Introduction
Esophageal cancer, which includes esophageal
squamous cell carcinoma (ESCC) and adenocarcinoma of the esophagus
(EADC), is one of the most prevalent types of malignant tumor
worldwide (1). Esophageal cancer
occupies the sixth position among malignant tumors worldwide with
regard to mortality and ranks fourth in China (2). ESCC is the most predominant esophageal
cancer type in Asia (3). Surgery,
radiotherapy and chemotherapy are the typical treatments for ESCC.
Due to a lack of effective clinical diagnosis methods for ESCC, it
is often diagnosed in the later stages of the disease. The 5-year
overall survival rate of ESCC is between 20 and 30% (4). Investigating the mechanism of ESCC
progression is essential to improve disease diagnosis, treatment
and prognosis.
MicroRNAs (miRNAs) are a large class of highly
conserved small non-coding RNAs that negatively regulate gene
expression through binding the 3′ untranslated region (3′-UTR) of
target mRNAs, causing mRNA degradation and therefore repression of
translation (5). In total >1,000
miRNAs have been identified in the human genome, which may regulate
the expression of <30% of all protein-coding genes (6). Numerous studies have identified that
aberrant expression of miRNAs is associated with tumorigenesis,
tumor progression and response to therapy (7). miR-375 was initially characterized as a
pancreatic islet-specific miRNA, and it regulates glucose
homeostasis and glucose-induced insulin secretion (8,9). Further
miRNA expression profiling studies revealed that miR-375 is
downregulated in multiple types of cancer and acts as a tumor
suppressor. For example, miR-375 was demonstrated to repress
gastric cancer growth by targeting Janus kinase 2 (10). It has also been reported that miR-375
inhibits the proliferation and invasion of human squamous cervical
cancer cells by repressing transcription factor SP1 (11). In ESCC, miR-375 is one of the most
frequently downregulated miRNAs (10). However, the tumor suppression
mechanism of miR-375 in ESCC has not been fully elucidated.
In the present study, it was identified that miR-375
is downregulated in ESCC tumor tissue and the ESCC cell line EC109.
It was also identified that miR-375 acts as a tumor suppressor in
EC109 cells, and that metadherin (MTDH) is a direct target. The
effect of miR-375 on ESCC cell proliferation and invasive ability
may be mediated by the downregulation of MTDH, suggesting that MTDH
is a potential therapeutic target for ESCC therapy.
Materials and methods
Patients and sample collection
A total of 10 patients who had been diagnosed with
ESCC were recruited to participate in the present study between May
2013 and October 2013 at Xiangya Hospital of Central South
University (Changsha, China). All patients underwent esophageal
cancer resection prior to administration of chemotherapy; following
resection, ESCC and corresponding non-cancerous tissues were
collected immediately and validated by pathologists. All patients
provided written informed consent. All experiments were performed
with the approval of the Ethics Committee of the Xiangya Hospital
of Central South University.
Cell lines and cell culture
The ESCC cell line EC109 and the human immortalized
normal esophageal epithelial cell line Het-1A were purchased from
Changsha Yingrun Biotechnology Co. Ltd. (Changsha, China). Cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and supplemented
with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin in a
humidified incubator with 5% CO2 at 37°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total miRNA was extracted using an miRNeasy kit
(Qiagen GmbH, Hilden, Germany), according to the manufacturer's
protocol. RNA was converted into cDNA using an miScriptRT kit
(Qiagen GmbH), according to the manufacturer's protocol. qPCR was
performed using an miScript SYBR Green PCR kit (Qiagen GmbH) in an
ABI 7,500 Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). PCR reactions were run at 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. The
detected expression of miRNAs was normalized using small nuclear
RNA U6 (U6) as an endogenous control. miR-375 and U6 primers
(miR-375 forward, 5′-gcagatttgttc gttcggctc-3′ and reverse,
5′-gtgcagggtccgaggt-3′; and U6 forward, 5′-ctcgcttcggcagcaca-3′ and
reverse 5′-aacgcttca cgaatttgcgt-3′) were purchased from Changsha
Yingrun Biotechnology Co. Ltd. The RT-qPCR assays were performed in
triplicate and the change in expression level was calculated using
the 2−ΔΔCq method (ABI 7500 Software v2.0.1; Applied
Biosystems; Thermo Fisher Scientific, Inc.) (12).
Proliferation assays
A Cell Counting kit-8 (CCK-8; Beyotime Institute of
Biotechnology, Haimen, China) assay was used to measure cell
proliferation. A total of 2×104 cells/well were plated
in 96-well plates and cultured under the aforementioned conditions.
Cell proliferation was monitored each day. To allow cell counting,
10 µl CCK-8 reagent was added to each well and the plates were
incubated at 37°C for 2 h. The absorbance values were subsequently
measured at 450 nm using a microplate reader (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). Each experiment was performed in
triplicate.
Transfection of miR-375 mimic
The miR-375 mimic (sense,
5′-uuuguucguucggcucgcguga-3′ and antisense, 5′-acgcgagcc
gaacgaacaaauu-3′) and a negative control (NC; sense, 5′-uuc
uccgaacgugucacgutt-3′ and antisense, 5′-acgugacacguucgg aga att-3′)
were purchased from GenePharm (Shanghai, China). Cells were plated
at 3×105 per well in 6-well plates and cultured for 24
h. The cells were subsequently transfected with miR-375 mimic or NC
RNA, to a final concentration of 75 nM, using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) and FBS-free Opti-MEM
medium (Gibco; Thermo Fisher Scientific, Inc.). After 6 h, the
medium was replaced with DMEM containing 10% FBS.
