Introduction
Bladder cancer is one of the most prevalent types of
cancer globally (1), with recurrence
rates as high as 67% in the first 12 months following treatment
(2). A substantial proportion
(16–25%) of urothelial cancers are invasive, infiltrating
high-grade carcinomas which will progress to metastasis (3), resulting in a poor 5-year survival
rate.
Intravesical chemotherapy effectively reduces and
delays tumor recurrence and progression (3). Drugs available at present for this
purpose include thiotepa, Adriamycin, epirubicin, doxorubicin (DOX)
and mitomycin C (MMC) (4). MMC has a
wide spectrum of antitumor activity (5,6).
Hydroxyca-mptothecin (HCPT) has a significant effect on the
proliferation and apoptosis of human bladder cancer and prostate
cancer cell lines in vitro (7,8). HCPT has
previously been demonstrated to have a marked effect on cell cycle
progression by forcing the cells into S arrest (9). However, due to the presence of cancer
stem cells in invasive bladder cancer, the majority of bladder
cancer cells demonstrate chemoresistance (10). Therefore, it is essential to identify
an effective chemosensitizer and multimodal strategy to treat
invasive bladder cancer.
Adenovirus, which is a well-characterized system
with deficient replication, has been widely used in cancer gene
therapy studies, including bladder cancer (11–14). Our
previous study constructed urothelium-specific recombinant
adenovirus type 5 [Ad5-UPII-adenoviral early region gene (E1A)]
with E1A (541 bp) and uroplakin II promoter (UPII promoter, 314
bp), which promoted the expression of the E1A gene and limited
replication of adenovirus to urothelial cells (15). Our previous studies demonstrated that
the oncolytic Ad5-UPII-E1A had a bladder cancer cell-specific
antitumor effect in vivo and in vitro (14–16).
Oncolytic adenovirus also functions as a chemosensitizer for
different antitumor mechanisms (17,18).
However, little is known about the anticancer effect of oncolytic
Ad5-UPII-E1A combined with MMC or HCPT.
Therefore, in the present study, the oncolytic
Ad5-UPII-E1A adenovirus was combined with MMC or HCPT to
investigate whether there was a synergistic inhibitory effect on
bladder cancer cell viability, and to screen for potential
mechanisms.
Materials and methods
Adenovirus vectors and cell lines
Urothelium-specific recombinant Ad5-UPII-E1A was
constructed with uroplakin II promoters driving expression of the
E1A gene of adenovirus serotype 5. Handling, replication,
amplification, purification and titration of Ad5-UPII-E1A were
performed as previously described (14). Our previous studies revealed that
Ad5-UPII-E1A with 10 multiplicity of infection (MOI) had the most
significant cytotoxicity within 96 h (16), therefore, all human bladder cancer
cells were infected with Ad5-UPII-E1A at a MOI of 10 in the present
study.
Human bladder cancer 5,637 cells were obtained from
the American Type Culture Collection (Manassas, VA, USA) and
maintained in RPMI-1640 medium supplemented with 10% fetal bovine
serum (both from Thermo Fisher Scientific, Inc., Waltham, MA, USA),
100 U/ml penicillin and 100 U/ml streptomycin at 37°C in 5%
CO2.
Chemotherapeutic agents
MMC and HCPT (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) were prepared as a stock solution at concentrations of 0.4
and 1.0 mg/ml, frozen in sterile 1.5 ml tubes individually at
−80°C, protected from light and used only once. The stock solutions
were serially diluted prior to their addition to cell cultures to
reach final concentrations of 0.05, 0.1 and 0.2 mg/ml MMC and 0.1,
0.2 and 0.4 mg/ml HCPT. Chemotherapy drugs were added 4 h following
the infection with Ad5-UPII-E1A.
Cell viability assay
Cell viability was quantified by MTT assay. Bladder
cancer 5,637 cells in 96-well plates at 2×103 cells per
well were infected with Ad5-UPII-E1A, chemotherapy drugs MMC and
HCPT, or infected by Ad5-UPII-E1A combined with either single
agent. Mock-treated cells (treated with equal amounts of PBS)
without any treatment were used as negative controls. Following
treatment for 24, 48, 72 or 96 h at 37°C, respectively, the medium
was replaced with 90 µl serum-free medium and 10 µl MTT solution (5
mg/ml in sterile PBS). Following incubation for 4 h at 37°C, the
MTT solution in the wells was replaced with 150 µl dimethyl
sulfoxide. The absorbance of the samples was measured using a
Bioelisa Reader (EXL-800, BioTek Instruments, Inc., Winooski, VT,
USA) at 490 nm. The percentage of cell viability was calculated
according to the formula 100% × (mean value A490 of infected
cells)/(mean value A490 of uninfected cells). Results are expressed
as the mean ± standard deviation (mean ± SD) for selected paradigms
performed in triplicate (n=3). The nature of the combined effect of
drugs with adenovirus was estimated by using previously published
methods (19). In brief, the expected
value of combination effect between treatment 1 and treatment 2 was
calculated as [(observed treatment 1 value)/(control value)] ×
[(observed treatment 2 value)/(control value)] × (control value),
and the combination index was calculated as the ratio of (expected
value)/(observed value). A ratio of >1 indicated a synergistic
effect, and a ratio of <1 indicated an antagonistic effect.
