Introduction
Tetrandrine (Tet), a natural product isolated from
Stephania tetrandra, has been reported to be an anti-tumor
and anti-inflammatory drug (1–8).
Previously, the present authors have demonstrated that Tet induces
cell death in 11 cancer cell lines (1). Tet directly neutralizes the lysosomal
acidity, blocks the autophagic flux in the degradation step and
causes energetic impairment, which eventually results in cell death
(1). By contrast, Tet also acts as an
autophagy agonist by inducing the synthesis of reactive oxygen
species (3,4,9–11), indicating that Tet may function as a
multi-target drug to regulate autophagy in cancer cells. Regardless
of the Tet target, the present authors and other groups have
previously consistently observed that Tet increases the number of
autophagosomes, levels of microtubule-associated protein 1 light
chain 3, type II (LC3-II) and the number of enhanced green
fluorescent protein-LC3 puncta in cancer cells (1,3,9).
DU145, a commonly used prostate cancer cell line for
research, is different from PC-3 and LNCaP cells in terms of
autophagy regulation (12). As
alternative transcripts of autophagy related 5 (ATG5) lack one or
two exons in DU145 cells, and this causes the autophagy pathway to
be genetically impaired (12). By
contrast, DU145 cells exhibited the highest sensitivity to Tet in
all of the 11 cancer cell lines that were previously tested by the
authors of the present study (1),
implying that impairment in autophagy may increase Tet-induced cell
death. In order to investigate the role of autophagy on Tet-induced
cell death in DU145 cells, the ultrastructural changes following
Tet treatment were observed by transmission electron microscopy
(TEM). Notably, Tet treatment triggered an accumulation of
autophagosomes in this cell line, indicating that alternative
autophagy was involved.
Materials and methods
Tetrandrine and other reagents
Tet (Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) was dissolved as previously described (1). In brief, Tet powder was dissolved in one
drop of 0.1 mM HCl and neutralized with NaHCO3 solution
to 7.4 pH Tet was subsequently diluted with phosphate-buffered
saline (PBS) for a final concentration of 10 mM. Chloroquine (CQ)
was purchased from Sigma-Aldrich (Merck Millipore, Darmstadt,
Germany) and dissolved in sterilized deionized water for a final
concentration of 100 mM. Rabbit anti-LC3B antibody was purchased
from Sigma-Aldrich, Merck Millipore. Mouse anti-β-actin antibody
was purchased from Santa Cruz Biotechnology Inc. Other primary
(β-actin; cat. no., 8457; dilution, 1:2,000; p62, cat. no., 8025;
dilution, 1:1,000) and anti-rabbit secondary antibody (cat. no.,
7074; dilution, 1:5,000) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Lipofectamine 3000 reagent and
LysoSensor Yellow/Blue DND-160 were obtained from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). The EGFP-LC3 plasmid was
obtained from Osaka University, Osaka, Japan.
Cell culture and transfection
DU145 and PC-3 cells (National Platform of
Experimental Cell Resources for Sci-Tech Beijing, China) were
cultured in RPMI-1640 medium (Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific,
Inc.), 100 units/ml penicillin and 0.1 mg/ml streptomycin (Thermo
Fisher Scientific, Inc.), and maintained in an atmosphere of 5%
CO2 at 37°C. The cells were digested with 0.25% of
trypsin-EDTA solution for passage, and the cells were serum starved
for 24 h prior to treatment.
For transfection, 1×105/ml cells were
seeded in a 6-well plate and transiently transfected with EGFP-LC3
plasmid at a final concentration of 50 nM by using Lipofectamine
3000 reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Following 6 h of incubation at 37°C, the
supernatant was replaced with RPMI-1640 medium containing 1% FBS
and incubated at 37°C for 48 h.
Cell viability measurement
Cells were digested and seeded
(1×104/well) onto a 96-well plate. Cell viability was
detected with Cell Counting Kit-8 (CCK-8) assay (Dojindo Molecular
Technologies, Inc. Kumamoto, Japan). Briefly, cells were treated
with Tet or CQ and incubated at 37°C in RPMI-1640 medium containing
10% (v/v) CCK-8 for 2 h. Optical densities were read at 450 nm by
using a microplate reader (Thermo Fisher Scientific, Inc.).
