Introduction
The prevalence of thyroid carcinoma has been
increasing worldwide. Thyroid carcinoma has become the most common
malignant tumor of the endocrine system; it is ranked second among
all female tumors, and in particular there has been a 5.7-fold
increase in papillary thyroid carcinoma (PTC) incidence (1,2). The
standard therapy for PTC involves surgical resection,
chemotherapeutics and radiotherapy (3,4). However,
there are few effective treatment measures for patients with
metastatic thyroid carcinoma. Autophagy is involved in numerous
physiopathological processes and in cancer autophagy has become a
novel target for the investigation of tumorigenesis (5–7).
Beclin-1 and microtubule-associated protein light
chain 3 (LC3) genes serve an important role in mammalian autophagy.
Beclin-1 interacts with several cofactors to regulate the lipid
kinase Vps-34 protein and promote formation of Beclin-1-Vps34-Vps15
core complexes, thereby inducing autophagy (8). Beclin-1 also acts as a tumor suppressor.
In mice, when heterozygous disruption occurs, autophagy is reduced,
cellular proliferation increased and spontaneous tumor development
occurs (9,10).
LC3 is comprised of LC3-I and LC3-II. LC3-II
correlates with autophagy, being recruited into autophagosomes.
Autophagy is used by organisms as a defense strategy to face
environmental stress. LC3-I is formed by removal of the C-terminal
22 amino acids from newly synthesized LC3, followed by conversion
of a fraction of LC3-I into LC3-II. LC3-II is the first mammalian
protein identified that specifically associates with autophagosome
membranes (11,12).
In the present study, Beclin-1 and LC3-II protein
levels were detected in human PTC and adjacent normal tissues, and
the changes and clinical significance of autophagy were
investigated in PTC.
Materials and methods
Patients and samples
A total of 50 human PTC samples and their paired
adjacent noncancerous tissue samples were obtained with informed
consent from patients of Tangshan Worker Hospital (Tangshan, China)
from April to November 2014. The patients in the present study
comprised 36 women and 14 men, with an age range of 17–67 years
(median, 47.12±15.37 years). The diagnosis of PTC was confirmed by
histopathology. The most representative PTC and their adjacent
normal tissues were selected for study. The tissue specimens were
snap-frozen immediately in liquid nitrogen following resection from
the patients, and the samples were frozen until the time of protein
extraction. None of the patients had received preoperative
chemotherapy, radiation therapy or other biological therapy prior
to the operation. All samples were deposited in the Central
Laboratory of the Tangshan Worker Hospital. Diagnosis and
histological typing of thyroid cancer was performed according to
the World Health Organization 2004 thyroid histology classification
standards (13). All cases of PTC
were staged according to the 2009 Tumor-Node-Metastasis (TNM)
classification (14). The
histopathological features included a papillary structure, ground
glass-like nuclei, nuclear overlap, folding of the nuclear membrane
and nuclear inclusion bodies. The present study was approved by the
Institutional Ethics Review Board of Tangshan Worker Hospital.
Immunohistochemical analysis
Rabbit anti-human Beclin-1 polyclonal antibody
(sc-11427), rabbit anti-human LC3-II polyclonal antibody (sc-28266)
and rabbit anti-human β-actin polyclonal antibody (sc-130656) were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA).
Immunohistochemical staining was performed on 4-µm
thick sections of the most representative tumor paraffin block.
Sections were dewaxed in 3% H2O2 for 10 min.
Antigen retrieval was performed with citrate buffer in a microwave
for 3 min. Following blocking with 5% Normal Goat Serum For
Blocking (PH0424; Phygene Life Sciences, Fuzhou, China) at room
temperature for 15 min, the primary antibodies (1:100) were
subsequently applied overnight at 4°C. Following washing in TBS
three times, the sections were incubated with a biotin-conjugated
secondary goat anti-rabbit antibody (1:1,000; ab6720; Abcam,
Cambridge, UK) at 37°C for 1 h. Following treatment with
3,3′-diaminobenzidine, sections were counterstained with
hematoxylin, dehydrated through graded alcohols, cleared with
dimethyl benzene and mounted with resin.