Flow cytometric analysis of
apoptosis
Apoptotic cells were measured using an Annexin
V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit
(Nanjing KeyGen Biotech Co., Ltd, Nanjing, China). The cell samples
were harvested using 0.25% EDTA-free trypsin after 48 h of
incubation, washed twice with ice-cold PBS and resuspended in 500
µl binding buffer. Cells were incubated with 5 µl annexin
V-FITC-labeled specific antibodies (from the apoptosis detection
kit) and 5 µl propidium iodide (PI) for between 15 and 20 min in
the dark, and detected using a FACSAria flow cytometer and FASCDiva
4.1 software (BD Biosciences, Franklin Lakes, NJ, USA). Each
experiment was performed in triplicate.
Cell cycle assays
Following transfection with miR-375 mimic or NC, the
cell samples were harvested as aforementioned and fixed with 75%
ethanol at 4°C overnight. The cells were washed in ice-cold PBS and
treated with RNase I and Triton-X-100, followed by PI staining for
30 min in the dark. Finally, cells were washed and resuspended in
PBS. Cell cycle distributions were detected using a BD Biosciences
FACSAria flow cytometer.
Invasion assays
The invasiveness of cells was determined using a
Transwell system [24-well plate, 8 µm pore polyethylene
terephthalate (PET) membrane; Corning Incorporated, Corning, NY,
USA]. The PET membrane was coated with Matrigel (dilution, 1:5; BD
Biosciences) in FBS-free DMEM. Following transfection, the cell
samples were harvested as aforementioned and resuspended in
FBS-free DMEM, for seeding in the upper chamber. DMEM containing
10% FBS was added to the lower chamber. Following incubation at
37°C for 24 h, cells that appeared on the lower surface of the
membrane were fixed with methanol for 15 min at 4°C, then stained
with 0.1% crystal violet solution and counted under a light
microscope.
Construction of vector and luciferase
reporter assay
The 3′-UTR of the MTDH gene (positions 1454–1461)
was predicted to be complementary to the sequence of miR-375
according to an analysis using the miRNA target gene prediction
database TargetScan 7.1 (13).
Fragments of sequence from wild-type and mutant MTDH 3′-UTR were
amplified by PCR and cloned into a pYr-MirTarget luciferase vector
(Changsha Yingrun Biotechnology Co. Ltd.), designated pYr-MTDH-3U
and PYr-MTDH-3Umt, respectively. The primers for the wild-type
3′-UTR, containing XhoI and NotI restriction sites, were as
follows: Forward, 5′-ccgctcgagtgtggcgccaagaacgaa-3′, and reverse,
5′-aaagcggccgcatactgccatgttcctgaa-3′. The primers for the mutant
3′-UTR were as follows: Forward, 5′-ttgtcagac
taggaaagctaaaaaaatggttttgctgaa-3′; reverse, 5′-ttcagcaaaactaaa
accatttttttagctttcctagtctgacaa-3′. HEK293 cells (American Type
Culture Collection, Manassas, VA, USA) were plated into a 96-well
plate and co-transfected with 100 ng pYr-MTDH-3U or pYr-MTDH-3Umt
and miR-375 mimic or NC (75 nM) using Lipofectamine 2000.
Luciferase activity was determined using a Dual-Luciferase Reporter
Assay kit (Promega Corporation, Madison, WI, USA), according to the
manufacturer's protocol. Each experiment was performed in
triplicate.
Western blot analysis
Cells were harvested and homogenized with lysis
buffer (Beyotime Institute of Biotechnology) 48 h
post-transfection. Protein concentrations were measured using a BCA
Protein Assay kit (Beyotime Institute of Biotechnology). Proteins
(between 30 and 50 µg) were separated by SDS-PAGE (10% gel) and
transferred onto nitrocellulose membranes (EMD Millipore,
Billerica, MA, USA). Subsequent to blocking with 5% skimmed milk
powder at room temperature for 2 h, the membranes were incubated
with primary antibodies specific to MTDH (catalog no., 14065S),
vascular endothelial growth factor C (VEGF-C; catalog no., 2445S),
cyclin D1 (catalog no., 2978T), epithelial (E-) cadherin (catalog
no., 3195S) and β-actin (catalog no., 3700S) (all dilution,
1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA). The
membranes were then incubated with corresponding horseradish
peroxidase-conjugated secondary antibodies (goat anti-mouse
secondary antibody for β-actin; dilution, 1:5,000; catalog no.,
sc-2005 or mouse anti-rabbit secondary antibody for MTDH, VEGF-C,
cyclin D1 and E-cadherin; catalog no., sc-2357; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The resulting immunoblots
were quantified using ImageJ2x software (National Institutes of
Health, Bethesda, MD, USA).
Statistical analysis
Data were expressed as the mean ± standard deviation
from at least three independent experiments and statistically
analyzed using a one-way analysis of variance test followed by
Tukey's multiple comparison test or an independent sample t-test.