Cell cycle distribution analysis
A total of 5,637 cells (3×106
cells/culture flask) was treated with Ad5-UPII-E1A alone or
combined with different concentrations of MMC (0.05, 0.1 and 0.2
mg/ml) or HCPT (0.1, 0.2 and 0.4 mg/ml) for 48, 72 and 96 h at 37°C
and was harvested using trypsin. The cells were washed with PBS and
then kept overnight at 4°C in 70% ethanol. The cells were then
collected and resuspended in PBS, propidium iodide (PI; 50 mg/ml;
Sigma-Aldrich; Merck KGaA) and RNase A (100 mg/ml; Thermo Fisher
Scientific, Inc.), and incubated at 37°C for 30 min. Cell cycle
distribution was evaluated by flow cytometry (Beckman Coulter Epics
XL; Beckman Coulter, Inc., Brea, CA, USA), and data were analyzed
by CytExpert software (edition 1.0.135.1; Beckman Coulter, Inc.,
Brea, CA, USA).
Apoptosis assays
Annexin V and PI double dyes and immunofluorescence
flow cytometry assay were used to examine the apoptotic rate of the
5,637 cell line following treatment with Ad5-UPII-E1A alone or
combined with different concentrations of MMC (0.05, 0.1 and 0.2
mg/ml) and HCPT (0.1, 0.2 and 0.4 mg/ml). After 72 h treatment with
adenovirus and chemotherapy, 5,637 cells (1×106) were
collected and resuspended in binding buffer (Wuhan Amyjet
Scientific, Inc., Wuhan, China), Annexin V-fluorescein
isothiocyanate (FITC) and PI (Vybrant Apoptosis Assay kit 2;
Molecular Probes; Thermo Fisher Scientific, Inc.) were added to
each sample according to the manufacturer's instructions and
incubated in the dark for 10 min at room temperature. The number of
apoptotic cells was evaluated by flow cytometry (Ex=488 nm; Em=530
nm; BD Biosciences, San Jose, CA, USA), and data were analyzed by
CytExpert software (edition 1.0.135.1; Beckman Coulter, Inc.).
Electron microscopy assay
For ultrastructural analyses, 5,637 cells were
treated with Ad5-UPII-E1A and/or drugs and incubated for 72 h. The
cells were washed with PBS, suspended in 2.5% glutaraldehyde,
post-fixed for 1 h in 1% osmium tetroxide at room temperature,
dehydrated in ethanol (15, 30, 50, 70 and 90%), and embedded in
epoxy resin (at room temperature for 12 h). Ultrathin sections were
cut (10-mm thick), stained with lead citrate and uranyl acetate
(for 15 min at room temperature) and observed with JEM-1230
transmission electron microscopy (TEM; Japan; magnification,
×3,000–40,000).
Western blot analysis for E1A
expression
To detect expression of the E1A gene and virus
replication in cells, 5,637 cells were infected with recombinant
Ad5-UPII-E1A at an MOI=10 only, or followed by combined treatment
with MMC or HCPT. After 72 h, cells were washed with PBS and then
lysed in radioimmunoprecipitation assay cell lysis buffer with the
fresh protease inhibitor phenylmethylsulfonyl fluoride
(Sigma-Aldrich; Merck KGaA) on ice. Total protein (40 µg) was
separated on 8–12% polyacrylamide gels and then transferred to
polyvinylidene fluoride membranes (0.22 µm). The membranes were
incubated with anti-Ad5 E1A mouse mono-clonal antibodies at a
dilution of 1:200 (cat. no. ab52523; Abcam, Cambridge, UK) at 4°C
overnight. Horseradish peroxidase-conjugated anti-mouse
immunoglobulin G (cat. no. TA130005, OriGene Technologies, Inc.,
Beijing, China) was used as a secondary antibody at a dilution of
1:500 at room temperature for 2 h. Reactivity was visualized using
an enhanced chemiluminescence system (Upstate Biotechnology, Inc.,
Lake Placid, NY, USA). Equal amounts of protein loading were
controlled by GAPDH in the sample and visualized with mouse
anti-GAPDH mAb (cat. no. TA802519; OriGene Technologies, Inc.) at a
dilution of 1:1,000 at 4°C overnight.
Statistical analysis
Data are presented as the mean ± SD. All experiments
were performed in triplicate. Statistical analysis was conducted
using SPSS software (version 17.0; SPSS, Inc., Chicago, IL, USA).
Significance was assessed using one-way analysis of variance and a
post hoc test (Tukey's test). P<0.05 was considered to indicate
a statistically significant difference.
