis. The low
expression level of miR-320a indicates a poor prognosis and rapid
metastasis for patients with salivary adenoid cystic carcinoma
(31). Furthermore, multivariate
analysis indicated that miR-320a expression level was an
independent indicator of lung metastasis (31). These findings suggested that miR-320a
was downregulated in these cancers, and may be a potential
prognostic factor.
miR-320a has previously been validated as a tumor
suppressor in numerous types of human cancer (26,28,32). For
example, in colon cancer, the overexpression of miR-320 inhibited
cell proliferation by the downregulation of β-catenin (28). Zhang et al (32) revealed that miR-320a expression level
restoration suppressed the migration and invasion abilities of
colon cancer by directly targeting neuropilin 1. In breast cancer,
re-expression of miR-320a significantly inhibited the proliferation
and invasion of breast cancer cells by the blockade of Ras-related
protein Rab-11A (26). Lu et
al (33) demonstrated that
enforced miR-320a expression was sufficient to sensitize
tamoxifen-resistant breast cancer cells to tamoxifen by targeting
the cyclic adenosine 3′, 5′-monophosphate-regulated phosphoprotein
and estrogen-related receptor γ, in addition to their downstream
effectors, c-Myc and Cyclin D1. In chronic myeloid leukemia,
miR-320a suppressed K562 cell growth and metastasis, and enhanced
apoptosis by directly targeting BCR, RhoGEF and GTPase activating
protein/ABL proto-oncogene 1, non-receptor tyrosine kinase
(34). Qi et al (35) revealed that the upregulation of
miR-320a significantly decreased nasopharyngeal carcinoma cell
proliferation, migration and invasion in vitro, and tumor
growth in vivo by negative regulation of B cell-specific
Moloney murine leukemia virus integration site 1. Together these
findings suggested that miR-320a acts as a tumor suppressor, and
may be worthy of investigation as a potential anticancer drug for
these cancer types.
Identification of cancer-specific miRNAs, in
addition to their target genes, is important for elucidating the
functions of miRNAs in carcinogenesis and may provide promising
therapeutic targets. In the present study, IGF-1R was identified as
a direct target gene of miR-320a in NSCLC cells. Firstly, it was
revealed that miR-320a could target the 3′UTR sequence of IGF-1R by
performing bioinformatic analysis. Furthermore, the luciferase
reporter assay demonstrated that miR-320a suppressed the luciferase
activity in cells transfected with PGL3-IGF-1R-3′UTR Wt compared
with PGL3-IGF-1R-3′UTR Mut, indicating that miR-320a directly
targeted the 3′UTR of IGF-1R. Additionally, the overexpression of
miR-320a reduced the IGF-1R expression level in NSCLC cells at the
mRNA and protein levels. Finally, knockdown of IGF-1R by siRNA had
similar effects compared with miR-320a overexpression in NSCLC
cells, which demonstrated that IGF-1R was the direct functional
target of miR-320a in NSCLC cells. IGF-1R is a transmembrane
tyrosine kinase receptor that contains two extracellular α subunits
and two transmembrane β subunits (27). Previous functional studies have
revealed that IGF-1R serves important functions in numerous
cancer-associated biological processes, including cell
proliferation, anti-apoptosis, vascularization, migration, invasion
and metastasis (36,37). Together these findings suggested that
IGF-1R is worth paying close attention to as a potential target for
the inhibition of NSCLC.
In conclusion, the present study demonstrated that
miR-320a was downregulated in NSCLC tissues and cell lines.
Overexpression of miR-320a inhibited the proliferation and invasion
of NSCLC cells by directly binding to the IGF-1R 3′UTR in
vitro. In the future, the aberrant expression of miR-320a may
be utilized in therapeutic interventions for patients with
NSCLC.
Acknowledgements
The present study was supported by The National
Natural Science Fund (grant no. 81472744).
References
|
1
|
Shi WY, Liu KD, Xu SG, Zhang JT, Yu LL, Xu
KQ and Zhang TF: Gene expression analysis of lung cancer. Eur Rev
Med Pharmacol Sci. 18:217–228. 2014.PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boffetta P and Nyberg F: Contribution of
environmental factors to cancer risk. Br Med Bull. 68:71–94. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Didkowska J, Manczuk M, McNeill A, Powles
J and Zatonski W: Lung cancer mortality at ages 35–54 in the
European Union: Ecological study of evolving tobacco epidemics.
BMJ. 331:189–191. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ridge CA, McErlean AM and Ginsberg MS:
Epidemiology of lung cancer. Semin Intervent Radiol. 30:93–98.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paliogiannis P, Attene F, Cossu A, Budroni
M, Cesaraccio R, Tanda F, Trignano M and Palmieri G: Lung cancer
epidemiology in North Sardinia, Italy. Multidiscip Respir Med.
8:452013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang B, Liu T, Wu T, Wang Z, Rao Z and
Gao J: microRNA-137 functions as a tumor suppressor in human
non-small cell lung cancer by targeting SLC22A18. Int J Biol
Macromol. 74:111–118. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Youlden DR, Cramb SM and Baade PD: The
International Epidemiology of lung cancer: Geographical
distribution and secular trends. J Thorac Oncol. 3:819–831. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoda S, Soejima K, Hamamoto J, Yasuda H,
Nakayama S, Satomi R, Terai H, Ikemura S, Sato T, Naoki K and
Betsuyaku T: Claudin-1 is a novel target of miR-375 in
non-small-cell lung cancer. Lung Cancer. 85:366–372. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
de Sanchez Cos J, Sojo Gonzélez MA,
Montero MV, Pérez Calvo MC, Vicente MJ and Valle MH: Non-small cell
lung cancer and silent brain metastasis. Survival and prognostic
factors. Lung Cancer. 63:140–145. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Crino L, Weder W, van Meerbeeck J and
Felip E: ESMO Guidelines Working Group: Early stage and locally
advanced (non-metastatic) non-small-cell lung cancer: ESMO clinical
practice guidelines for diagnosis, treatment and follow-up. Ann
Oncol. 21:(Suppl 5). v103–v115. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lewis BP, Shih IH, Jones-Rhoades MW,
Bartel DP and Burge CB: Prediction of mammalian microRNA targets.
