Introduction
Colorectal cancer (CRC) is considered to be one of
the most frequent causes of cancer-related morbidity and mortality
worldwide (1,2). Although comprehensive strategies in the
treatment of CRC have been developed for a number of years, the
five-year survival rate of metastatic colon cancer is only 10%
(3). In the clinical treatment of
metastatic colon cancer, oxaliplatin is commonly used as an
essential chemotherapeutic agent, and not only improves the
response rate of patients, but also prolongs patients'
progression-free survival. Nevertheless, more than 40% of patients
still develop significant resistance (3,4).
Therefore, how to overcome resistance to oxaliplatin is a key
scientific problem to be solved in the treatment of metastatic
colon cancers.
Connexins are integral membrane proteins, six of
which make up a hemi-channel. Two hemi-channels in neighboring
cells dock together to form an integral gap junction (GJ). The GJ
enables cells to exchange ions and small molecules (with a molecule
weight less than 1 kDa) directly, including calcium, glutathione,
cyclic adenosine monophosphate and cyclic guanosine monophosphate.
Molecules transferred through the GJ are essential for numerous
physiological and pathological events (5,6). The
connexin gene family constitutes 21 isoforms, the most significant
of which is Cx43 (6). The loss of
Cx43 is extremely common in the development of cancers and is even
involved in advanced stages of tumor progression. The deficiency of
Cx43 also contributes to the resistance of chemotherapeutic agents
(7). It has been reported that
decreased levels of Cx43 GJ are associated with the resistance of
temozolomide and cisplatin targeting glioblastoma or lung
adenocarcinoma (8,9). Furthermore, Sirnes et al
indicated that Cx43 may be a CRC tumor suppressor protein that
predicts clinical outcome (7). Thus,
we supposed that the resistance of oxaliplatin to metastatic colon
cancers might be involved in the alternation of Cx43 GJ function,
and explored the mechanisms involved in this.
Reactive oxygen species are one of the significant
elements that may be transferred between neighboring cells through
the GJ (5). More significantly, Lin
et al observed that ROS-related mechanisms increased
oxaliplatin sensitivity in human colorectal cancers (10). Thus, in the present study, we present
the first evidence that ROS distribution mediated by Cx43
contributes to the resistance of oxaliplatin in the treatment of
human CRC.
Materials and methods
Cell line and cell culture
The human colon cancer cell line COLO320 was
obtained from the American Type Culture Collection (Manassas, VA,
USA) and cultured in RPMI-1640 medium (Invitrogen Life
Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (Invitrogen Life Technologies) and 100 U/ml
penicillin-streptomycin (Invitrogen Life Technologies) in a 5%
CO2 incubator with 90% humidity at 37°C (Thermo Fisher
Scientific, Waltham, MA, USA).
Colony-forming assay
Colony-forming assay was used to detect cytotoxicity
dependent on GJ. At high cell density culture, cells were seeded at
100,000 cells/cm2. When cells were exposed to drugs, the
cultures were 70 to 100% confluent and GJs were formed. At low cell
density culture, cells were seeded at 10,000 cells/cm2.
When cells were exposed to drugs, GJs were not formed. Cells were
treated with oxaliplatin (0–125 µM, Sigma-Aldrich, St. Louis, MO,
USA) for 24 h, and then washed with RPMI-1640 medium. Next, cells
were harvested by trypsinization (Invitrogen Life Technologies),
counted, diluted and seeded into six-well dishes at 100
cells/cm2. After 7 days, cells were stained with crystal
violet (Sigma-Aldrich) and assessed. Colonies containing more than
50 cells were scored (11).
Chemical treatment, lactate
dehydrogenase (LDH) and ROS assay
COLO320 cells were pretreated with connexin mimetic
peptide Gap26 at 300 µM for 1 h (Sigma-Aldrich) to inhibit Cx43
channel function, as well as retinoic acid (RA) at 10 µM for 24 h
(Sigma-Aldrich) to enhance Cx43 channel function prior to the
different assays. N-acetyl-L-cysteine (NAC; 5 mM, 24 h;
Sigma-Aldrich) and apocynin (AP; 10 µM, 24 h; Sigma-Aldrich) were
used to scavenge ROS. The corresponding solvent of these chemicals
was dimethyl sulfoxide (DMSO; Sigma-Aldrich). Cell growth was
determined via a colony-forming assay. LDH assays (Roche
Diagnostics, Indianapolis, IN, USA) were carried out according to
the manufacturer's instructions. Intracellular ROS production was
estimated using 2,7-dichlorofluorescein diacetate
(Sigma-Aldrich).
Cx43 knockdown with small interfering
RNA (siRNA) transfection
Two specific siRNAs (GCT GGT TAC TGG TGA CAGA and
CCG CAA TTA CAA CAA GCAA; siRNA1-Cx43 and siRNA2-Cx43) targeting
the Cx43 gene were designed to decrease Cx43 expression.