Immunostaining was semiquantitatively evaluated by 2
independent observers. Beclin-1 and LC3-II were considered positive
by cytoplasmic and/or cytomembrane staining. A total of 10 high
power fields were randomly observed, and 100 cells in each view
were counted using a BX-60 microscope (Olympus Corporation, Tokyo,
Japan). Comprehensive evaluation was according to the intensity and
percentage of the stained tumor cells. The staining intensity was
classified into 4 grades: 0 (no staining), 1 (yellow), 2 (deep
yellow) and 3 (brown). The percentage of positive cells was scored
in 4 grades: 0 (0–10%), 1 (11–25%), 2 (26–50%) and 3 (51–100%). The
immunohistochemical expression level was based on the total points.
Total points = staining intensity total + total percentage of
positive cells. The specimens were classified into two groups:
Negative expression, 0–1 points; positive expression, 2–6
points.
Western blotting
Protein was extracted from nitrogen frozen tissue
fragments of samples. The tissues were homogenized in 1 ml
radioimmunoprecipitation assay buffer, treated with protease
inhibitor cocktail, incubated for 20 min on ice and subsequently
centrifuged at 12,000 × g for 15 min at 4°C. The supernatant was
collected, boiled for 15 min and preserved at −80°C. Proteins were
separated by 10% SDS-PAGE (Beyotime Institute of Biotechnology,
Haimen, China), followed by electroblotting to a nitrocellulose
membrane (EMD Millipore, Billerica, MA, USA), After 1 h incubation
in blocking solution (TBS containing 0.1% Tween-20 and 5% nonfat
milk), membranes were incubated with primary antibodies,
anti-beclin-1 (1:1,000) or anti-LC3-II (1:1,000) overnight at 4°C.
Anti-β-actin antibody (1:1,000) was used as a loading control.
Subsequently, membranes were incubated with a secondary antibody
(horseradish peroxidase-conjugated goat anti-rabbit antibody;
1:1,000; ab6721; Abcam) 37°C for 2 h. Following washing three times
with TBS for 15 min at room temperature, the membranes were treated
with a chemiluminescence detection kit (Fast Western Blot kit, ECL
Substrate; #35055; Pierce; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). Protein bands were quantified using densitometry.
Statistical analysis
Statistical evaluation was performed using the SPSS
version 13.0 software package (SPSS, Inc., Chicago, IL, USA). The
association between Beclin-1 and LC3-II protein expression and
clinicopathological features was analyzed by χ2 test.
Spearman's test was used to evaluate the correlation between
Beclin-1 and LC3-II protein expressions. P<0.05 was considered
to indicate a statistically significant difference.
Results
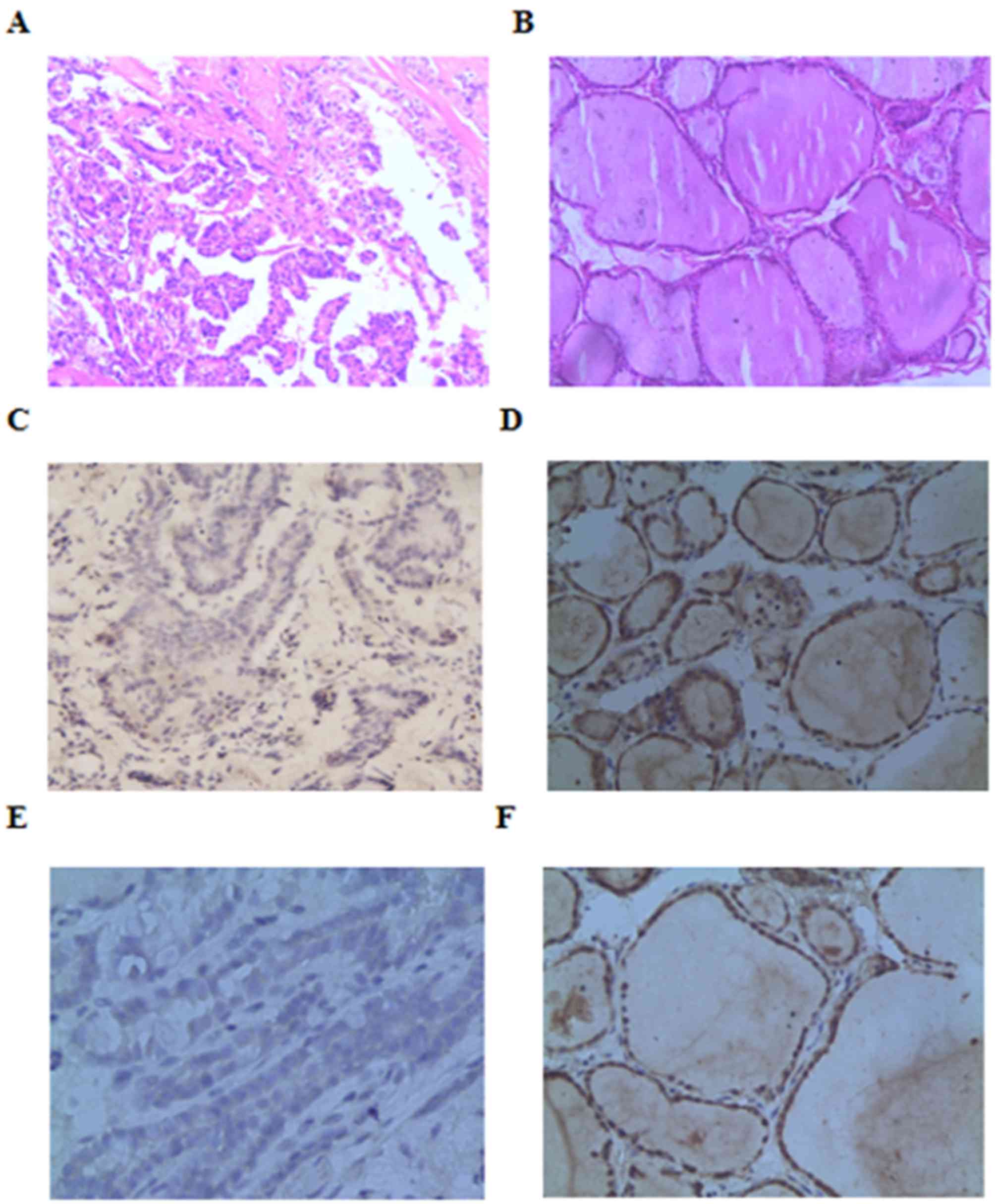
