Introduction
Indoleamine 2,3-dioxygenase (IDO) is an
intracellular enzyme that acts as an immunosuppressant (1). IDO serves a crucial role in the
induction of tolerance during infection, cancer and autoimmune
diseases (2,3). IDO expression is induced by interferon-γ
(IFN-γ) during inflammation (4). The
IDO pathway includes two related but distinct enzymes encoded by
genes IDO-1 and IDO-2 (5). The two
IDO enzymes may have a differential range of expression and
signaling pathways, and differ in their selectivity for certain
inhibitors (6,7). However, their specific contribution to
immune regulation is not fully understood.
Primary biliary cirrhosis (PBC) is an inflammatory
autoimmune disease that targets the biliary epithelial cells (BEC)
in the liver. Impaired IDO generation contributes to the progress
of autoimmunity in PBC (8). Defects
in the regulatory cell niche are vital to the advancement of
autoimmunity, but PBC is characterized by a quantitative decline in
the cluster of differentiation (CD)4+CD25+
forkhead box p3 (Foxp3+) lymphocyte [regulatory T-cell
(T-reg)] compartment that may be a key point in the pathogenesis of
disease (9). It has been speculated
that the destruction of the biliary tract in PBC is mediated by
auto-reactive CD4+, as well as CD8+ T cells
(10,11).
Overexpression of IDO may induce immunosuppression
and tolerance; this has been established following studies in
multiple animal models (1,12,13). IDO
has been shown to modulate immune response in two ways. Firstly, it
depletes tryptophan, which produces a cellular stress reaction
through the general control non-repressed 2 (GCN2) kinase pathway
(14), leading to the production of
tryptophan metabolites (15). These
metabolites contribute to the immunosuppressive effects of IDO by
inhibiting T-cell responses (16).
Secondly, IDO induces immunosuppression by producing kynurenine
(Kyn), a ligand for the aryl hydrocarbon receptor (15); this promotes differentiation of
Foxp3+ T-regs and decreases the immunogenicity of
dendritic cells (DCs) (12,13). It has been established that human T
cells are responsive to the anti-proliferative and cytotoxic
effects of supplemented Kyns (16,17). Kyns
suppress proliferation and have pro-apoptotic properties, primarily
in T-helper (Th) 1 lymphocytes, which respond to antigen
presentation (18,19). The mechanisms by which tryptophan
metabolites affect T cells are presently ambiguous; they may have
direct toxic effects or could bind to the receptor that induces
T-cell death (20).
The mutual outcome of tryptophan deprivation and
increased Kyn concentration depends on the GCN2 kinase-mediated
downregulation of the TCRζ-chain in CD8+ cells, reducing
their cytotoxic effects (21).
Furthermore, the depletion of tryptophan, together with the
increased generation of tryptophan metabolites, polarizes the
differentiation of naïve CD4+CD25− T cells
towards a regulatory phenotype (21).
IDO is upregulated by certain cytokines and
inflammatory molecules; however, its most potent stimulator is
IFN-γ (4). Transforming growth
factor-β (TGF-β) serves a pivotal role in establishing tolerance
and preventing autoimmunity (22).
IDO is an essential intermediary that links TGF-β production by DCs
with T-reg differentiation and the induction of tolerance (23). TGF-β (24), IL-10 (25) and nitric oxide (26) are negative regulators of IDO. With the
exception of IL-10, expression of these regulators in the liver of
PBC patients is increased (27,28).
In the present study, we hypothesized that IDO may
have a potential role in the pathogenesis of human PBC. High IDO
activity in the biliary epithelial H69 cell line may offer an
insight into the role of IDO in the pathogenesis of PBC. Thus, the
following were investigated: i) The functional enzymatic activity
of IDO in the cell line and serum samples of PBC patients and
controls; ii) the expression of IDO in control and PBC liver
tissues; and iii) whether Kyns modulate the human CD4+ T
cells polarization toward a T-reg phenotype.
Materials and methods
Patients
The present study was conducted with local regional
ethics committee approval, and written informed consent for
publication of this study was obtained from all patients. Samples
included serum collected for the purposes of research and excess
liver biopsy tissue from diagnostic procedures. The mean age of
patients was 58 years, with an age range between 35 and 66 years.
All samples were anonymized. To discount the variable effects due
to gender difference, all patients with PBC and all healthy
controls included in the study were female.
Cell culture
The H69 immortalized biliary epithelial cell (iBEC)
line was created and obtained from Grubman et al (29) (Tufts University School of Medicine,
Boston, MA, USA) from human intrahepatic biliary epithelial cells
and was used for the current study. These cells exhibit
characteristics of normal human biliary epithelium, with expression
of cytokeratin (CK)7 and 19. The iBECs were cultured in
75-cm2 flasks, at 37°C in an atmosphere containing 5%
CO2, in a 3:1 mix of Dulbecco's modified Eagle's medium
and Nutrient Mixture F12 Ham (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) supplemented with 1.8×10−4 M adenine (24.3
mg/l), 2×10−9 M triiodothyronine (1.345 µg/l),
5.5×10−6 M epinephrine (1.0 mg/l) (all from
Sigma-Aldrich; Merck KGaA), ITS-X supplement (10 mg/l insulin, 5.5
g/l transferrin, 2.0 g/l ethanolamine and 6.7 µg/l sodium selenite;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 1 µM
hydrocortisone solution (362.46 µg/l), 10% heat-inactivated fetal
bovine serum and 100 U/ml each of penicillin and streptomycin (all
from Sigma-Aldrich; Merck KGaA).
H69 cells treated with IFN-γ
H69 cells were treated with 20 ng/ml IFN-γ. Once
confluent (>80%), the cells were then left post- IFN-γ treatment
for 0, 6, 12, 24, 48, 72, 96 and 120 h prior to RNA isolation or
harvesting of supernatants.