All statistics were performed using SPSS software (version 17.0;
SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to indicate
a statistically significant difference, and P<0.01 was
considered to indicate a highly significant difference.
Results
miR-375 is downregulated in clinical
esophageal cancer tissues and the ESCC cell line EC109
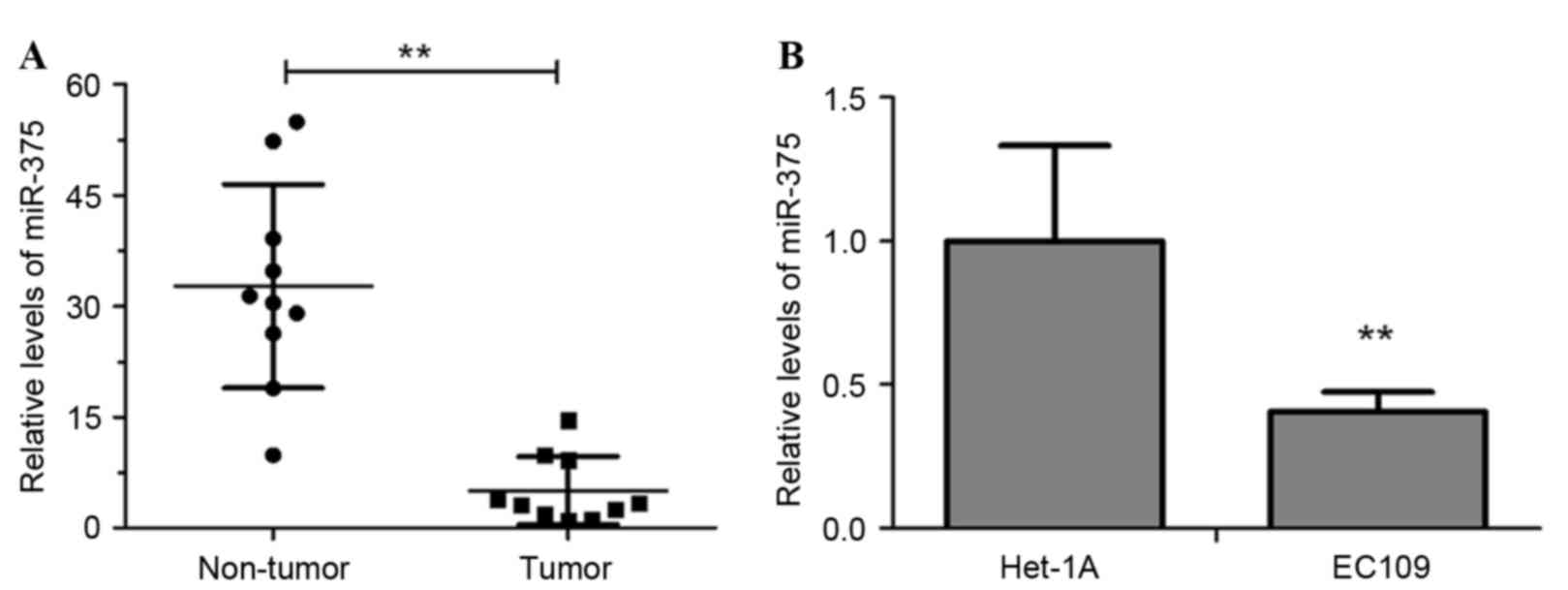
The level of miR-375 in clinical esophageal
carcinoma and adjacent non-tumor tissues from 10 ESCC patients were
measured using RT-qPCR. The results revealed that the miR-375 level
was significantly decreased in tumor tissues compared with in
non-tumor tissues (Fig. 1A;
P<0.01). Additionally, miR-375 expression in the ESCC cell line
EC109 was significantly downregulated compared with that in Het-1A,
a normal human esophageal endothelial cell line (Fig. 1B, P<0.01). These results suggested
that downregulation of miR-375 may be associated with the
pathogenesis of esophageal cancer.