Beclin-1 and LC3-II immunohistochemistry
demonstrated 18 patients with positive expression of Beclin-1
protein and 15 patients with positive expression of LC3-II protein
in PTC (Fig. 1). Western blot
analysis was performed to confirm the specificity of Beclin-1 and
LC3-II antibody, As presented in Fig.
2, lanes 1 and 2 exhibited faint bands for Beclin-1 and LC3-II
in PTC samples, and normal thyroid tissue samples yielded stronger
bands for Beclin-1 and LC3-II. Beclin-1 and LC3-II expression in
PTC tissues was significantly reduced compared with in normal
tissue (Table I). Expression of
Beclin-1 and LC3-II was associated with lymph node metastasis in
PTC, but had no association with age, sex, tumor size, number of
tumors and TNM stage (Table II).
 | Table I.Expression of Beclin-1 and LC3-II in
PTC and normal tissue (n=50). |
Table I.
Expression of Beclin-1 and LC3-II in
PTC and normal tissue (n=50).
|
|
| Beclin-1 | LC3-II |
|---|
|
|
|---|
| Group | Cases | Negative | Positive | χ2 | P-value | Negative | Positive | χ2 | P-value |
|---|
| PTC | 50 | 32 | 18 | 19.869 | <0.001 | 35 | 15 | 29.743 | <0.001 |
| Normal | 50 | 10 | 40 |
|
| 8 | 42 |
|
|
| Table II.Association between Beclin-1 and
LC3-II protein expression levels and clinicopathological
characteristics of papillary thyroid carcinoma. |
Table II.
Association between Beclin-1 and
LC3-II protein expression levels and clinicopathological
characteristics of papillary thyroid carcinoma.
|
|
| Beclin-1 | LC3-II |
|---|
|
|
|---|
| Variables | Cases | Negative | Positive | χ2 | P-value | Negative | Positive | χ2 | P-value |
|---|
| Age, years |
|
|
| 3.766 | 0.067 |
|
| 0.004 | 0.948 |
|
<45 | 17 | 14 | 3 |
|
| 12 | 5 |
|
|
| ≥45 | 33 | 18 | 15 |
|
| 23 | 10 |
|
|
| Gender |
|
|
| 0.001 | 0.979 |
|
| 0.019 | 1.000 |
| Male | 14 | 9 | 5 |
|
| 10 | 4 |
|
|
|
Female | 36 | 23 | 13 |
|
| 25 | 11 |
|
|
| Tumor size, cm |
|
|
| 2.335 | 0.126 |
|
| 2.068 | 0.215 |
|
<1 | 29 | 16 | 13 |
|
| 18 | 11 |
|
|
| ≥1 | 21 | 16 | 5 |
|
| 17 | 4 |
|
|
| Tumor number |
|
|
| 3.979 | 0.056 |
|
| 0.680 | 0.507 |
|
Single | 36 | 20 | 16 |
|
| 24 | 12 |
|
|
|
Multiple | 14 | 12 | 2 |
|
| 11 | 3 |
|
|
| TNM stage |
|
|
| 1.666 | 0.398 |
|
| 3.488 | 0.087 |
| I and
II | 43 | 26 | 17 |
|
| 28 | 15 |
|
|
| III and
IV | 7 | 6 | 1 |
|
| 7 | 0 |
|
|
| Lymph node
involvement |
|
|
| 5.640 | 0.026 |
|
| 6.320 | 0.019 |
|
Present | 16 | 14 | 2 |
|
| 15 | 1 |
|
|
|
Absent | 34 | 18 | 16 |
|
| 20 | 14 |
|
|
Spearman's test was used to evaluate the correlation
between Beclin-1 and LC3-II protein expression in PTC. The results
revealed Beclin-1 and LC3-II expression was positively correlated
in PTC (r=0.327; P=0.020).