H69 cells treated with TGF-β type-1
receptor serine/threonine kinase (ALK5) inhibitor
H69 cells were incubated with an ALK5 inhibitor
(SB-505124; catalog no. 3263; TOCRIS Bioscience, Bristol, UK) at an
optimal concentration of 1 µΜ (30)
for 1 h prior to stimulation with IFN-γ for 6 and 24 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA isolation was performed according to the method
developed by Chomczynski and Sacchi (31). TRIzol reagent (Sigma-Aldrich; Merck
KGaA) was used according to the manufacturer's instructions. RNA
was quantified and quality-assessed using a NanoDrop
spectrophotometer (NanoDrop ND-1000; Thermo Fisher Scientific,
Logan, UT, USA). Isolated RNA was reverse transcribed to cDNA using
the AffinityScript Multi Temperature cDNA Synthesis kit (Agilent
Technologies, Inc., Santa Clara, CA, USA) according to the
manufacturer's protocol. The qPCR experiments in this project
utilized TaqMan chemistry and were performed in a MicroAmp Optical
96-well plate (both from Applied Biosystems; Thermo Fisher
Scientific, Inc.) on a StepOnePlus Real-Time PCR machine (Applied
Biosystems; Thermo Fisher Scientific, Inc.), with each well
containing 1 µl TaqMan primer-probe (all primers were
exon-spanning), 1 µl cDNA, 10 µl Master Mix and 8 µl RNase-free
water. Primers used were for GAPDH (catalog no. Hs02758991_g1),
IDO-1 (catalog no. Hs00984148_m1), IDO-2 (catalog no.
Hs01589373_m1), TGF-β1 (catalog no. Hs00998133_m1), TGF-β2 (catalog
no. Hs00234244_m1) and were all purchased from Applied Biosystems;
Thermo Fisher Scientific, Inc. The thermocycling conditions were as
follows: 50°C for 1 min, 95°C for 10 min, followed by 40 cycles of
95°C for 15 sec and 60°C for 1 min. Data were normalized to the
expression of GAPDH mRNA using the 2−ΔΔCq method
(32) and analyzed using REST 2009
software (Qiagen GmbH, Hilden, Germany).
Immunohistochemistry
Formalin-fixed, paraffin-embedded liver biopsy
sections from patients were deparaffinized in xylene and
rehydrated. Antigen retrieval was conducted by pressure-cooking
(pressure, 15PSI; temperature, 121°C) for 1 min in Tris/EDTA (pH
8.0). To block endogenous peroxidase activity, 3%
H2O2 in methanol was utilized and a biotin
block was performed according to the manufacturer's protocol
(Avidin/Biotin Blocking kit; catalog no. SP-2001; Vector
Laboratories, Ltd., Peterborough, UK). Sections were incubated
either with anti-IDO antibody (catalog no. ab55305; dilution,
1:100) or anti-Foxp3 antibody (catalog no. ab20034; dilution,
1:100) (both from Abcam, Cambridge, UK) in normal swine serum at
4°C, in a humidified chamber overnight. Following washing with 0.1%
Tween-20 in PBS, sections were incubated with biotinylated
anti-mouse secondary antibody (catalog no. BA-9200; dilution,
1:200; Vector Laboratories, Ltd.) for 1 h at room temperature in a
humidified chamber. Sections were then stained with Vectastain
peroxidase ABC kit (Vector Laboratories, Ltd.), according to the
manufacturer's protocol. Staining was developed with
3,3′-diaminobenzidine (Sigma-Aldrich; Merck KGaA) and
counterstained with Mayer's hematoxylin (Dako; Aglient
Technologies, Inc.), according to the manufacturers' protocols.
Sections stained with no primary antibody was used as a negative
control. Visualization was performed using an optical microscope
(Provis AX-70; Olympus Corporation, Tokyo, Japan).
High-performance liquid chromatography
(HPLC)
HPLC was performed on a Shimadzu LC-10ADVP system
(Shimadzu Corporation, Kyoto, Japan). The separation was performed
isocratically using a Cronusil-S ODS1 (250×4.6-mm) column
(SMI-LabHut, Ltd., Gloucester, UK) at 25°C at a flow rate of 1
ml/min. The mobile phase contained 15 mM sodium acetate (pH 5.0)
and acetonitrile [94:6% (v/v)]. Following injection (25 µl), the
eluted Kyn was monitored at 360 nm between 0 and 8.5 min, and after
8.5 min, tryptophan was monitored at 278 nm. A working-top standard
solution containing 10 µM Kyn and 100 µM tryptophan was made in
mobile phase and further diluted to create a standard curve.
Standard or sample (200 µl) was mixed with 50 µl sulfosalicylic
acid, incubated on ice for 15 min and clarified by centrifugation
for 15 min at 18,000 × g at 4°C. Supernatant (25 µl) was injected
into the HPLC system. Kyn typically eluted at ~6.5 min and
tryptophan at 9.4 min. Standard curve and sample quantification was
based on peak area using LCSolution software (version 1.11SP1;
Shimadzu Corporation).
Colorimetric assay
Kyn was measured spectrophotometrically, as
described previously (33). A total
of 75 µl 30% trichloroacetic acid was added to 100 µl of the
culture supernatant, vortexed and centrifuged at 10,000 × g for 5
min at 4°C. Following this, a 75-µl volume of the supernatant was
then added to an equal volume of Ehrlich's reagent (100 mg
dimethylbenzaldehyde and 5 ml glacial acetic acid) in a microtiter
plate well. Optical density was measured at 492-nm using a
Multiskan MS microplate reader (Thermo Labsystems, Santa Rosa, CA,
USA). A standard curve of defined Kyn concentration (0–100 µM)
permitted analysis of unknowns.
Tryptophan metabolites
3-Hydroxykynurenine (3-HK), Kyn and
3-hydroxyanthranilic acid (3-HAA) (all from Sigma-Aldrich; Merck
KGaA) were dissolved in RPMI-1640 medium and added to
CD4+ T cells in culture at concentrations between 0 and
100 µM (pH 7.5–8.5).
