Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of
the most common malignant neoplasms of the pancreas and presents
with poor prognosis (5-year survival rate, <5%) (1). Chemoradiation is the conventional option
for patients with PDAC. However, due to the inherent
chemoresistance and radioresistance of PDAC, such combined modality
therapy consistently fails to improve outcomes (2).
Cancer stem cells (CSCs) are capable of unlimited
self-renewal, and through asymmetric division, they give rise to
further differentiated cells. CSCs are resistant to chemotherapy
and radiation therapy, compared to differentiated cells; a number
of previous studies revealed that tumor recurrence or metastasis
following anticancer treatment could be attributed to CSCs
(3–5).
The existence of cancer stem cells was first shown in the context
of acute myelogenous leukemia, and subsequently verified in breast
and brain tumors. In 2007, Li et al (3) reported that the cluster of
differentiation (CD)44+CD24+
epithelial-specific antigen+ pancreatic cancer cells
exhibited the stem cell properties of self-renewal, the ability to
produce differentiated progeny and increased expression of the
developmental signaling molecule sonic hedgehog. These cells
exhibited the following main characteristics: Tumorigenic capacity;
specific molecular markers; and responsibility for the maintenance
of tumor growth and resistant to chemo- or radiation therapy. Dou
et al (6) used the
cell-surface markers CD44+, CD24+ and
CD133+ to identify cancer stem-like cells in murine
melanoma B16F10 cells, and revealed that
CD44+CD24+CD133+ cells exhibited
biological properties of cancer stem-like cells and behaved
similarly to CSCs. In addition, previous studies (7,8) identified
that chemoradiation resistance in PDAC cells may be linked to
pancreatic CSCs (PCSCs). Therefore, understanding the nascency and
regulation of PCSCs may be critical for the identification of more
effective treatments for patients with PDAC.
Reactive oxygen species (ROS) regulate a broad array
of signal transduction pathways in multiple biological processes,
including cell growth, differentiation, gene expression and
apoptosis. ROS production contributes to tumor cell apoptosis
following exposure to infrared and other stressors, including high
glucose, angiotensin and tumor necrosis factor-α (9).
In the present study, PANC-1 cells were isolated and
sorted into CD44+CD24+,
CD44−CD24+, CD44+CD24−
and CD44−CD24− using flow cytometry. The
sensitizer enhancement ratio (SER) was then examined in the four
subsets. At the same time, the effect of radiation on cell
apoptosis, cycle distribution and the level of intracellular ROS
was examined. In addition, it was also investigated whether ROS and
cell cycle were able to affect radioresistance.
The results demonstrated that decreased levels of
ROS and apoptosis in CD44+CD24+ cells may
contribute to their resistance to radiation.
Materials and methods
Reagents
Dulbecco's modified Eagle's medium (DMEM), DMEM:
Nutrient mixture F-12 (DMEM/F-12) and B-27 supplement were
purchased from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Annexin V-fluorescein isothiocyanate (FITC) and propidium
iodide (PI) were purchased from Nanjing KeyGen Biotech. Co. Ltd.
(Nanjing, China). Fetal bovine serum (FBS) was purchased from
Zhejiang Tianhang Biological Technology Co., Ltd. (Huzhou, China).
Trypsin was purchased from Sigma-Aldrich (Merck KGaA, Darmstadt,
Germany). Anti-human CD24 (cat. no. 173–820) and FITC-anti-human
CD44 (cat. no. 193–040) were purchased from Ancell Corporation
(Bayport, MN, USA). The 2′,7′-dichlorofluorescin diacetate
(DCFH-DA) probe was purchased from Sigma-Aldrich (Merck KGaA). The
Cell Lab Quanta SC flow cytometer was purchased from Beckman
Coulter, Inc. (Brea, CA, USA). Medical linear accelerators were
purchased from Siemens AG (Munich, Germany).
Cell culture
The human pancreatic cancer PANC-1 cell line was
purchased from the Cell Bank of the Institute of Biochemistry and
Cell Biology, Shanghai Institutes for Biological Sciences
(Shanghai, China). Cells were cultured in DMEM supplemented with
10% FBS and penicillin/streptomycin at 37°C in a humidified 5%
CO2 atmosphere.
Radiation
Cells were seeded onto 6-well tissue culture plates
and incubated for 12 h as described previously, then treated with a
single dose of radiation with 6 MV X-ray at room temperature. The
initial dose rate was 300 cGy/min (SSD, 100 cm; gantry, 0°; and
radiation field, 15×15 cm).