Autophagy assays
DU145 cells were transfected with EGFP-LC3 plasmid
and cultured at 37°C for 48 h. The EGFP-LC3 puncta formation in
each group were observed and counted using fluorescence microscopy
(Leica Microsystems GmbH, Wetzlar, Germany) (13,14).
TEM
TEM was performed as previously described (1). In brief, the cells were digested, washed
with pre-cold PBS, centrifuged at 1,000 × g for 5 min at 4°C and
fixed with 2% glutaraldehyde at 4°C overnight. The cell pellets
were post-fixed with 1% osmium tetroxide for 1 h. The samples were
subsequently dehydrated and cut into ultrathin 40-nm sections. The
subcellular structures of cells in each group were observed with an
electron microscope (JEOL Ltd., Tokyo, Japan).
Lysosome pH quantification
Lysosomal pH was quantified as previously described
(1). A total of 1×105/ml
cells were seeded in a 96-well plate, treated with Tet and washed
with PBS. The cells were subsequently incubated in 5 µM of
LysoSensor Yellow/Blue DND-160 (Thermo Fisher Scientific, Inc.) at
room temperature for 5 min. The supernatants were removed and
replaced with 100 µl RPMI-1640 medium. Cells used for the standard
curve were incubated at room temperature with 10 µM monensin and 20
µM nigericin in 20 mM 2-(N-morpholino) ethanesulfonic acid, 110 mM
KCl and 20 mM NaCl at pH 3.6 to 6.9 for 10 min. Fluorescence in
each well was assessed based on the ratio of light excited at 329
nm vs. 384 nm.
Western blotting
Total protein from each group was extracted with
radioimmunoprecipitation assay (RIPA) lysis buffer (50 mM Tris-HCl,
150 mM NaCl, 1 mM EDTA, 1% NP-40 and 0.1% sodium dodecyl sulfate).
The lysates were centrifuged at 8,000 × g at 4°C for 5 min and the
supernatants were collected and subsequently quantified. The
lysates in each group were diluted to the same concentration with
RIPA lysis buffer, mixed with loading buffer and denatured at 95°C
for 5 min. A total of 20 µg of the samples were loaded onto 12%
SDS-PAGE and separated with electrophoresis. The proteins were then
blotted onto a polyvinylidene difluoride membrane and blocked with
5% non-fat milk for 1 h at room temperature. The membrane was
incubated with diluted primary antibodies at 4°C overnight. The
membrane was washed with TBST and subsequently incubated with
diluted secondary antibody for 1 h at room temperature. Following
four washes, the blots were measured by using horseradish
peroxidase substrate (Thermo Fisher Scientific, Inc.) and captured
with a G:BOX Chemi Gel Documentation System (Syngene, Frederick,
MD, USA). The relative density was calculated with Quantity One
Software (v4.6.2, Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Statistical analysis
Student's t-test was performed to determine the
statistical significance between two groups by using SPSS version
19.0 software (IBM SPSS, Armonk, NY, USA). P<0.05 was considered
to indicate a statistically significant difference. Data are
presented as the mean ± standard deviation.