Cell. 115:787–798. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Broderick JA and Zamore PD: MicroRNA
therapeutics. Gene Ther. 18:1104–1110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu G, Jia Z and Dou Z: miR-24-3p regulates
bladder cancer cell proliferation, migration, invasion and
autophagy by targeting DEDD. Oncol Rep. 37:1123–1131.
2017.PubMed/NCBI
|
|
17
|
Zu C, Liu T and Zhang G: MicroRNA-506
inhibits malignancy of colorectal carcinoma cells by targeting
LAMC1. Ann Clin Lab Sci. 46:666–674. 2016.PubMed/NCBI
|
|
18
|
Bao J, Zou JH, Li CY and Zheng GQ: miR-194
inhibits gastric cancer cell proliferation and tumorigenesis by
targeting KDM5B. Eur Rev Med Pharmacol Sci. 20:4487–4493.
2016.PubMed/NCBI
|
|
19
|
Qi Z, Cai S, Cai J, Chen L, Yao Y, Chen L
and Mao Y: miR-491 regulates glioma cells proliferation by
targeting TRIM28 in vitro. BMC Neurol. 16:2482016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Heist RS: First-line systemic therapy for
non-small cell lung cancer. Hematol Oncol Clin North Am. 31:59–70.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ge H, Li B, Hu WX, Li RJ, Jin H, Gao MM
and Ding CM: MicroRNA-148b is down-regulated in non-small cell lung
cancer and associated with poor survival. Int J Clin Exp Pathol.
8:800–805. 2015.PubMed/NCBI
|
|
24
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mishra PJ and Merlino G: MicroRNA
reexpression as differentiation therapy in cancer. J Clin Invest.
119:2119–2123. 2009.PubMed/NCBI
|
|
26
|
Wang B, Yang Z, Wang H, Cao Z, Zhao Y,
Gong C, Ma L, Wang X, Hu X and Chen S: MicroRNA-320a inhibits
proliferation and invasion of breast cancer cells by targeting
RAB11A. Am J Cancer Res. 5:2719–2729. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang H, Yu J, Wang L, Ding D, Zhang L, Chu
C, Chen Q, Xu Z, Zou Q and Liu X: miR-320a is an independent
prognostic biomarker for invasive breast cancer. Oncol Lett.
8:1043–1050. 2014.PubMed/NCBI
|
|
28
|
Sun JY, Huang Y, Li JP, Zhang X, Wang L,
Meng YL, Yan B, Bian YQ, Zhao J, Wang WZ, et al: MicroRNA-320a
suppresses human colon cancer cell proliferation by directly
targeting β-catenin. Biochem Biophys Res Commun. 420:787–792. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shang C, Zhang H, Guo Y, Hong Y, Liu Y and
Xue Y: MiR-320a down-regulation mediates bladder carcinoma invasion
by targeting ITGB3. Mol Biol Rep. 41:2521–2527. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo T, Feng Y, Liu Q, Yang X, Jiang T,
Chen Y and Zhang Q: MicroRNA-320a suppresses in GBM patients and
modulates glioma cell functions by targeting IGF-1R. Tumour Biol.
35:11269–11275. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun L, Liu B, Lin Z, Yao Y, Chen Y, Li Y,
Chen J, Yu D, Tang Z, Wang B, et al: MiR-320a acts as a prognostic
factor and inhibits metastasis of salivary adenoid cystic carcinoma
by targeting ITGB3. Mol Cancer. 14:962015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, He X, Liu Y, Ye Y, Zhang H, He P,
Zhang Q, Dong L, Liu Y and Dong J: microRNA-320a inhibits tumor
invasion by targeting neuropilin 1 and is associated with liver
metastasis in colorectal cancer. Oncol Rep. 27:685–694.
2012.PubMed/NCBI
|
|
33
|
Lu M, Ding K, Zhang G, Yin M, Yao G, Tian
H, Lian J, Liu L, Liang M, Zhu T and Sun F: MicroRNA-320a
sensitizes tamoxifen-resistant breast cancer cells to tamoxifen by
targeting ARPP-19 and ERRγ. Sci Rep. 5:87352015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xishan Z, Ziying L, Jing D and Gang L:
MicroRNA-320a acts as a tumor suppressor by targeting BCR/ABL
oncogene in chronic myeloid leukemia. Sci Rep. 5:124602015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qi X, Li J, Zhou C, Lv C and Tian M:
MicroRNA-320a inhibits cell proliferation, migration and invasion
by targeting BMI-1 in nasopharyngeal carcinoma. FEBS Lett.
588:3732–3738. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Werner H and LeRoith D: The role of the
insulin-like growth factor system in human cancer. Adv Cancer Res.
68:183–223. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pollak M: The insulin and insulin-like
growth factor receptor family in neoplasia: An update. Nat Rev
Cancer. 12:159–169. 2012.PubMed/NCBI
|