Results
MMC and HCPT combined with
Ad5-UPII-E1A synergistically inhibits cell viability yin a dose-
and time-dependent manner
To evaluate the cytotoxic effects of the combination
treatment, the viability of the human bladder cancer 5,673 cell
line was depicted by a dose-response curve (Fig. 1). MTT assays were performed according
to a standard operating procedure. Drug concentration was optimized
to a certain extent so that it would not generate an extensive
cytotoxic effect alone. Dose ranges of the MMC (0.05, 0.1 and 0.2
mg/ml) and HCPT (0.1, 0.2 and 0.4 mg/ml), as well as 10 MOI
Ad5-UPII-E1A were used. The nature of the combined effect of drugs
with adenovirus was estimated as described in material and methods.
Therefore, compared with either chemotherapy drug alone (Fig. 1A and C), MMC and HCPT combined with
Ad5-UPII-E1A synergistically inhibited cell growth in a dose- and
time-dependent manner in bladder cancer 5,637 cells (Fig. 1B and D).
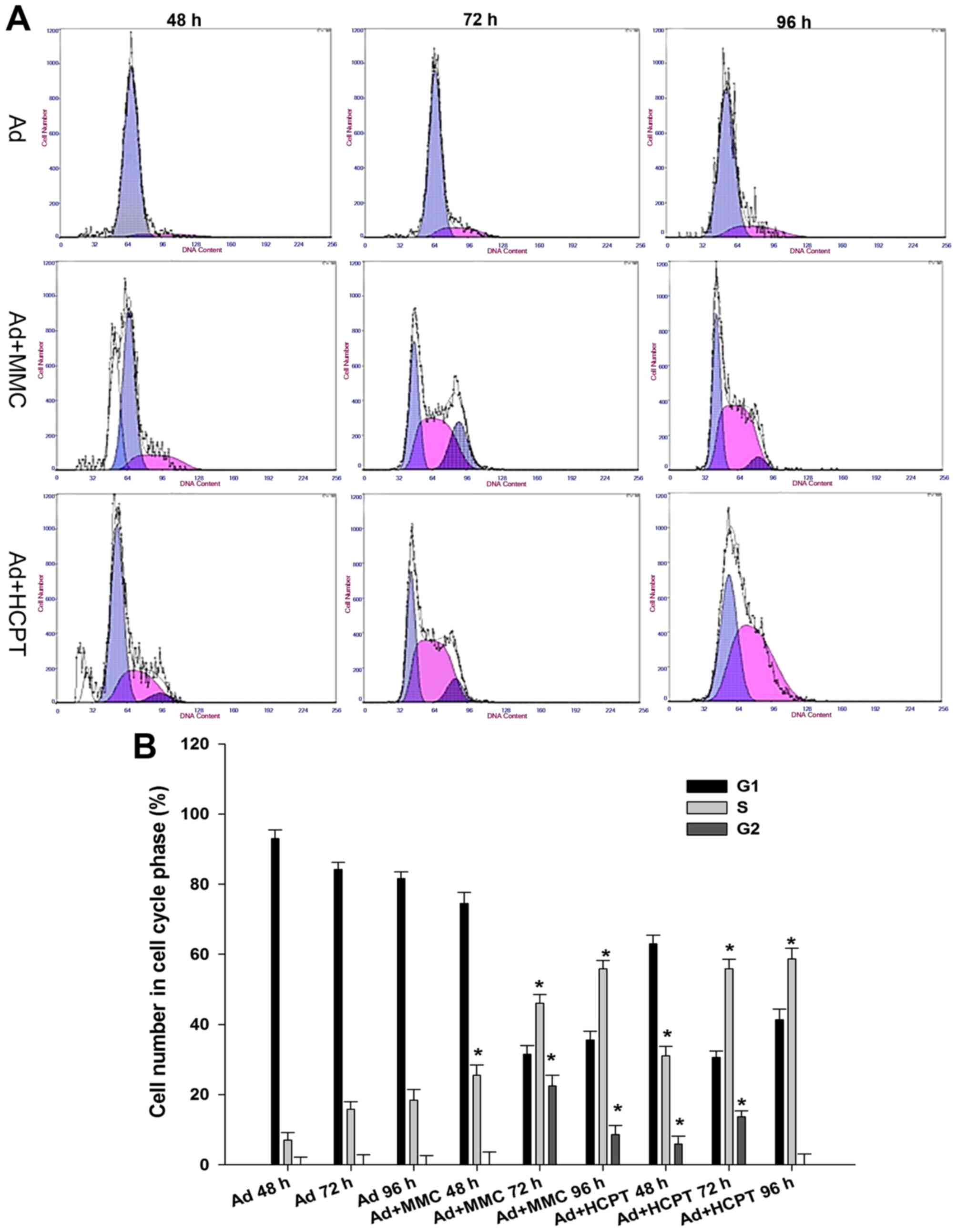
A total of 5,637 bladder cancer cells
is arrested at the G1 or S phase when treated with adenovirus or
combination therapy
In order to decipher the suppressive mechanisms of
Ad5-UPII-E1A and MMC and HCPT on bladder cancer cells, the changes
of cell cycle distribution were monitored using flow cytometry.