Flow cytometric analysis and
fluorescence-activated cell sorting (FACS)
Cells in the exponential growth phase were
dissociated by trypsin-EDTA solution (trypsin, 0.25%; EDTA, 0.02%)
for 2–5 min at 37°C. Cells were then transferred to a 5-ml tube,
washed twice with PBS and 2% heat-inactivated calf serum (Zhejiang
Tianhang Biological Technology Co., Ltd., Huzhou, China),
centrifuged for 5 min at 800 × g at 37°C, re-suspended in 100 µl
(per 106 cells) of PBS, and counted by flow cytometry.
The previously described anti-human CD24 or FITC anti-human CD44
antibodies were diluted to 1:100, added and incubated for 30 min at
4°C, and then washed twice with PBS. FACS was performed, and data
were analyzed with the Cell Quest software (version 3.0; BD
Biosciences, Franklin Lakes, NJ, USA). Using forward and side
scatter profiles, debris and dead cells were gated out. Cells were
routinely sorted twice and reanalyzed for purity.
CD44+CD24+, CD44−CD24+,
CD44+CD24− and
CD44−CD24− were obtained. Cells were then
cultured in DMEM/F-12 supplemented with FBS and
penicillin/streptomycin/B-27 supplement at 37°C in a humidified 5%
CO2 atmosphere.
Clonogenic assay
Cells in the exponential growth phase were seeded
onto 6-well tissue culture plates (105 cells/well) with
triplicate repeats for each cell group. Following 24 h, cells were
treated with a single dose of radiation (0, 2, 4, 6 or 8 Gy) at
room temperature, then incubated for 14 days without changing the
culture medium. Cells were then fixed with methanol and stained
with 0.05% crystal violet (Nanjing KeyGen Biotech Co., Ltd.),
according to the manufacturer's protocol. The number of colonies
with >50 cells were counted under a dissecting microscope
(magnification, ×400). Cell survival was determined by a colony
formation assay. The plating efficiency (PE) and survival fraction
(SF) were calculated as follows: PE (%) = (number of
colonies/inoculating cell number) × 100; SF = number of colonies
counted [cells seeded × (PE/100)]. All experiments were repeated
three times. According to the target model [S = 1 - (1 -
eD/D0 N], in which S was the cell survival
rate, D was the dose, D0 was the mean
lethal dose and N was the extrapolation number), cell survival
curves were drawn using GraphPad Prism version 6 software (GraphPad
Software, Inc., La Jolla, CA, USA). The SER was calculated as SER =
Dq (CD44+CD24+)/Dq
(CD44+CD24−,
CD44−CD24+,
CD44−CD24−), where Dq was the
quasi-threshold dose (Dq = D0 × lnN), as
previously described (10).
Cell cycle and apoptosis analysis
The sorted cells were exposed to radiation dosages
(2 Gy). The cells were collected following 48 h of radiation. For
the detection of apoptotic cells, the cells were trypsinized and
stained with acridine orange (Nanjing KeyGen Biotech Co., Ltd.),
and the cells were observed and counted under a fluorescence
microscope (magnification, ×400). The cells used for the apoptosis
analysis were stained with PI and Annexin V. The cells used for
cell cycle analysis were stained with PI subsequent to ethanol
fixation. Each analysis was performed four times.
Measurement of intracellular ROS
The sorted cells were exposed to radiation dosages
(2 Gy). The cells were collected following 48 h of radiation. The
production of intracellular ROS was measured by performing flow
cytometry using the oxidation-sensitive probe DCFH-DA. Briefly, 10
mM DCFH-DA stock solution (in methanol) was diluted 4,000-fold in
cell culture medium without serum or other additives to yield a 2.5
mM working solution. Following the exposure of human umbilical
endothelial cells to silica nanoparticles for 3 and 24 h,
respectively, the cells in 6-well plates were washed twice with PBS
and incubated in 2 ml working solution of DCFH-DA at 37°C in the
dark for 30 min. The cells were then washed twice with cold PBS and
re-suspended in the PBS for analysis of intracellular ROS by FACS.
Experiments were repeated four times.