Nonspecific siRNA-Cx43 was used as a negative control (NC). The
siRNA and NC were purchased from Guangzhou RiboBio Co., Ltd.
(Guangzhou, China). Lipofectamine 2000 (Invitrogen Life
Technologies) was used to transfect siRNA and NC into the cells
according to the manufacturer's instructions (12).
‘Parachute’ dye-coupling assay
The ‘parachute’ dye-coupling method was used to
detect the GJ function. Donor cells were labeled with 5 µM CM-DiI
(Invitrogen Life Technologies) and 5 µM calcein-acetoxymethyl ester
(Invitrogen Life Technologies). CM-DiI did not spread to coupled
cells, but calcein-acetoxymethyl ester was able to transform into
the neighboring cells through the GJ. Donor cells were seeded onto
the receiver cells at a 1:150 donor/receiver ratio to form the GJ.
Four h later, GJ function was examined with a fluorescence
microscope (Eclipse E800, Nikon Corporation, Tokyo, Japan). The
average number of receiver cells containing dye per donor cell was
counted and normalized to that of control cultures (13).
Western blotting
Cells were washed three times with wash buffer [0.01
mol/l phosphate-buffered saline, 0.138 mol/l NaCl, 0.02%
NaN3 (pH 7.4)] and then incubated with lysis buffer for
2 h (Nanjing Keygen Biotech Co., Ltd., Nanjing, China) at 4°C using
0.05 ml/cm2. The bicinchoninic acid method (Nanjing
Keygen Biotech Co., Ltd.) was used to measure protein
concentrations. Cell lysates (30 µg) were separated by SDS-PAGE on
10% Tris-glycine mini-gels (Invitrogen Life Technologies) and
transferred onto polyvinylidene difluoride membrane (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). We used 5% non-fat dry milk
(Sigma-Aldrich) to block the membranes at room temperature for 30
min. After that, the membranes were immunoblotted using anti-Cx43
antibody (1:4,000; mouse monoclonal Cx43 antibody raised against
humans; cat. no. C8093; Sigma-Aldrich) overnight at 4°C. After
several washes, the membranes were incubated for 1 h at room
temperature with anti-mouse horseradish peroxidase (HRP)-conjugated
secondary antibodies (1:4,000; goat polyclonal antibody raised
against mouse IgG; cat. no. M6898; Sigma-Aldrich). Mouse monoclonal
anti-β-actin antibody raised against humans (cat. no. A1978;
Sigma-Aldrich) was immunoblotted (1:10,000) at 4°C overnight.
Anti-mouse HRP-conjugated secondary antibodies (goat polyclonal
antibody raised against mouse IgG; cat. no. M6898; Sigma-Aldrich)
was used at 1:10,000 for 1 h at room temperature. All protein bands
were detected with an enhanced chemiluminescence system (KGP1125;
Nanjing KeyGen Biotech. Co., Ltd.) and estimated using Alpha View
software (version 2.2.14407; ProteinSimple, Santa Clara, CA,
USA).
Statistical analysis
Statistical analysis was performed using SPSS 15.0
software (SPSS, Inc., Chicago, IL, USA). Multiple comparisons among
groups were analyzed using one-way analysis of variance, followed
by Tukey post hoc comparisons. P<0.05 was considered to indicate
a statistically significant difference.
Results
Effects of oxaliplatin on cell
survival, LDH release and intracellular ROS are all dependent on
cell density
In order to explore the effects of GJ function on
oxaliplatin cytotoxicity, COLO320, a type of human colon cancer
cell line, was cultured at low and high density. At low-density
cell culture, the cells were not in contact with each other and no
GJs were formed when exposed to different chemicals. However, at
high-density cell culture, cells were confluent, which allowed
significant formation of GJs. This method was therefore used to
explore the function of the GJ. Fig. 1A
and B reveal that when cells were in contact with one another
at high-density cell culture, oxaliplatin cytotoxicity was
substantially greater than at low-density cell culture. This
manifested as lower clonogenic survival and a greater extent of LDH
release in the high-density condition than in the low-density
condition. These results indicated that oxaliplatin cytotoxicity
was density-dependent, being more significant at high-density cell
culture where GJs were formed. As previously reported,
density-dependent cytotoxicity was always relative with ‘death
signals’ transferred between the neighboring cells via GJs: The
most important ‘death signals’ was ROS (5). Thus, we determined the intercellular ROS
of COLO320 at low- and high-density cell cultures when exposed to
oxaliplatin. Fig. 1C reveals that
intercellular ROS were increased more notably at high-density cell
culture than at low-density cell culture, which suggests that ROS
transfer mediated by GJ may play a significant part in oxaliplatin
cytotoxicity.
