Introduction
Chronic myelogenous leukemia (CML) is a clonal
myeloproliferative disorder that is derived from abnormal
pluripotent bone marrow hematopoietic stem cells, characterized by
the Philadelphia chromosome and/or the breakpoint cluster region
protein (BCR)-Abelson murine leukemia viral oncogene homolog 1
(ABL) fusion gene (1). Imatinib
(Gleevec), a small-molecule BCR-ABL tyrosine kinase inhibitor
(TKI), markedly improved the estimated overall survival rate in
patients with chronic-phase (CP)-CML to between 80 and 85% at 7–10
years (2,3). However, although imatinib eradicated
mature BCR-ABL+ CML cells, it was not able to achieve
similar effects in CML leukemia stem cells (LSCs) since BCR-ABL
tyrosine kinase is dispensable for CML LSC survival and
maintenance, rendering these cells able to survive in the presence
of TKIs and to eventually promote relapse (4,5).
Therefore, the elucidation of the biological characteristics of the
LSC is required.
Microvesicles (MVs), lipid-bilayer vesicles formed
by directly budding off from the cell membrane, represent a
heterogeneous population of vesicles with a diameter of between 100
and 1,000 nm (6,7). MVs are able to selectively package
complex biological information from parental cells, including mRNA,
microRNA (miRNA) and proteins, which serve an important role in
intercellular communication (8).
Previous studies have suggested that MVs are involved in
inflammation, immune regulation, tumor metastasis and angiogenesis,
and may also be indicators for the diagnosis and prognosis of
disease (9–12).
miRNAs, a large family of small (between 22 and 24
nucleotides in length) non-coding RNAs, decrease gene expression
levels by regulating the translation of certain mRNAs into protein
and serve an important role in disease progression and
carcinogenesis. An increasing number of studies have revealed that
genetic exchange of miRNA between cells may be accomplished through
MVs (13). For instance, MVs derived
from endothelial progenitor cells protect the kidneys from acute
ischemic injury by miRNA-dependent reprogramming of resident renal
cells (14). Additionally, embryonic
stem cell-derived MVs may be useful therapeutic tools for
transferring miRNA to cells and important mediators of signaling
within stem cell niches (15).
Although a number of cell-derived MVs are currently recognized and
studied, MVs derived from primary CML LSCs remain unknown, and
there is a lack of comprehensive information concerning miRNA from
LSC-secreted MVs.
In the present study, CML LSCs were studied and MVs
secreted from LSCs were characterized in terms of surface markers
and miRNA profiles. Furthermore, the miRNA profiles in MVs of CML
blasts were compared with those of the AML stem cell line KG-1a.
The results of the present study revealed that these miRNAs were
primarily associated with MVs and stem cells, and may affect
characteristics of LSCs.
Materials and methods
Patient samples
Fresh peripheral blood (PB) or bone marrow (BM)
samples were obtained from healthy donors and patients with CML
(patient characteristics are listed in Table I). PB cells were also acquired from
leftover material from PB transplant harvests of healthy donors.
Written informed consent was obtained and the present study was
approved by the Ethics Committee of Union Hospital, Tongji Medical
College, Huazhong University of Science and Technology (Wuhan,
China). Mononuclear cells were extracted by Ficoll-Paque density
gradient centrifugation (Lymphoprep™, 1.073 g/ml; Tianjinhaoyang
Biological Manufacture Co., Ltd, Tianjin, China) according to the
manufacturer's protocol.
 | Table I.Characteristics of patients with
CML. |
Table I.
Characteristics of patients with
CML.
| Patient | Sex | Age, years | Diagnosis | White blood cell
count (g/l) | Treatment |
|---|
| CP-CML 1 | F | 45 | CP-CML | 111.70 | De novo |
| CP-CML 2 | M | 27 | CP-CML | 262.26 | De novo |
| CP-CML 3 | M | 39 | CP-CML | 289.50 | De novo |
| CP-CML 4 | M | 50 | CP-CML | 236.57 | De novo |
| CP-CML 5 | M | 81 | CP-CML | 366.99 | De novo |
| CP-CML 6 | F | 29 | CP-CML | 196.20 | De novo |
| CP-CML 7 | F | 47 | CP-CML | 269.60 | De novo |
| CP-CML 8 | M | 53 | CP-CML | 198.20 | De novo |
| BC-CML 1 | F | 56 | BC-CML |
16.80 | IFN, IM |
| BC-CML 2 | F | 52 | BC-CML |
30.72 | IM, Dasa |
| BC-CML 3 | M | 47 | BC-CML |
42.40 | IM |
| BC-CML 4 | M | 43 | BC-CML |
35.20 | De novo |
Cluster of differentiation (CD)34+ cell
isolation and cell culture. CD34+ cells (>92% pure)
were isolated using immunomagnetic separation by positive selection
of cells (human CD34 microbead kit; Miltenyi Biotec GmbH, Bergisch
Gladbach, Germany), according to the manufacturer's protocol.
Primary CD34+ cells were cultured in StemSpan serum-free
medium (Stemcell Technologies, Inc., Vancouver, BC, Canada) which
was supplemented with a growth factor mixture, including 100 ng/ml
each of Fms-related tyrosine kinase-3 ligand and stem cell factor,
and 20 ng/ml each of interleukin (IL)-3, IL-6 and
granulocyte-colony stimulating factor (Peprotech, Inc., Rocky Hill,
NJ, USA). Primary cells were counted using a hemocytometer and
subsequently cultured at density of 5×104 cells/ml.
After 48 h, cells were collected for flow cytometry or miRNA
experiments and culture medium was used for MV isolation. The KG-1a
cells, a human acute myelogenous leukemia (AML) cell line, were
stored long-term and passaged in the Institute of Hematology, Union
Hospital, Tongji Medical College, Huazhong University of Science
and Technology (Wuhan, China). The KG-1a cell line, with ~95% CD34
expression, was routinely cultured in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and supplemented
with 10% fetal bovine serum (FBS); FBS-derived MVs were removed
using a differential centrifugation method, including 750 × g for
15 min, followed by 1,500 × g for 20 min and then 16,000 × g for 45
min, at 4°C. The cells were cultured at 37°C in a humidified
atmosphere containing 5% CO2. The culture medium was
collected and replaced with fresh medium every 48 h.
Preparation of MVs
MVs were isolated from the conditioned medium of
CD34+ cells and KG-1a cells by continuous differential
centrifugation as described previously (9). At the time of culture medium harvest,
MVs were prepared by differential centrifugation as described in
the previous section. Pellets of MVs were washed in ice-cold PBS
(particles <0.1 µm were removed with a membrane filter prior to
use) for flow cytometric analysis.