Compared with the control group, when 5,637 cells were infected
with Ad5-UPII-E1A (10 MOI) for 48, 72 and 96 h, the proportions of
G0/G1 phase cells were 92.85±1.4, 83.39±2.31 and 80.8±1.83%,
respectively, and the proportion of S phase cells did not exceed
20% in these groups (Fig. 2A). Cell
cycle was arrested in the G1 phase. However, in the combination
group, following treatment of 5,637 cells with 0.1 mg/ml MMC or
HCPT combined with Ad5-UPII-E1A (10 MOI), the proportion of S phase
cells increased with time, and the proportion of G2/M phase cells
decreased with time (Fig. 2A). Cell
cycle was arrested in the S phase. When 5,637 cells were treated
with 0.1 mg/ml MMC combined with Ad5-UPII-E1A (10 MOI) for 72 h,
the proportion of cells in the S phase increased along with time,
and the proportion of G2/M phase cells decreased to 22.4±1.32%
(Fig. 2A). When 5,637 cells were
treated with two chemotherapy drugs at a concentration of 0.1 mg/ml
combined with Ad5-UPII-E1A, the cell cycle was blocked at the S
phase (Fig. 2A). The difference of
cell proportion in the S phase between the combined group and the
Ad5-UPII-E1A group was significant (P<0.01; Fig. 2B).
Chemotherapy enhances
Ad5-UPII-E1A-induced necrosis in 5,637 cells
To investigate whether the adenovirus and
chemotherapy-mediated cell death in bladder cancer cells was due to
an apoptotic mechanism, apoptosis was analyzed by flow cytometry
with FITC and PI double staining. In cell scatter plots, there were
almost no apoptotic cells in the control group, and normal cells
accounted for 99.6% of all cells (Fig.
3A). The apoptotic rate was 12.93% in 5,637 cells when treated
with Ad5-UPII-E1A alone for 72 h (Fig.
3A). However, when 5,637 cells were treated with Ad5-UPII-E1A
combined with MMC, the apoptotic rate was 23.8 (0.05 mg/ml), 9.13
(0.1 mg/ml) and 2.6% (0.2 mg/ml; P<0.05; Fig. 3A, the lower right quadrant), while the
proportion of necrotic cells gradually increased by 12.6, 13.8 and
79.8%, respectively (P<0.05; Fig.
3A, the upper right quadrant). This indicated that a low dose
of chemotherapy combined with Ad5-UPII-E1A induced an apoptotic
effect, while a high dose of chemotherapy combined with
Ad5-UPII-E1A induced necrosis in a dose-dependent manner. The
apoptotic and necrotic trends of the HCPT combined with
Ad5-UPII-E1A groups were similar, but the apoptosis and necrosis
effect were more evident compared with MMC (Fig. 3A). For instance, when comparing the
two drugs at the same concentrations of 0.1 mg/ml, 9.13% of cells
were apoptotic following treatment with MMC combined with
Ad5-UPII-E1A, and 13.8% of cells were necrotic (Fig. 3A). However, in the HCPT combined with
Ad5-UPII-E1A group, 19.73% of cells were apoptotic and 16.5% were
necrotic (Fig. 3A). The
quantification of the results is depicted in Fig. 3B.
Apoptosis was most notable in the HCPT
combined with Ad5-UPII-E1A group by TEM
To analyze whether chemo drugs MMC and HCPT combined
with Ad5-UPII-E1A promoted apoptosis, TEM was used to capture
images of 5,637 cells following treatment with Ad5-UPII-E1A and
drugs for 72 h. As shown in Fig. 4,
virus particles were observed in cells from the two combined groups
(Fig. 4A and B) and Ad5-UPII-E1A
group (Fig. 4C). The distribution and
quantity of virus particles was not visibly different. Recombinant
adenovirus was widely present in the cytoplasm, either scattered or
regularly spaced (Fig. 4D and E). The
early apoptotic phenotype, including vacuolization, cell shrinkage
and the advanced apoptotic phenotype of budding and edge set, and
the apoptosis bodies, were all observed in cells of the combined
groups (Fig. 4A and B). The
phenomenon of apoptosis was most notable in the group treated with
HCPT combined with Ad5-UPII-E1A for 72 h, and apoptosis occurred in
a time-dependent manner (Fig.
4A).
