Introduction
Bladder cancer is one of the most common urothelial
malignancies (1). Superficial bladder
cancer can be treated by the transurethral resection of bladder
tumors; however, 50–70% of noninvasive cancers recur and 15–30% of
cases progress to invasive muscle disease (1,2). Although
systemic chemotherapy is typically used in the treatment of the
invasive bladder cancer, recurrence and progression rates remain
high. Bladder cancer is therefore resistant to conventional
therapeutic methods, and novel therapeutic agents for the treatment
of bladder cancer are required (1).
The reduced expression in immortalized cells (REIC)
gene is also known as Dickkopf-3 (Dkk-3). A previous study from our
group reported that REIC/Dkk-3 is a tumor suppressor gene that may
be a target for future treatments (2). The expression of REIC/Dkk-3 is
downregulated in a wide range of human cancer types, including
bladder cancer, which suggests that REIC/Dkk-3 is a promising
therapeutic gene (2,3). With the expectation that the
overexpression of REIC/Dkk-3 could promote anticancer effects, our
group previously developed an adenovirus vector encoding the human
REIC/Dkk3 gene (Ad-REIC) and demonstrated that the Ad-REIC agent
induced cancer-specific apoptosis in a number of cancer cell lines
(4), and the phosphorylation of c-Jun
N-terminal kinase (JNK) was revealed to be an important step in
inducing cancer cell apoptosis (3–5). In
addition, Ad-REIC inhibits cell motility and invasion in various
cancer cells; however, the molecular mechanisms underlying these
multiple therapeutic effects remain unclear (4).
Cluster of differentiation (CD)147 [also known as
extracellular matrix metalloproteinase inducer (EMMPRIN) or
basigin] is a cell surface glycoprotein belonging to the
immunoglobulin superfamily. CD147 was demonstrated to be the most
frequently upregulated mRNA and protein in micro-metastatic cells
isolated from the bone marrow of patients with cancer, suggesting
that it serves a key role in tumorigenesis and metastasis (6). The expression of the CD147 protein in
cancer cells, including urinary bladder, breast, lung, oral cavity,
esophageal and skin cancer, as well as malignant lymphoma cells,
has been observed to be significantly increased compared with that
in the corresponding normal tissue (6). The elevated expression of CD147 is
correlated with tumor progression in bladder cancer, glioma, giant
cell tumor of bone, laryngeal squamous cell carcinoma, ovarian
carcinoma and melanoma, indicating that CD147 serves a significant
role in the progression of cancer (6–11). It has
previously been demonstrated that CD147 is involved in the cell
glycolytic metabolic pathways that enable cancer cells to divide
and rapidly proliferate (12–14), providing an insight into the molecular
mechanisms underlying the role that CD147 serves in cancer
progression. In addition, the high expression of CD147 on the
surface of cancer cells stimulates adjacent cancer cells and
fibroblasts to produce matrix metalloproteinases (6).
Since CD147 has rapidly emerged as a strong promoter
of the development and progression of cancer, the present study
investigated the regulation of CD147 expression by the
Ad-REIC-mediated therapeutic overexpression of REIC/Dkk-3, and
examined the signaling pathways associated with the downregulation
of CD147.
Materials and methods
Cells and cell culture
Human urothelial cancer cell lines (HT1376, RT4,
T24, UMUC3, J82, 5637, TCCsup), a mouse bladder cancer cell line
(MBT2) and human embryonic kidney cells (HEK293) were obtained from
the American Type Culture Collection (Manassas, VA, USA). The KK47
human bladder cancer cell line was kindly provided by Professor S.
Naito (Department of Urology, University of Kyushu, Fukuoka, Japan)
(15). The cells were grown in
RPMI-1640 medium (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
supplemented with 10% (v/v) fetal bovine serum (FBS) (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), penicillin (100
IU/ml) and streptomycin (100 µg/ml) (2). Human normal bladder urothelial cells
(HUC; cat. no., #4320) were purchased from ScienCell™ Research
Laboratories, Inc. (San Diego, CA, USA) and were cultivated using
the medium recommended by the supplier (16).
Construction and production of
Ad-REIC
In order to generate Ad-REIC vectors,
pShuttle-SGE-REIC plasmids were digested with the restriction
enzymes I-CeuI and PI-SceI and inserted into the
Adeno-X Viral DNA (Clontech Laboratories, Inc., Mountainview, CA,
USA), as previously described (3,17).
Briefly, the recombinant adenoviral DNA with the full-length human
REIC/Dkk-3 gene was linearized by digestion with PacI and
transfected into HEK293 cells. At 7–10 days following transfection,
the HEK293 cells were harvested, and a viral solution was obtained
by three freeze/thaw cycles. The recovered virus solution was used
to propagate sufficient viruses in HEK293 cells for further
studies. All virus particles were purified by CsCl density gradient
ultracentrifugation and were stored at −80°C. An adenovirus vector
carrying the LacZ gene (Ad-LacZ) was used as a control, as
described previously (3,5). The adenoviral vectors were produced
using replication-defective adenoviruses of serotype 5.