Statistical analysis
All analyses were performed using SPSS 16.0 (SPSS,
Inc., Chicago, IL, USA). Data were expressed as the mean ± standard
deviation. Statistically significant differences were determined by
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Presence of CD44 and CD24 on the cell
surface of pancreatic carcinoma cell lines
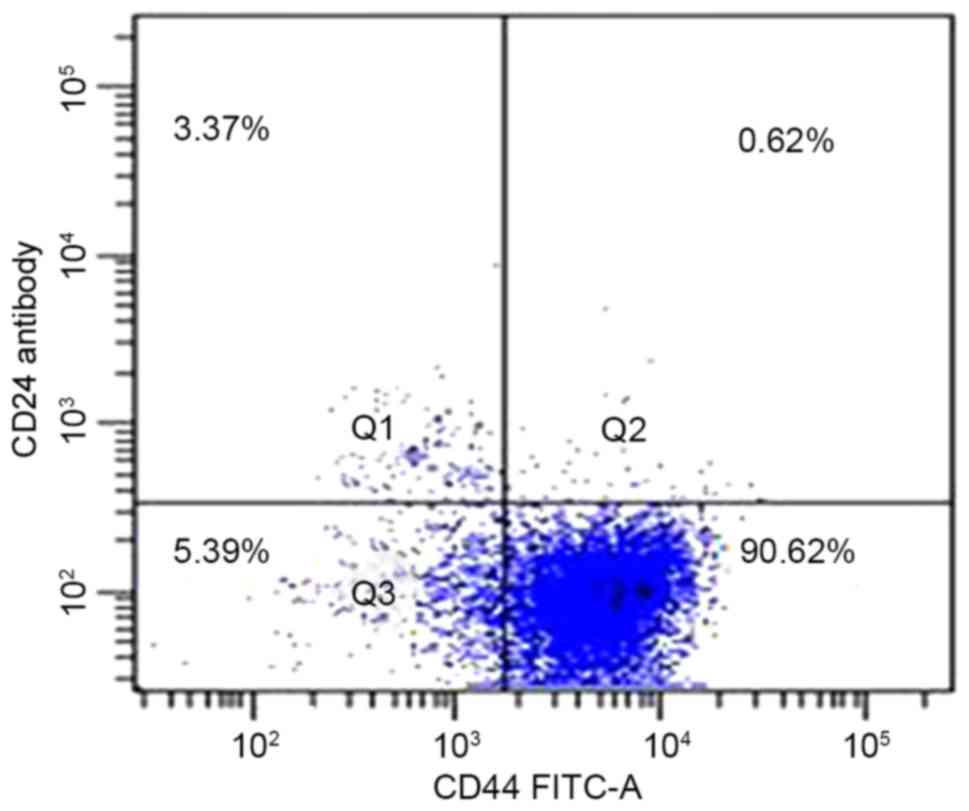
Flow cytometric analysis was used to determine the
presence of CD44 and CD24 on the cell surface of the pancreatic
adenocarcinoma PANC-1 cell line. A total of 92% of cells expressed
the cell surface marker CD44, and 4.7% expressed CD24;
CD44+CD24+, CD44+CD24−,
CD44−CD24+ and