Flow cytometric analysis
To analyze CD123 expression, CD34+ cells
were stained with anti-CD34-phycoerythrin (PE) antibody (1:50; cat.
no., 130-098-140; Miltenyi Biotec GmbH) and anti-CD123-PE-cyanine 7
antibody (1:50; cat. no., 306009; BioLegend, Inc., San Diego, CA,
USA) per tube. A previously described method (16) was utilized to analyze the phenotype of
MVs. In brief, standard microbeads with a diameter of 1 or 3 µm
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) were used to set
the upper size limit for MVs. MVs isolated from a 10 ml supernatant
of CD34+ cell-conditioned medium were resuspended in PBS
(extra particles with 0.1 µm membrane filter were removed prior to
use) and then stained with calcein-acetoxymethyl ester (AM) (5
µg/ml; cat. no., 65-0853-39; Thermo Fisher Scientific, Inc.), CD34
and CD123 for 20 min at ambient temperature. The stained MVs were
diluted in 300 µl PBS and analyzed using a FACSAria II flow
cytometer with FACSDiva version 6.1.3 software (BD Pharmingen, San
Diego, CA, USA). MVs were defined as calcein-AM-positive
events.
miRNA expression profiling
miRNA expression profiles were conducted on 4
samples [CP-CML CD34+ cells (C-C); CP-CML
CD34+ cell-derived MVs (C-MV); KG-1a cells (K-C); KG-1a
cell-derived MVs (K-MV)] using miRCURY™ locked nucleic acid (LNA)
array (7th generation, version 18.0; Exiqon A/S, Vedbaek, Denmark)
containing probes for 3,100 miRNAs. Total RNA was isolated using
TRIzol (Thermo Fisher Scientific, Inc.) and purified using an
RNeasy mini kit (Qiagen GmbH, Hilden, Germany) according to the
manufacturer's protocol. RNA was quantified using a NanoDrop
spectrophotometer (ND-1000; NanoDrop Technologies; Thermo Fisher
Scientific, Inc.) and RNA integrity was determined by gel
electrophoresis. Following RNA extraction from the samples, miRNAs
were labelled using the miRCURY™ Hy3™/Hy5™ Power Labeling kit
(Exiqon A/S), according to the manufacturer's protocol.
Subsequently, the Hy3™-labeled samples were hybridized on the
miRCURY™ LNA array. Following hybridization, the slides were washed
five times with the Wash Buffer kit (Exiqon A/S) and dried by
centrifugation at 200 × g for 5 min. The slides were scanned using
the Axon GenePix 4000B microarray scanner (Axon Instruments;
Molecular Devices, LLC, Sunnyvale, CA, USA). Scanned images were
then imported into GenePix Pro 6.0 software (Axon Instruments;
Molecular Devices, LLC) for grid alignment and data extraction. The
4 replicated spots for each probe were averaged. Expressed results
were normalized using the median normalization method
(foreground-background signal intensity)/median of samples with
intensity ≥30) and subsequently identified differentially expressed
(DE) miRNAs between two samples were filtered by fold change
(threshold, ≥2.0).
Validation of microarray results
A total of 4 DE miRNAs identified by miRNA
microarray were selected for further validation by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR). RNA
(400 ng) was reverse transcribed with a reaction mix of of dNTPs
(HyTest Ltd., Turku, Finland), RT buffer, MMLV reverse
transcriptase, RNase inhibitor (all from Epicentre; Illumina, Inc.,
San Diego, CA, USA), specific primers (Balige Co., Shanghai, China)
and RNAase-free water to 20 µl. The reaction was performed at 16°C
for 30 min, followed by a 40-min incubation at 42°C and finally
85°C for 5 min using an Applied Biosystems 9700 Real-Time PCR
instrument (Thermo Fisher Scientific, Inc.).
SYBR Green miRNA assays with commercially available
primers (Table II) for
hsa-miR-627-5p, hsa-miR-483-5p, hsa-miR-638 and hsa-miR-1290 (all
materials from Guangzhou RiboBio Co., Ltd., Guangzhou, China) were
used according to the manufacturer's protocol. Relative expression
was calculated with the 2−∆∆Cq method with an Applied
Biosystems ViiA 7 Real-Time PCR system (Thermo Fisher Scientific,
Inc.) (17).
| Table II.Primers used in Real-time PCR. |
Table II.
Primers used in Real-time PCR.
| Gene | Primer
sequence |
|---|
| hsa-miR-483-5p |
| F |
5′-AGGGAAGACGGGAGAAGAGA-3′ |
| R |
5′-GTGCGTGTCGTGGAGTCG-3′ |
| hsa-miR-627-5p |
| F |
5′-GGGGGTGAGTCTCTAAGAAA-3′ |
| R |
5′-CAGTGCGTGTCGTGGAGT-3′ |
| hsa-miR-1290 |
| F |
5′-GGGGTGGATTTTTGGAT-3′ |
| R |
5′-CAGTGCGTGTCGTGGAGT-3′ |
| hsa-miR-638 |
| F |
5′-AAGGATCGCGGGCGGGT-3′ |
| R |
5′-GTGCGTGTCGTGGAGTCG-3′ |
Target gene prediction
TargetScan, miRBase and miRanda databases were used
to predict the target genes of DE miRNAs. Among the putative
targets, the overlapping genes from these databases were
determined.
Bioinformatics and statistical
analysis
Gene Ontology (GO) analysis and pathway annotation
were used to examine the gene pool of DE miRNAs. GO terms were
analyzed using the GO database. Fisher's exact test was used to
identify any overlap between the DE list and the GO annotation list
greater than was expected by chance. The P-value denoted the
statistical significance of GO term enrichment in the DE genes.
Pathway analysis is a functional analysis mapping genes to Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathways. The P-value
denoted the statistical significance of the pathway associated with
the conditions. P<0.05 was considered to indicate a
statistically significant difference.
Results
CML CD34+ blasts and
blast-derived-MVs express increased levels of CD123
A previous study demonstrated that IL-3 receptor α
(CD123) expression is elevated in CML progenitor and stem cells
compared with healthy donors (18),
which led to the collection of samples from 4 patients with blast
crisis (BC)-CML, 8 patients with CP-CML and 4 cases of healthy
donor stem cell residue in the present study. Multicolor flow
cytometry was used to determine the surface marker profile of CML
CD34+ cells and cell-derived MVs. The results of the
present study indicated that, compared with healthy
CD34+ cell samples, CD123 expression levels of BC-CML
CD34+ cells were significantly increased. There was also
a marked difference between BC-CML and CP-CML CD34+
cells, as presented in Fig. 1:
Compared with MVs from healthy CD34+ samples, CD123
expression levels of BC-CML CD34+ cell-derived MVs were
significantly increased, whereas expression levels of CP-CML
CD34+ cell-derived MVs slightly increased. As presented
in Fig. 2, BC-CML and CP-CML
CD34+ cell-derived MVs also demonstrated significant
differences. The results of the present study revealed that the
expression levels of CD123 in CD34+ cells and MVs
gradually increased in association with disease progression.