GJs affect cell survival, LDH release
and intracellular ROS of COLO320 cells exposed to oxaliplatin
Density-dependent cell cytotoxicity was always
regulated by the GJs, and Cx43 contributed to the cytotoxicity of
antitumor drugs. In order to explore the effects of Cx43 GJ on
oxaliplatin cytotoxicity targeting colon cancer cells, we used
different methods with distinct mechanisms to manipulate the
function of GJs composed of Cx43. Fig.
2A shows that the survival fraction of COLO320 cells detected
with colony-forming assay was decreased in different degrees when
subjected to oxaliplatin; however, at high-density cell culture
(where GJs formed), the cells were much more sensitive to this
commonly used chemotherapy drug. Moreover, the cytotoxicity of
oxaliplatin was attenuated subsequent to pretreatment with Gap26, a
specific inhibitor of Cx43 GJ, but exacerbated following RA
treatment. In contrast, at low-density cell culture (where GJs were
not formed), the cytotoxicity of oxaliplatin demonstrated no
significant difference when COLO320 cells were pretreated with
Gap26 or RA. When cells were pretreated with oxaliplatin, LDH
release and intracellular ROS of COLO320 cells increased
significantly. At high-density cell culture, the levels of LDH
release and intracellular ROS fluctuated as Cx43 GJ function
changed, and this was modulated by Gap26 and RA. However, at
low-density cell culture, the change in Cx43 GJ function had no
influence on the levels of LDH release and intracellular ROS. DMSO,
the vehicle control of Gap26 and RA, had no effect on the
parameters (Fig. 2B and C).
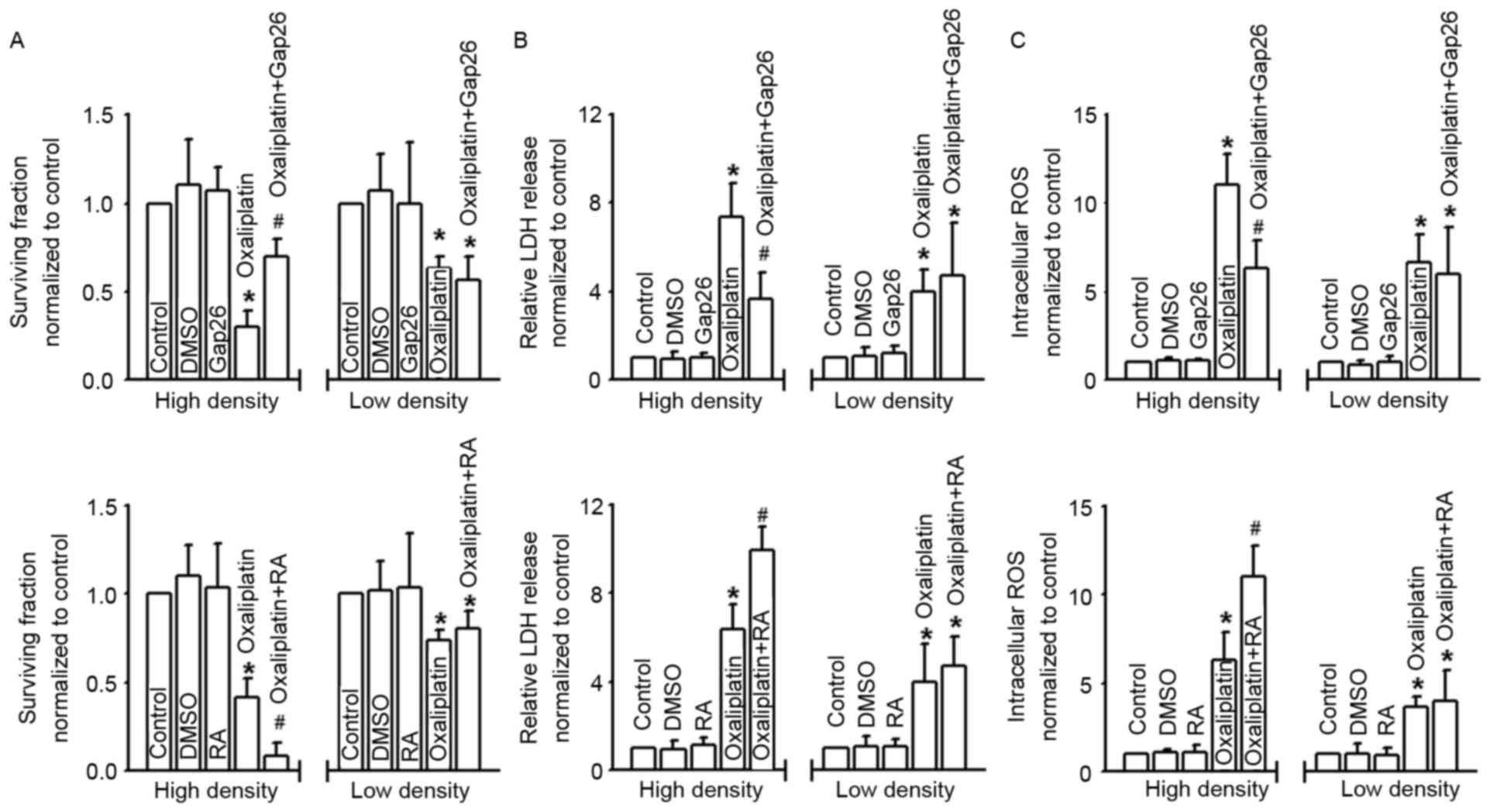 | Figure 2.Gap junctions composed of Cx43
regulate the cytotoxicity of oxaliplatin. (A-C) Clonogenic
survival, LDH release and intracellular ROS of COLO320 cells
exposed to oxaliplatin (100 µM, 24 h), at low or high density cell
culture and co-incubated with or without Gap26 and RA. n=4–6;
*P<0.05, compared with control group; #P<0.05,
compared with oxaliplatin group at high-density cell culture. Cx43,
connexin43; LDH, lactate dehydrogenase; ROS, reactive oxygen
species; DMSO, dimethyl sulfoxide; RA, retinoic acid. |
Cx43 gene knockdown attenuates COLO320