Isolation of human CD4+
T-cells
Isolation of human CD4+ T cells was
performed using Rosette Sep (Stemcell Technologies, Vancouver, BC,
Canada) human CD4+ T-cell enrichment cocktail at 50
µl/ml of whole blood (500 µl/10 ml), according to the
manufacturer's protocol. Briefly, the whole blood was taken from a
healthy donor and incubated with the cocktail for 20 min at room
temperature. The sample was diluted with an equal volume of PBS
(plus 2% FBS) and mixed gently. This was layered on top of the
density medium and centrifuged at 1,200 × g for 20 min at room
temperature. The enriched cells were isolated from the density
medium-plasma interface and washed with PBS with 2% FBS. Isolated
CD4+ T cells (1×105 cells/well) were
stimulated with anti-human CD3 and CD28 beads at the ratio of one
bead per cell (Dynabeads Human T-activator; Invitrogen; Thermo
Fisher Scientific, Inc.) in the presence of tryptophan metabolites,
3-HK, 3-HAA and L-Kyn (Sigma-Aldrich; Merck KGaA) at 50 µΜ
concentration, with 10 ng/ml TGF-β1 (R&D Systems Europe, Ltd.,
Abingdon, UK) used as a control; cells were incubated for 5
days.
Intracellular staining of T-regulatory
cells
Intracellular staining for Foxp3 was performed using
a Foxp3 intracellular staining kit (eBioscience; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. This
kit has been formulated and optimized to stain intracellular
antigen Foxp3 (clone: PCH101; allophycocyanin, catalog no.
17-4776-42; eBioscience™, Invitrogen; Thermo Fisher Scientific,
Inc.). Cells were stained for intracellular staining. Briefly,
cells suspended in FACS tubes were pelleted by centrifugation at
500 × g for 5 min at 4°C. Following removal of the supernatant,
cells were fixed in a fixation/permeabilization buffer (1-part
fixation/permeabilization concentrate to 3 parts
fixation/permeabilization diluent) for 30 min at 4°C. Cells were
then washed and centrifuged in permeabilization buffer at 500 × g
for 5 min (at 4°C) followed by 1-h incubation with the primary
antibody. Finally, cells were washed twice in permeabilization
buffer prior to re-suspension in FACS buffer (PBS with 2% FBS) and
analyzed using a FACSCanto II flow cytometer and BD FACSDiva™
software, version 7.0 (both from BD Biosciences, Franklin Lakes,
NJ, USA).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA). Unless
otherwise noted, data represent the mean value of an experiment
conducted in triplicate ± standard error of the mean. For
comparison between 2 groups, unpaired two-tailed Students t-tests
or Mann Whitney unpaired t-test were performed. For ≥3 groups, a
one-way analysis of variance test was performed and, if
significant, a post-hoc Tukey's analysis was performed. P<0.05
was considered to indicate a statistically significant
difference.
Results
Expression of IDO in iBECs
A prior study established an in vitro model
using iBECs to investigate the link between senescence and
epithelial cell de-differentiation (30). The present study assessed the
expression of mRNA encoding IDO-1, IDO-2, TGF-β1 and TGF-β2
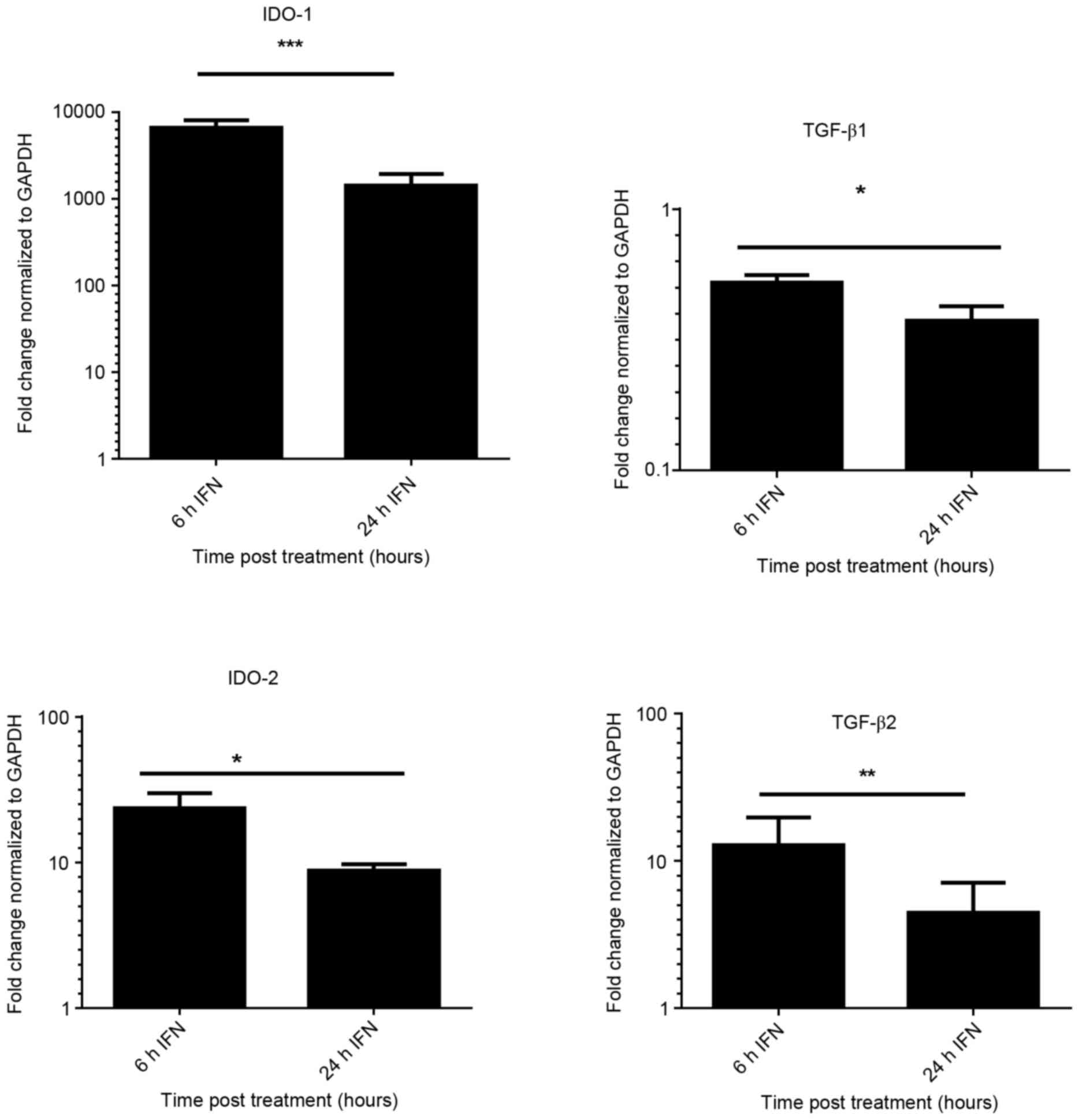
following stimulation of iBEC with IFN-γ, for 6 and 24 h. Fig. 1 demonstrates the significant increase
(P<0.05) in the expression of IDO-1 (6,644-fold) with IFN-γ at 6
h. The results shown were normalized against the housekeeping gene
GAPDH. High levels of IDO-1 expression were sustained at 24 h in
IFN-γ treated cells; whereas no detectable expression of IDO-1 was
found in untreated iBECs (data not shown).