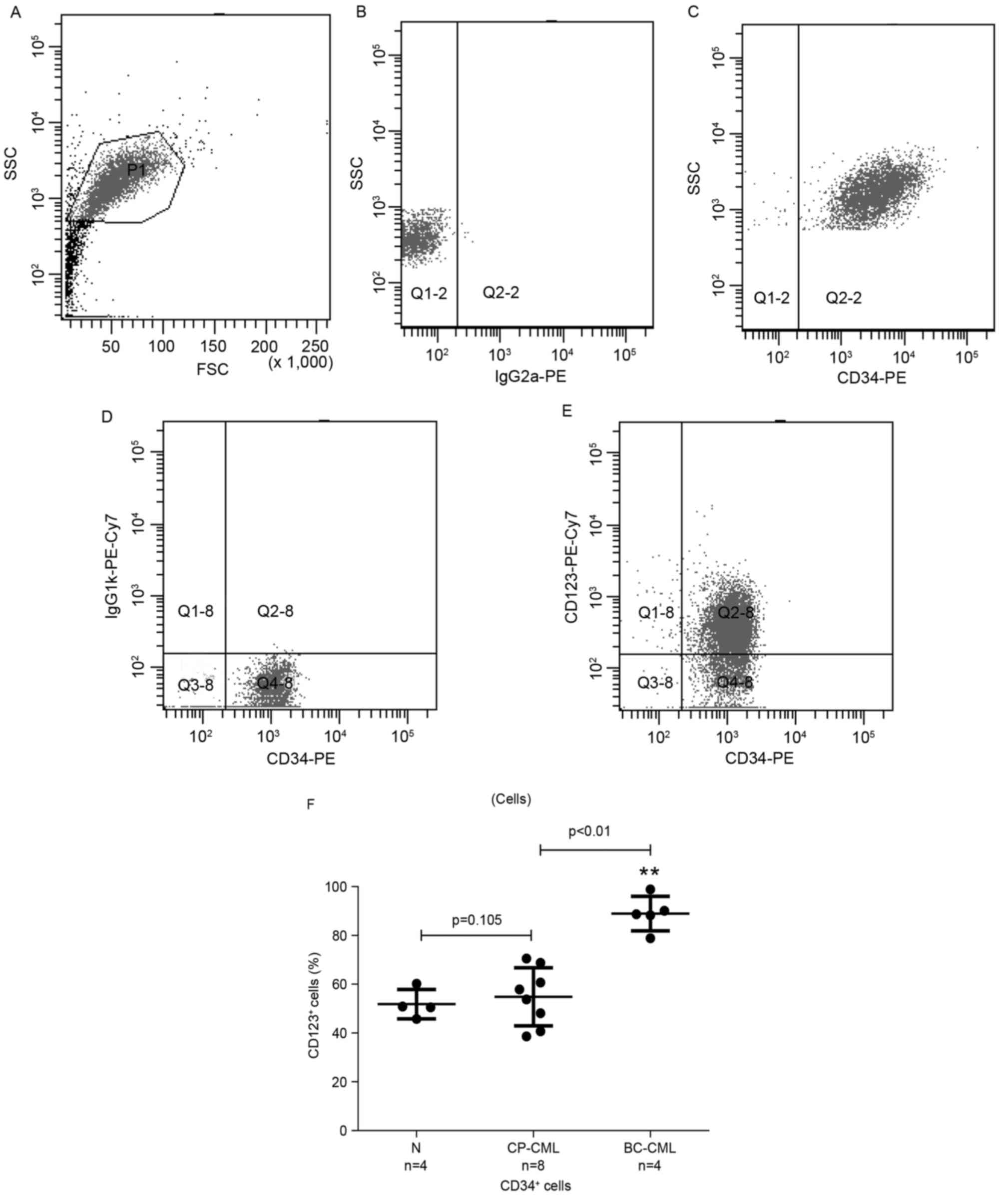 | Figure 1.CD123 expression is significantly
increased in CD34+ cells of patients with CP-CML and
BC-CML, compared with healthy donors. The gating strategy applied
to determine the proportion of CD123+ cells in
CD34+ subsets was: (A) P1, active cells; (B) isotype
control; (C) Q2-2, CD34+ blasts; (D) isotype control;
(E) Q2-8, CD34+ CD123+ blasts. (F) Proportion
of CD123+ cells within CD34+ populations from
healthy donors, and patients with CP-CML and BC-CML determined
using multicolor flow cytometry. Each dot represents an individual
tissue sample. P=0.105, N vs. CP-CML; **P<0.01, CP-CML vs.