CD44−CD24− were 0.6±0.2, 89.3±2.6, 4.1±1.3
and 6.0±1.7%, respectively. Typical samples are shown in Fig. 1.
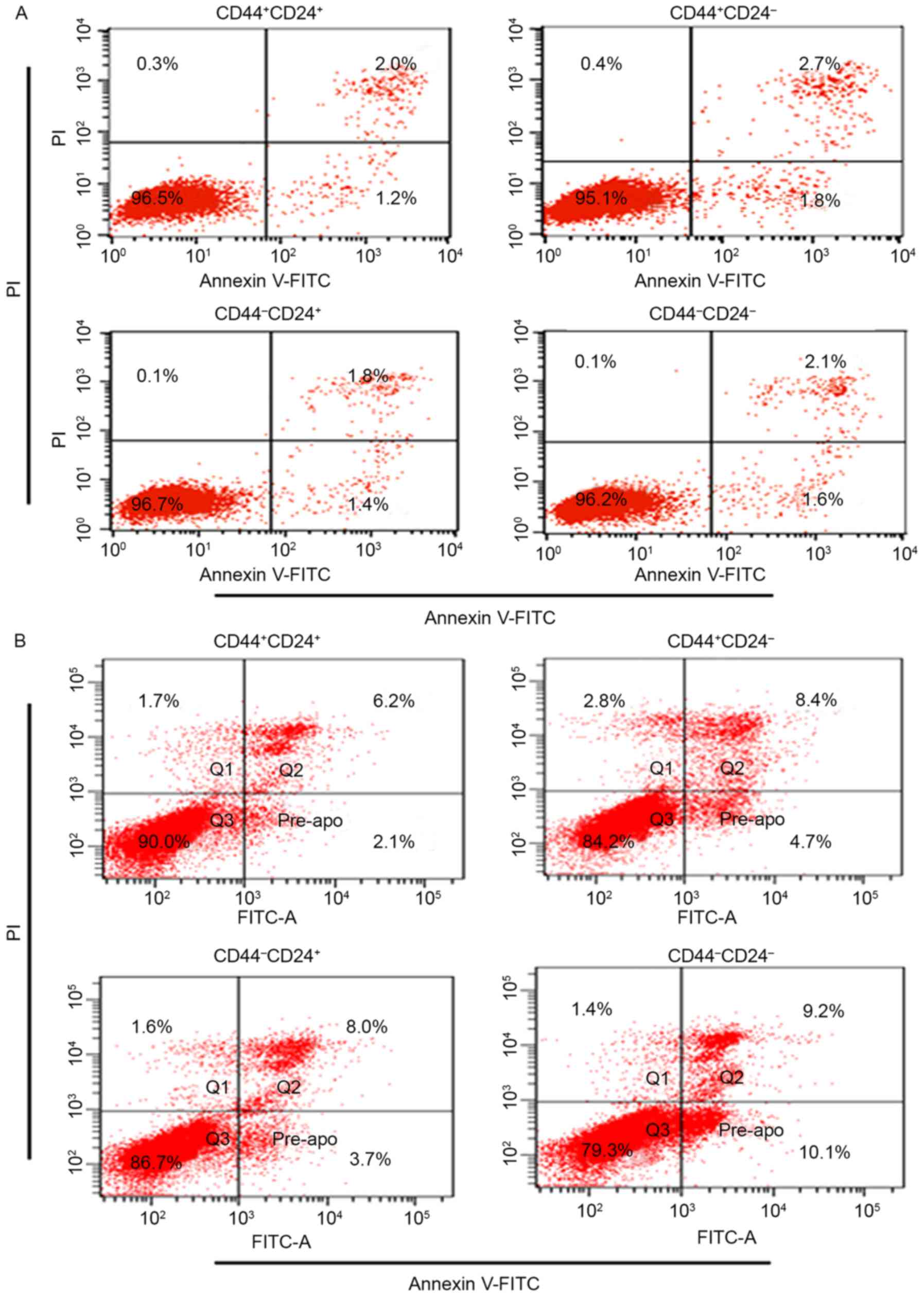
Apoptosis of sorted cells
Prior to radiation, the percentage of apoptosis for
CD44+CD24+ (3.3±0.6%),
CD44+CD24− (4.0±0.8%),
CD44−CD24+ (3.4±0.7%) and
CD44−CD24− (3.5±0.8) was not significantly
different (P>0.05; Fig. 2A).
Following 48 h of radiation, the results revealed
that radiation induced a lower percentage of apoptosis in
CD44+CD24+ when compared with others (6.8±1.1
vs. 10.4±2.7%, P<0.01; 6.8±1.1 vs. 13±3.1%, P<0.01; 6.8±1.1
vs. 26.3±2.4%, P<0.01). The differences were significant
(Fig. 2B).