miR-375 suppresses cell growth and
induces cell cycle arrest in EC109 cells
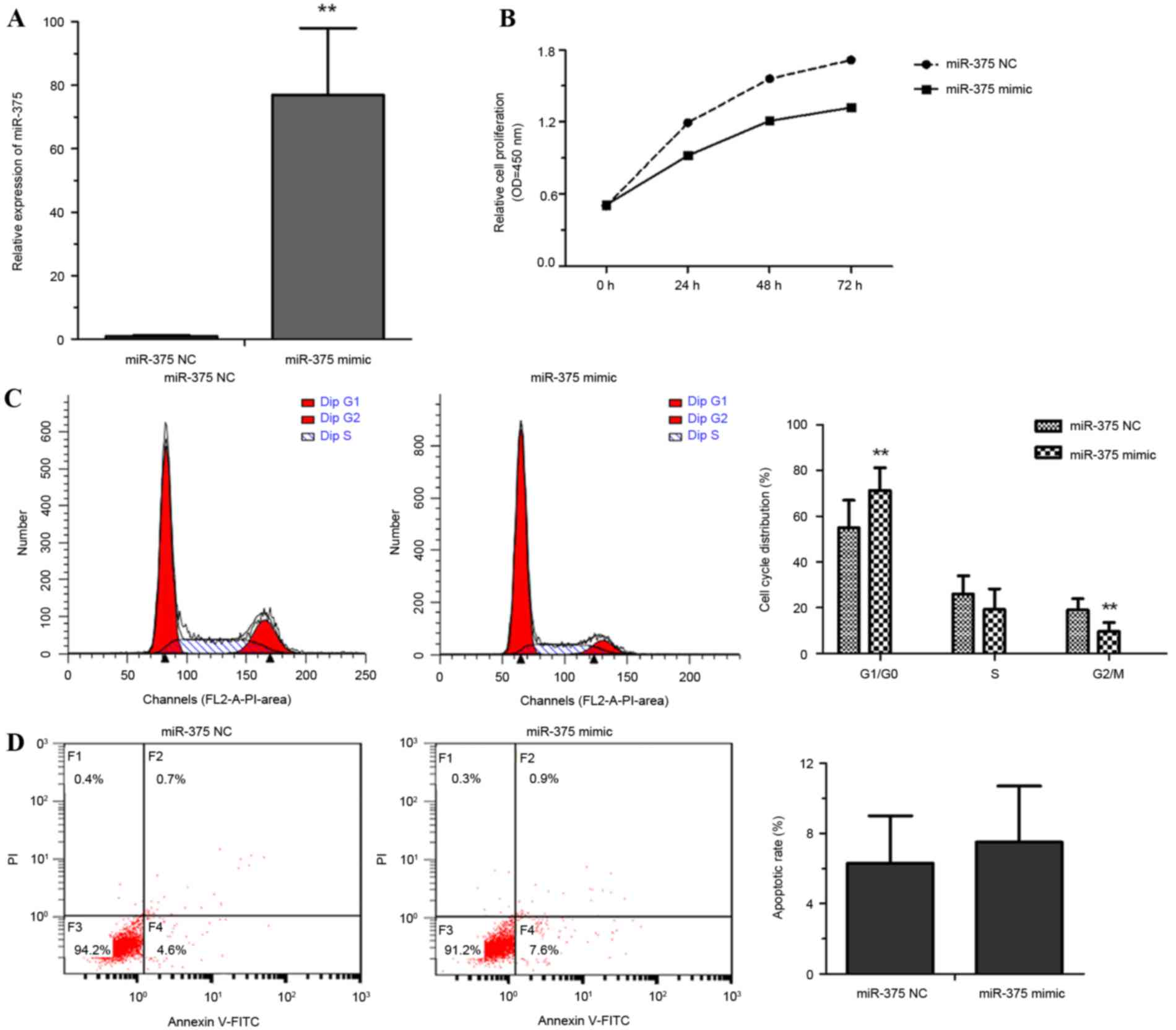
To explore the roles of miR-375 in esophageal cancer
progression, EC109 cells were transfected with miR-375 mimic or a
NC. Cell proliferation rate, the proportion of apoptotic cells and
cell cycle distribution were measured. Overexpression of miR-375 by
miR-375 mimic transfection was confirmed using RT-qPCR (Fig. 2A). miR-375 overexpression
significantly inhibited the proliferation of EC109 cells (Fig. 2B). Flow cytometric analysis revealed
that overexpression of miR-375 modulated the cell cycle by inducing
G1 arrest (Fig. 2C). No significant
difference between the NC and miR-375 mimic group in the rate of
apoptosis was identified (Fig. 2D).
These results suggested that the overexpression of miR-375
inhibited cell proliferation through triggering cell cycle arrest
rather than apoptosis.
miR-375 suppresses invasion in EC109
cells
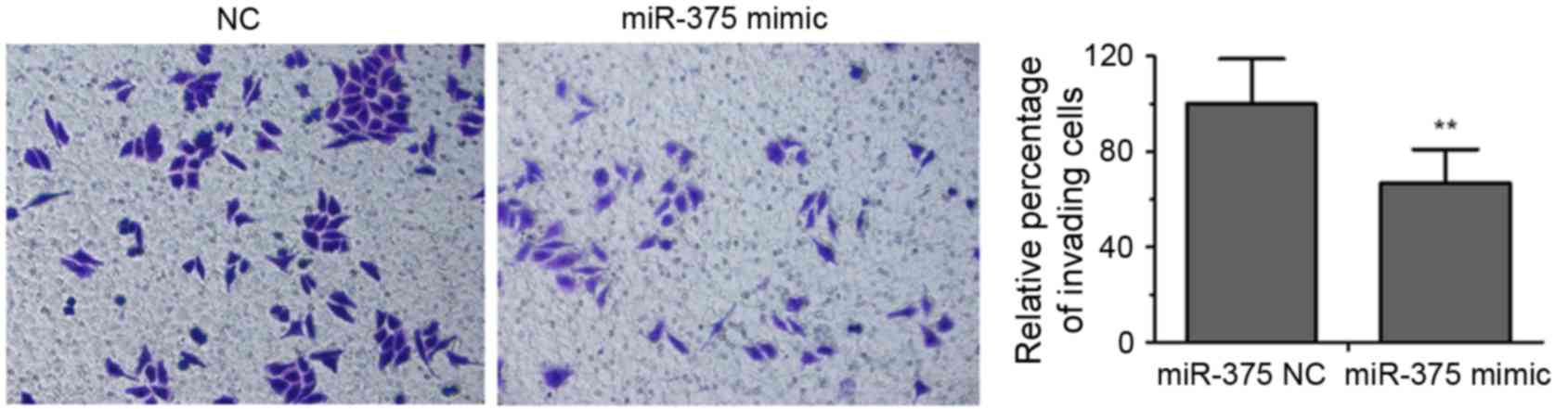
EC109 cells transfected with miR-375 mimic or an NC
were analyzed using Transwell invasion assays. As presented in
Fig. 3, overexpression of miR-375
repressed the invasive ability of EC109 cells by ~67% compared with
the NC group (P<0.01).