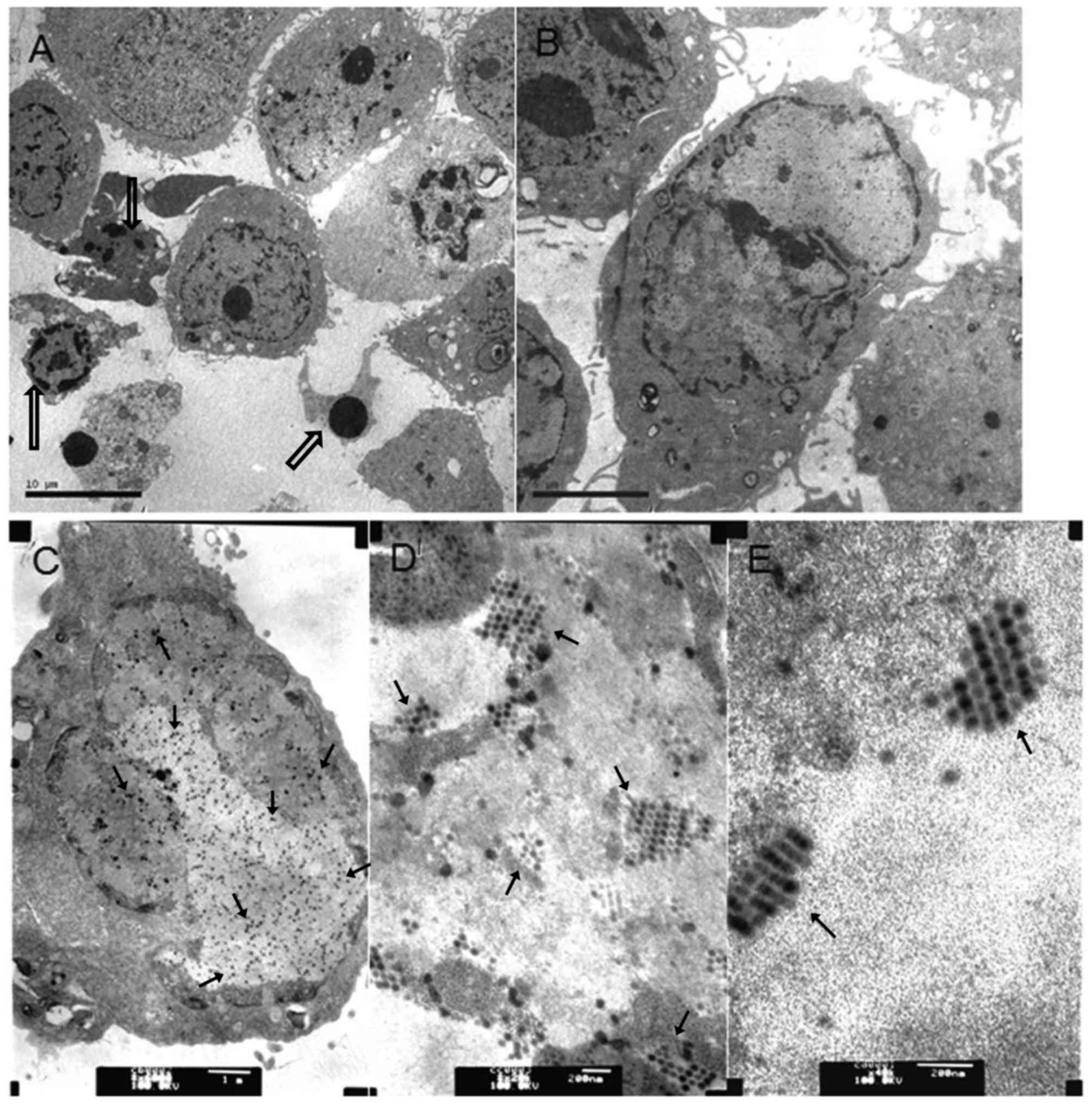 | Figure 4.Virus particles in bladder cancer
cells infected with Ad5-UPII-E1A or Ad5-UPII-E1 Acombined with MMC
or HCPT. (A) A total of 5,637 cells was treated with 0.4 mg/ml HCPT
and Ad5-UPII-E1A (10 MOI) for 72 h (magnification, ×3,000). (B) A
total of 5,637 cells was treated with 0.2 mg/ml MMC combined with
Ad5-UPII-E1A (10 MOI) for 72 h (magnification, ×6,000). (C) A total
of 5,637 cells was treated with Ad5-UPII-E1A alone, and the uniform
distribution of virus particles within the cell is visible
(magnification, ×20,000). (D) Distribution of virus particles
within the cell (magnification, ×40,000). (E) Virus particle
morphology (magnification, ×40,000). Hollow arrows indicate the
fragmented nuclei, the gathering of nuclearchromatin and the
formation of apoptotic bodies. Regular arrows indicate the virus
particles. Ad5-UPII-E1A, urothelium-specific recombinant adenovirus
type 5; MMC, mitomycin; HCPT, hydroxycamptothecin; 10 MOI, 10
multiplicity of infection. |
Ad5-UPII-E1A expresses higher levels
of E1A protein following exposure to MMC and HCPT
For Ad5-UPII-E1A, the E1A expression vector contains
the entire E1A coding sequence under the regulation of the UPII
promoter (16). To assess whether the
expression of Ad5-UPII-E1A vector was increased following treatment
with MMC or HCPT, expression of E1A protein from infected 5,637
cells was examined using western blot analysis. As shown in
Fig. 5, when 5,637 cells were treated
by Ad5-UPII-E1A (10 MOI) combined with different doses of MMC and
HCPT for 72 h, Ad5-UPII-E1A expressed higher levels of E1A protein
compared with the Ad5-UPII-E1A alone group. Furthermore, the
increase of E1A was dosage-dependent in the Ad5-UPII-E1A and MMC
combination group (Fig. 5). Multiple
E1A protein species were produced from this vector, as predicted,
ranging in size from ~30–50 kDa. E1A-hybridizing bands were
detected in infected 5,637 cells, thus, Ad5-UPII-E1A was able to
effectively transduce 5,637 cells with the addition of chemotherapy
drugs, resulting in the production of multiple E1A protein
species.