cell cytotoxicity of oxaliplatin
Figs. 1 and 2 illustrate that alternation of Cx43 GJ
function affected the cytotoxicity of oxaliplatin in COLO320 cells.
In order to confirm this result, we designed two different Cx43
siRNAs (siRNA1-Cx43 and siRNA2-Cx43) to knock down the expression
of Cx43 specifically (Fig. 3A). As
the expression of Cx43 was decreased, Cx43 GJ function was also
notably decreased (Fig. 3B). Cx43
knockdown did not influence COLO320 survival fraction, LDH release
and intracellular ROS per se, but Cx43 knockdown attenuated the
cytotoxicity induced by oxaliplatin on COLO320, and increased the
levels of oxaliplatin-induced LDH release and intracellular ROS,
all of which indicated that Cx43 GJ function played a significant
part in the cytotoxicity of oxaliplatin (Fig. 3C to E).
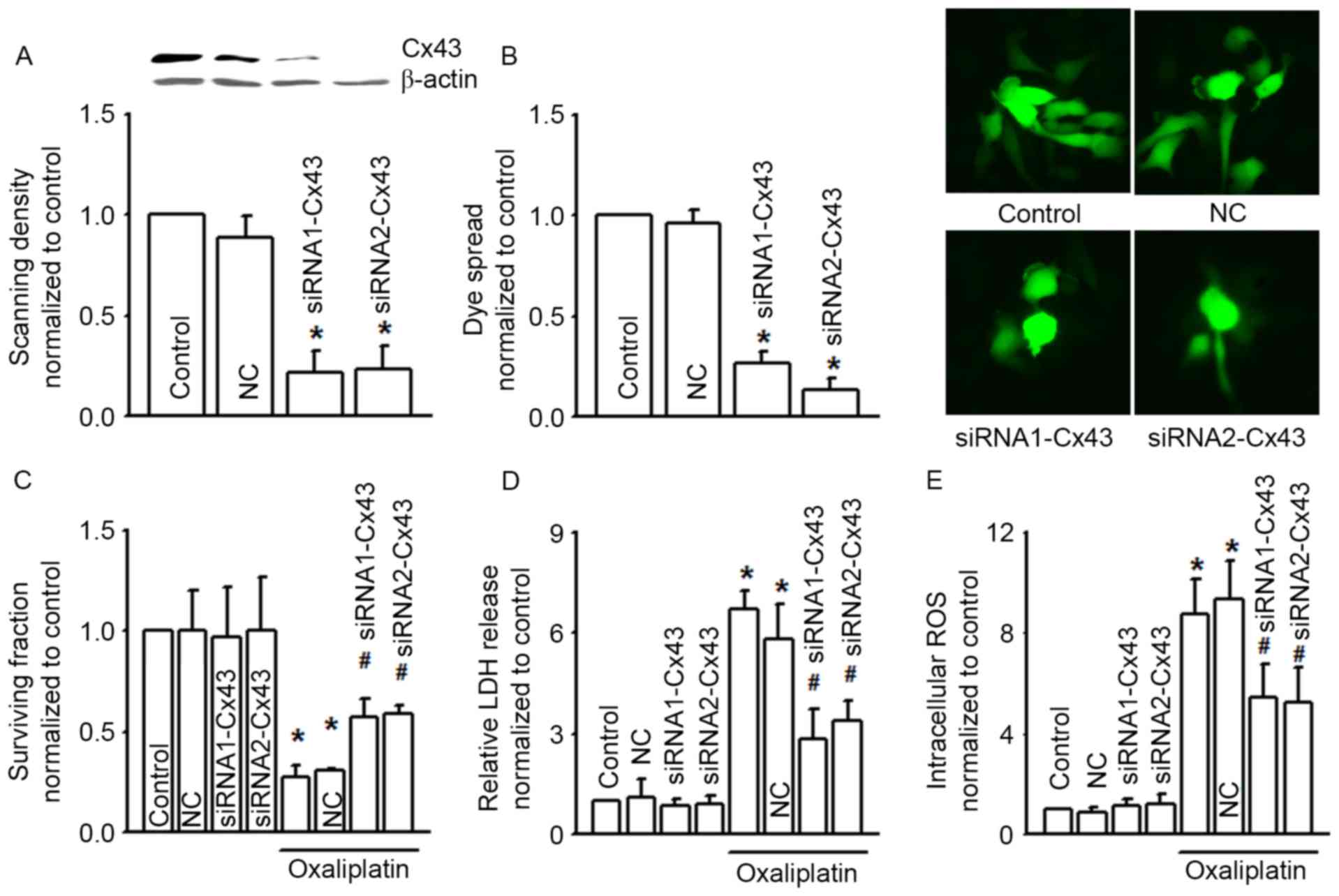 | Figure 3.siRNA alters clonogenic survival, LDH
release and intracellular ROS of COLO320 cells exposed to
oxaliplatin (100 µM, 24 h). (A) Two different specific siRNAs
(siRNA1-Cx43, siRNA2-Cx43) decreased Cx43 expression. n=3;
*P<0.05, compared with NC group. (B) The two different specific
siRNAs attenuated dye coupling of COLO320 cells. n=3; *P<0.05,
compared with NC group. (C-E) Clonogenic survival, LDH release and
intracellular ROS of COLO320 cells exposed to oxaliplatin (100 µM,
24 h), when pretreated with the two specific siRNAs. n=4–6;
*P<0.05, compared with control group; #P<0.05,
compared with NC group. siRNA, small interfering RNA; LDH, lactate
dehydrogenase; ROS, reactive oxygen species; Cx43, connexin43; NC,
negative control. |
The antioxidants NAC and AP attenuate
COLO320 cell cytotoxicity of oxaliplatin
It has been reported that the level of intracellular
ROS affects the cytotoxicity of oxaliplatin (10). Our results (Figs. 1C, 2C
and 3E) also revealed that Cx43 GJ
function alternation changed the level of oxaliplatin-induced
intracellular ROS. Thus, we supposed that modulation of the level
of intracellular ROS by Cx43 GJ might contribute to the
cytotoxicity of oxaliplatin. Fig. 4A
demonstrates that two antioxidants, NAC and AP, effectively
attenuated intracellular ROS increase induced by oxaliplatin.