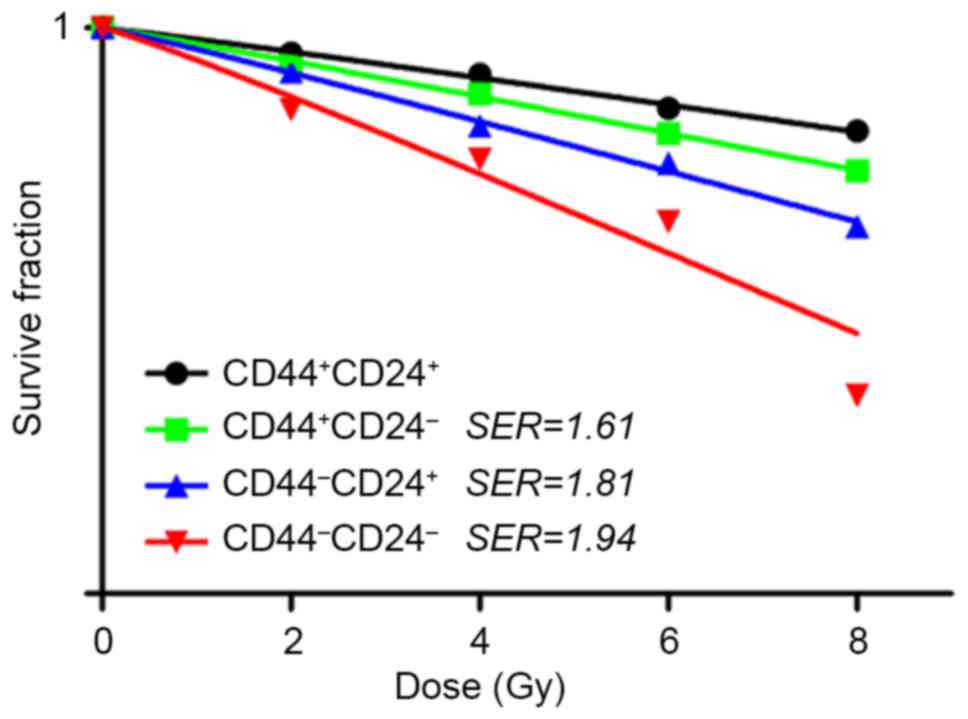
Clonogenic survival rates
Sorted cells were exposed to various radiation
dosages (0, 2, 4, 6 and 8 Gy) and their survival fractions were
measured based on colony formation. The dose-response curves for
the cell-killing effects of radiation are shown in Fig. 3. The survival fractions of
CD44−CD24− were decreased exponentially as
the dose of radiation increased. The SERs for
CD44+CD24−, CD44−CD24+
and CD44−CD24− were 1.61, 1.81 and 1.94,
respectively.
Cell cycle analysis
Prior to radiation, the percentage of G0/G1 was
highest in CD44+CD24+ (63.8±1.7 vs.
58.2±2.2%, P<0.01; 63.8±1.7 vs. 53.4±2.7%, P<0.01; 63.8±1.7
vs. 50.1±3.4%, P<0.01; Table
I).
 | Table I.Cell cycle distribution for four
types of cells. |
Table I.
Cell cycle distribution for four
types of cells.
|
| Cell cycle
distribution |
|---|
|
|
|
|---|
| Sorted cells | G0/G1 phase | S phase | G2/M phase |
|---|
| Radiation | − | + | − | + | − | + |
|
CD44+CD24+ | 63.8±1.7 |
67.2±3.4a | 20.2±3.3 | 18.9±2.1 | 16.0±1.6 | 13.9±2.5 |
|
CD44+CD24− |
58.2±2.2b | 57.7±2.9 | 24.1±4.2 |
22.2±3.1b |
17.7±2.4b | 18.1±3.7 |
|
CD44−CD24+ | 53.4±2.7 | 50.3±4.6 |
25.1±1.9a |
23.7±1.6b | 21.5±2.7 |
26.0±2.3a |
|
CD44−CD24− | 50.1±3.4 |
42.8±2.7b |
25.8±2.5b | 27.7±1.9 | 24.1±3.8 | 29.5±4.1 |
Following 48 h of radiation, the percentage of G0/G1
was also highest in CD44+CD24+ (67.2±3.4 vs.
57.7±2.9%, P<0.01; 67.2±3.4 vs. 50.3±4.6%, P<0.01; 67.2±3.4
vs. 42.8±2.7%, P<0.01). The difference was significant (Table I).
Intracellular ROS
Prior to radiation, the level of intracellular ROS
was lowest in CD44−CD24− (1,306±205 vs.
1,483±172, P<0.05; 1,306±196 vs. 1,586±201, P<0.05; 1,306±216
vs. 1,592±216, P<0.05; Fig.
4A).