BC-CML. CD, cluster of differentiation; CP, chronic-phase; CML,
chronic myelogenous leukemia; BC, blast crisis; N, healthy donors;
SSC, side scatter; FSC, forward scatter; PE, phycoerythrin; Cy7,
cyanine 7; Q, quadrant; Ig, immunoglobulin. |
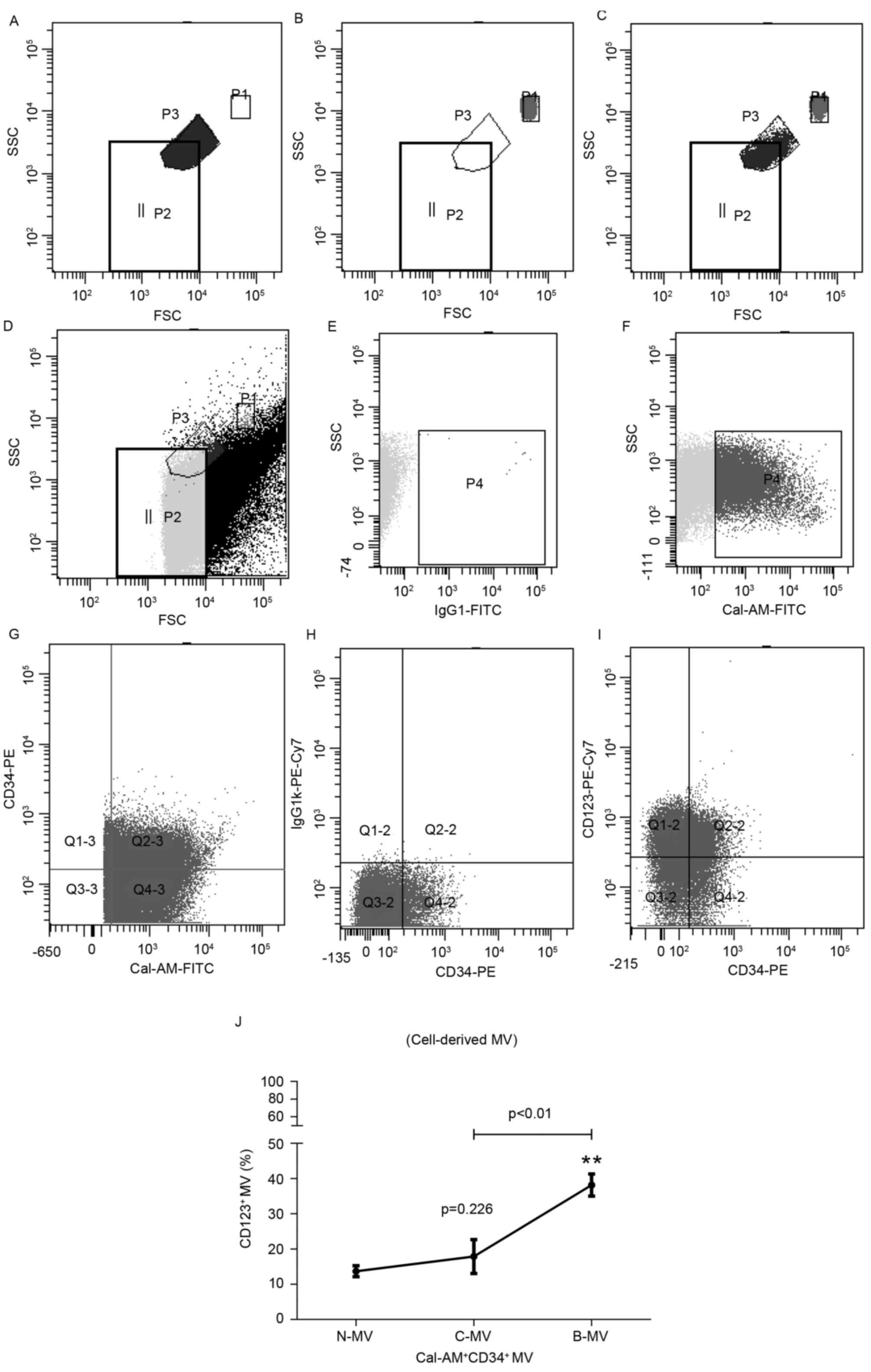 | Figure 2.CD123 expression is significantly
increased in CD34+ blast-derived MVs of patients with
CP-CML and BC-CML compared with healthy donors. Flow cytometric
analysis and representative cytograms of
CD123+CD34+ blast-derived MVs in patients
with CML. (A) Fluorosphere beads of a known size were used as a
standard to set the MV gate. P3 represents 1 µm beads. (B) P1
represents 3 µm beads. (C) P1 and P3 represent 3 µm and 1 µm beads,
respectively. (D) Based on the calibrating beads above, P2
represents the MV gate. (E) IgG1-FITC was set as the negative
control for calcein-AM+ MVs. (F) Events in the MV gate
(P4) were analyzed to differentiate calcein-AM+ MVs from
the background signal. (G) Q2-3,
calcein-AM+CD34+ MVs; (H) CD123 isotype
control, Q2-2,
calcein-AM+CD34+CD123− MVs. (I)
Q2-2, calcein-AM+CD34+CD123+ MVs.
(J) The proportion of CD123+ MV within
calcein-AM+CD34+ populations from N-MV and
patients with C-MV and B-MV was determined using multicolor flow
cytometry. P=0.226, N-MV vs. C-MV; **P<0.01, C-MV vs. B-MV. CD,
cluster of differentiation; CP, chronic-phase; CML, chronic
myelogenous leukemia; BC, blast crisis; MV, microvesicle; N-MV,
healthy; C-MV, chronic-phase CML; B-MV, blast crisis CML; SSC, side
scatter; FSC, forward scatter; PE, phycoerythrin; Cy7, cyanine 7;
Q, quadrant; Ig, immunoglobulin; FITC, fluorescein isothiocyanate;
Cal-AM, calcein acetoxymethyl ester. |
Comparison of MV-derived and
cell-derived miRNA profiles
To validate the hypothesis that miRNAs in the MVs
enabled communication and reflected genetic changes within LSCs,
miRNA profiles of the MVs extracted from CML CD34+ blast
cells and KG-1a were determined. It was hypothesized that there
were differences in the miRNA expression levels between MVs and
LSCs, therefore the RNA of CML blasts and KG-1a cells was also
extracted and hybridized (CML CD34+ blast cells, CML
blast-derived MV, KG-1a cells and KG-1a-derived MV, n=2). Each
sample was analyzed in duplicate under similar conditions. DE
miRNAs were screened using the following criteria: Normalized
intensity of each miRNA >30, fold changes of
C-MV/CD34+ cells and K-MV/KG-1a >2.0. The present
study identified that 15 miRNAs were elevated in C-MV and K-MV by
setting screening criteria (Table
III). The 5 most significantly upregulated miRNAs were human
(hsa)-miR-4732-5p, miR-1290, hsa-miR-4750-5p, hsa-miR-1908-5p and
hsa-miR-483-5p, suggesting that these MV miRNAs serve an important
role in CD34+ blasts.
| Table III.miRNAs increased in CD34+
blast-derived MVs and KG-1a cell-derived MVs. |
Table III.
miRNAs increased in CD34+
blast-derived MVs and KG-1a cell-derived MVs.