Moreover, pretreatment of NAC and AP increased the survival
fraction of COLO320 cells, but decreased LDH release when exposed
to oxaliplatin. Both of the above indicate that inhibition of
intracellular ROS decreased the cytotoxicity of oxaliplatin
(Fig. 4B and C).
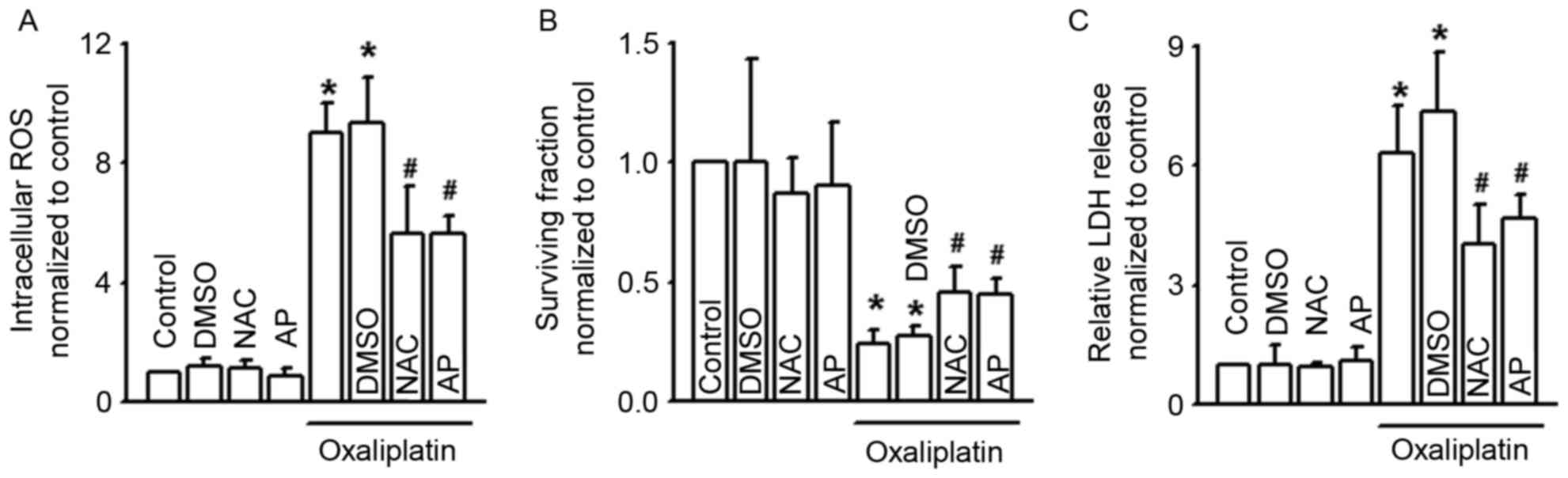 | Figure 4.NAC and AP alter the clonogenic
survival, LDH release and intracellular ROS of COLO320 cells
exposed to oxaliplatin. (A-C) Intracellular ROS, clonogenic
survival and LDH release of COLO320 cells exposed to oxaliplatin
(100 µM, 24 h) when pretreated with NAC (5 mM, 24 h) and AP (10 µM,
24 h). n=4–6; *P<0.05, compared with control group;
#P<0.05, compared with oxaliplatin group. NAC,
N-acetyl-L-cysteine; AP, apocynin; LDH, lactate dehydrogenase; ROS,
reactive oxygen species; DMSO, dimethyl sulfoxide. |
Discussion
The present study investigated the effect of Cx43 GJ
on the cytotoxicity of oxaliplatin in colon cancer cells, as well
as the mechanisms of its involvement in intracellular ROS
alternation. We demonstrated that oxaliplatin worked in a cell
density-dependent manner. At high-density cell culture (where GJs
were formed), the cytotoxicity of oxaliplatin was attenuated
subsequent to a decrease in Cx43 GJ function with the specific
inhibitor of Cx43 GJ, Gap26, but exacerbated as Cx43 GJ function
increased with the enhancer, RA. However, Gap26 and RA had no
effect at low-density cell culture (where no GJs were formed). In
conclusion, Cx43 plays a significant role in the cytotoxicity of
oxaliplatin in COLO320, a colon cancer cell line. Significantly, we
noted that intracellular ROS induced by oxaliplatin was also
modulated by Cx43 GJs: when Cx43 GJ function was enhanced,
intracellular ROS induced by oxaliplatin were increased; in
contrast, intracellular ROS were decreased as Cx43 GJ function was
attenuated. These results indicated that intracellular ROS
alternation modulated by Cx43 GJs might contribute to the
cytotoxicity of oxaliplatin in colon cancer cells.
CRC remains one of the leading causes of
cancer-related mortality worldwide (14). Although therapeutic strategies have
been improved in recent years, the five-year survival rate remains