Results
DU145 cells are more sensitive to
tetrandrine (Tet) compared with PC-3 cells
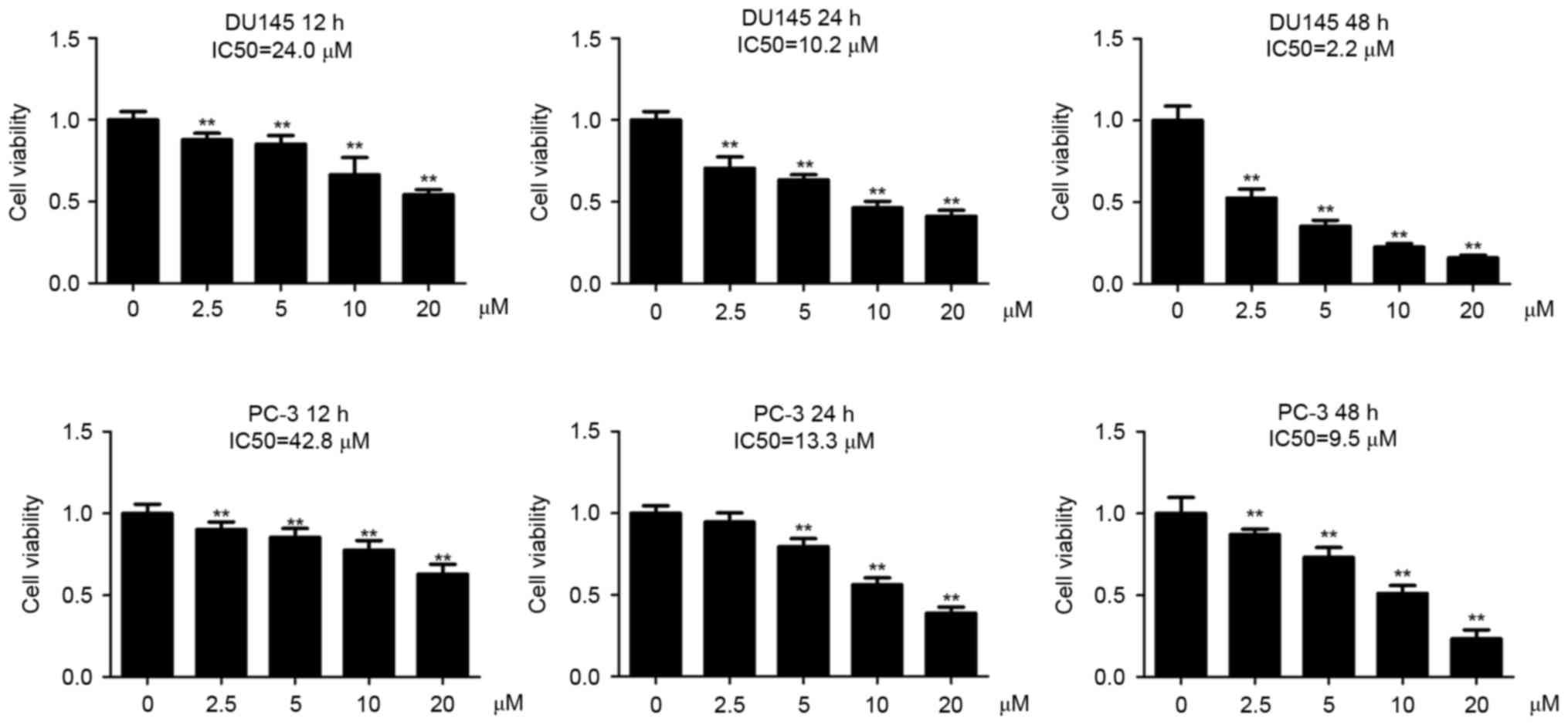
The present authors have previously observed that
DU145 cells are more sensitive to Tet treatment than PC-3 cells at
48 h post-incubation (1). In the
present study, the DU145 and PC-3 cells were incubated with a
series of Tet concentrations for 12, 24 and 48 h, respectively. It
was observed that Tet treatment inhibited cell viability in the two
cell lines in a dose- and time-dependent manner (Fig. 1). The half maximal inhibitory
concentration (IC50) values of Tet in DU145 cells were
24.0 (12 h), 10.2 (24 h) and 2.6 µM (48 h), whereas in PC-3 cells
the values were 42.8 (12 h), 13.3 (24 h) and 9.5 µM (48 h),
indicating that DU145 cells were more sensitive to Tet treatment
compared with PC-3 cells (Fig.
1).