| miRNA | C-MV/C-C, fold
change | K-MV/K-C, fold
change |
|---|
|
hsa-miR-4732-5p | 3.15 | 20.74 |
| hsa-miR-1290 | 3.13 |
8.33 |
|
hsa-miR-4750-5p | 2.92 |
5.46 |
|
hsa-miR-1908-5p | 2.78 |
5.26 |
| hsa-miR-483-5p | 2.63 |
6.00 |
| hsa-miR-638 | 2.58 |
4.65 |
| hsa-miR-3960 | 2.58 |
3.23 |
| hsa-miR-4516 | 2.44 | 12.97 |
| hsa-miR-1469 | 2.42 |
5.66 |
| hsa-miR-4467 | 2.41 |
4.12 |
|
hsa-miR-4707-5p | 2.20 |
7.75 |
| hsa-miR-4285 | 2.13 |
7.43 |
|
hsa-miR-4787-5p | 2.12 |
3.90 |
|
hsa-miR-4708-3p | 2.06 |
4.73 |
| hsa-miR-627-5p | 2.04 |
2.65 |
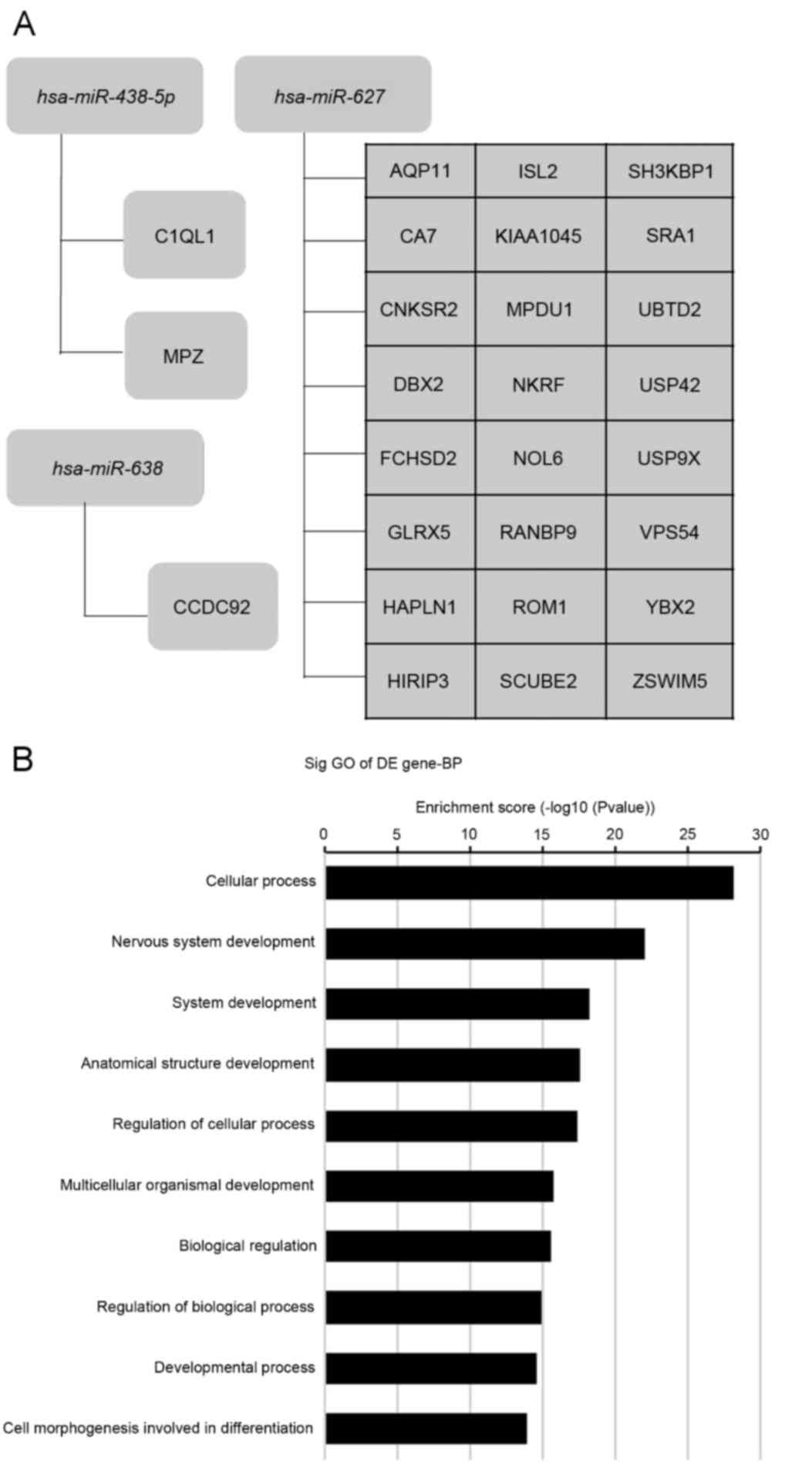
miRNA-target genes and GO terms
In order to investigate the significantly
dysregulated MV miRNAs derived from CD34+ blasts,
potential target analysis was performed. Using TargetScan, miRanda
and miRBase, it was determined that 9,036, 2,412 and 2,291 target
gene prediction records were identified, respectively, for the 15
overexpressed miRNAs. A total of 27 target gene results were
screened through overlapping each dataset. These results suggested
that these miRNAs affect various cellular biological processes
including transcription, metabolism, molecular signaling pathway,
proliferation, differentiation and apoptosis via the regulation of
these target genes. These miRNAs may additionally be involved in
tumorigenesis. Fig. 3A presents a
list of associated genes targeted by DE miRNAs.
GO terms may be assigned to the
potential targets
In order to understand the function of the involved
genes, GO terms were divided into three groups including molecular
function (MF), biological process (BP) and cellular component (CC).
In the present study, focus was placed on the BP function of the
involved genes. Fig. 3B presents
significantly distinct GO terms of DE gene-related biological
processes. The results suggested that the miRNAs of blast-derived
MVs were associated with cell metabolism, cell cycle and cell
adhesion.