Assay for human telomerase reverse
transcriptase (hTERT) promoter-driving activity and CD147 band
density
The hTERT promoter-driving activity was analyzed as
previously described (16). HUC,
HT1376, RT4, T24, UMUC3, J82, 5637, KK47, TCCsup and MBT2 cells
(4×105) were plated in 6-well plates in culture medium containing
charcoal-stripped FBS. The cells were cultured for 24 h, and the
transient transfection of the luciferase-encoding plasmid with the
advanced two-step transcriptional amplification (TSTA) system were
performed using the Lipofectamine transfection reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). The advanced TSTA system is a
transcriptional system that can robustly enhance the hTERT
promoter-driven and cancer-specific expression of genes (16). In the luciferase assay, the effector
plasmid was co-transfected with the reporter plasmid, which was
derived from the Dual-Luciferase Reporter Assay kit (Promega
Corporation, Madison, WI, USA). The cells were incubated for 48 h,
harvested and assayed for luciferase activity using the luciferase
assay kit and a luminescence microplate reader, according to the
manufacturer's protocol.
Western blotting
Western blotting was performed as previously
described (2,3,17). KK47
cells (5×105) were seeded in flat-bottom 6-well plates, incubated
for 24 h and then sampled. In some experiments, the cells were
treated with Ad-LacZ or Ad-REIC at a multiplicity of infection
(MOI) of 100 in 0.5 ml of complete medium for 1 h, prior to the
addition of 1.5 ml of fresh medium and incubation of the cells for
24 h. Subsequently, the floating dead cells were removed and
attached cells were lysed with ice-cold lysis buffer to extract
proteins. The insoluble fragments were removed by centrifugation
and the supernatants were adjusted to an equal protein
concentration in each experiment. The samples were separated on a
7.5% SDS-PAGE gel and transferred onto a polyvinylidene fluoride
membrane (EMD Millipore, Billerica, MA, USA) for western blotting.
Following the transfer, the membranes were blocked for 1 h with 5%
nonfat milk powder, 6% glycine and 0.1% Tween-20 in Tris-buffered
saline (TBST) at room temperature. The membranes were incubated for
1 h at room temperature with the following primary antibodies at
1:1,000 dilution: CD147 (cat. no., EPR4052; Abcam, Cambridge, UK),
REIC/Dkk-3 (mouse monoclonal, raised in our laboratory) (17), phospho-p38 mitogen-activated protein
kinase (MAPK; cat. no., #4511), phospho-p44/42 MAPK (Erk1/2; cat.
no., #4370), phospho-JNK (cat. no., #9251), phospho-c-Jun (cat.
no., #2361), c-Myc (cat. no., #9402) and β-actin (cat. no., #4967;
all from Cell Signaling Technology, Inc., Danvers, MA, USA). After
three washes in TBST, the membranes were incubated with a
horseradish peroxidase-conjugated secondary antibody (dilution,
1:5,000; cat. no., NA931; GE Healthcare Life Sciences, Little
Chalfont, UK) for 1 h at room temperature. The bound antibodies
were visualized by the enhanced chemiluminescence detection method
(Amersham ECL kit; GE Healthcare Life Sciences) using medical X-ray
film (2). In order to determine band
density by western blotting, the films were scanned and a
densitometric analysis was performed using ImageJ software (version
1.44p, available from: https://imagej.nih.gov/ij/; National Institutes of
Health, Bethesda, MD, USA).
Cell viability assay
KK47 bladder cancer cells (5×105) were seeded in
6-well plates and incubated for 24 h at 37°C. The cells were
treated with Ad-LacZ or Ad-REIC at 100 MOI in 0.5 ml of complete
medium for 1 h, 1.5 ml of fresh medium was added, and the cells
were then incubated for 72 h. Following removal of the floating
dead cells by exchanging medium, the attached cells were detached
by trypsin and the number of cells was counted using a disposable
hemocytometer without staining.
Statistical analysis
The data are presented as the mean ± standard
deviation from five experiments. The unpaired Student's t-test was
performed to analyze the statistical significance of differences
between two groups. A regression analysis was performed to examine
the correlation between two parameters. P<0.05 was considered to
indicate a statistically significant difference. Statistical
analyses were performed using StatView version 4.5 software (Abacus
Concepts, Piscataway, NJ, USA).