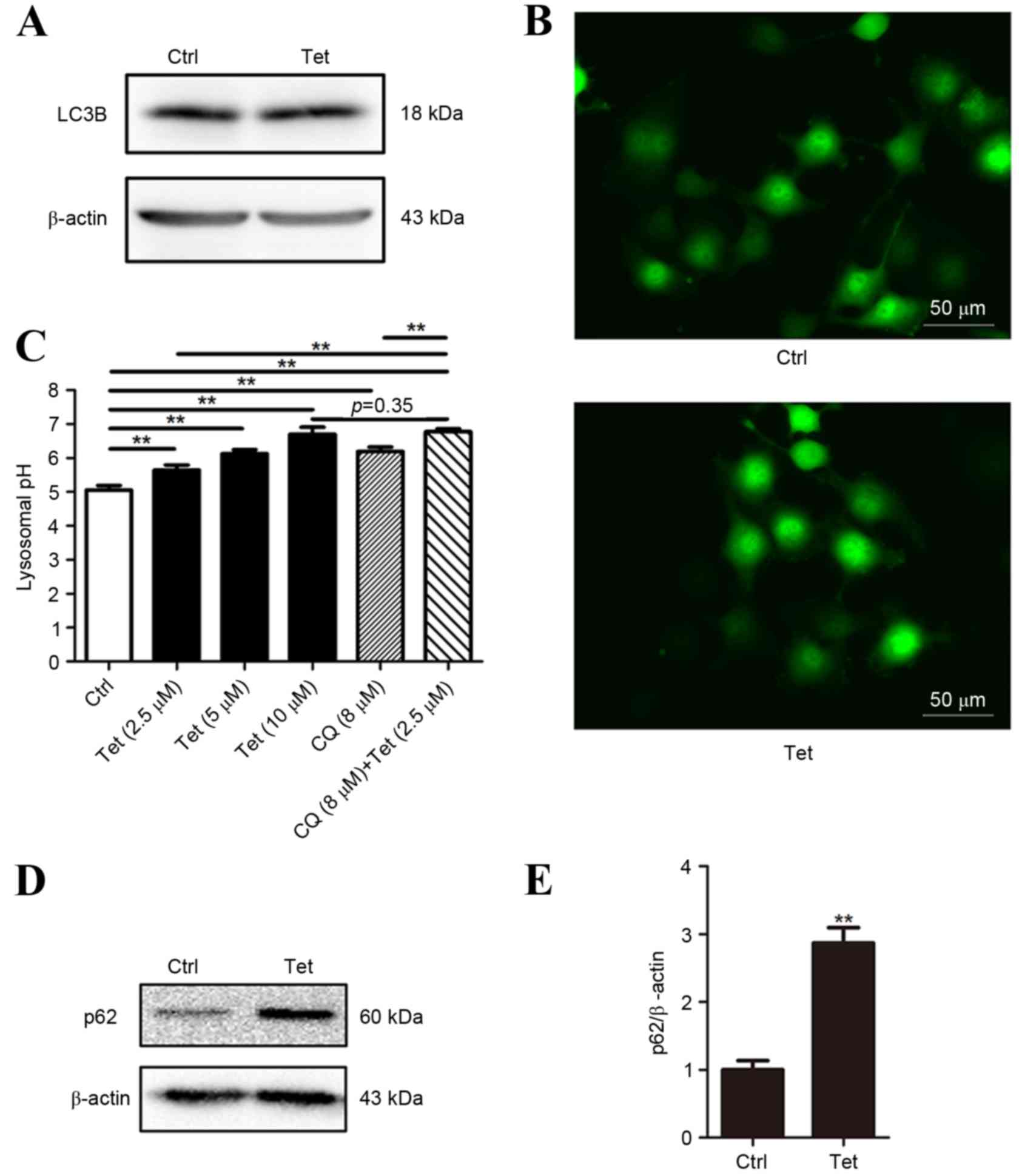
Tetrandrine fails to induce LC3
conversion in DU145 cells
Autophagy has an important role in Tet-induced cell
death (1,3). In order to explore the role of autophagy
in Tet-induced cell death of DU145 cells, the LC3 conversion was
measured using western blotting. The DU145 cells were treated with
10 µM Tet for 12 h. At 10 µM, Tet was able to induce a significant
increase in LC3-II and EGFP-LC3 puncta formation in PC-3 cells
(1). It was observed that in DU145
cells LC3-II expression was undetectable in baseline and Tet
treatment conditions (Fig. 2A). This
observation was not unexpected as the lack of one or two exons in
ATG5 interferes with the formation of LC3-II in DU145 cells
(12). Consistently, EGFP-LC3 puncta
was not be observed in Tet-treated DU145 cells (Fig. 2B).