Increase in IDO-2 expression was observed in cells
treated with IFN-γ (P<0.01). Expression of TGF-β2 was also
upregulated (12-fold) upon treatment with IFN-γ at 6 h, but
expression was reduced at 24 h. There was no significant increase
in expression of TGF-β1 following stimulation with IFN-γ; however,
TGF-β2 expression was significantly increased.
To determine whether IDO-1 expression is dependent
on the TGF-β pathway, cells were incubated with an ALK5 inhibitor
(SB-505124) at an optimal concentration of 1 µΜ (30) for 1 h prior to stimulation with IFN-γ
for 6 and 24 h. The expression of IDO-1 was not significantly
different in the treated or the untreated group (P>0.05, data
not shown). These findings suggest that IDO-1 expression in iBEC is
inducible, with a marked increase following IFN-γ stimulation, but
independent of the TGF-β pathway.
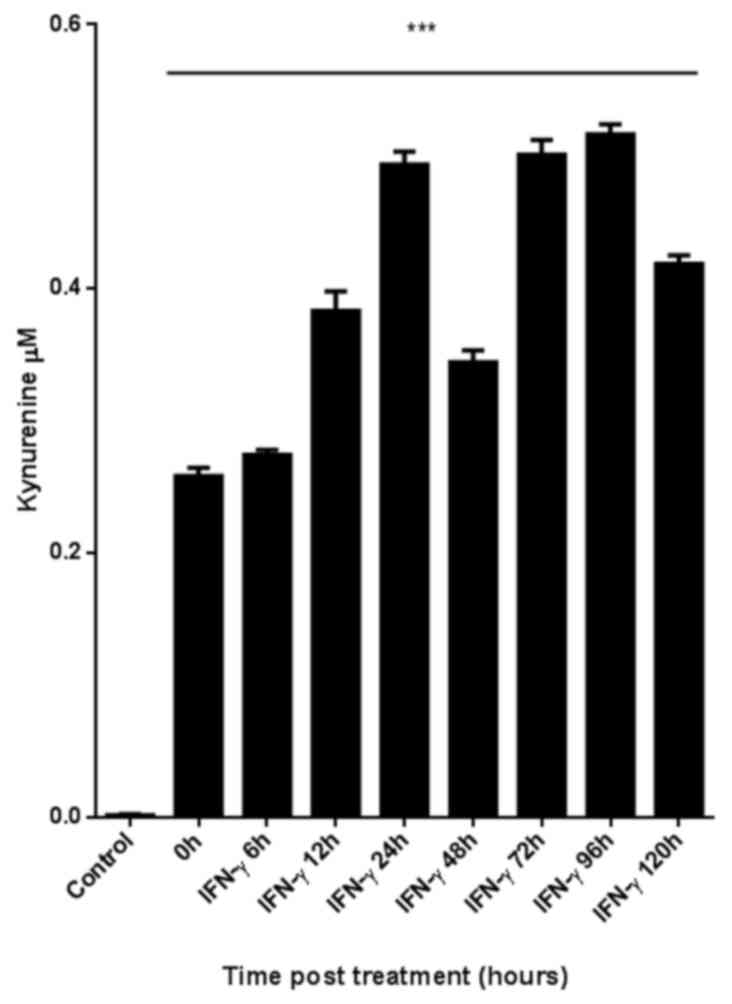
Enzymatic activity of IDO is
upregulated by IFN-γ stimulation in iBECs
Certain studies have revealed an inconsistency
between the expression of IDO and its enzymatic activity (34,35); thus,
the present study examined the enzymatic activity of IDO in iBECs.
A colorimetric assay was used to measure the levels of Kyn in the
culture supernatant (Fig. 2). Cells
were stimulated with IFN-γ for up to 120 h. The supernatant of
unstimulated iBEC produced no Kyn. The difference in Kyn levels
between the IFN-γ-stimulated and the unstimulated iBECs was
significant (P<0.05).