Results
Expression of CD147 in human
urothelial cancer cell lines
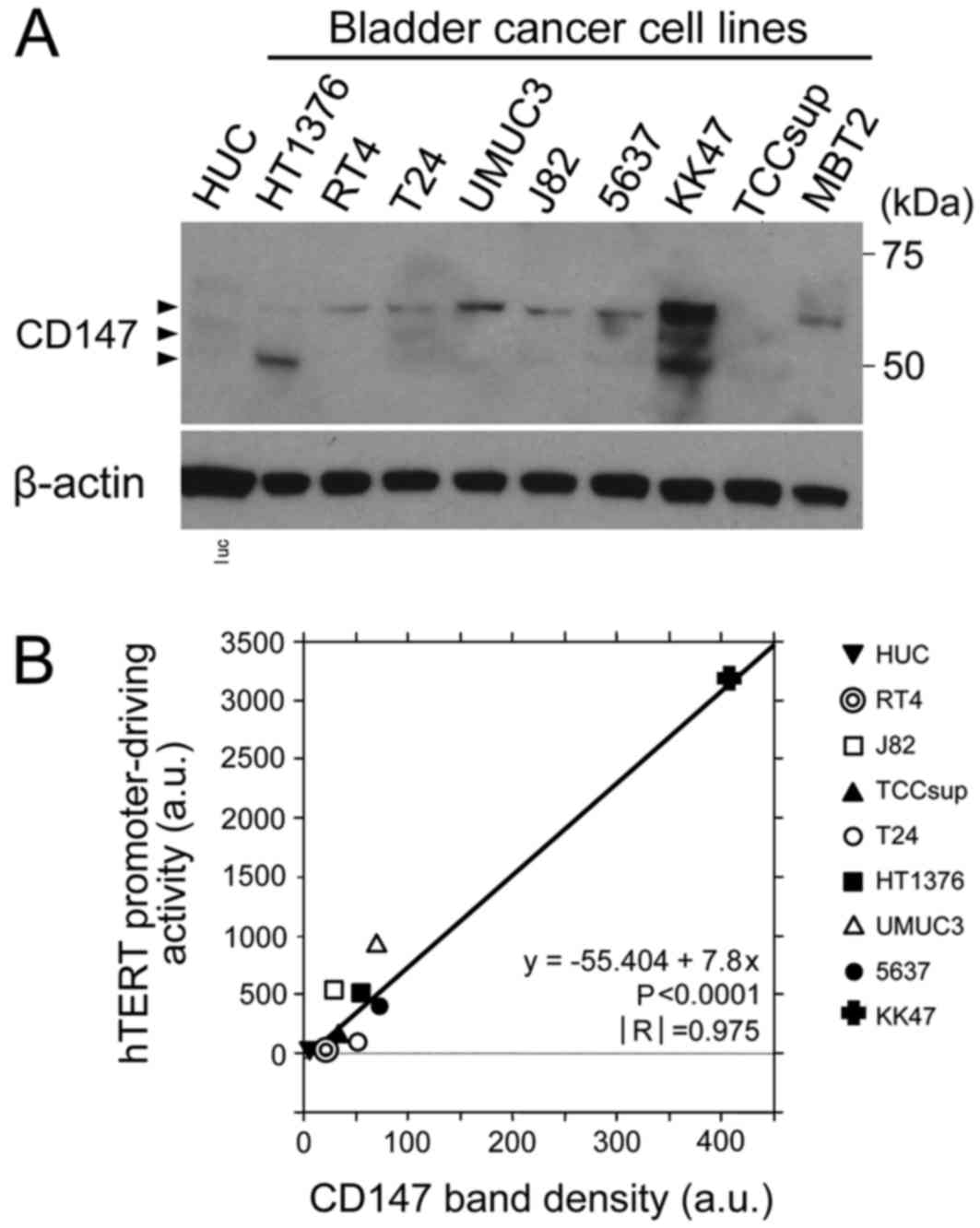
The expression of CD147 protein was first examined
by western blotting in a number of urothelial cancer cell lines and
a human normal bladder urothelial cell line (HUC). In the majority
of the urothelial cancer cell lines investigated, the CD147 protein
bands were identified at a molecular weight of 50–60 kDa (Fig. 1A); as previously reported, the protein
exhibited multiple bands of different molecular sizes (6). In the KK47 human bladder cancer cell
line, the expression level of CD147 was markedly increased compared
with that in other urothelial cancer cell lines (Fig. 1A). CD147 expression was not observed
in the HUC cells.
Correlation between the expression of
CD147 and the hTERT promoter-driving activity in human urothelial
cells
Since CD147 functions as an oncoprotein (6–11), the
association between CD147 expression and malignancy-associated
factors was examined in a number of urothelial cancer cell lines
and normal HUC cells. The hTERT promoter-driving activity was
measured as an index of the malignancy using the hTERT
promoter-driven advanced TSTA system (16). A significant correlation was observed
between the CD147 band density and the value of the hTERT
promoter-driving activity, as determined by the regression analysis
(Fig. 1B). The results indicated that
high CD147 expression levels were positively associated with the
promoter-driving activity of hTERT, and were correlated with hTERT
expression levels and cancer malignancy (16).