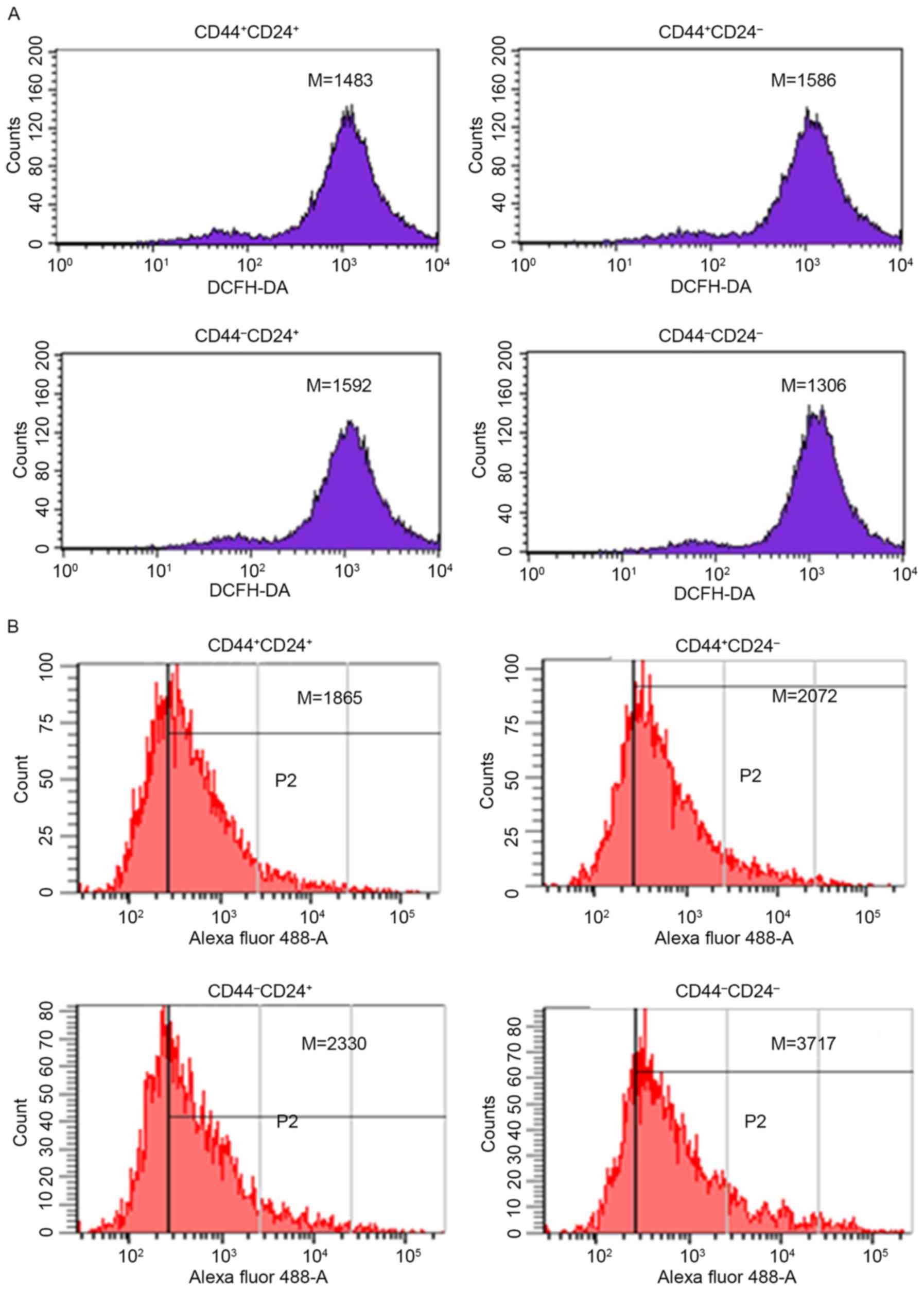 | Figure 4.Level of intracellular ROS for sorted
cells. (A) The level of intracellular ROS for sorted cells prior to
radiation: CD44+CD24+ (1,483±172);
CD44+CD24− (1,586±201);
CD44−CD24+ (1,592±216); and
CD44−CD24− (1,306±205). (B) The level of
intracellular ROS for sorted cells following 48 h of radiation:
CD44+CD24+ (1,865±315);
CD44+CD24− (2,072±167);
CD44−CD24+ (2,330±208); and
CD44−CD24− (3,717±236). Experiments were
repeated four times. ROS, reactive oxygen species; CD cluster of
differentiation; M, median; DCFH-DA, 2′,7′-dichlorofluorescin
diacetate. |
Following 48 h of radiation, the level of
intracellular ROS was lowest in CD44+CD24+
(1,865±315 vs. 2,072±167, P<0.01; 1,865±315 vs. 2,330±208,
P<0.01; 1,865±315 vs. 3,717±236, P<0.01; Fig. 4B).
Discussion
Pancreatic cancer is a devastating disease with a
median survival of only ~6 months (11,12).
Pancreatic cancer is resistant to almost all conventional
therapies. It is imperative to understand the unique logical
characteristics of pancreatic cancer and the scientific mechanisms
for its obstinate malignancy. The cancer stem cell hypothesis is
important to improve understanding of the intrinsic biological
characteristics of pancreatic cancer. This indicates that tumor
progression is initiated and driven by a small subset of
undifferentiated cells with the ability of self-renewal and
differentiation into different integrated and heterogeneous tumor
populations (13,14).