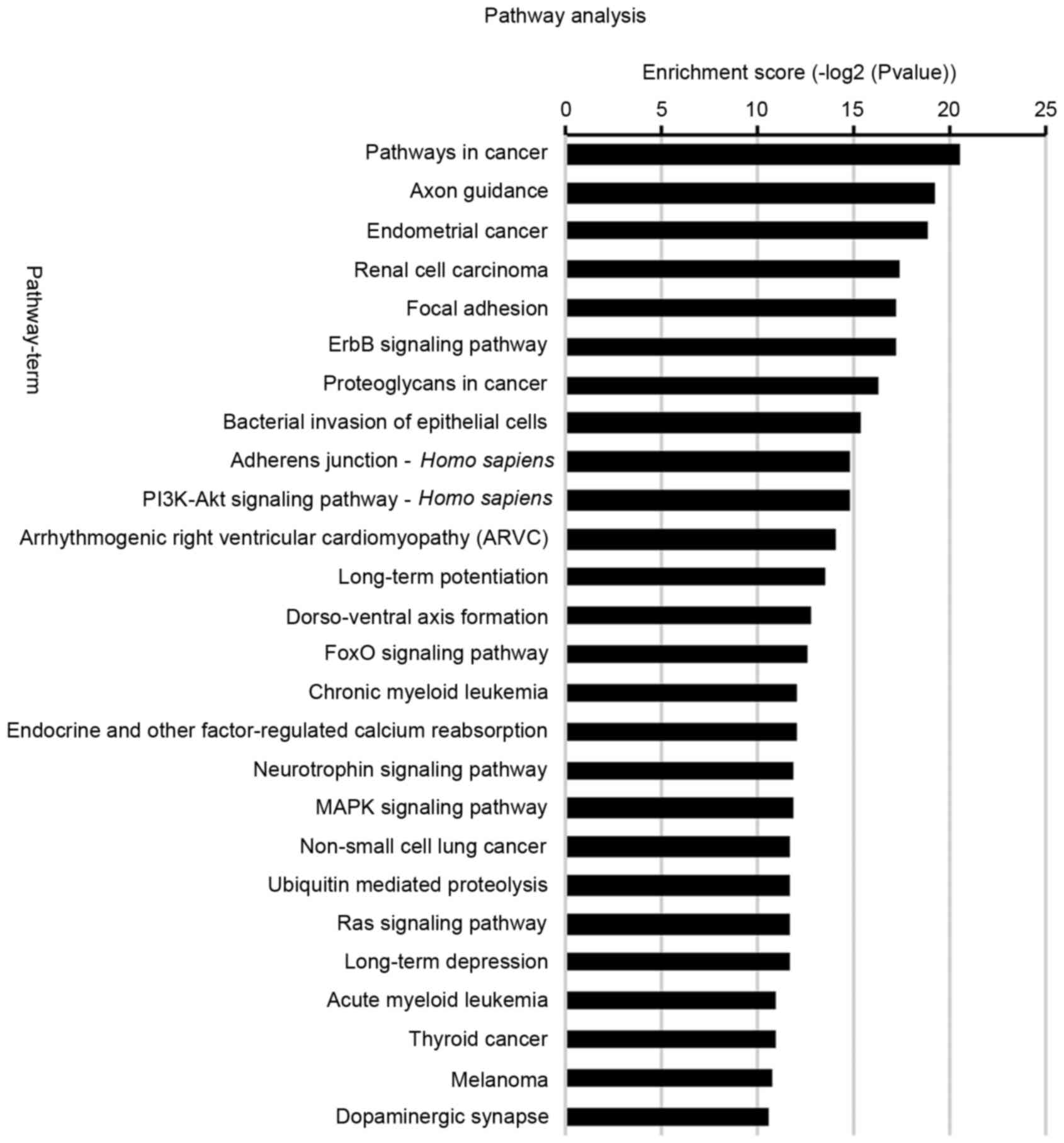
KEGG pathway
The activation and inactivation of certain
intracellular signaling pathways serve critical roles in tumor stem
cell biology. To better understand the function of potential
targets, signaling pathways were analyzed by KEGG (26 signaling
pathways; Fig. 4). A total of 65
distinct pathways with enrichment test P<0.01 were identified
according to the KEGG pathway database. The pathways associated
with the upregulated miRNAs in MVs (P<0.05) included those in
cancer, CML and AML, in addition to ErbB signaling and the
phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling
pathway. The signaling pathways identified were primarily
associated with regulation of cell viability, apoptosis, metabolism
and tumorigenesis.
RT-qPCR
In order to validate the microarray results, RT-qPCR
was performed to determine the upregulated miRNAs. A total of 4
miRNAs were selected for RT-qPCR: Hsa-miR-627-5p, hsa-miR-483-5p,
hsa-miR-638 and hsa-miR-1290 (Fig.
5). Similar results were revealed in the DE miRNAs as in the
miRNA microarray.
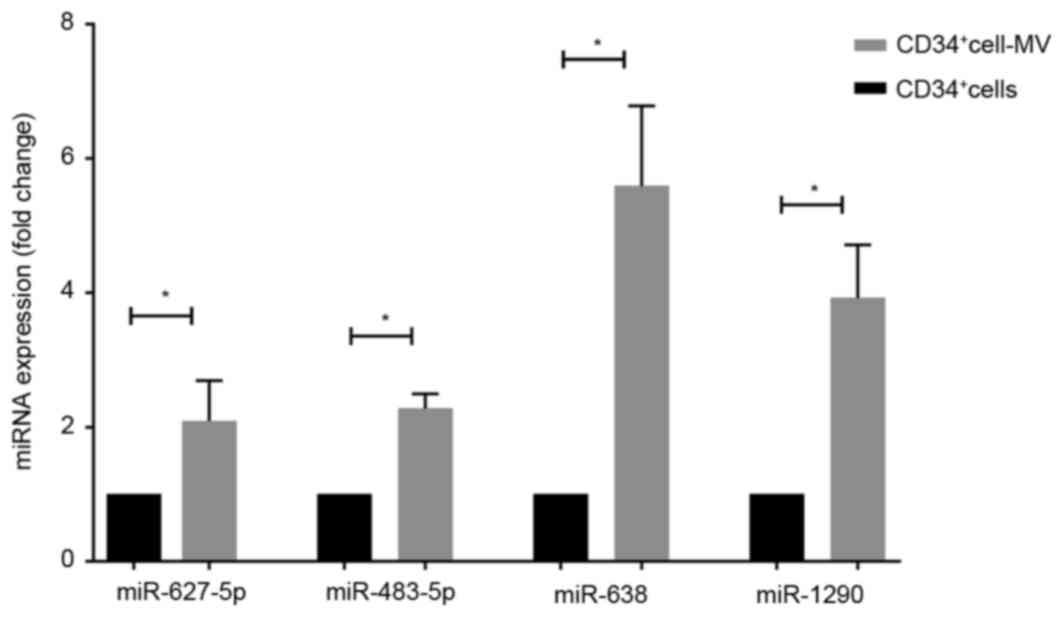 | Figure 5.Validation of miRNA expression levels
using the reverse transcription-quantitative polymerase chain
reaction. Expression levels of miR-627-5p, miR-483-5p, miR-638 and
miR-1290 in an independent set of primary CD34+ blasts
and blast-derived MVs were determined. miRNA expression in blasts
was set as 1; the relative expression of hsa-miR-627-5p,
hsa-miR-483-5p, hsa-miR-638 and hsa-miR-1290 in MVs was 2.09±0.35,
2.14±0.01, 5.59±0.69 and 3.92±0.46, respectively. Results are
presented as the mean ± standard deviation. *P<0.05. miRNA,
microRNA; CD, cluster of differentiation. MV, microvesicle; hsa,
human. |
Discussion
Despite the second-generation TKIs markedly
improving the outcome of patients with CML, the remaining LSCs
cause disease recurrence in patients with CML (19). The biological characteristics of LSCs
are quiescence, multi-differentiation and self-renewal (3); however, current strategies have
difficulty accessing and eradicating LSCs (20). Similarly to the results of the present
study, it had been previously demonstrated that cells may
selectively package miRNAs into MVs, which are secreted under
various pathophysiological conditions, and the content of MVs is
considerable (21,22). Our previous study identified that
BCR-ABL1 mRNA remains detectable within MVs, although the
intracellular copy is 0 (strict complete molecular remission)
(23). It was revealed that when
maternal cells remain in the bone marrow niche, MVs can be detected
and may magnify characteristics of maternal cells in the peripheral
blood. Consequently, the aim of the present study was to elucidate
the characteristics and molecular profiles of MVs released from
specific CD34+ blasts in patients with CML.