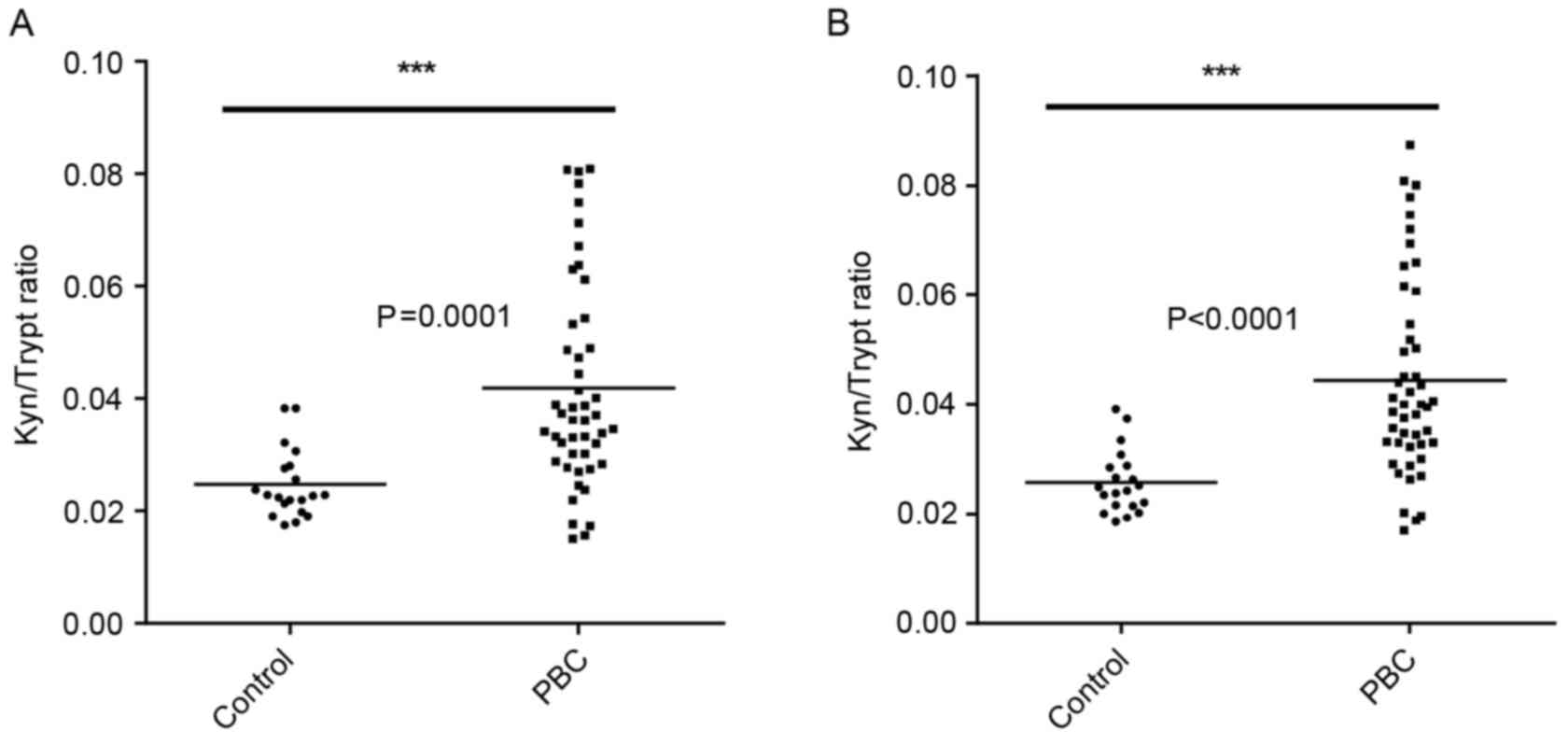
IDO activity in PBC patient sera
IDO activity was measured by assessing the ratio of
the enzyme substrate tryptophan and its product Kyn (Fig. 3). Higher IDO activity was observed in
the sera of PBC patients (n=47) compared with the healthy controls
(n=24). Statistically significant differences were found between
the two groups when measuring the area and height of peaks as
determined by HPLC (Fig. 3).
Expression of IDO in BECs from the
patients with PBC
To assess IDO expression in PBC, an
immunohistochemical assay was performed. Expression of IDO in the
liver tissue was examined in samples from healthy controls (n=5)
and patients with PBC (n=5). No expression was observed in the
cholangiocytes of healthy controls (Fig.
4A). Fig. 4B-D reveals IDO
expression in late-stage PBC. Discrete staining is revealed in the
BECs (Fig. 4B). IDO expression was
increased in periportal hepatocytes, in conjunction with notable
interface hepatitis (Fig. 4C and D).
Sections from matched PBC patients stained for Foxp3 are presented
in Fig. 4E and F; Foxp3+
cells were morphologically small lymphocytes. There was an evident
increase in the number of inflammatory cells, particularly those
with interface hepatitis and IDO expression.
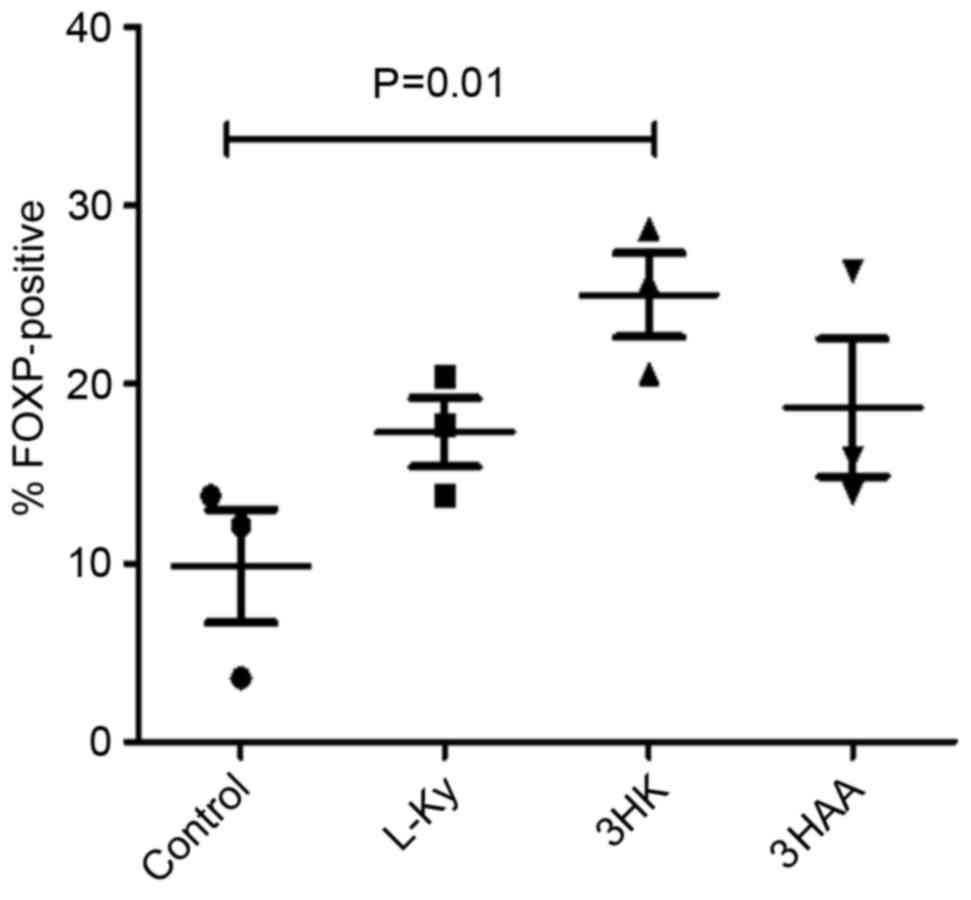
Tryptophan metabolites induce the
T-reg phenotype from the whole human CD4+ T cell
population
IDO may induce T-regs, at least in part, through
certain tryptophan metabolites. To test this, human CD4+
T cells were stimulated with anti-CD3/CD28 beads in the presence of
50 µM of the Kyns, L-Kyn, 3-HK and 3-HAA for 5 days. T-reg
development was assessed by analyzing Foxp3 expression with flow
cytometry.