Emerging evidence (14–21) has
indicated that pancreatic adenocarcinoma is hierarchically
organized and sustained by a distinct subpopulation of CSCs, which
are responsible for tumor growth, metastasis and resistance to
therapy. In addition, elimination of these cells is possible and
may subsequently improve the effect of clinical treatment. However,
in previous studies, direct evidence for the existence of CSCs in
human pancreatic cancer was not consistent (22,23). To
isolate and identify pancreatic cancer stem cells more efficiently,
new methods are required for additional studies.
In the present study, the PANC-1 cells were isolated
and sorted into CD44+CD24+,
CD44−CD24+, CD44+CD24−
and CD44−CD24− by flow cytometry. The results
revealed that 92% of cells expressed CD44 and 4.7% expressed CD24.
The survival fractions of CD44−CD24−
decreased exponentially as the dose of radiation increased. The
SERs for CD44+CD24−,
CD44−CD24+ and
CD44−CD24− were 1.61, 1.81 and 1.94,
respectively. Prior to radiation, no significant differences in
apoptosis were observed among the four groups. However, the results
indicated that radiation induced a significantly lower percentage
of apoptosis in CD44+CD24+ when compared with
others. Prior to or following radiation, the percentage of G0/G1
cells was significantly highest in
CD44+CD24+, indicating that the relative
stationary cell cycle is critical for radiation resistance.
In addition, the level of intracellular ROS was
revealed to be lowest in CD44−CD24− prior to
radiation. However, following radiation, the level of intracellular
ROS was lowest in CD44+CD24+. In contrast to
general cancer cells in which ROS levels are increased, CSCs
exhibited reduced levels of ROS. The maintenance of low ROS levels
is essential to preserve CSC self-renewal and stemness. Ishimoto
et al (24) revealed that
CD44+ gastrointestinal CSCs exhibited an enhanced
capacity of glutathione synthesis and defense against ROS by
activation of the cystine-glutamate exchange transporter. These
properties contribute to the radioresistance of stem cells, since
radiation exerts a cytotoxic effect through the generation of free
radicals, and the critical mediator ROS is decreased in stem cells
(25). CSCs are more radioresistant
compared with non-CSCs, and this is partly attributable to the
lower ROS levels and enhanced ROS defenses observed in CSCs
(26).
In summary, the results from the present study have
significant implications for the treatment of pancreatic cancer.
The present study demonstrated that
CD44+CD24+ pancreatic cancer cells possessed
stem cell properties. These cells are more resistant to standard
therapies used to treat pancreatic cancer, including radiotherapy.
An improved understanding of pancreatic cancer stem cells may not
only affect the ability to improve understanding of current
therapeutics, but expression studies of pancreatic cancer stem
cells may help in the identification of novel therapeutic
targets.
Acknowledgements
The present study was supported by the Project of
Lianyungang City Development Commission (grant no. SH1218).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gutt R, Liauw SL and Weichselbaum RR: The
role of radiotherapy in locally advanced pancreatic carcinoma. Nat
Rev Gastroenterol Hepatol. 7:437–447. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li CW, Heidt DG, Dalerba P, Burant CF,
Zhang L, Adsay V, Wicha M, Clarke MF and Simeone DM: Identification
of pancreatic cancer stem cells. Cancer Res. 67:1030–1037. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang P, Zhang J, Zhang L, Zhu Z, Fan J,
Chen L, Zhuang L, Luo J, Chen H, Liu L, et al: MicroRNA 23b
regulates autophagy associated with radioresistance of pancreatic
cancer cells. Gastroenterology. 145:1133–1143.e12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maugeri-Saccà M, Vigneri P and De Maria R:
Cancer stem cells and chemosensitivity. Clin Cancer Res.
17:4942–4947. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dou J, Pan M, Wen P, Li Y, Tang Q, Chu L,
Zhao F, Jiang C, Hu W, Hu K and Gu N: Isolation and identification
of cancer stem-like cells from murine melanoma cell lines. Cell Mol
Immunol. 4:467–472. 2007.PubMed/NCBI
|
|
7
|
Du ZY, Qin RY, Wei CF, Wang M, Shi C, Tian
R and Peng C: Pancreatic cancer cells resistant to
chemoradiotherapy rich in ‘stem-cell-like’ tumor cells. Dig Dis
Sci. 56:741–750. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Al-Assar O, Demiciorglu F, Lunardi S,
Gaspar-Carvalho MM, McKenna WG, Muschel RM and Brunner TB:
Contextual regulation of pancreatic cancer stem cell phenotype and
radioresistance by pancreatic stellate cells. Radiother Oncol.