less than 10% for metastatic colon cancer (3). Oxaliplatin is one of the most commonly
used chemotherapy drugs in colon cancer therapies. It is a
third-generation platinum-based anti-neoplastic agent commonly used
in the treatment of CRC (15,16). Oxaliplatin application in the clinic
has brought about notable improvements in the response rate and
progression-free survival in advanced colon cancer, but ~40% of
patients still develop resistance (3,17).
Resistance generation limits oxaliplatin application in the course
of clinical treatment. Thus, it is of vital clinical significance
to investigate the resistance of chemotherapy drugs.
It has been reported that loss of GJs in the
development of cancers contributes to the generation of resistance
to chemotherapy drugs (18–20). Enhancement of GJ function increases
the cytotoxicity of chemotherapy drugs (21). Conversely, inhibition of GJ function
decreases the cytotoxicity of chemotherapy drugs (22,23).
However, the mechanisms of these processes remain unclear. GJ
mediates the direct transfer of small molecules or electrical
charge between neighboring cells, generating various effects on the
different physiological and pathological issues processes. The most
significant of the transferred molecular signals is called the
‘death signal’, which results in the amplification of cytotoxicity
or apoptosis (11,24). Chemotherapy drugs attack the cancer
cells and result in cell death directly, and the attacked cells
generate various types of toxic products, called death signals. The
GJ is the main channel of death signals transferred between the
neighboring cells. Death signals not only attack the neighboring
cells directly, but also activate various signal pathways,
resulting indirectly in cytotoxicity or apoptosis (11,25). In
summary, the effects of chemotherapy drugs are amplified by the
transfer of death signals between the neighboring cells through the
GJ, which is called the ‘bystander effect’ (26,27). The
present study also supports this hypothesis, as the cytotoxicity of
oxaliplatin was attenuated subsequent to Cx43 GJ function decrease
with the specific inhibitor of Cx43 GJ, Gap26, but exacerbated as
Cx43 GJ function increased with the enhancer, RA.