MTDH is a direct target gene of
miR-375
To further explore the molecular mechanism of
miR-375 in ESCC progression, computational algorithms were applied
(using TargetScan) to predict potential target mRNAs of miR-375. It
was identified that the MTDH mRNA 3′-UTR contained highly conserved
binding sites for miR-375 (Fig. 4A).
Dual-luciferase assays were performed to confirm this prediction.
HEK293 cells were co-transfected with pYr-MTDH-3U and miR-375
mimic. The assessment of luciferase activity demonstrated that
miR-375 mimic significantly decreased the luciferase activity of
pYr-MTDH-3U compared with the NC (Fig.
4B; P<0.01). No significant difference in the luciferase
activity of pYr-MTDH-3Umt was identified (Fig. 4C). miR-375 mimic downregulated the
expression of MTDH in EC109 cells (P<0.01), further confirming
that MTDH is a target gene of miR-375. The expression of the MTDH
downstream protein VEGF-C (P<0.01) and cell cycle regulator
cyclin D1 (P<0.05) were decreased by miR-375 mimic (Fig. 4D). By contrast, miR-375 mimic
significantly upregulated expression of the epithelial marker
E-cadherin (Fig. 4D; P<0.01),
which may inhibit tumor cell epithelial-mesenchymal transition
(EMT) and invasion (14).
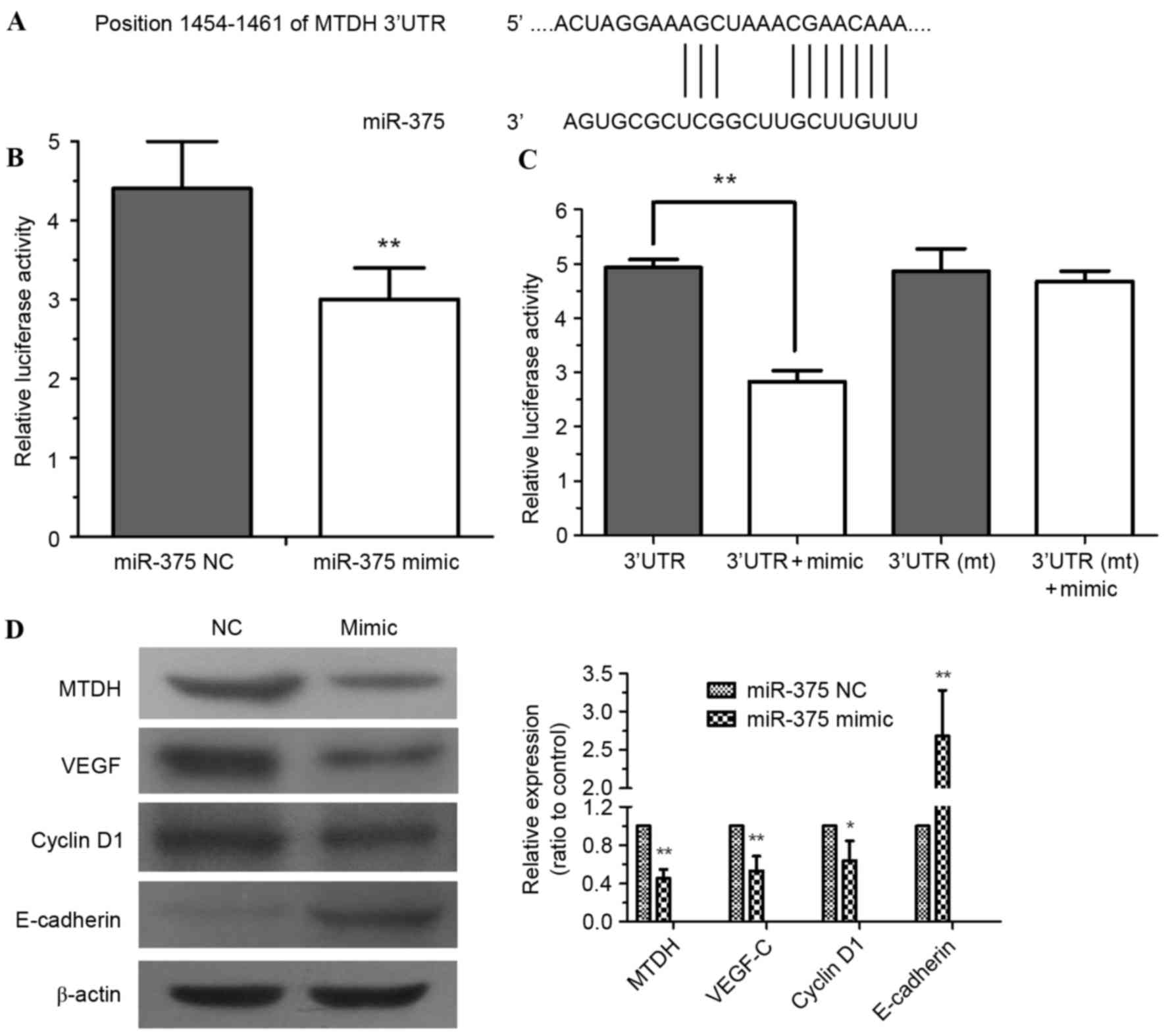 | Figure 4.MTDH is a direct target gene of
miR-375. (A) The 3′-UTR of MTDH mRNA includes a highly conserved
binding site for miR-375. (B) HEK293 cells were co-transfected with
pYr-MTDH-3U and an miR-375 mimic or NC. At 24 h after transfection,
luciferase activity was detected. (C) Dual-luciferase assays
demonstrated miR-375 mimic-mediated repression of pYr-MTDH-3U
luciferase; however, no significant difference from pYr-MTDH-3Umt
luciferase was identified. (D) miR-375 modulated the expression of
MTDH, cyclin D1, VEGF-C and E-cadherin in EC109 cells. β-Actin
served as a loading control. Experiments were performed in
triplicate and data are expressed as the mean ± standard deviation,
**P<0.01 vs. NC. MTDH, metadherin; miR-375, microRNA-375;
3′-UTR, 3′-untranslated region; NC, negative control; mt, mutated;
VEGF-C, vascular endothelial growth factor. |
Discussion
ESCC has a poor prognosis and high mortality rate
(2). Cancer metastasis is the primary
cause of cancer-associated mortality; therefore, preventing
metastasis may be an effective strategy for cancer therapy. A
number of studies have provided novel insights into the underlying
molecular mechanism of regulation of gene expression by miRNAs. It
has been reported that the aberrant expression of miRNA is
associated with tumorigenesis, tumor progression, cancer prognoses
and response to therapy (7,15,16). In
the present study, it was demonstrated that miR-375 was
downregulated in ESCC tumor tissue samples and the ESCC cell line
EC109 compared with non-tumor tissues and Het-1A, a normal
esophageal endothelial cell line, respectively. Furthermore,
overexpression of miR-375 in EC109 cells was demonstrated to
inhibit cell proliferation and invasion, and induce cell cycle