Discussion
Autophagy is an evolutionarily conserved processes
that regulates cell fate. Under certain circumstances, autophagy
constitutes a stress adaptation that avoids cell death. Portions of
the cytoplasm are sequestered within double-membrane cytosolic
vesicles termed autophagosomes, the autophagosomes are delivered to
a lysosome, degradation into amino acids occurs and nucleotides are
released into the cytoplasm. Under various nutrient limitations,
autophagy may become essential for viability (15–17).
Emerging evidence has correlated impaired autophagy with tumor
progression (18). Autophagy is
considered to play a dual role in the process of tumor formation. A
lack of autophagy genes would destroy the environmental balance and
may lead to the occurrence of tumors; furthermore, autophagy can
promote tumor cells resistance to stress, meaning improved survival
of cancer cells (19). It is thought
that autophagy is a type of anti-tumor mechanism (20).
Beclin-1 is an autophagy-specific protein that
regulates autophagosome formation. The human Beclin-1 gene is
located on chromosome 17q21. In human pancreatic cancer cells,
wogonin is able to activate Beclin-1 and the phosphoinositide
3-kinase (PI3K) signaling pathway, inducing reactive oxygen
species-mediated autophagy (21).
Beclin-1 also serves a role in the occurrence and development of
tumors by regulating autophagy. Research has revealed that Beclin-1
protein is almost undetectable in human breast cancer cell lines
(22). In human thyroid papillary
carcinoma cell lines TPC-1 and 8505-C low levels of autophagosomes
were observed (23). However, certain
experiments have demonstrated the opposite result. In the present
study, it was observed that the expression level of Beclin-1 was
significantly downregulated in human PTC tissues compared with the
normal thyroid tissue; inducing autophagy decline serves a role in
the incidence and development of PTC (24,25).
Analysis of the association between Beclin-1 protein and general
clinical and pathological factors demonstrated that Beclin-1
protein expression decreased more significantly in patients with
lymph node metastases, but had no association with age, sex, tumor
size, the number of tumors and TNM stage. The decline of autophagy
activity may be involved in tumor metastasis. In pcDNA3.1-Bec
transfected CaSki cells, the expression of Beclin-1 protein was
upregulated, and led to arrest in the G0/G1 phase of the cell cycle
(26). In CaSki cells, the apoptosis
signaling induced by anti-cancer drugs may be enhanced by
overexpression of Beclin-1. The results suggest that Beclin-1
serves a significant role in the regulation of potent anti-tumor
activity (26). In a study of tongue
squamous cell carcinoma cell lines SCC9 and SCC15, it was
demonstrated that knockdown of Beclin-1 promoted proliferation,
migration and invasion, while overexpression of Beclin-1 inhibited
proliferation and migration in the two lines cell (27). The data also confirmed that Beclin-1
inhibited TSCC xenograft growth in vivo (27). These results indicate that
autophagy-regulating gene Beclin-1 may be a potential target for
cancer gene therapy.