111:243–251. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tothova Z, Kollipara R, Huntly BJ, Lee BH,
Castrillon DH, Cullen DE, McDowell EP, Lazo-Kallanian S, Williams
IR, Sears C, et al: FoxOs are critical mediators of hematopoietic
stem cell resistance to physiologic oxidative stress. Cell.
128:325–339. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xia S, Zhao Y, Yu S and Zhang M: Activated
PI3K/Akt/COX-2 pathway induces resistance to radiation in human
carvival cancer HeLa cells. Cancer Biother Radiopharm. 25:317–323.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Philip PA, Mooney M, Jaffe D, Eckhardt G,
Moore M, Meropol N, Emens L, O'Reilly E, Korc M, Ellis L, et al:
Consensus report of the national cancer institute clinical trials
planning meeting on pancreas cancer treatment. J Clin Oncol.
27:5660–5669. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang J, Dhakal I, Yan H, Phillips M and
Kesteloot H: SEER Cancer Registries: Trends in pancreatic cancer
incidence in nine SEER cancer registries, 1973–2002. Ann Oncol.
18:1268–1279. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gellmini S, Mangoni M, Serio M, Romagnani
P and Lazzeri E: The critical role of SDF-1/CXCR4 axis in cancer
and cancer stem cells metastasis. J Endocrinol Invest. 31:809–819.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dirks P: Cancer stem cells: Invitation to
a second round. Nature. 466:40–41. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li YF, Xiao B, Tu SF, Wang YY and Zhang
XL: Cultivation and identification of colon cancer stem
cell-derived spheres from the Colo205 cell line. Braz J Med Biol
Res. 45:197–204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakanishi Y, Seno H, Fukuoka A, Ueo T,
Yamaga Y, Maruno T, Nakanishi N, Kanda K, Komekado H, Kawada M, et
al: Dclk1 distinguishes between tumor and normal stem cells in the
intestine. Nat Genet. 45:98–103. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen J, Li YJ, Yu TS, McKay RM, Burns DK,
Kernie SG and Parada LF: A restricted cell population propagates
glioblastoma growth after chemotherapy. Nature. 488:522–526. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Machado HL, Kittrell FS, Edwards D, White
AN, Atkinson RL, Rosen JM, Medina D and Lewis MT: Separation by
cell size enriches for mammary stem cell repopulation activity.
Stem Cell Transl Med. 2:199–203. 2013. View Article : Google Scholar
|
|
20
|
Driessens G, Beck B, Caauwe A, Simons BD
and Blanpain C: Defining the mode of tumour growth by clonal
analysis. Nature. 488:527–530. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hinohara K, Kobayashi S, Kanauchi H,
Shimizu S, Nishioka K, Tsuji E, Tada K, Umezawa K, Mori M, Ogawa T,
et al: ErbB receptor tyrosine kinase/NF-κB signaling controls
mammosphere formation in human breast cancer. Proc Natl Acad Sci
USA. 109:pp. 6584–6589. 2012; View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang P, Wang CY, Gou SM, Wu HS, Liu T and
Xiong JX: Isolation and biological analysis of tumor stem cells
from pancreatic adenocarcinoma. World J Gastroentero. 14:3903–3907.
2008. View Article : Google Scholar
|
|
23
|
Lee HJ, You DD, Choi DW, Choi YS, Kim SJ,
Won YS and Moon HJ: Significance of CD133 as a cancer stem cell
markers focusing on the tumorigenicity of pancreatic cancer cell
lines. J Korean Surg Soc. 81:263–270. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ishimoto T, Nagano O, Yae T, Tamada M,
Motohara T, Oshima H, Oshima M, Ikeda T, Asaba R, Yagi H, et al:
CD44 variant regulates redox status in cancer cells by stabilizing
the xCT subunit of system xc(−) and thereby promotes tumor growth.
Cancer Cell. 19:387–400. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schieber MS and Chandel NS: ROS links
glucose metabolism to breast cancer stem cell and EMT phenotype.
Cancer Cell. 23:265–267. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rycaj K and Tang DG: Cancer stem cells and
radioresistance. Int J Radiat Biol. 90:615–621. 2014. View Article : Google Scholar : PubMed/NCBI
|