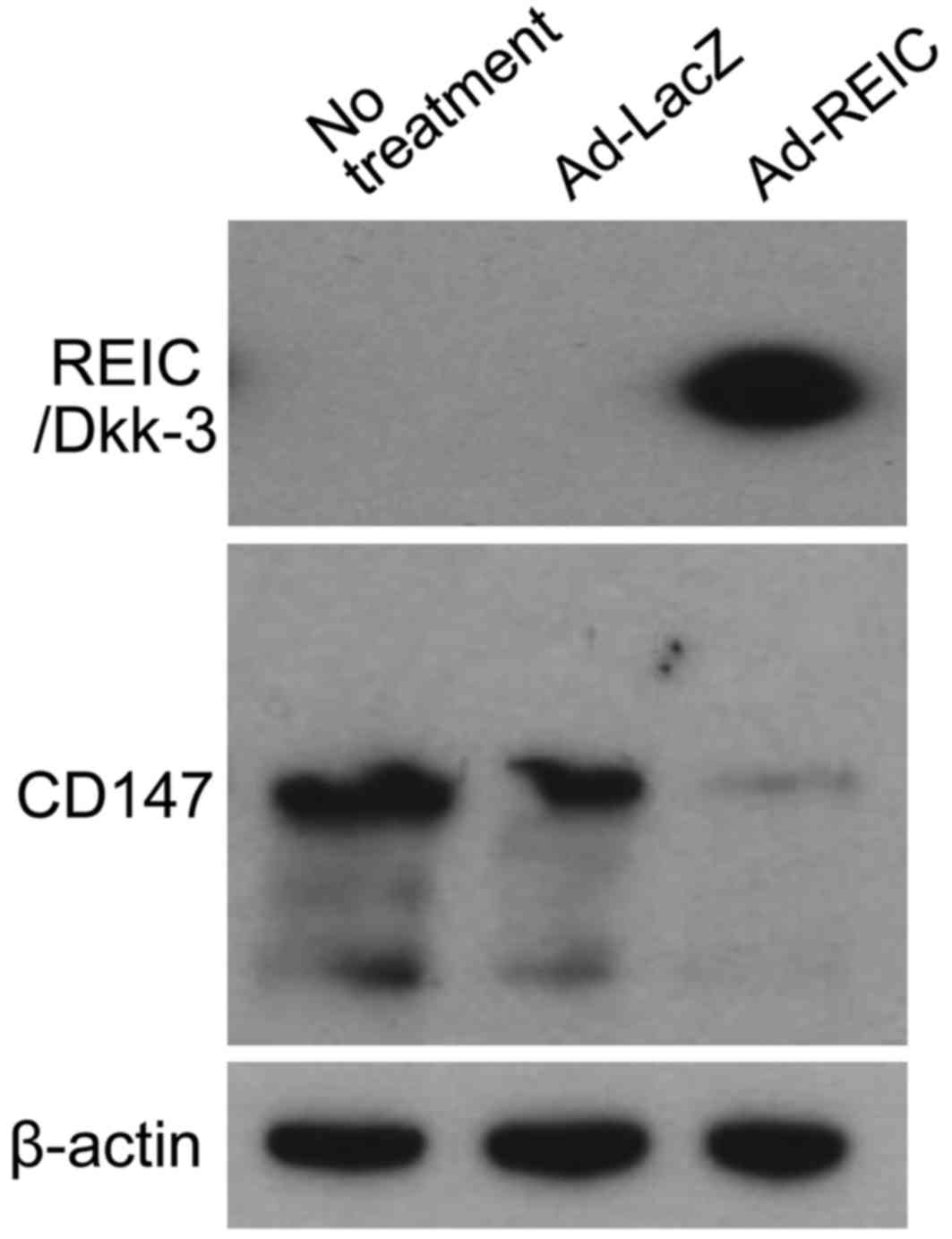
Ad-REIC treatment downregulates the
expression of CD147 in human bladder cancer KK47 cells
The expression levels of REIC/Dkk-3 and CD147
following Ad-REIC treatment were examined by western blotting
(Fig. 2). REIC/Dkk-3 protein was
expressed following Ad-REIC but not Ad-LacZ treatment. The
expression of CD147 was clearly observed in untreated and
Ad-LacZ-treated KK47 cells, whereas Ad-REIC treatment markedly
downregulated the expression of CD147 in these cells.
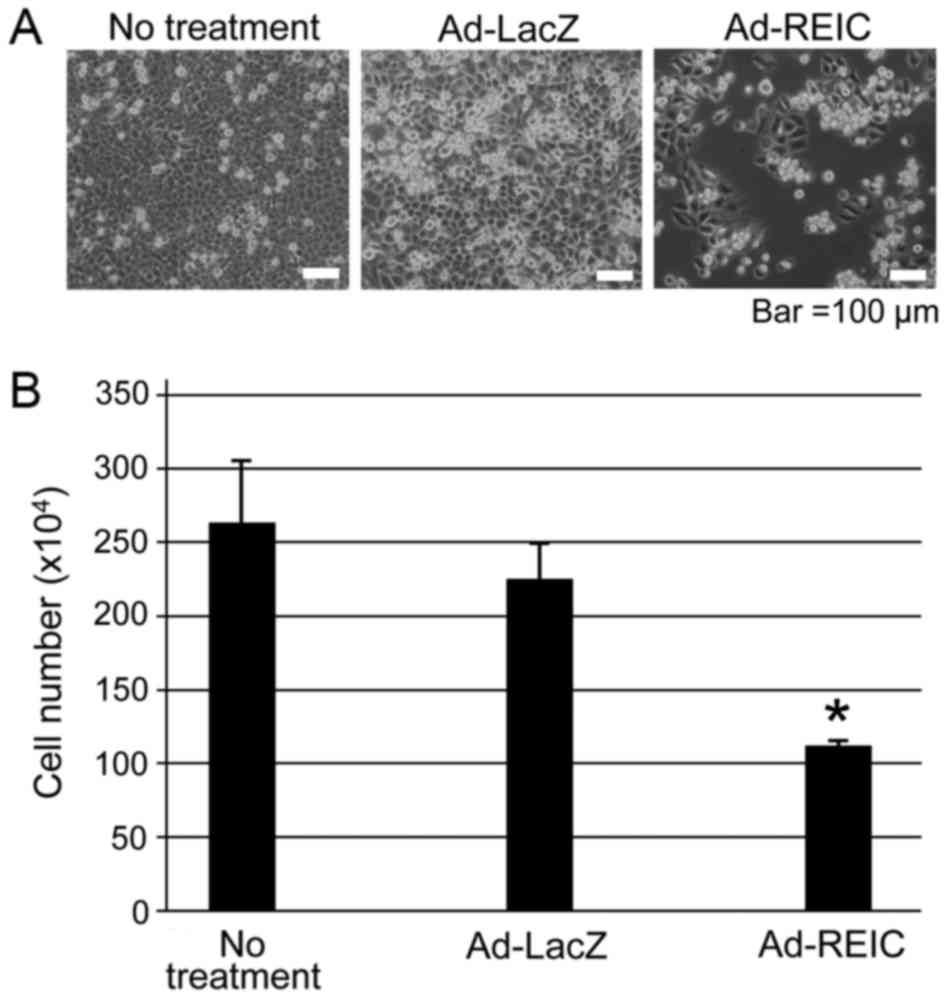
Ad-REIC significantly reduces the
viability of KK47 bladder cancer cells
KK47 cancer cells were treated with Ad-REIC to
assess the anti-proliferative effects of the agent. A marked
induction of apoptosis was observed in the Ad-REIC treatment group
in comparison to the other treatment groups (Fig. 3A). The attached viable cells were
counted following removal of floating dead cells. There number of
cells in the Ad-REIC treatment group was significantly lower
compared with that in than the other groups (P<0.05; Fig. 3B), indicating that Ad-REIC
significantly decreased cell viability in the bladder cancer cell
line.