There was no significant difference in the
proportion of CD4+ cells expressing Foxp3 in the
presence of L-Kyn and 3-HAA compared with the control after 5 days
(Fig. 5); however, 3-HK was
significantly upregulated in the fraction of CD4+ cells
expressing Foxp3.
Discussion
To the best of our knowledge, the current study
establishes the expression of IDO in H69 iBECs for the first time,
and reveals the increased rate of conversion of tryptophan to Kyn
in patients with PBC. Furthermore, clinical biopsies from PBC
patients demonstrated that the expression of IDO was observed not
only in BECs, as described previously (8), but also in hepatocytes.
The transcriptional profile of IDO in iBECs was
assessed following direct stimulation with IFN-γ, and levels were
found to be significantly increased. IFN-γ induced IDO expression
in the iBECs. This suggests that the expression of IDO in iBECs is
highly responsive to IFN-γ stimulation. Data from the present study
is in agreement with earlier studies, which revealed that IFN-γ can
induce IDO expression in several cell types and is produced by
inflammatory cells in response to immune activation (36–38). This
expression occurs during infectious, autoimmune and malignant
diseases (2,3).
Although the majority of past research has focused
on IDO as the central and immunobiologically relevant enzyme that
catalyzes the conversion of tryptophan to Kyn, there are two
additional enzymes, IDO-2 and tryptophan-2,3-dioxygenase (TDO),
which can catalyze the same enzymatic reaction. TDO was first
detected in the liver; however, little is known about the role of
TDO in tumorigenesis or its association with IDO (39). IDO-2 was recently identified and has
slightly different expression pattern to IDO-1 (6). In the present study, the regulation of
IDO-2 was examined in response to IFN-γ. Unstimulated BECs do not
express IDO-2; however, a significant increase in IDO-2 expression
was observed following treatment with IFN-γ.
TGF-β is an immunoregulatory cytokine produced by
activated T cells that mediates its effect through TGF-β receptors
1 and 2. A previous study demonstrated that CD8− DCs can
induce IDO in response to TGF-β which can modify the cells from
immunogenic into a tolerogenic state (40). Upon examination of the expression of
TGF-β receptors 1 and 2, TGF-β receptor 1 levels were not
significantly altered, whereas TGF-β receptor 2 levels were
significantly increased in response to IFN-γ. Furthermore,
incubation of cells with the non-toxic pharmacological TGF-β
receptor 1 inhibitor, SB-505124, followed by stimulation with IFN-γ
had no effect on IDO-1 expression (data not shown). This suggests
that, in BECs, the induction of IDO is IFN-γ-dependent, but
independent of the TGF-β pathway.
The enzymatic activity of IDO was examined in H69
cells in the present study. Increased Kyn production and reduced
substrate tryptophan demonstrated that IDO was enzymatically
functional in H69 cells. Additionally, significantly higher IDO
activity was observed in sera derived from PBC patients compared
with that from healthy controls. However, this pathway is extremely
responsive to non-specific inflammation and is induced in all
states of chronic inflammation, as IDO is an extremely sensitive
IFN-γ responsive gene.
Liver sections from PBC patients were examined and
it was confirmed that IDO is expressed in hepatocytes, as it is in
other epithelial cells (41,42). Low-level expression of IDO was
identified within BECs and strong expression was observed in
hepatocytes, which indicates its potential role in PBC. Liver
sections from matched patients with PBC expressed Foxp3. There was
an increase in the number of inflammatory cells, particularly those
exhibiting interface hepatitis and IDO expression. IDO was
recognized in early stage PBC samples and within damaged
cholangiocyte aggregates, identifying its role in immunogenicity
and indicating its involvement in PBC (8).
The tryptophan metabolite 3-HK significantly
upregulated Foxp3 in human CD4+ T cells, thus polarizing
the response towards a T-reg phenotype. In agreement with the data
from the current study, prior research by Fallarino et al
(21), found that tryptophan
catabolites downregulate the expression of T-cell receptor-ζ-chain
and induce a regulatory phenotype in naïve T cells.
IDO activity is increased in autoimmune disorders
and inflammation (43). In animal
models of autoimmunity, such as that of experimental autoimmune
encephalomyelitis (EAE), the presence of IDO-produced tryptophan
metabolites results in the skewing of a Th1 immune response towards
a Th2 response, which results in reduced inflammation and tissue
damage. Notably, a tryptophan metabolite, 3,4,'-dimethoxycinnamoyl
anthranilic acid, was able to reverse paralysis in mice with EAE
(44). IDO immunosuppressive action
serves as a negative feedback effect induced by hyper-activation of
T-cell immunity. In the present study, although the Kyn/tryptophan
ratio was increased in PBC patients, it is possible the level of
IDO expression was not sufficient to produce the desired
immunosuppression. This raises the possibility that Kyn metabolites
could be used in disease resolution.
In conclusion, the present study demonstrated that
expression of IDO in iBECs was stimulated by IFN-γ. The induction
of IDO at the tissue level may serve a role in the pathogenesis of
PBC. The effect of tryptophan metabolites on human CD4+
T cells in inducing polarization towards a T-reg phenotype suggests
that there exists a therapeutic opportunity for the management of
PBC. Furthermore, tryptophan catabolism could act as a potential
biomarker to monitor disease progression in PBC.
Acknowledgements
The present study was supported by grants from the
Marie Curie Innovative Doctoral Programme (grant no.
FP7-PEOPLE-2013-ITN), and Medical Research Council UK Primary
Biliary Cirrhosis Programme (grant no. MR/L001489/1).