arrest. MTDH was identified as a target gene of miR-375; the
results of the present study indicate that miR-375 acts as a tumor
suppressor in EC109 cells by modulating the expression of MTDH.
Previous studies have identified several miRNAs
suitable for use in distinguishing ESCC from non-tumor tissue. Guo
et al (4) reported four miRNAs
(miR-100, −99a, −29c and −140) that were downregulated and three
miRNAs (miR-25, −424 and −151) that were upregulated in ESCC. Yan
et al (17) confirmed that
miR-340 is significantly downregulated in ESCC and inhibits ESCC
cell growth and invasion by targeting phosphoserine
aminotransferase 1. miR-204 was demonstrated to repress EC109 EMT
and invasion by targeting box protein M1 (18). In addition, the circulating levels of
miR-21 and miR-375 in patients with ESCC may act as reliable
prognostic markers for ESCC (19).
It has been demonstrated that miR-375 predominantly
serves as a tumor suppressor, particularly in tumors of the
digestive tract (10). In
hepatocellular carcinoma, ectopic expression of miR-375 decreases
cell growth and invasion, and induces G1 arrest and apoptosis
(20). miR-375 has been reported as
one of the most frequently downregulated miRNAs in esophageal
cancer (21). Similarly, in the
present study, it was demonstrated that miR-375 was significantly
decreased in ESCC tumor tissue and the ESCC line EC109.
Overexpression of miR-375 in EC109 repressed cell growth and
invasion; however, it did not induce apoptosis in EC109 cells.
miR-375-induced G1 cell cycle arrest was the primary cause for the
inhibition of EC109 cell proliferation.
Although several studies have explored the role of
miRNAs in esophageal cancer, the identified potential targets
require further confirmation. For example, overexpression of
miR-145, −133a or −133b in ESCC may inhibit cell proliferation and
invasion; they all may repress the expression of Fascin homolog 1
(22). miR-375 inhibits ESCC cell
proliferation through targeting insulin-like growth factor receptor
1 and 3-phosphoinositide-dependent protein kinase-1, which are
involved in the potentially carcinogenic phosphoinositide 3-kinase
(PI3K)/protein kinase B (AKT) signaling pathway (10). To investigate the mechanism of miR-375
function in ESCC cells, computational algorithms were used to
predict possible target mRNAs of miR-375. The intersection of
algorithms and a luciferase reporter assay indicated that MTDH was
a target gene of miR-375. MTDH was originally identified as an
oncogene (23) upregulated in
numerous types of malignancy, including breast cancer,
neuroblastoma, non-small cell lung cancer and cancers of the
digestive tract (24–26). A number of studies have demonstrated
that the upregulation of MTDH promotes tumor progression through
modulating signaling pathways including the PI3K/AKT, Wnt/β-catenin
and nuclear factor κB signaling pathways (24). In the present study, overexpression of
miR-375 in EC109 cell significantly decreased the expression of
MTDH.