 | Figure 5.Expression of E1A in the 5,637 cell
line following treatment with MMC or HCPT. A total of 5,637 cells
was infected with recombinant Ad5-UPII-E1A (10 MOI) alone or with
Ad5-UPII-E1A combined with MMC or HCPT for 72 h. The results
revealed that, following exposure to Ad5-UPII-E1A (10 MOI) combined
with different doses of MMC and HCPT for 72 h, E1A was increased
compared with Ad5-UPII-E1A alone, and is increase was dependent on
the dose of MMC and HCPT. GAPDH was used as a loading control. E1A,
adenoviral early region gene; MMC, mitomycin; HCPT,
hydroxycamptothecin; Ad5-UPII-E1A, urothelium-specific recombinant
adenovirus type 5; 10 MOI, 10 multiplicity of infection. |
Discussion
Oncolytic adenoviruses, which are considered to be
an important method of cancer gene therapy, have been an effective
treatment strategy to date (11).
Furthermore, tissue-specific adenovirus may function as a type of
tumor-specific chemotherapy sensitizer, since it is highly targeted
to tumor tissues through tissue-specific promoters and enhancers
(20). It has been reported that the
adenovirus E1A gene may induce sensitivity to DNA-damaging agents,
including cisplatin (CDDP), DOX and γ irradiation on squamous cell
carcinoma cells. Ganjavi et al (21) demonstrated that Ad-wtp53 significantly
increased sensitivity of the cell lines [Saos-2 (p53-/−), HOS
(R156P), KHOS/NP (R156P) and MNNG (R156P, F270L)] to CDDP and DOX,
chemotherapeutic agents commonly used in the treatment of
osteosarcoma.
The present study revealed that chemotherapy
combined with Ad5-UPII-E1A more effectively killed bladder cancer
cells than the groups treated with MMC/HCPT or Ad5-UPII-E1A alone,
as indicated in Fig. 1. When 5673
cells were treated with HCPT (0.2 mg/ml) for 96 h, cell viability
was 63.98%, however, when 5,673 cells were treated with adenovirus
plus HCPT (0.1 mg/ml) for 48 h, cell viability declined to 63.51%,
indicating that pre-treatment of cells with Ad5-UPII-E1A sensitized
cells to chemotherapy-induced cell death, and therefore it may be
possible to reduce the drug dosage. In combination therapy, reduced
dosage of chemotherapy drugs may achieve an improved therapeutic
effect, which may alleviate the side effects of chemotherapy drugs.
Ad5-UPII-E1A increased the sensitivity of bladder cancer cells to
chemotherapy, and E1A protein expression sensitized tumor cells to
chemotherapy drugs. One of the molecular mechanisms by which E1A
induces chemosensitization is downregulation of erb-b2 receptor
tyrosine kinase 2/proto-oncogene Neuoverexpression (22,23).
Inhibition of protein kinase Band activation of p38 was reported to
provide a general cellular mechanism for E1A-mediated chemo
sensitization (24,25). Regulation of certain critical tumor
suppressors was also proposed as being involved in E1A-induced
chemo sensitization, including p53 and p19ARF (26), the pro-apoptotic protein B-cell
lymphoma-2-associated X protein, caspase 9 and an as-of-yet
unidentified inhibitor that ordinarily provides protection against
cell death (27,28). A forkhead box O3 a dependent mechanism
of E1A-induced chemo sensitization was also previously reported
(29). The mechanisms underlying E1A
protein-induced sensitization of bladder cancer cells to HCPT and
MMC will be discussed in the near future.
Next, the mechanisms underlying the antitumor effect
of the combined treatment were investigated. The cell cycle and
apoptosis distribution, determined by flow cytometric analysis,
demonstrated that chemotherapy promoted oncolytic
adenovirus-induced apoptosis. When bladder cancer cells were
treated with Ad5-UPII-E1A alone, the cell cycle was arrested in the
G1 phase. However, when cells were treated with Ad5-UPII-E1A and
HCPT or MMC, the cell cycle was arrested in the S phase,
demonstrating that the drugs have a stronger cell cycle blocking
function than Ad5-UPII-E1A. Pharmacological studies have
demonstrated that MMC is able to block the cell cycle at the G1 and
S phase in the late period (30).
HCPT has a cytotoxic activity against S-phase cells (31), which was consistent with the results
of the present study.