Glossary
Abbreviations
Abbreviations:
|
IDO
|
indoleamine 2,3-dioxygenase
|
|
PBC
|
primary biliary cirrhosis
|
|
iBEC
|
immortalized, but non-malignant,
biliary epithelial cell line
|
|
HPLC
|
high-performance liquid
chromatography
|
|
IFN-γ
|
interferon-γ
|
|
TGF-β
|
transforming growth factor-β
|
|
BEC
|
biliary epithelial cells
|
|
GCN2
|
general control non-repressed 2
|
|
3-HAA
|
3-hydroxyanthranilic acid
|
|
Kyn
|
kynurenine
|
|
TDO
|
tryptophan-2,3-dioxygenase
|
|
EAE
|
experimental autoimmune
encephalomyelitis
|
References
|
1
|
Mellor AL and Munn DH: IDO expression by
dendritic cells: Tolerance and tryptophan catabolism. Nat Rev
Immunol. 4:762–774. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Uyttenhove C, Pilotte L, Théate I,
Stroobant V, Colau D, Parmentier N, Boon T and Van den Eynde BJ:
Evidence for a tumoral immune resistance mechanism based on
tryptophan degradation by indoleamine 2,3-dioxygenase. Nat Med.
9:1269–1274. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Curti A, Aluigi M, Pandolfi S, Ferri E,
Isidori A, Salvestrini V, Durelli I, Horenstein AL, Fiore F,
Massaia M, et al: Acute myeloid leukemia cells constitutively
express the immunoregulatory enzyme indoleamine 2,3-dioxygenase.
Leukemia. 21:353–355. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Däubener W and MacKenzie CR: IFN-gamma
activated indoleamine 2,3-dioxygenase activity in human cells is an
antiparasitic and an antibacterial effector mechanism. Adv Exp Med
Biol. 467:517–524. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ball HJ, Sanchez-Perez A, Weiser S, Austin
CJ, Astelbauer F, Miu J, McQuillan JA, Stocker R, Jermiin LS and
Hunt NH: Characterization of an indoleamine 2,3-dioxygenase-like
protein found in humans and mice. Gene. 396:203–213. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Metz R, Duhadaway JB, Kamasani U,
Laury-Kleintop L, Muller AJ and Prendergast GC: Novel tryptophan
catabolic enzyme IDO2 is the preferred biochemical target of the
antitumor indoleamine 2,3-dioxygenase inhibitory compound
D-1-methyl-tryptophan. Cancer Res. 67:7082–7087. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lob S, Konigsrainer A, Schafer R,
Rammensee HG, Opelz G and Terness P: Levo- but not dextro-1-methyl
tryptophan abrogates the IDO activity of human dendritic cells.
Blood. 111:2152–2154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oertelt-Prigione S, Mao TK, Selmi C,
Tsuneyama K, Ansari AA, Coppel RL, Invernizzi P, Podda M and
Gershwin ME: Impaired indoleamine 2,3-dioxygenase production
contributes to the development of autoimmunity in primary biliary
cirrhosis. Autoimmunity. 41:92–99. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lan RY, Cheng C, Lian ZX, Tsuneyama K,
Yang GX, Moritoki Y, Chuang YH, Nakamura T, Saito S, Shimoda S, et
al: Liver-targeted and peripheral blood alterations of regulatory T
cells in primary biliary cirrhosis. Hepatology. 43:729–737. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shimoda S, Van de Water J, Ansari A,
Nakamura M, Ishibashi H, Coppel RL, Lake J, Keeffe EB, Roche TE and
Gershwin ME: Identification and precursor frequency analysis of a
common T cell epitope motif in mitochondrial autoantigens in
primary biliary cirrhosis. J Clin Invest. 102:1831–1840. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kita H, Matsumura S, He XS, Ansari AA,
Lian ZX, Van de Water J, Coppel RL, Kaplan MM and Gershwin ME:
Quantitative and functional analysis of PDC-E2-specific
autoreactive cytotoxic T lymphocytes in primary biliary cirrhosis.
J Clin Invest. 109:1231–1240. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grohmann U, Fallarino F and Puccetti P:
Tolerance, DCs and tryptophan: Much ado about IDO. Trends Immunol.
24:242–248. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Munn DH, Sharma MD and Mellor AL: Ligation
of B7-1/B7-2 by human CD4+ T cells triggers indoleamine
2,3-dioxygenase activity in dendritic cells. J Immunol.
172:4100–4110. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Munn DH, Sharma MD, Baban B, Harding HP,
Zhang Y, Ron D and Mellor AL: GCN2 kinase in T cells mediates
proliferative arrest and anergy induction in response to
indoleamine 2,3-dioxygenase. Immunity. 22:633–642. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mezrich JD, Fechner JH, Zhang X, Johnson
BP, Burlingham WJ and Bradfield CA: An interaction between
kynurenine and the aryl hydrocarbon receptor can generate
regulatory T cells. J Immunol. 185:3190–3198. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Terness P, Bauer TM, Röse L, Dufter C,
Watzlik A, Simon H and Opelz G: Inhibition of allogeneic T cell
proliferation by indoleamine 2,3-dioxygenase-expressing dendritic
cells: Mediation of suppression by tryptophan metabolites. J Exp
Med. 196:447–457. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Frumento G, Rotondo R, Tonetti M, Damonte
G, Benatti U and Ferrara GB: Tryptophan-derived catabolites are
responsible for inhibition of T and natural killer cell
proliferation induced by indoleamine 2,3-dioxygenase. J Exp Med.
196:459–468. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fallarino F, Grohmann U, Vacca C, Bianchi
R, Orabona C, Spreca A, Fioretti M and Puccetti P: T cell apoptosis
by tryptophan catabolism. Cell Death Differ. 9:1069–1077. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fallarino F, Grohmann U, Vacca C, Orabona
C, Spreca A, Fioretti MC and Puccetti P: T cell apoptosis by
kynurenines. Adv Exp Med Biol. 527:183–190. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Seman M, Adriouch S, Scheuplein F, Krebs
C, Freese D, Glowacki G, Deterre P, Haag F and Koch-Nolte F:
NAD-induced T cell death: ADP-ribosylation of cell surface proteins
by ART2 activates the cytolytic P2X7 purinoceptor. Immunity.