By optimizing flow cytometric analysis, the results
of the present study demonstrated that CD34 and CD123 were
selectively packaged on MVs from CD34+ blasts. In
addition, the expression of CD123 in CD34+ cells and MVs
increased gradually along with disease progression. Thus,
CD34+ cell-derived MVs may be a predictor of disease
progression and may be used as a minimal residual disease marker. A
previous study has revealed that CD123 is a specific surface marker
of acute and chronic leukemia stem and progenitor cells (18). The present study may provide novel
perspectives for the study of the biological characteristics of
LSCs and the surface marker of MVs may provide the basis for the
sorting of the MV subpopulation.
In the present study, it was hypothesized that
miRNAs were contained in MVs, which enabled communication and
influenced genetic changes within patients with CML. miRNA
expression patterns were determined in primary CD34+
cells and MV purified from CD34+ cells from patients
with CML. miRNAs in KG-1a cells and the associated MVs were also
determined, for comparison. The results of the present study
revealed that 15 miRNAs were significantly increased in MVs with
respect to corresponding cells. Target gene prediction was
performed to further determine the function of miRNAs. GO and
pathway analysis demonstrated that the upregulated miRNAs in MVs
may be associated with cell development, morphology,
differentiation, metabolism and cell cycle of CD34+
blasts. KEGG pathway demonstrated that upregulated miRNAs may be
involved in tumorigenesis, chronic and acute myeloid leukemia and
signaling pathways including ErbB, PI3K/Akt and forkhead box O
(FOXO).
Of the 15 upregulated miRNAs in MVs, miR-1290 was
the primary modulator and modulates 4,019 target genes. The targets
of miR-1290, including B-cell lymphoma/leukemia (Bcl)2, lysine
methyltransferases, tumor protein p63-regulated 1, chronic
lymphocytic leukemia upregulated (CLLU) 1 and Bcl tumor suppressor
7A, were identified to be involved in multiple cancers. The Bcl2
gene is a proto-oncogene and inhibits apoptosis. Bcl2 serves an
important role in the pathogenesis of breast cancer and increased
expression levels of Bcl2 in breast cancer indicates a poor
prognosis (24). CLLU1, located on
chromosomal locus 12q22, encodes chronic lymphocytic leukemia
(CLL)-specific transcripts and patients with increased expression
levels of CLLU1 usually exhibit decreased progression-free survival
and overall survival times (25).
Huang et al (26) hypothesized
that exosomes may be selectively enriched with miR-1290 and
miR-375, which allow exosomes to be prognostic markers for advanced
prostate cancer. With the exception of hsa-miR-1290, other
upregulated miRNAs were also predicted to serve important roles in
cancer. For example, miR-638 may be stably detected in human
plasma, and the miR-92/miR-638 ratio in plasma may be a useful
indicator to distinguish between patients with leukemia and the
healthy control group (27). Jaiswal
et al (28) identified that
miR-1246, miR-1308, miR-638 and other human miRNAs may be
selectively enriched in microparticles, transferring them to the
recipient cells. Similarly, the results of the present study
identified that MVs, which cannot be distinguished from
microparticles at present, package miR-638. In addition,
upregulated miRNAs in MVs were identified to be poor prognostic
tumor markers, which is consistent with a previous study (29), which revealed that miR-483-5p and
miR-195 were associated with poor prognosis in adrenocortical
adenoma. Thus, it can be hypothesized that CD34+ blasts
release MVs which contain elevated miRNAs and may be beneficial for
the survival of blasts themselves; however, this requires
additional study. Following GO analysis, KEGG was used to analyze
the pathways which involved the predicted miRNA target genes. ErbB,
PI3K/Akt and FOXO signaling pathways were included. A previous
study demonstrated that these pathways serve a critical role in CML
(30) and Naka et al (31) identified that the transforming growth
factor β (TGFβ)/FOXO signaling pathway may be involved in
leukemia-initiating cells. Furthermore, in CML FOXO3a−/−
rat experiments, TGF-β inhibitor combined with imatinib was
demonstrated to be an effective treatment.
To the best of our knowledge, the present study is
the first to identify the DE miRNAs between CD34+ blasts
and blast-derived MVs in CML. Additionally, the present study
optimized the flow cytometric method of blast-derived MVs. Using
complex pathway analysis, distinct KEGG pathways and GO terms were
characterized by altered gene expression and miRNA regulation may
be identified. The results of the present study provide evidence
that may increase the understanding of physiological functions of
stem cell-derived MVs and the potential roles of CD34+
blast-derived MVs in CML-associated processes. However, the
interactions between miRNAs and their targets in the present study
are only bioinformatically predicted, and further study is required
to validate them.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81370659). The
authors thank the Key Laboratory for Molecular Diagnosis of Wuhan
Central Hospital for the detection and analysis of MVs.
References
|
1
|
Sawyers CL: Chronic myeloid leukemia. N
Engl J Med. 340:1330–1340. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kantarjian H and Cortes J: Considerations
in the management of patients with Philadelphia chromosome-positive
chronic myeloid leukemia receiving tyrosine kinase inhibitor
therapy. J Clin Oncol. 29:1512–1516. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
O'Hare T, Zabriskie MS, Eiring AM and
Deininger MW: Pushing the limits of targeted therapy in chronic
myeloid leukaemia. Nat Rev Cancer. 12:513–526. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morotti A, Panuzzo C, Fava C and Saglio G:
Kinase-inhibitor-insensitive cancer stem cells in chronic myeloid
leukemia. Expert Opin Biol Ther. 14:287–299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Crews LA and Jamieson CH: Selective
elimination of leukemia stem cells: Hitting a moving target. Cancer
Lett. 338:15–22. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Raposo G and Stoorvogel W: Extracellular
vesicles: Exosomes, microvesicles, and friends. J Cell Biol.