In the present study, reduced expression of LC3-II
protein was observed in human PTC tissues compared with normal
thyroid tissue. The analysis of the association between LC3-II
protein and general clinical and pathological factors revealed that
LC3-II protein expression was inversely correlated with lymph node
metastases, but had no association with age, sex, tumor size,
number of tumors and TNM stage. Beclin-1 and LC3-II were positively
correlated in PTC. This finding may present novel strategies for
the development of therapies for PTC. LC3 is a yeast autophagy gene
autophagy-related protein 8 homolog in mammalian cells, and is a
specific autophagic marker in mammalian cells (28). LC3 is processed from LC3-I to the
membrane-bound form LC3-II. LC3-II is located on the membrane of
autophagosomes (29). The results of
a previous study of triple-negative breast cancer suggested that
expression of LC3 in triple-negative breast cancer was associated
with increased distant metastases, and LC3 negativity was a
significant independent prognostic factor of disease-free survival
(30). Beclin-1 and LC3 autophagic
genes are altered in several human types of cancer (31). The lowest expression of LC3-II protein
is observed in melanoma metastases; LC3 messenger RNA significantly
decreased with tumor progression, and the expression of LC3-II
protein was inversely correlated to thickness, ulceration and
mitotic rate (32).
In conclusion, compared with normal thyroid tissue,
Beclin-1 and LC3-II expression in PTC was reduced, particularly in
patients with lymph node metastasis. Furthermore, it was observed
that clinical characteristics, including patient age, sex, tumor
size and the number of tumors did not affect Beclin-1 and LC3-II
expression. The results of the present study indicate that a
decline autophagy activity may be associated with PTC metastasis.
However, the regulation of autophagy is a two-way process, and its
regulatory mechanism in tumors was not completely clear. As a form
of programmed cell death, autophagy is regulated by common
signaling pathways with apoptosis, including PI3K/Akt and B-cell
lymphoma 2 family members (33). The
results of the present study may provide initial evidence for the
further investigation of Beclin-2 and LC3-II in PTC. Autophagy may
be a novel target for the prevention or treatment of PTC.
References
|
1
|
Qian BY, He M, Gao M and Chen KX: Thyroid
cancer incidence in the city of Tianjin during 2002–2006 and its
secular trend in recent 26 year. Chin J Gen Surg. 4:275–278.
2011.
|
|
2
|
La Vecchia C, Malvezzi M, Bosetti C,
Garavello W, Bertuccio P, Levi F and Negri E: Thyroid cancer
mortality and incidence: A global overview. Int J Cancer.
136:2187–2195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grant CS: Recurrence of papillary thyroid
cancer after optimized surgery. Gland Surg. 4:52–62.
2015.PubMed/NCBI
|
|
4
|
Jeong GC, Song M, Park HJ, Min JJ, Bom HS,
Cho SG, Park KS, Kang SR, Kim J, Song HC and Kwon SY: Iodine uptake
patterns on post-ablation whole body scans are related to elevated
serum thyroglobulin levels after radioactive iodine therapy in
patients with papillary thyroid carcinoma. Nucl Med Mol Imaging.
50:329–336. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Perlmutter DH: The role of autophagy in
alpha-1-antitrypsin deficiency: A specific cellular response in
genetic diseases associated with aggregation-prone proteins.
Autophagy. 2:258–263. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Terman A and Brunk UT: Autophagy in
cardiac myocyte homeostasis, aging, and pathology. Cardiovasc Res.
68:355–365. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fogel AI, Dlouhy BJ, Wang C, Ryu SW,
Neutzner A, Hasson SA, Sideris DP, Abeliovich H and Youle RJ: Role
of membrane association and Atg14-dependent phosphorylation in
beclin-1-mediated autophagy. Mol Cell Biol. 33:3675–3688. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qu X, Yu J, Bhagat G, Furuya N, Hibshoosh
H, Troxel A, Rosen J, Eskelinen EL, Mizushima N, Ohsumi Y, et al:
Promotion of tumorigenesis by heterozygous disruption of the beclin
1 autophagy gene. J Clin Invest. 112:1809–1820. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pirtoli L, Cevenini G, Tini P, Vannini M,
Oliveri G, Marsili S, Mourmouras V, Rubino G and Miracco C: The
prognostic role of Beclin 1 protein expression in high-grade
gliomas. Autophagy. 5:930–936. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Klionsky DJ, Abdalla FC, Abeliovich H,
Abraham RT, Acevedo-Arozena A, Adeli K, Agholme L, Agnello M,
Agostinis P, Aguirre-Ghiso JA, et al: Guidelines for the use and
interpretation of assays for monitoring autophagy. Autophagy.
8:445–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
DeLellis RA, Lloyd R, Heitz P and Eng C:
Pathology and Genetics of Tumours of Endocrine Organs (IARC WHO
Classification of Tumours). 1st. IARC Press; Lyon: 2004
|
|
14
|
Sobin LH, Gospodarowicz MK and Wittekind
C: TNM Classification of Malignant Tumours. 7th. Wiley-Blackwell;
Oxford: 2009
|
|
15
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: Crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thorburn A: Apoptosis and autophagy:
Regulatory connections between two supposedly different processes.