As autophagy mainly depends on the function of the
lysosome, lysosomal pH of DU145 cells treated with a range of Tet
concentrations (2.5 to 10 µM) was measured. The pH value increased
in a concentration-dependent manner (Fig.
2C), indicating that Tet neutralized lysosomal acidity in DU145
cells.
De-acidulation of lysosomes results in the blockade
of autophagic flux. As LC3 lipidation in DU145 cells is defective,
assays based on LC3 turnover cannot be performed. Therefore, the
expression of p62 (a protein that is degraded through autophagy and
is upregulated when the autophagic flux is blocked) was analyzed.
p62 protein in Tet-treated DU145 cells was upregulated, indicating
that the autophagic flux may be blocked (Fig. 2D and E).
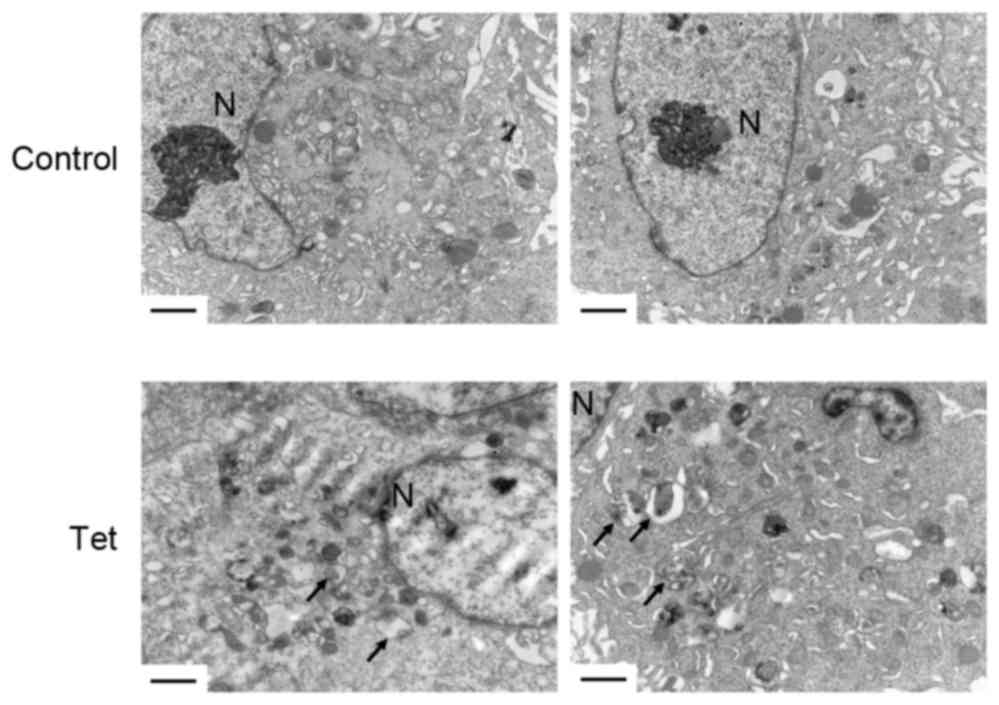
Tet triggers autophagosome
accumulation in DU145 cells
In order to investigate the role of autophagy in
DU145 cells, the ultrastructural changes of DU145 cells upon Tet
treatment for 12 h were observed. Notably, Tet triggered a large
amount of autophagosome accumulation in the cytoplasm (Fig. 3). There were no significant
differences in the shapes of the autophagosomes in DU145 cells
compared with autophagosomes in other cell lines induced by Tet
treatment, indicating that Tet may have triggered an alternative
autophagy in DU145 cells.