Thus far, the intrinsic quality of death signals has
not been identified. Although the possibility of the involvement of
calcium or other cell metabolites has been discussed for a number
of years, direct proof is still lacking. We observed that ROS not
only enhanced oxaliplatin sensitivity to CRC, but also contributed
to epithelial-mesenchymal transition and cancer metastasis, which
indicated that ROS may play an essential role in the development of
cancers (10). Luo et al
reported that GJs composed of Cx32 regulated the distribution of
ROS (5). Therefore, we considered
that ROS distribution modulated by GJs might contribute to the
cytotoxicity of oxaliplatin. This hypothesis was supported by our
finding that GJ function inhibition decreased the level of ROS and
the cytotoxicity of oxaliplatin. Our results also indicated that
ROS may be significant death signals.
As reported in previous studies, downregulation of
Cx43 by siRNA resulted in a more aggressive growth of breast cancer
cells, and Cx43 knockout mice were susceptible to chemicals
inducing lung adenomas (22,23). In contrast, enhancement of Cx43
expression promoted cancer cell restoration and differentiation in
various types of tissue (27). Hence,
it may be noted that Cx43 is one of the most significant tumor
suppressors, and is associated with malignant behaviors of cancers.
Our study demonstrates for the first time that GJs composed of Cx43
affect the cytotoxicity of oxaliplatin by modulating the levels of
ROS in colon cancer cells. Moreover, Cx43 GJ function enhancement
amplified the oxaliplatin cytotoxicity of colon cancer cells. This
not only explains the possible mechanisms of resistance generation
from another aspect, but also provides a potential target in colon
cancer prevention and therapy.
References
|
1
|
Hirschi B, Gallmeier E, Ziesch A,
Marschall M and Kolligs FT: Genetic targeting of B-RafV600E affects
survival and proliferation and identifies selective agents against
BRAF-mutant colorectal cancer cells. Mol Cancer. 13:1222014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arnold M, Sierra MS, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global patterns and trends in
colorectal cancer incidence and mortality. Gut. 66:686–691. 2017.
View Article : Google Scholar
|
|
3
|
Tan S, Peng X, Peng W, Zhao Y and Wei Y:
Enhancement of oxaliplatin-induced cell apoptosis and tumor
suppression by 3-methyladenine in colon cancer. Oncol Lett.
9:2056–2062. 2015.PubMed/NCBI
|
|
4
|
Fan F, Gray MJ, Dallas NA, Yang AD, Van
Buren G II, Camp ER and Ellis LM: Effect of chemotherapeutic stress
on induction of vascular endothelial growth factor family members
and receptors in human colorectal cancer cells. Mol Cancer Ther.
7:3064–3070. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo C, Yuan D, Li X, Yao W, Luo G, Chi X,
Li H, Irwin MG, Xia Z and Hei Z: Propofol attenuated acute kidney
injury after orthotopic liver transplantation via inhibiting gap
junction composed of connexin 32. Anesthesiology. 122:72–86. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen X, Kong X, Zhuang W, Teng B, Yu X,
Hua S, Wang S, Liang F, Ma D, Zhang S, et al: Dynamic changes in
protein interaction between AKAP95 and Cx43 during cell cycle
progression of A549 cells. Sci Rep. 6:212242016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sirnes S, Bruun J, Kolberg M, Kjenseth A,
Lind GE, Svindland A, Brech A, Nesbakken A, Lothe RA, Leithe E and
Rivedal E: Connexin43 acts as a colorectal cancer tumor suppressor
and predicts disease outcome. Int J Cancer. 131:570–581. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gielen PR, Aftab Q, Ma N, Chen VC, Hong X,
Lozinsky S, Naus CC and Sin WC: Connexin43 confers Temozolomide
resistance in human glioma cells by modulating the mitochondrial
apoptosis pathway. Neuropharmacology. 75:539–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu M, Zhang C, Li L, Dong S, Zhang N and
Tong X: Cx43 reverses the resistance of A549 lung adenocarcinoma
cells to cisplatin by inhibiting EMT. Oncol Rep. 31:2751–2758.