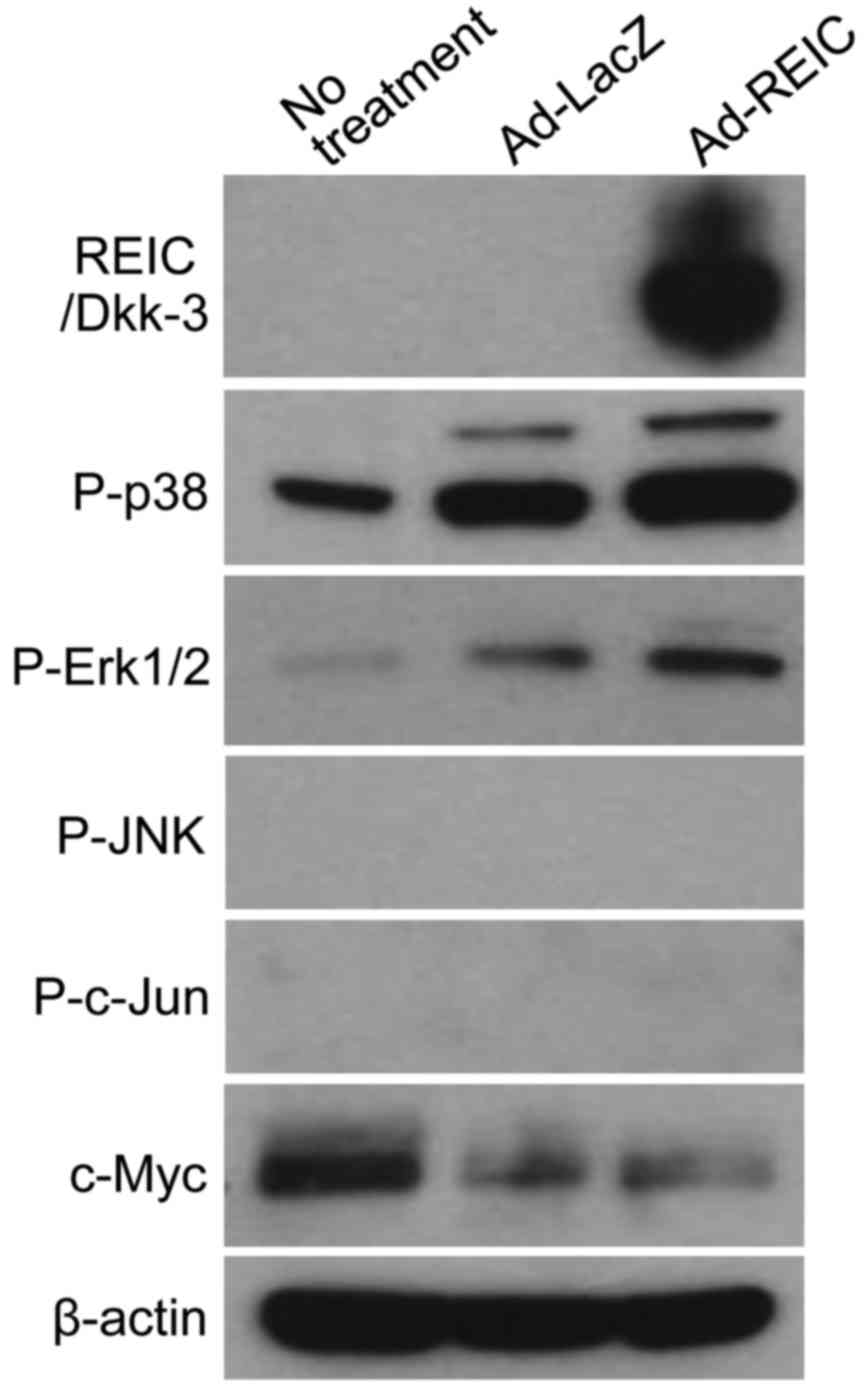
Ad-REIC downregulates the expression
of CD147 without dependence on the MAPK and c-Myc signaling
pathway
With regard to the regulation of the expression of
CD147, several signaling pathways are reported to be associated
with its gene transcription (18–23). In
particular, the expression of CD147 is reported to be positively
regulated by p38-, Erk1/2- and JNK-dependent MAPK signaling
pathways and the c-Myc protein (18,19,21,22).