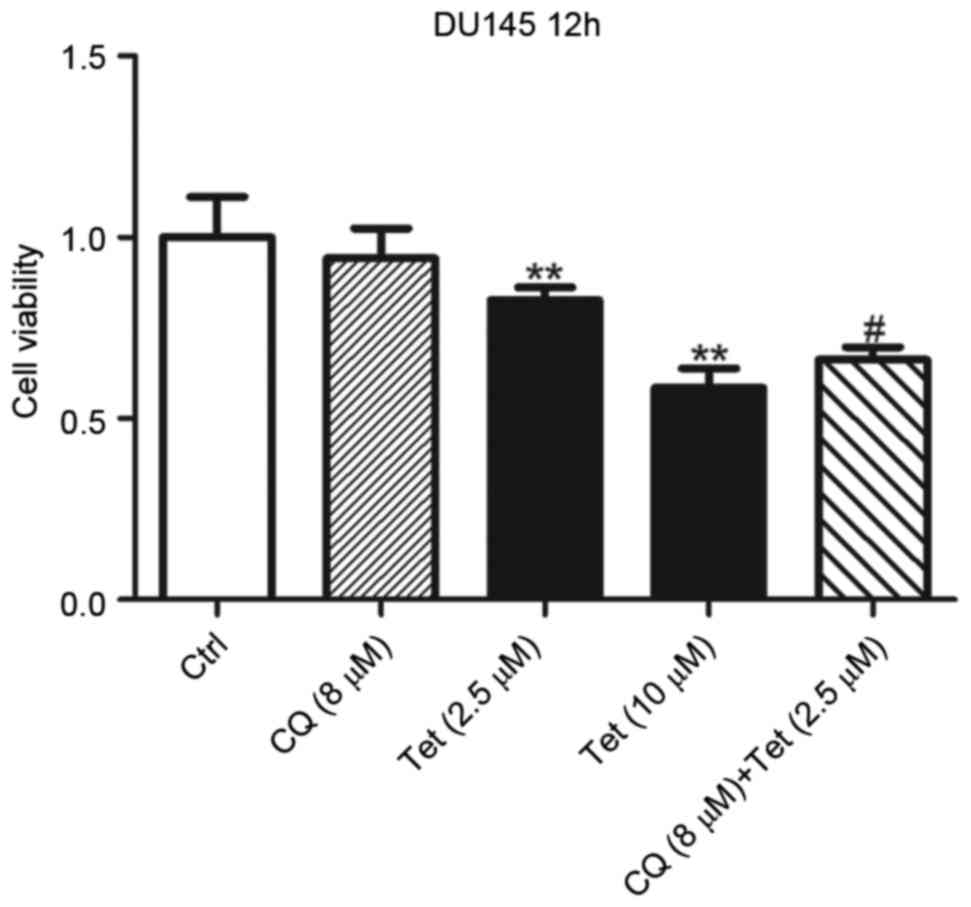
Lysosomal de-acidulation is required
in tetrandrine-induced cell death
The present authors have previously reported that
Tet is a potent lysosomal inhibitor in a number of cell lines,
which is closely associated with autophagy and its anti-tumor
effects (1). In the present study,
whether Tet-induced lysosomal de-acidulation contributed to its
anti-tumor effects in DU145 cells was investigated. To reduce
Tet-induced cell death, DU145 cells were treated with 2.5 µM Tet
for 12 h. At 2.5 µM, Tet exhibited mild inhibitory effects on DU145
cells (Fig. 1), and the lysosomal pH
was increased to 5.64. DU145 cells were subsequently co-treated
with 8 µM CQ to increase the lysosomal pH to 6.78 (Fig. 2C). Treatment with 8 µM CQ alone for 12
h did not result in a significant change in viability of DU145
cells (Fig. 4). However, the presence
of CQ synergized Tet-induced cell death (Fig. 4). These results indicated that high
lysosomal pH is required in Tet-induced cell death of DU145
cells.
Discussion
LC3 conversion from type I to type II has been
employed as a biomarker to evaluate the level of autophagy in
previous studies (15,16). In the present study, a notable finding
is that Tet triggered LC3-independent autophagy in ATG5 defective
prostate cancer cells. This alternative autophagy was characterized
as the accumulation of autophagosomes without affecting LC3
conversion. Therefore, evaluation of autophagy using LC3-based
assays should be cautiously interpreted. Tet induced lysosomal
de-acidulation in DU145 cells, which may result in autophagic flux
inhibition and may contribute to its anti-tumor effects.