The fact that theapoptotic effect was increased by
combined treatment demonstrated that Ad5-UPII-E1A combined with MMC
or HCPT had amore comprehensive inhibitory effect on bladder cancer
cells. Greater apoptosis and necrosis effects were observed in the
combination therapy group under an electron microscope. The
observation of cell phenotypes in the early or late-stage of
apoptosis demonstrated that addition of MMC and HCPT enhanced
Ad5-UPII-E1A-induced apoptosis. The phenomenon of apoptosis was
most notable in the group treated with a combination of HCPT and
Ad5-UPII-E1A for 72 h, compared with MMC combined therapy, which
may provide a reference for clinical application. The results of
the present study were similar to those of other studies that
demonstrated the existence of a cooperative or enhanced curative
effect when chemotherapy was combined with adenovirus treatment
(32,33). The present study confirmed that a
combination of chemotherapy with adenovirus resulted in an enhanced
antitumor effect through the induction of apoptosis.
By contrast, the effect of MMC and HCPT on
Ad5-UPII-E1A was investigated. Compared with the group treated only
with Ad5-UPII-E1A, MMC and HCPT promoted adenoviral E1A protein
expression. This may be caused by the increased mRNA transcription
levels of E1A, and the potential underlying mechanisms will be
explored in the future.
Although combination therapy was demonstrated to
result in decreased 5,637 cell viability and increased apoptosis,
with this increase depending on the dose of MMC and HCPT, the
underlying mechanisms remain to be reported. Additional studies are
required to validate the synergistic effect of Ad5-UPII-E1A and
chemotherapy combination treatment on animals and in clinical
trials, and to investigate the mechanisms underlying the
synergistic sensitization. In conclusion, combined therapy using
Ad5-UPII-E1A and MMC or HCPT may be a promising treatment for
patients with bladder cancer.
Acknowledgements
The present study was supported by the National
High-Tech R&D Program of China (863 program; grant no.
2008AA02Z421), the National Natural Science Foundation of China
(grant nos. 81172437/H1619 and 81372733/H1619) and the Natural
Science Foundation of Gansu Province (grant no. 1506RJZA234).
References
|
1
|
Cheung G, Sahai A, Billia M, Dasgupta P
and Khan MS: Recent advances in the diagnosis and treatment of
bladder cancer. BMC Med. 11:132013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sylvester RJ, van der Meijden AP,
Oosterlinck W, Witjes JA, Bouffioux C, Denis L, Newling DW and
Kurth K: Predicting recurrence and progression in individual
patients with stage Ta T1 bladder cancer using EORTC risk tables: A
combined analysis of 2596 patients from seven EORTC trials. Eur
Urol. 49:466–477. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Voutsinas GE and Stravopodis DJ: Molecular
targeting and gene delivery in bladder cancer therapy. J Buon. 14
Suppl 1:69–78. 2009.
|
|
4
|
Babjuk M, Oosterlinck W, Sylvester R,
Kaasinen E, Böhle A and Palou-Redorta J; European Association of
Urology (EAU), : EAU guidelines on non-muscle-invasive urothelial
carcinoma of the bladder. Eur Urol. 54:303–314. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Crooke ST and Bradner WT: Mitomycin C: A
review. Cancer Treat Rev. 3:121–139. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Verweij J and Pinedo HM: Mitomycin C:
Mechanism of action, usefulness and limitations. Anticancer Drugs.
1:5–13. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu Y, Zeng FQ, Wang YB and Wang L:
Hydroxycamptothecin promotes the apoptosis of prostate cancer cell
line PC-3. Zhonghua Nan KeXue. 13:890–894. 2007.(In Chinese).
|
|
8
|
Fan J, Tang X and Zhang X:
10-Hydroxycamptothecin induces apoptosis in human T24 urinary
bladder cancer cells. Zhonghua Yi XueZaZhi. 78:301–304. 1998.(In
Chinese).
|
|
9
|
Liu LF, Desai SD, Li TK, Mao Y, Sun M and
Sim SP: Mechanism of action of camptothecin. Ann N Y Acad Sci.
922:1–10. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dean M, Fojo T and Bates S: Tumour stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bochner BH: Gene therapy in bladder
cancer. Curr Opin Urol. 18:519–23. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wada Y, Gotoh A, Shirakawa T, Hamada K and
Kamidono S: Gene therapy for bladder cancer using adenoviral
vector. Mol Urol. 5:47–52. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chester JD, Kennedy W, Hall GD, Selby PJ
and Knowles MA: Adenovirus-mediated gene therapy for bladder
cancer: Efficient gene delivery to normal and malignant human
urothelial cells in vitro and ex vivo. Gene Ther. 10:172–179. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang F, Wang Z, Tian H, Qi M, Zhai Z, Li
S, Li R, Zhang H, Wang W, Fu S, et al: Biodistribution and safety
assessment of bladder cancer specific recombinant oncolytic
adenovirus in subcutaneous xenografts tumor model in nude mice.