200:373–383. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van der Pol E, Boing AN, Harrison P, Sturk
A and Nieuwland R: Classification, functions, and clinical
relevance of extracellular vesicles. Pharmacol Rev. 64:676–705.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen X, Liang H, Zhang J, Zen K and Zhang
CY: Secreted microRNAs: A new form of intercellular communication.
Trends Cell Biol. 22:125–132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hong CS, Muller L, Whiteside TL and
Boyiadzis M: Plasma exosomes as markers of therapeutic response in
patients with acute myeloid leukemia. Front Immunol. 5:1602014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mfonkeu JB Pankoui, Gouado I, Kuaté H
Fotso, Zambou O, Zollo PH Amvam, Grau GE and Combes V: Elevated
cell-specific microparticles are a biological marker for cerebral
dysfunctions in human severe malaria. PLoS One. 5:e134152010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haflidadóttir BS and Ceder Y: Exosomal
microRNAs as potential biomarkers in castration-resistant prostate
cancer. Eur Urol. 67:42–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Belov L, Matic KJ, Hallal S, Best OG,
Mulligan SP and Christopherson RI: Extensive surface protein
profiles of extracellular vesicles from cancer cells may provide
diagnostic signatures from blood samples. J Extracell Vesicles.
5:253552016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moldovan L, Batte K, Wang Y, Wisler J and
Piper M: Analyzing the circulating microRNAs in
exosomes/extracellular vesicles from serum or plasma by qRT-PCR.
Methods Mol Biol. 1024:129–145. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cantaluppi V, Gatti S, Medica D,
Fiqliolini F, Bruno S, Dereqibus MC, Sordi A, Biancone L, Tetta C
and Camussi G: Microvesicles derived from endothelial progenitor
cells protect the kidney from ischemia-reperfusion injury by
microRNA-dependent reprogramming of resident renal cells. Kidney
Int. 82:412–427. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan A, Farber EL, Rapoport AL, Tejada D,
Deniskin R, Akhmedov NB and Farber DB: Transfer of microRNAs by
embryonic stem cell microvesicles. PLoS One. 4:e47222009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun L, Wang HX, Zhu XJ, Wu PH, Chen WQ,
Zou P, Li QB and Chen ZC: Serum deprivation elevates the levels of
microvesicles with different size distributions and selectively
enriched proteins in human myeloma cells in vitro. Acta Pharmacol
Sin. 35:381–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T))method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nievergall E, Ramshaw HS, Yong AS, Biondo
M, Busfield SJ, Vairo G, Lopez AF, Hughes TP, White DL and Hiwase
DK: Monoclonal antibody targeting of IL-3 receptor α with CSL362
effectively depletes CML progenitor and stem cells. Blood.
123:1218–1228. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Copland M, Hamilton A, Elrick LJ, Baird
JW, Allan EK, Jordanides N, Barow M, Mountford JC and Holyoake TL:
Dasatinib (BMS-354825) targets an earlier progenitor population
than imatinib in primary CML but does not eliminate the quiescent
fraction. Blood. 107:4532–4539. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou H and Xu R: Leukemia stem cells: The
root of chronic myeloid leukemia. Protein Cell. 6:403–412. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He WA, Calore F, Londhe P, Canella A,
Guttridge DC and Croce CM: Microvesicles containing miRNAs promote
muscle cell death in cancer cachexia via TLR7. Proc Natl Acad Sci
USA. 111:pp. 4525–4529. 2014; View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Camussi G, Deregibus MC, Bruno S,
Cantaluppi V and Biancone L: Exosomes/microvesicles as a mechanism
of cell-to-cell communication. Kidney Int. 78:838–848. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu XJ, Li QB, Zeng C, Zhong Z, You Y and
Zou P: Detection of leukemia-derived microparticles in the
monitoring of chronic myeloid leukemia. Zhonghua Xue Ye Xue Za Zhi.
35:138–141. 2014.(In Chinese). PubMed/NCBI
|
|
24
|
Bouchalova K, Svoboda M, Kharaishvili G,
Vrbkova J, Bouchal J, Trojanec R, Koudelakova V, Radova L, Cwiertka
K, Hajduch M and Kolar Z: BCL2 is an independent predictor of
outcome in basal-like triple-negative breast cancers treated with
adjuvant anthracycline-based chemotherapy. Tumour Biol.
36:4243–4252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gonzalez D, Else M, Wren D, Usai M, Buhl
AM, Parker A, Oscier D, Morgan G and Catovsky D: CLLU1 expression
has prognostic value in chronic lymphocytic leukemia after
first-line therapy in younger patients and in those with mutated
IGHV genes. Haematologica. 98:274–278. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang X, Yuan T, Liang M, Du M, Xia S,
Dittmar R, Wang D, See W, Costello BA, Quevedo F, et al: Exosomal
miR-1290 and miR-375 as prognostic markers in castration-resistant
prostate cancer. Eur Urol. 67:33–41. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pallasch CP, Patz M, Park YJ, Hagist S,
Eggle D, Claus R, Debey-Pascher S, Schulz A, Frenzel LP, Claasen J,
et al: miRNA deregulation by epigenetic silencing disrupts
suppression of the oncogene PLAG1 in chronic lymphocytic leukemia.
Blood. 114:3255–3264. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jaiswal R, Luk F, Gong J, Mathys JM, Grau
GE and Bebawy M: Microparticle conferred microRNA
profiles-implications in the transfer and dominance of cancer
traits. Mol Cancer. 11:372012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Soon PS, Tacon LJ, Gill AJ, Bambach CP,
Sywak MS, Campbell PR, Yeh MW, Wong SG, Clifton-Bligh RJ, Robinson
BG and Sidhu SB: miR-195 and miR-483-5p identified as predictors of
poor prognosis in adrenocortical cancer. Clin Cancer Res.
15:7684–7692. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang WZ, Pu QH, Lin XH, Liu MY, Wu LR, Wu
QQ, Chen YH, Liao FF, Zhu JY and Jin XB: Silencing of miR-21
sensitizes CML CD34+ stem/progenitor cells to imatinib-induced
apoptosis by blocking PI3K/AKT pathway. Leuk Res. 39:1117–1124.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Naka K, Hoshii T, Muraguchi T, Tadokoro Y,
Kondo Y, Nakao S, Motoyama N and Hirao A: TGF-beta-FOXO signalling
maintains leukaemia-initiating cells in chronic myeloid leukaemia.
Nature. 463:676–680. 2010. View Article : Google Scholar : PubMed/NCBI
|