It has been reported that serum starvation, CQ
treatment and rapamycin treatment all failed to induce LC3-I to
LC3-II conversion in DU145 cells (12). Due to the lack of functional products
of ATG5, the LC3-I to LC3-II conversion is blocked (12). Therefore, the EGFP-LC3 marked
autophagosomes could not be observed in DU145 cells (Fig. 2B). By contrast, a recent study
reported that sorafenib induces EGFP-LC3 foci formation in DU145
cells (17). The results of the
present study indicated that Tet could not induce EGFP-LC3 puncta
formation, but it increased the number of autophagosomes in DU145
cells. An alternative autophagy has been previously been discovered
in ATG5 genetic knockout mouse embryonic fibroblasts (18). The results of the present study
demonstrated that this type of autophagy may naturally exist in
cancer cells. The present authors postulate that this alternative
autophagy may universally exist in different types of cells.
Therefore, only assessing autophagy by using experiments based on
LC3 turnover may underestimate its actual level.
The present authors previously confirmed that Tet is
a known lysosome inhibitor and is able to block autophagic flux
(1). Like other cancer cell lines, it
was observed in the present study that the lysosomal acidity was
neutralized following Tet incubation in DU145 cells. Therefore, the
present authors hypothesize that the autophagic flux may be
blocked, although assays based on LC3 turnover or monomeric red
fluorescent protein-EGFP-LC3 co-localization could not be performed
in DU145 cells. The increased p62 levels induced by Tet supported
this hypothesis.
The role of autophagy in chemotherapy remains
unclear. It may either lead to cell death or maintain cell survival
against chemotherapeutic drugs. In DU145 cells, sorafenib blocks
the autophagic flux, whereas transiently re-expressing ATG5 in
DU145 cells antagonizes sorafenib-induced cell death (17). Therefore, the present authors
postulate that the autophagic flux may be relatively inadequate in
DU145 cells. Blockade of autophagic flux with Tet resulted in cell
death. This may explain why DU145 cells exhibited the highest
sensitivity towards Tet in all of the 11 cell lines tested
(1). It has been reported that CQ
enhances the anti-tumor effects in cancer cells (19). The results of the present study
indicated that lysosomal pH is able to affect Tet-induced cell
death. Further alkalizing lysosomes with nontoxic concentrations of
CQ enhanced Tet-induced cell death, indicating that low acidity of
the lysosomal lumen is associated with Tet-induced cell death.
Collectively, the present study provided evidence
that Tet induced alternative autophagy in DU145 cells. Detecting
autophagy in tumor tissue may assist in selecting lysosome
inhibitors for chemotherapy treatment in prostate cancer.
Acknowledgements
The authors would like to thank Dr Tamotsu Yoshimori
at Osaka University (Osaka, Japan) for providing the EGFP-LC3
plasmid. The present study was supported by grants from the China
Postdoctoral Science Foundation (No. 2016M601065).
References
|
1
|
Qiu W, Su M, Xie F, Ai J, Ren Y, Zhang J,
Guan R, He W, Gong Y and Guo Y: Tetrandrine blocks autophagic flux
and induces apoptosis via energetic impairment in cancer cells.
Cell Death Dis. 5:e11232014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ma JW, Zhang Y, Li R, Ye JC, Li HY, Zhang
YK, Ma ZL, Li JY, Zhong XY and Yang X: Tetrandrine suppresses human
glioma growth by inhibiting cell survival, proliferation and tumour
angiogenesis through attenuating STAT3 phosphorylation. Eur J
Pharmacol. 764:228–239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang H, Liu T, Li L, Wang Q, Yu C, Liu X
and Li W: Tetrandrine is a potent cell autophagy agonist via
activated intracellular reactive oxygen species. Cell Biosci.
5:42015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu FS, Yu CS, Chen JC, Yang JL, Lu HF,
Chang SJ, Lin MW and Chung JG: Tetrandrine induces apoptosis via
caspase-8, −9, and −3 and poly (ADP ribose) polymerase dependent
pathways and autophagy through beclin-1/LC3-I, II signaling
pathways in human oral cancer HSC-3 cells. Environ Toxicol.