Curr Gene Ther. 12:67–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang D, Wang Z, Tian J, He X, Chowdhury
WH, Zhang X, Li S and Rodriguez R: Prostate stem cell antigen
enhancer and uroplakin II promoter based bladder cancer targeted
tissue-specific vector. Urol Oncol. 28:164–169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhai Z, Wang Z, Fu S, Lu J, Wang F, Li R,
Zhang H, Li S, Hou Z, Wang H and Rodriguez R: Antitumor effects of
bladder cancer-specific adenovirus carrying E1A-andro gen receptor
in bladder. cancer. 19:1065–1074. 2012.
|
|
17
|
Guo W, Zhu H, Zhang L, Davis J, Teraishi
F, Roth JA, Stephens C, Fueyo J, Jiang H, Conrad C and Fang B:
Combination effect of oncolytic adenovirotherapy and TRAIL gene
therapy in syngeneic murine breast cancer models. Cancer GeneTher.
13:82–90. 2006.
|
|
18
|
Alonso MM, Gomez-Manzano C, Jiang H,
Bekele NB, Piao Y, Yung WK, Alemany R and Fueyo J: Combination of
the oncolytic adenovirus ICOVIR-5 with chemotherapy provides
enhanced anti-glioma effect in vivo. Cancer Gene Ther. 14:756–761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou JR, Yu L, Mai Z and Blackburn GL:
Combined inhibition of estrogen-dependent human breast carcinoma by
soy and tea bioactive components in mice. Int J Cancer. 108:8–14.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sánchez-Prieto R, Quintanilla M, Cano A,
Leonart ML, Martin P, Anaya A and Ramón y Cajal S: Carcinoma cell
lines become sensitive to DNA-damaging agents by the expression of
the adenovirus E1A gene. Oncogene. 13:1083–1092. 1996.PubMed/NCBI
|
|
21
|
Ganjavi H, Gee M, Narendran A, Parkinson
N, Krishnamoorthy M, Freedman MH and Malkin D: Adenovirus-mediated
p53 gene therapy in osteosarcoma cell lines: Sensitization to
cisplatin and doxorubicin. Cancer Gene Ther. 13:415–419. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brader KR, Wolf JK, Hung MC, Yu D,
Crispens MA, van Golen KL and Price JE: Adenovirus E1A expression
enhances the sensitivity of an ovarian cancer cell line to multiple
cytotoxic agents through an apoptotic mechanism. Clin Cancer Res.
3:2017–2024. 1997.PubMed/NCBI
|
|
23
|
Ueno NT, Bartholomeusz C, Herrmann JL,
Estrov Z, Shao R, Andreeff M, Price J, Paul RW, Anklesaria P, Yu D,
Hung MC, et al: E1A-mediated paclitaxel sensitization in
HER-2/neu-overexpressing ovarian cancer SKOV3.ip1 through apoptosis
involving the caspase-3 pathway. Clin Cancer Res. 6:250–259.
2000.PubMed/NCBI
|
|
24
|
Liao Y and Hung MC: Regulation of the
activity of p38 mitogen-activated protein kinase by Akt in cancer
and adenoviral protein E1A-mediated sensitization to apoptosis. Mol
Cell Biol. 23:6836–6848. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liao Y and Hung MC: A new role of protein
phosphatase 2a in adenoviral E1A protein-mediated sensitization to
anticancer drug-induced apoptosis in human breast cancer cells.
Cancer Res. 64:5938–5942. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
de Stanchina E, McCurrach ME, Zindy F,
Shieh SY, Ferbeyre G, Samuelson AV, Prives C, Roussel MF, Sherr CJ
and Lowe SW: E1A signaling to p53 involves the p19(ARF) tumor
suppressor. Genes Dev. 12:2434–2442. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Duelli DM and Lazebnik YA: Primary cells
suppress oncogene-dependent apoptosis. Nat Cell Biol. 2:859–862.
2000. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
McCurrach ME, Connor TM, Knudson CM,
Korsmeyer SJ and Lowe SW: bax-deficiency promotes drug resistance
and oncogenic transformation by attenuating p53-dependent
apoptosis. Proc Natl Acad Sci USA. 94:pp. 2345–2349. 1997;
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Su JL, Cheng X, Yamaguchi H, Chang YW, Hou
CF, Lee DF, Ko HW, Hua KT, Wang YN, Hsiao M, et al: FOXO3a
dependent mechanism of E1A-Induced chemosensitization. Cancer Res.
71:6878–6887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Olivier M and Theillet C: Mitomycin C
induced apoptosis: influence of cell cycle phase. Biology of the
Cell. 88:82–82a. 1996. View Article : Google Scholar
|
|
31
|
Hu W, Zhang C, Fang Y and Lou C:
Anticancer properties of 10-hydroxycamptothecin in a murine
melanoma pulmonary metastasis model in vitro and in vivo. Toxicol
In Vitro. 25:513–520. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tanaka M and Grossman HB: Connexin 26 gene
therapy of human bladder cancer: Induction of growth suppression,
apoptosis, and synergy with Cisplatin. Hum Gene Ther. 12:2225–2236.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Y, Yu DC, Chen Y, Amin P, Zhang H,
Nguyen N and Henderson DR: A hepatocellular carcinoma-specific
adenovirus variant, CV890, eliminates distant human liver tumors in
combination with doxorubicin. Cancer Res. 61:6428–6436.
2001.PubMed/NCBI
|