The associations between CD147 expression, phosphorylation of MAPK
pathway components, and the expression of c-Myc following Ad-REIC
treatment was examined. In addition, since JNK-dependent signaling
has been reported to be a critical step in Ad-REIC-induced cancer
cell death (4,5), the phosphorylation of JNK and c-Jun was
investigated. Western blotting was performed to analyze the
expression levels of the indicated factors following Ad-LacZ and
Ad-REIC treatment (Fig. 4). The
expression of REIC/Dkk-3 protein was confirmed following Ad-REIC
treatment. Unexpectedly, no positive association was observed
between CD147 level and MAPK signaling or c-Myc expression. MAPK
signaling via p38 and Erk1/2 was markedly activated following
Ad-REIC treatment, indicating that the agent affected the intrinsic
cell survival response against cell death signaling. The results
indicated that the downregulation of CD147 by Ad-REIC occurs
without the inhibition of MAPK signaling or the suppression of
c-Myc. Furthermore, the levels of phosphorylated JNK and c-Jun were
not upregulated following Ad-REIC treatment, indicating that JNK
signaling was not significantly activated by this agent in KK47
bladder cancer cells.
Discussion
The expression of CD147 is significantly upregulated
in a number of types of malignancy (6). It is a key molecule that promotes the
proliferation, invasion and metastasis of cancer cells (6–11). CD147
is an important promoter of cell growth through the reprogramming
of glucose metabolism, including glycolysis (12,24–26). These
findings indicate that cancer cells use the CD147-dependent
glycolytic metabolic pathways in order to divide and rapidly
proliferate (12–14). Thus, inhibiting or blocking the
expression of CD147 to reduce the glycolytic capacity of the
targeted cancer cells represents an attractive cancer therapeutic
strategy (27–29). In order to further elucidate the
therapeutic mechanism of Ad-REIC agents for cancer gene therapy,
the effects of Ad-REIC on the expression of CD147 and its
associated signaling molecules in human bladder cancer cells were
investigated in the present study. In vitro Ad-REIC
treatment was revealed to significantly downregulate the expression
of CD147 and to inhibit the proliferation of bladder cancer cells,
which suggests an association between the downregulation of CD147
and the anti-cancer effects of the agents.
The molecular mechanisms underlying Ad-REIC-induced
apoptosis have been previously investigated and the phosphorylation
(activation) of JNK was demonstrated to be a critical step in
cancer cell death (5). REIC/Dkk-3
protein is a secretory protein and its overexpression in response
to Ad-REIC treatment efficiently leads to endoplasmic reticulum
(ER) stress-induced apoptosis in cancer cells (4). ER stress-induced apoptosis is triggered
due to a failure in the folding of large amounts of REIC/Dkk-3
protein in the lumen of the ER, and the phosphorylation of JNK
occurs downstream of ER stress signaling (4,5). The
levels of phosphorylated JNK and c-Jun following Ad-REIC treatment
in bladder cancer cells were therefore analyzed in the present
study. Surprisingly, treatment with the Ad-REIC agent did not lead
to activation of JNK signaling in KK47 cells. The induction of
apoptosis and the inhibition of proliferation were observed in KK47
cells, indicating that the activation of JNK may not be essential
for the anti-proliferative effects of Ad-REIC. It is possible that
cancer suppressive mechanisms other than JNK signaling may underlie
the effects of Ad-REIC and that CD147 may be a novel target of
cancer therapeutic agents. Different cancer cells have their own
genetic characterization; however, the same phenomenon has been
observed in human prostate cancer LNCap cell lines (17).
With regard to the regulation of CD147 expression,
several signaling pathways are reported to be associated with its
transcription (18–23). The core promoter and transcription
factor-binding sites of CD147 were previously analyzed and
identified as targets of the cancer-associated transcription
factors c-Myc and Sp1 (22,23). Since the expression of CD147 is known
to be positively regulated by p38-, Erk1/2- and JNK-dependent MAPK
signaling and c-Myc protein, the associations between CD147
expression, MAPK pathway activation and the expression of c-Myc
following Ad-REIC treatment were examined in the present study.
Unexpectedly, no positive correlation was observed between the
expression of CD147 and the possible regulators that were assessed.
These results indicate that another signaling pathway that was not
examined in the present study could be responsible for the
downregulation of CD147.
Our group recently conducted a phase I/IIa study of
in situ gene therapy for patients with prostate cancer,
using an Ad-REIC agent produced according to good manufacturing
practice guidelines (30). Patients
with hormonal therapy-resistant prostate cancer with or without
metastasis were enrolled and evaluated to investigate the tumor
inhibitory effects of Ad-REIC treatment. The survival time of one
patient with progressive lymph node metastases was favorable, and
significant tumor killing effects were demonstrated in
Ad-REIC-injected lesions, with apparent reductions in tumor volume
(31). Although experimental studies
with cancer-bearing mouse models are required to investigate the
downregulation of CD147 in Ad-REIC-injected tumors, the therapeutic
effects observed in the treated legions could be partially
explained by the inhibition of the CD147 oncoprotein by the Ad-REIC
agent.