31:395–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen JC, Hwang JH, Chiu WH and Chan YC:
Tetrandrine and caffeine modulated cell cycle and increased glioma
cell death via caspase-dependent and caspase-independent apoptosis
pathways. Nutr Cancer. 66:700–706. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen Y, Li P, Yang S, Tong N, Zhang J and
Zhao X: Tetrandrine enhances the anticancer effects of arsenic
trioxide in vitro. Int J Clin Pharmacol Ther. 52:416–424. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qin R, Shen H, Cao Y, Fang Y, Li H, Chen Q
and Xu W: Tetrandrine induces mitochondria-mediated apoptosis in
human gastric cancer BGC-823 cells. PLoS One. 8:e764862013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu SJ and Ng LT: Tetrandrine inhibits
proinflammatory cytokines, iNOS and COX-2 expression in human
monocytic cells. Biol Pharm Bull. 30:59–62. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gong K, Chen C, Zhan Y, Chen Y, Huang Z
and Li W: Autophagy-related gene 7 (ATG7) and reactive oxygen
species/extracellular signal-regulated kinase regulate
tetrandrine-induced autophagy in human hepatocellular carcinoma. J
Biol Chem. 287:35576–35588. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang AC, Lien JC, Lin MW, Yang JS, Wu PP,
Chang SJ and Lai TY: Tetrandrine induces cell death in SAS human
oral cancer cells through caspase activation-dependent apoptosis
and LC3-I and LC3-II activation-dependent autophagy. Int J Oncol.
43:485–494. 2013.PubMed/NCBI
|
|
11
|
Liu T, Men Q, Wu G, Yu C, Huang Z, Liu X
and Li W: Tetrandrine induces autophagy and differentiation by
activating ROS and Notch1 signaling in leukemia cells. Oncotarget.
6:7992–8006. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ouyang DY, Xu LH, He XH, Zhang YT, Zeng
LH, Cai JY and Ren S: Autophagy is differentially induced in
prostate cancer LNCaP, DU145 and PC-3 cells via distinct splicing
profiles of ATG5. Autophagy. 9:20–32. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kimura S, Noda T and Yoshimori T:
Dissection of the autophagosome maturation process by a novel
reporter protein, tandem fluorescent-tagged LC3. Autophagy.
3:452–460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klionsky DJ, Abdalla FC, Abeliovich H,
Abraham RT, Acevedo-Arozena A, Adeli K, Agholme L, Agnello M,
Agostinis P, Aguirre-Ghiso JA, et al: Guidelines for the use and
interpretation of assays for monitoring autophagy. Autophagy.
8:445–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Klionsky DJ, Abeliovich H, Agostinis P,
Agrawal DK, Aliev G, Askew DS, Baba M, Baehrecke EH, Bahr BA,
Ballabio A, et al: Guidelines for the use and interpretation of
assays for monitoring autophagy in higher eukaryotes. Autophagy.
4:151–175. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kharaziha P, Chioureas D, Baltatzis G,
Fonseca P, Rodriguez P, Gogvadze V, Lennartsson L, Björklund AC,
Zhivotovsky B, Grandér D, et al: Sorafenib-induced defective
autophagy promotes cell death by necroptosis. Oncotarget.
6:37066–37082. 2015.PubMed/NCBI
|
|
18
|
Nishida Y, Arakawa S, Fujitani K,
Yamaguchi H, Mizuta T, Kanaseki T, Komatsu M, Otsu K, Tsujimoto Y
and Shimizu S: Discovery of Atg5/Atg7-independent alternative
macroautophagy. Nature. 461:654–658. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mei L, Chen Y, Wang Z, Wang J, Wan J, Yu
C, Liu X and Li W: Synergistic anti-tumour effects of tetrandrine
and chloroquine combination therapy in human cancer: A potential
antagonistic role for p21. Br J Pharmacol. 172:2232–2245. 2015.
View Article : Google Scholar : PubMed/NCBI
|