To further investigate the mechanism of miR-375 on
tumor growth and invasiveness, the expression levels of VEGF-C,
E-cadherin and cyclin D1 were examined. The results demonstrated
that cyclin D1 and VEGF-C were downregulated, whereas E-cadherin
was upregulated in miR-375-overexpressing EC109 cells. The majority
of tumors and metastases begin as avascular masses limited in size
(27), therefore VEGF-C, an inducer
of angiogenesis, is a critical factor for tumor growth and
development (28). It has been
confirmed that MTDH can directly upregulate VEGF-C expression by
enhancing VEGF-C promoter activity and activating the PI3K/AKT
signaling pathway (29,30). Cyclin D1 is essential for G1 phase
progression and is frequently overexpressed in a broad range of
human cancers (31). The
overexpression of MTDH was observed in chronic lymphocytic leukemia
and was identified to be associated with the Rai stage
classification. Inhibition of MTDH expression resulted in decreased
levels of cyclin D1 via suppression of the Wnt/β-catenin signaling
pathway (32). The epithelial marker
E-cadherin serves a key role in EMT; its expression represses tumor
invasion and metastasis (33). Li
et al (34) have reported that
overexpression of MTDH could promote EMT via downregulating
E-cadherin and increasing other EMT indicators. This demonstrates
the involvement of MTDH in tumorigenesis and supports the results
of the present study, suggesting that downregulation of miR-375 may
contribute to ESCC tumorigenesis via the resulting upregulation of
MTDH.
In summary, the present study has revealed that
miR-375 is underexpressed in ESCC tumor tissue samples and the ESCC
cell line EC109, compared with normal tissue and cells. miR-375 has
been demonstrated to act as a tumor suppressor in ESCC cells that
inhibited cell proliferation and invasion by repressing the
expression of its direct target MTDH, an oncogene associated with
tumorigenesis in numerous types of cancer. Therefore, the
miR-375/MTDH axis may represent a potential target for the
treatment of ESCC.
References
|
1
|
Kamangar F, Qiao YL, Schiller JT, Dawsey
SM, Fears T, Sun XD, Abnet CC, Zhao P, Taylor PR and Mark SD: Human
papillomavirus serology and the risk of esophageal and gastric
cancers: Results from a cohort in a high-risk region in China. Int
J Cancer. 119:579–584. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arnold M, Soerjomataram I, Ferlay J and
Forman D: Global incidence of oesophageal cancer by histological
subtype in 2012. Gut. 64:381–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lagergren J: Oesophageal cancer in 2014:
Advances in curatively intended treatment. Nat Rev Gastroenterol
Hepatol. 12:74–75. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo Y, Chen Z, Zhang L, Zhou F, Shi S,
Feng X, Li B, Meng X, Ma X, Luo M, et al: Distinctive microRNA
profiles relating to patient survival in esophageal squamous cell
carcinoma. Cancer Res. 68:26–33. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen K and Rajewsky N: The evolution of
gene regulation by transcription factors and microRNAs. Nat Rev
Genet. 8:93–103. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fendler A, Jung M, Stephan C, Honey RJ,
Stewart RJ, Pace KT, Erbersdobler A, Samaan S, Jung K and Yousef
GM: miRNAs can predict prostate cancer biochemical relapse and are
involved in tumor progression. Int J Oncol. 39:1183–1192.
2011.PubMed/NCBI
|
|
8
|
Poy MN, Eliasson L, Krutzfeldt J, Kuwajima
S, Ma X, Macdonald PE, Pfeffer S, Tuschl T, Rajewsky N, Rorsman P
and Stoffel M: A pancreatic islet-specific microRNA regulates
insulin secretion. Nature. 432:226–230. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van de Bunt M, Gaulton KJ, Parts L, Moran
I, Johnson PR, Lindgren CM, Ferrer J, Gloyn AL and McCarthy MI: The
miRNA profile of human pancreatic islets and beta-cells and
relationship to type 2 diabetes pathogenesis. PLoS One.
8:e552722013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yan JW, Lin JS and He XX: The emerging
role of miR-375 in cancer. Int J Cancer. 135:1011–1018. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang F, Li Y, Zhou J, Xu J, Peng C, Ye F,
Shen Y, Lu W, Wan X and Xie X: miR-375 is down-regulated in
squamous cervical cancer and inhibits cell migration and invasion
via targeting transcription factor SP1. Am J Pathol. 179:2580–2588.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:2015. View Article : Google Scholar
|
|
14
|
Li D, Li X, Cao W, Qi Y and Yang X:
Antagonism of microRNA-99a promotes cell invasion and
down-regulates E-cadherin expression in pancreatic cancer cells by
regulating mammalian target of rapamycin. Acta Histochem.