In conclusion, a novel therapeutic mechanism
underlies the effects of the Ad-REIC agent developed for cancer
gene therapy. In addition to the previously reported activation of
the JNK signaling pathway (5), the
downregulation of the cancer-progression factor CD147 could be one
of the major therapeutic effects of Ad-REIC gene therapy. The
mechanisms and the variety of roles of CD147 in cancer progression
make the oncoprotein an attractive target in the field of cancer
therapy. Further studies are required to elucidate the mechanisms
that regulate the expression of CD147.
Acknowledgements
This study was supported by scientific research
grants [Japan Society for the Promotion of Science Grants-in-Aid
for Scientific Research (KAKENHI); grant nos. JP15H04297,
JP15K10590 and JP16K11004] from the Ministry of Education, Culture,
Sports, Science and Technology of Japan and Takeda Science
Foundation. The authors would like to thank Mr. Hideo Ueki and Ms.
Fusaka Oonari (Okayama University) for their valuable assistance.
Okayama University and Momotaro-Gene Inc. are applying for patents
on the Ad-REIC agents. Drs Masami Watanabe and Yasutomo Nasu are
the inventors of the patents and own stock in Momotaro-Gene
Inc.
References
|
1
|
Dobruch J and Herr H: Should all patients
receive single chemotherapeutic agent instillation after bladder
tumour resection? BJU Int. 104:170–174. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hirata T, Watanabe M, Kaku H, Kobayashi Y,
Yamada H, Sakaguchi M, Takei K, Huh NH, Nasu Y and Kumon H:
REIC/Dkk-3-encoding adenoviral vector as a potentially effective
therapeutic agent for bladder cancer. Int J Oncol. 41:559–564.
2012.PubMed/NCBI
|
|
3
|
Watanabe M, Sakaguchi M, Kinoshita R, Kaku
H, Ariyoshi Y, Ueki H, Tanimoto R, Ebara S, Ochiai K, Futami J, et
al: A novel gene expression system strongly enhances the anticancer
effects of a REIC/Dkk-3-encoding adenoviral vector. Oncol Rep.
31:1089–1095. 2014.PubMed/NCBI
|
|
4
|
Watanabe M, Nasu Y and Kumon H:
Adenovirus-mediated REIC/Dkk-3 gene therapy: Development of an
autologous cancer vaccination therapy (Review). Oncol Lett.
7:595–601. 2014.PubMed/NCBI
|
|
5
|
Abarzua F, Sakaguchi M, Takaishi M, Nasu
Y, Kurose K, Ebara S, Miyazaki M, Namba M, Kumon H and Huh NH:
Adenovirus-mediated overexpression of REIC/Dkk-3 selectively
induces apoptosis in human prostate cancer cells through activation
of c-Jun-NH2-kinase. Cancer Res. 65:9617–9622. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nabeshima K, Iwasaki H, Koga K, Hojo H,
Suzumiya J and Kikuchi M: Emmprin (basigin/CD147): Matrix
metalloproteinase modulator and multifunctional cell recognition
molecule that plays a critical role in cancer progression. Pathol
Int. 56:359–367. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhong WD, Chen QB, Ye YK, Han ZD, Bi XC,
Dai QS, Liang YX, Zeng GH, Wang YS, Zhu G, et al: Extracellular
matrix metalloproteinase inducer expression has an impact on
survival in human bladder cancer. Cancer Epidemiol. 34:478–482.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pan Y, He B, Chen J, Sun H, Deng Q, Wang
F, Ying H, Liu X, Lin K, Peng H, et al: Gene therapy for colorectal
cancer by adenovirus-mediated siRNA targeting CD147 based on loss
of the IGF2 imprinting system. Int J Oncol. 47:1881–1889.
2015.PubMed/NCBI
|
|
9
|
Bovenzi CD, Hamilton J, Tassone P, Johnson
J, Cognetti DM, Luginbuhl A, Keane WM, Zhan T, Tuluc M, Bar-Ad V,
et al: Prognostic indications of elevated MCT4 and CD147 across
cancer types: A meta-analysis. Biomed Res Int. 2015:2424372015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hatanaka M, Higashi Y, Kawai K, Su J, Zeng
W, Chen X and Kanekura T: CD147-targeted siRNA in A375 malignant
melanoma cells induces the phosphorylation of EGFR and
downregulates cdc25C and MEK phosphorylation. Oncol Lett.
11:2424–2428. 2016.PubMed/NCBI
|
|
11
|
Liang YX, Lu JM, Mo RJ, He HC, Xie J,
Jiang FN, Lin ZY, Chen YR, Wu YD, Luo HW, et al: E2F1 promotes
tumor cell invasion and migration through regulating CD147 in
prostate cancer. Int J Oncol. 48:1650–1658. 2016.PubMed/NCBI
|
|
12
|
Le Floch R, Chiche J, Marchiq I, Naiken T,
Ilc K, Murray CM, Critchlow SE, Roux D, Simon MP and Pouysségur J:
CD147 subunit of lactate/H+ symporters MCT1 and hypoxia-inducible
MCT4 is critical for energetics and growth of glycolytic tumors.