Apoptosis. 13:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mizushima N and Klionsky DJ: Protein
turnover via autophagy: Implications for metabolism. Annu Rev Nutr.
27:19–40. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Helgason GV, Holyoake TL and Ryan KM: Role
of autophagy in cancer prevention, development and therapy. Essays
Biochem. 55:133–151. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Katheder NS, Khezri R, O'Farrell F,
Schultz SW, Jain A, Rahman MM, Schink KO, Theodossiou TA, Johansen
T, Juhász G, et al: Microenvironmental autophagy promotes tumour
growth. Nature. 541:417–420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morselli E, Galluzzi L, Kepp O, Vicencio
JM, Criollo A, Maiuri MC and Kroemer G: Anti- and pro-tumor
functions of autophagy. Biochim Biophys Acta. 1793:1524–1532. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li SJ, Sun SJ, Gao J and Sun FB: Wogonin
induces Beclin-1/PI3K and reactive oxygen species-mediated
autophagy in human pancreatic cancer cells. Oncol Lett.
12:5059–5067. 2016.PubMed/NCBI
|
|
22
|
Wang Riwei and Wu Aiguo: Autophagy gene
Beclinl and breast cancer. Chin J Gen Surg. 21:591–596. 2012.
|
|
23
|
Lin CI, Whang EE, Abramson MA, Jiang X,
Price BD, Donner DB, Moore FD Jr and Ruan DT: Autophagy: A new
target for advanced papillary thyroid cancer therapy. Surgery.
146:1208–1214. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li X, Xu H and Ma H: Beclin 1 is highly
expressed in papillary thyroid carcinoma and correlates with lymph
node metastasis. Acta Chir Belg. 113:175–181. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yeşil C, Kandemir O, Haksever H and
Dabakoğlu T: Is BECLIN-1 Immunoreactivity more effective than
HBME-1 in diagnosis of papillary thyroid cancer? Acta Chir Belg.
115:299–305. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun Y, Liu JH, Jin L, Lin SM, Yang Y, Sui
YX and Shi H: Over-expression of the Beclin1 gene upregulates
chemosensitivity to anti-cancer drugs by enhancing therapy-induced
apoptosis in cervix squamous carcinoma CaSki cells. Cancer Lett.
294:204–210. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Weng J, Wang C, Wang Y, Tang H, Liang J,
Liu X, Huang H and Hou J: Beclin1 inhibits proliferation, migration
and invasion in tongue squamous cell carcinoma cell lines. Oral
Oncol. 50:983–990. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kraft C, Kijanska M, Kalie E, Siergiejuk
E, Lee SS, Semplicio G, Stoffel I, Brezovich A, Verma M, Hansmann
I, et al: Binding of the Atg1/ULK1 kinase to the ubiquitin-like
protein Atg8 regulates autophagy. EMBO J. 31:3691–3703. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang H, Cheng D, Liu W, Peng J and Feng
J: Protein kinase C inhibits autophagy and phosphorylates LC3.
Biochem Biophys Res Commun. 395:471–476. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He JH, Luo RZ, Cai MY, Li M, Lu JB and
Yuan ZY: Decreased expression of light chain 3 (LC3) increased the
risk of distant metastasis in triple-negative breast cancer. Med
Oncol. 30:4682013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Miracco C, Cevenini G, Franchi A, Luzi P,
Cosci E, Mourmouras V, Monciatti I, Mannucci S, Biagioli M, Toscano
M, et al: Beclin 1 and LC3 autophagic gene expression in cutaneous
melanocytic lesions. Hum Pathol. 41:503–512. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Marino ML, Pellegrini P, Di Lernia G,
Djavaheri-Mergny M, Brnjic S, Zhang X, Hägg M, Linder S, Fais S,
Codogno P and De Milito A: Autophagy is a protective mechanism for
human melanoma cells under acidic stress. J Biol Chem.
287:30664–30676. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Boya P, González-Polo RA, Casares N,
Perfettini JL, Dessen P, Larochette N, Métivier D, Meley D,
Souquere S, Yoshimori T, et al: Inhibition of macroautophagy
triggers apoptosis. Mol Cell Biol. 25:1025–1040. 2005. View Article : Google Scholar : PubMed/NCBI
|