116:723–729. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bartels CL and Tsongalis GJ: MicroRNAs:
Novel biomarkers for human cancer. Clin Chem. 55:623–631. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Komatsu S, Ichikawa D, Takeshita H,
Tsujiura M, Morimura R, Nagata H, Kosuga T, Iitaka D, Konishi H,
Shiozaki A, et al: Circulating microRNAs in plasma of patients with
oesophageal squamous cell carcinoma. Br J Cancer. 105:104–111.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan S, Jiang H, Fang S, Yin F, Wang Z, Jia
Y, Sun X, Wu S, Jiang T and Mao A: MicroRNA-340 inhibits esophageal
cancer cell growth and invasion by targeting phosphoserine
aminotransferase 1. Cell Physiol Biochem. 37:375–386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun Y, Yu X and Bai Q: miR-204 inhibits
invasion and epithelial-mesenchymal transition by targeting FOXM1
in esophageal cancer. Int J Clin Exp Pathol. 8:12775–12783.
2015.PubMed/NCBI
|
|
19
|
Komatsu S, Ichikawa D, Takeshita H,
Konishi H, Nagata H, Hirajima S, Kawaguchi T, Arita T, Shiozaki A,
Fujiwara H, et al: Prognostic impact of circulating miR-21 and
miR-375 in plasma of patients with esophageal squamous cell
carcinoma. Expert Opin Biol Ther. 12 Suppl 1:S53–S59. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He XX, Chang Y, Meng FY, Wang MY, Xie QH,
Tang F, Li PY, Song YH and Lin JS: MicroRNA-375 targets AEG-1 in
hepatocellular carcinoma and suppresses liver cancer cell growth in
vitro and in vivo. Oncogene. 31:3357–3369. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mathé EA, Nguyen GH, Bowman ED, Zhao Y,
Budhu A, Schetter AJ, Braun R, Reimers M, Kumamoto K, Hughes D, et
al: MicroRNA expression in squamous cell carcinoma and
adenocarcinoma of the esophagus: Associations with survival. Clin
Cancer Res. 15:6192–6200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kano M, Seki N, Kikkawa N, Fujimura L,
Hoshino I, Akutsu Y, Chiyomaru T, Enokida H, Nakagawa M and
Matsubara H: miR-145, miR-133a and miR-133b: Tumor-suppressive
miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int J
Cancer. 127:2804–2814. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kang DC, Su ZZ, Sarkar D, Emdad L, Volsky
DJ and Fisher PB: Cloning and characterization of HIV-1-inducible
astrocyte elevated gene-1, AEG-1. Gene. 353:8–15. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brown DM and Ruoslahti E: Metadherin, a
cell surface protein in breast tumors that mediates lung
metastasis. Cancer Cell. 5:365–374. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li WF, Ou Q, Dai H and Liu CA:
Lentiviral-mediated short hairpin RNA knockdown of MTDH inhibits
cell growth and induces apoptosis by regulating the PTEN/AKT
pathway in hepatocellular carcinoma. Int J Mol Sci. 16:19419–19432.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang C, Zheng S, Liu Q, Liu T, Lu M, Dai
F, Gao X, Sheyhidin I and Lu X: Metadherin is required for the
proliferation, migration, and invasion of esophageal squamous cell
carcinoma and its meta-analysis. Transl Res. 166:614–626.e2. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Holash J, Maisonpierre PC, Compton D,
Boland P, Alexander CR, Zagzag D, Yancopoulos GD and Wiegand SJ:
Vessel cooption, regression, and growth in tumors mediated by
angiopoietins and VEGF. Science. 284:1994–1998. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Eriksson A, Cao R, Pawliuk R, Berg SM,
Tsang M, Zhou D, Fleet C, Tritsaris K, Dissing S, Leboulch P and
Cao Y: Placenta growth factor-1 antagonizes VEGF-C-induced
angiogenesis and tumor growth by the formation of functionally
inactive PlGF-1/VEGF heterodimers. Cancer Cell. 1:99–108. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu GC, Yu CY, She L, Tan HL, Li G, Ren
SL, Su ZW, Wei M, Huang DH, Tian YQ, et al: Metadherin regulation
of vascular endothelial growth factor expression is dependent upon
the PI3K/Akt pathway in squamous cell carcinoma of the head and
neck. Medicine (Baltimore). 94:e5022015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Emdad L, Lee SG, Su ZZ, Jeon HY, Boukerche
H, Sarkar D and Fisher PB: Astrocyte elevated gene-1 (AEG-1)
functions as an oncogene and regulates angiogenesis. Proc Natl Acad
Sci USA. 106:pp. 21300–21305. 2009; View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fu M, Wang C, Li Z, Sakamaki T and Pestell
RG: Minireview: Cyclin D1: Normal and abnormal functions.
Endocrinology. 145:5439–5447. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li PP, Feng LL, Chen N, Ge XL, Lv X, Lu K,
Ding M, Yuan D and Wang X: Metadherin contributes to the
pathogenesis of chronic lymphocytic leukemia partially through
Wnt/β-catenin pathway. Med Oncol. 32:4792015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cano A, Pérez-Moreno MA, Rodrigo I,
Locascio A, Blanco MJ, Del Barrio MG, Portillo F and Nieto MA: The
transcription factor snail controls epithelial-mesenchymal
transitions by repressing E-cadherin expression. Nat Cell Biol.
2:76–83. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Kong X, Huo Q, Guo H, Yan S, Yuan C,
Moran MS, Shao C and Yang Q: Metadherin enhances the invasiveness
of breast cancer cells by inducing epithelial to mesenchymal
transition. Cancer Sci. 102:1151–1157. 2011. View Article : Google Scholar : PubMed/NCBI
|