Proc Natl Acad Sci USA. 108:16663–16668. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aït-Ali N, Fridlich R, Millet-Puel G,
Clérin E, Delalande F, Jaillard C, Blond F, Perrocheau L, Reichman
S, Byrne LC, et al: Rod-derived cone viability factor promotes cone
survival by stimulating aerobic glycolysis. Cell. 161:817–832.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marchiq I, Albrengues J, Granja S,
Gaggioli C, Pouysségur J and Simon MP: Knock out of the
BASIGIN/CD147 chaperone of lactate/H+ symporters disproves its
pro-tumour action via extracellular matrix metalloproteases (MMPs)
induction. Oncotarget. 6:24636–24648. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kimiya K, Naito S, Soejima T, Sakamoto N,
Kotoh S, Kumazawa J and Tsuruo T: Establishment and
characterization of doxorubicin-resistant human bladder cancer cell
line, KK47/ADM. J Urol. 148:441–445. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ueki H, Watanabe M, Kaku H, Huang P, Li
SA, Ochiai K, Hirata T, Noguchi H, Yamada H, Takei K, et al: A
novel gene expression system for detecting viable bladder cancer
cells. Int J Oncol. 41:135–140. 2012.PubMed/NCBI
|
|
17
|
Mori A, Watanabe M, Sadahira T, Kobayashi
Y, Ariyoshi Y, Ueki H, Wada K, Ochiai K, Li SA and Nasu Y: The
downregulation of the expression of CD147 by tumor suppressor
REIC/Dkk-3, and its implication in human prostate cancer cell
growth inhibition. Acta Med Okayama. 71:135–142. 2017.PubMed/NCBI
|
|
18
|
Hagemann T, Wilson J, Kulbe H, Li NF,
Leinster DA, Charles K, Klemm F, Pukrop T, Binder C and Balkwill
FR: Macrophages induce invasiveness of epithelial cancer cells via
NF-kappa B and JNK. J Immunol. 175:1197–1205. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang Z, Wang C, Wei L, Wang J, Fan Y,
Wang L, Wang Y and Chen T: Resveratrol inhibits EMMPRIN expression
via P38 and ERK1/2 pathways in PMA-induced THP-1 cells. Biochem
Biophys Res Commun. 374:517–521. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ge H, Zhang JF, Guo BS, He Q, Wang BY, He
B and Wang CQ: Resveratrol inhibits macrophage expression of
EMMPRIN by activating PPARgamma. Vascul Pharmacol. 46:114–121.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu H, Xu XF, Zhao Y, Tang MC, Zhou YQ and
Gao FH: NS-398 promotes pancreatic cancer cell invasion by CD147
and MMP-2 via the activation of P38. Mol Med Rep. 13:2208–2214.
2016.PubMed/NCBI
|
|
22
|
Kong LM, Liao CG, Zhang Y, Xu J, Li Y,
Huang W, Zhang Y, Bian H and Chen ZN: A regulatory loop involving
miR-22, Sp1, and c-Myc modulates CD147 expression in breast cancer
invasion and metastasis. Cancer Res. 74:3764–3778. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang L, Major T and Bocan T:
Characterization of the promoter of human extracellular matrix
metalloproteinase inducer (EMMPRIN). Gene. 282:75–86. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Baba M, Inoue M, Itoh K and Nishizawa Y:
Blocking CD147 induces cell death in cancer cells through
impairment of glycolytic energy metabolism. Biochem Biophys Res
Commun. 374:111–116. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang Q, Li J, Xing J, Li W, Li H, Ke X,
Zhang J, Ren T, Shang Y, Yang H, et al: CD147 promotes
reprogramming of glucose metabolism and cell proliferation in HCC
cells by inhibiting the p53-dependent signaling pathway. J Hepatol.
61:859–866. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marchiq I, Le Floch R, Roux D, Simon MP
and Pouyssegur J: Genetic disruption of lactate/H+ symporters
(MCTs) and their subunit CD147/BASIGIN sensitizes glycolytic tumor
cells to phenformin. Cancer Res. 75:171–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Muramatsu T: Basigin (CD147), a
multifunctional transmembrane glycoprotein with various binding
partners. J Biochem. 159:481–490. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fu ZG, Wang L, Cui HY, Peng JL, Wang SJ,
Geng JJ, Liu JD, Feng F, Song F, Li L, et al: A novel
small-molecule compound targeting CD147 inhibits the motility and
invasion of hepatocellular carcinoma cells. Oncotarget.
7:9429–9447. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Walter M, Simanovich E, Brod V, Lahat N,
Bitterman H and Rahat MA: An epitope-specific novel anti-EMMPRIN
polyclonal antibody inhibits tumor progression. Oncoimmunology.
5:e10780562015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kumon H, Ariyoshi Y, Sasaki K, Sadahira T,
Araki M, Ebara S, Yanai H, Watanabe M and Nasu Y: Adenovirus vector
carrying REIC/Dkk-3 gene: Neoadjuvant intraprostatic injection for
high-risk localized prostate cancer undergoing radical
prostatectomy. Cancer Gene Ther. 23:400–409. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kumon H, Sasaki K, Ariyoshi Y, Sadahira T,
Ebara S, Hiraki T, Kanazawa S, Yanai H, Watanabe M and Nasu Y:
Ad-REIC gene therapy: Promising results in a patient with
metastatic CRPC following chemotherapy. Clin Med Insights Oncol.
9:31–38. 2015. View Article : Google Scholar : PubMed/NCBI
|