2014.PubMed/NCBI
|
|
10
|
Lin S, Lei K, Du W, Yang L, Shi H, Gao Y,
Yin P, Liang X and Liu J: Enhancement of oxaliplatin sensitivity in
human colorectal cancer by hypericin mediated photodynamic therapy
via ROS-related mechanism. Int J Biochem Cell Biol. 71:24–34. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Q, You T, Yuan D, Han X, Hong X, He
B, Wang L, Tong X, Tao L and Harris AL: Cisplatin and oxaliplatin
inhibit gap junctional communication by direct action and by
reduction of connexin expression, thereby counteracting cytotoxic
efficacy. J Pharmacol Exp Ther. 333:903–911. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan D, Wang Q, Wu D, Yu M, Zhang S, Li L,
Tao L and Harris AL: Monocyte-endothelial adhesion is modulated by
Cx43-stimulated ATP release from monocytes. Biochem Biophys Res
Commun. 420:536–541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo C, Yuan D, Zhao W, Chen H, Luo G, Su G
and Hei Z: Sevoflurane ameliorates intestinal
ischemia-reperfusion-induced lung injury by inhibiting the
synergistic action between mast cell activation and oxidative
stress. Mol Med Rep. 12:1082–1090. 2015.PubMed/NCBI
|
|
14
|
Lee W, Belkhiri A, Lockhart AC, Merchant
N, Glaeser H, Harris EI, Washington MK, Brunt EM, Zaika A, Kim RB
and El-Rifai W: Overexpression of OATP1B3 confers apoptotic
resistance in colon cancer. Cancer Res. 68:10315–10323. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leone F, Artale S, Marino D, Cagnazzo C,
Cascinu S, Pinto C, Fornarini G, Tampellini M, Di Fabio F,
Sartore-Bianchi A, et al: Panitumumab in combination with
infusional oxaliplatin and oral capecitabine for conversion therapy
in patients with colon cancer and advanced liver metastases. The
MetaPan study. Cancer. 119:3429–3435. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Taieb J, Zaanan A, Le Malicot K, Julié C,
Blons H, Mineur L, Bennouna J, Tabernero J, Mini E, Folprecht G, et
al: Prognostic effect of BRAF and KRAS mutations in patients with
stage III colon cancer treated with leucovorin, fluorouracil, and
oxaliplatin with or without cetuximab: A post hoc analysis of the
PETACC-8 trial. JAMA Oncol. 1–11. 2016.PubMed/NCBI
|
|
17
|
Anderson BJ and Peterson LL: Systemic
capillary leak syndrome in a patient receiving adjuvant oxaliplatin
for locally advanced colon cancer. J Oncol Pharm Pract. 22:725–728.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Segretain D, Decrouy X, Dompierre J,
Escalier D, Rahman N, Fiorini C, Mograbi B, Siffroi JP, Huhtaniemi
I, Fenichel P and Pointis G: Sequestration of connexin43 in the
early endosomes: An early event of Leydig cell tumor progression.
Mol Carcinog. 38:179–187. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Leithe E, Sirnes S, Omori Y and Rivedal E:
Downregulation of gap junctions in cancer cells. Crit Rev Oncog.
12:225–256. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mesnil M, Crespin S, Avanzo JL and
Zaidan-Dagli ML: Defective gap junctional intercellular
communication in the carcinogenic process. Biochim Biophys Acta.
1719:125–145. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kardami E, Dang X, Iacobas DA, Nickel BE,
Jeyaraman M, Srisakuldee W, Makazan J, Tanguy S and Spray DC: The
role of connexins in controlling cell growth and gene expression.
Prog Biophys Mol Biol. 94:245–264. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shao Q, Wang H, McLachlan E, Veitch GI and
Laird DW: Down-regulation of Cx43 by retroviral delivery of small
interfering RNA promotes an aggressive breast cancer cell
phenotype. Cancer Res. 65:2705–2711. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Avanzo JL, Mesnil M, Hernandez-Blazquez
FJ, Mackowiak II, Mori CM, da Silva TC, Oloris SC, Gárate AP,
Massironi SM, Yamasaki H and Dagli ML: Increased susceptibility to
urethane-induced lung tumors in mice with decreased expression of
connexin43. Carcinogenesis. 25:1973–1982. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
He B, Tong X, Wang L, Wang Q, Ye H, Liu B,
Hong X, Tao L and Harris AL: Tramadol and flurbiprofen depress the
cytotoxicity of cisplatin via their effects on gap junctions. Clin
Cancer Res. 15:5803–5810. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hong X, Wang Q, Yang Y, Zheng S, Tong X,
Zhang S, Tao L and Harris AL: Gap junctions propagate opposite
effects in normal and tumor testicular cells in response to
cisplatin. Cancer Lett. 317:165–171. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao Y, Liu B, Wang Q, Yuan D, Yang Y,
Hong X, Wang X and Tao L: Propofol depresses the cytotoxicity of
X-ray irradiation through inhibition of gap junctions. Anesth
Analg. 112:1088–1095. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sanson M, Marcaud V, Robin E, Valéry C,
Sturtz F and Zalc B: Connexin 43-mediated bystander effect in two
rat glioma cell models. Cancer Gene Ther. 9:149–155. 2002.
View Article : Google Scholar : PubMed/NCBI
|