19:571–582. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fallarino F, Grohmann U, You S, McGrath
BC, Cavener DR, Vacca C, Orabona C, Bianchi R, Belladonna ML, Volpi
C, et al: The combined effects of tryptophan starvation and
tryptophan catabolites down-regulate T cell receptor zeta-chain and
induce a regulatory phenotype in naive T cells. J Immunol.
176:6752–6761. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rubtsov YP and Rudensky AY: TGFbeta
signalling in control of T-cell-mediated self-reactivity. Nat Rev
Immunol. 7:443–453. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Belladonna ML, Volpi C, Bianchi R, Vacca
C, Orabona C, Pallotta MT, Boon L, Gizzi S, Fioretti MC, Grohmann U
and Puccetti P: Cutting edge: Autocrine TGF-beta sustains default
tolerogenesis by IDO-competent dendritic cells. J Immunol.
181:5194–5198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan W, Collado-Hidalgo A, Yufit T, Taylor
M and Varga J: Modulation of cellular tryptophan metabolism in
human fibroblasts by transforming growth factor-beta: Selective
inhibition of indoleamine 2,3-dioxygenase and tryptophanyl-tRNA
synthetase gene expression. J Cell Physiol. 177:174–186. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
MacKenzie CR, González RG, Kniep E, Roch S
and Däubener W: Cytokine mediated regulation of
interferon-gamma-induced IDO activation. Adv Exp Med Biol.
467:533–539. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thomas SR, Mohr D and Stocker R: Nitric
oxide inhibits indoleamine 2,3-dioxygenase activity in
interferon-gamma primed mononuclear phagocytes. J Biol Chem.
269:14457–14464. 1994.PubMed/NCBI
|
|
27
|
Martinez OM, Villanueva JC, Gershwin ME
and Krams SM: Cytokine patterns and cytotoxic mediators in primary
biliary cirrhosis. Hepatology. 21:113–119. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Neuman M, Angulo P, Malkiewicz I,
Jorgensen R, Shear N, Dickson ER, Haber J, Katz G and Lindor K:
Tumor necrosis factor-alpha and transforming growth factor-beta
reflect severity of liver damage in primary biliary cirrhosis. J
Gastroenterol Hepatol. 17:196–202. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Grubman SA, Perrone RD, Lee DW, Murray SL,
Rogers LC, Wolkoff LI, Mulberg AE, Cherington V and Jefferson DM:
Regulation of intracellular pH by immortalized human intrahepatic
biliary epithelial cell lines. Am J Physiol. 266:G1060–G1070.
1994.PubMed/NCBI
|
|
30
|
Brain JG, Robertson H, Thompson E,
Humphreys EH, Gardner A, Booth TA, Jones DE, Afford SC, von
Zglinicki T, Burt AD and Kirby JA: Biliary epithelial senescence
and plasticity in acute cellular rejection. Am J Transplant.
13:1688–1702. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chomczynski P and Sacchi N: Single-step
method of RNA isolation by acid guanidinium
thiocyanate-phenol-chloroform extraction. Anal Biochem.
162:156–159. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Takikawa O, Kuroiwa T, Yamazaki F and Kido
R: Mechanism of interferon-gamma action. Characterization of
indoleamine 2,3-dioxygenase in cultured human cells induced by
interferon-gamma and evaluation of the enzyme-mediated tryptophan
degradation in its anticellular activity. J Biol Chem.
263:2041–2048. 1988.PubMed/NCBI
|
|
34
|
Laich A, Neurauter G, Widner B and Fuchs
D: More rapid method for simultaneous measurement of tryptophan and
kynurenine by HPLC. Clin Chem. 48:579–581. 2002.PubMed/NCBI
|
|
35
|
Hwu P, Du MX, Lapointe R, Do M, Taylor MW
and Young HA: Indoleamine 2,3-dioxygenase production by human
dendritic cells results in the inhibition of T cell proliferation.
J Immunol. 164:3596–3599. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Billiau A and Matthys P: Interferon-gamma:
A historical perspective. Cytokine Growth Factor Rev. 20:97–113.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sakash JB, Byrne GI, Lichtman A and Libby
P: Cytokines induce indoleamine 2,3-dioxygenase expression in human
atheroma-associated cells: Implications for persistent
Chlamydophila pneumoniae infection. Infect Immun. 70:3959–3961.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Munn DH, Sharma MD, Lee JR, Jhaver KG,
Johnson TS, Keskin DB, Marshall B, Chandler P, Antonia SJ and
Burgess R: Potential regulatory function of human dendritic cells
expressing indoleamine 2,3-dioxygenase. Science. 297:1867–1870.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Knox WE and Mehler AH: The conversion of
tryptophan to kynurenine in liver I. The coupled tryptophan
peroxidase-oxidase system forming formylkynurenine. J Biol Chem.
187:419–430. 1950.PubMed/NCBI
|
|
40
|
Viñals F and Pouysségur J: Transforming
growth factor beta1 (TGF-beta1) promotes endothelial cell survival
during in vitro angiogenesis via an autocrine mechanism implicating
TGF-alpha signaling. Mol Cell Biol. 21:7218–7230. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shirey KA, Jung JY, Maeder GS and Carlin
JM: Upregulation of IFN-gamma receptor expression by
proinflammatory cytokines influences IDO activation in epithelial
cells. J Interferon Cytokine Res. 26:53–62. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zegarra-Moran O, Folli C, Manzari B,
Ravazzolo R, Varesio L and Galietta LJ: Double mechanism for apical
tryptophan depletion in polarized human bronchial epithelium. J
Immunol. 173:542–549. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Schröcksnadel K, Wirleitner B, Winkler C
and Fuchs D: Monitoring tryptophan metabolism in chronic immune
activation. Clin Chim Acta. 364:82–90. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Platten M, Ho PP, Youssef S, Fontoura P,
Garren H, Hur EM, Gupta R, Lee LY, Kidd BA, Robinson WH, et al:
Treatment of autoimmune neuroinflammation with a synthetic
tryptophan metabolite. Science. 310:850–855. 2005. View Article : Google Scholar : PubMed/NCBI
|