Introduction
Esophageal cancer, the sixth leading cause of
cancer-related death, has currently ranked the eighth in the
malignant tumors worldwide (1). There
were 455,800 new esophageal cancer cases and 400,200 deaths
estimated in 2012. The incidence rates of esophageal cancer vary
internationally, the highest rates are found in Eastern Asia and in
Eastern and Southern Africa and the lowest rates are found in
Western Africa. There are two main histological types of esophageal
cancer, including squamous cell carcinoma and adenocarcinoma
(2). Although, the diagnosis and
treatments for esophageal cancer update constantly (3,4), the
overall 5-year survival rate is only around 20% (5). Therefore, it is important and urgent to
elucidate an exactly novel molecular mechanism underlying
esophageal cancer formation, which may provide new strategies for
the diagnosis and treatments of esophageal cancer in future
healthcare.
TG-interacting factor (TGIF) belongs to the
three-amino acid loop extension (TALE) subfamily of atypical
homeodomain proteins (6).
Heterozygous loss of TGIF gene causes holoprosencephaly in humans
(7). It has been reported that TGIF
is involved in at least three signaling pathways, including
retinoic acid (RA) (6), transforming
growth factor β (TGF-β) (8), and
wnt/β-catenin signaling pathways (9).
A number of studies have reported that TGIF plays an important role
in the initiation, development, and progression of breast cancer
(9), lung cancer (10–12),
hepatocellular carcinoma (13,14), upper
urinary tract urothelial carcinoma (15,16),
leukemia (17–19), and oral squamous cell carcinoma
(20,21). Nakakuki et al reported that
frequent amplifications of TGIF were observed in esophageal
squamous cell carcinoma (ESCC) (22),
which suggests that TGIF might be associated with esophageal
tumorigenesis. But, the potential role of TGIF in the proliferation
and tumorigenicity of esophageal cancer cells is not clear. In the
present study, we knocked down TGIF of EC109 cells with short
hairpin RNA (shRNA) lentiviruses and observed the capabilities of
proliferation and tumorigenicity of stable TGIF-knocked down EC109
cells in vitro and in vivo. We also observed the
effects of TGIF knockdown on cisplatin-induced apoptosis in EC109
cells.
Materials and methods
Cell culture and infection
The esophageal cancer cell line of EC109 was
obtained from the Cell Resource Center, Peking Union Medical
College (which is the headquarters of National Infrastructure of
Cell Line Resource in Beijing, China) and cultured in RPMI-1640
medium with 10% of fetal bovine serum (FBS), 100 µg/ml of
streptomycin, 100 U/ml of penicillin, and 2 mM of L-glutamine at
37°C in a 5% CO2 incubator. TGIF shRNA (h) lentiviral
particles (sc-36659-V) and control shRNA lentiviral particles-A
(sc-108080) were obtained from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). The infection of lentiviral particles was
performed in accordance with the manufacturer's instructions. The
infected cells were maintained in RPMI-1640 full medium with 10
µg/ml of puromycin (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). The efficiency of the shRNA lentiviruses targeting TGIF
was measured at the level of protein expression. Stable clones were
named EC109-shRNA-TGIF and EC109-shRNA-control, respectively.
Measurement of cell proliferation
Cell proliferation was measured according to the
methods of our previous study (10).
Briefly, 4×104 of cells were seeded into each well of
12-well plate in triplicate. Cells were harvested and counted by
using a CASY Cell Counter (Schärfe System GmbH, Reutlingen,
Germany) at 24, 48, 72, and 96 h, respectively.
Colony formation assay
Colony formation assay in soft agar was performed as
previously described (11). Briefly,
500 cells were suspended in 0.35% low-melting point agarose and
plated onto 0.6% low-melting point agarose in 6-well plates. Cells
were maintained at 37°C in a 5% CO2 incubator for 18
days. Colonies of ten randomly selected views were counted by an
inverted microscope (Leica DM IL LED; Leica Microsystems GmbH,
Wetzlar, Germany) at ×100 magnification (11).
Tumor xenograft assay
All of the animal experiments were conducted in the
light of the Guide for the Care and Use of Laboratory Animals. The
Ethics Committee of Henan Center for Disease Control and Prevention
approved this study. Tumor formation assay in nude mice was carried
out in accordance with our previous study (11). In brief, male BALB/c nude mice aged 4
weeks were provided by Vital River Laboratory Animal Technology
Co., Ltd. (Beijing, China). Cells (5×106) in 150 µl of
PBS were subcutaneously injected into the back neck of each mouse.
Mice were monitored every day and sacrificed at 14 days
postinjection (11).
Cell cycle assay
The detailed methods of cell cycle assay were
described previously (10). Briefly,
cells were plated in 100-mm dishes and harvested when growing to
70–80% confluence. Cells were washed with PBS and fixed in 70% of
ethanol at −20°C overnight, following by washing with PBS twice,
suspending in 0.5 ml of PBS containing 100 µg/ml of RNase
(Invitrogen Life Technologies, Carlsbad, CA, USA) and 50 µg/ml of
propidium iodide (PI; Sigma-Aldrich, St. Louis, MO, USA), and
incubating at room temperature for 30 min in the dark. The
distribution of cell cycle was measured by EPICS XL-MCL ADC flow
cytometry (Beckman Coulter, Inc., Brea, CA, USA).
Cisplatin treatment
Cells were plated in 60-mm dishes and treated with
12.5 µg/ml of cisplatin for different time-points indicated. Cells
were harvested and washed with PBS, and then were used to analyze
protein expression and apoptosis.
Detection of apoptosis
The apoptosis was detected by using the Muse Annexin
V and Dead Cell Assay kit (MCH100105; EMD Millipore, Billerica, MA,
USA) in Muse™ Cell Analyzer according to the manufacturer's
instructions. In details, cells were detached by trypsinization and
suspended in at least 1% FBS. The cell samples were incubated with
Muse™ Annexin V and Dead Cell Reagent at room temperature for 20
min in the dark. The apoptosis rate was analyzed by using the Muse
Cell Analyzer.
Western blot assay
Western blot analysis was conducted as previously
described (10). The primary
antibodies used were listed as following: TGIF (sc-9084), Akt
(sc-8312), β-catenin (sc-7199), CDK4 (sc-260), cyclin A (sc-751),
cyclin B1 (sc-752), cyclin D1 (sc-718), p21 (sc-397), p53
(sc-6243), and β-actin (sc-47778) were obtained from Santa Cruz
Biotechnology, Inc., and Rb (#9313S), phospho-Rb (#8516S), c-Myc
(#13987S), p65 (#8242S), ERK1/2 (#4695S), Axin1 (#2087S), PARP
(#9532S), Bax (#5023S), caspase-3 (#9664S), and caspase-9 (#9508S)
were obtained from Cell Signaling Technology, Inc. (Danvers, MA,
USA). The secondary antibodies (peroxidase-coupled goat
anti-rabbit-IgG and goat anti-mouse IgG) were obtained from
ZSGB-BIO (Beijing, China). The membrane was developed by a Bio-Rad
Clarity™ Western ECL substrate in the ChemiDoc™ XRS+ Imaging system
(both from Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
Numeric data were presented as mean ± standard
deviation (SD) or median. Student's t-test, one-way analysis of
variance and Mann-Whitney U test were performed to estimate
statistical significance among groups by using the SPSS 13.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
statistically significant. All the tests were two-sided.
Results
The efficiency of TGIF knockdown in
EC109 cells
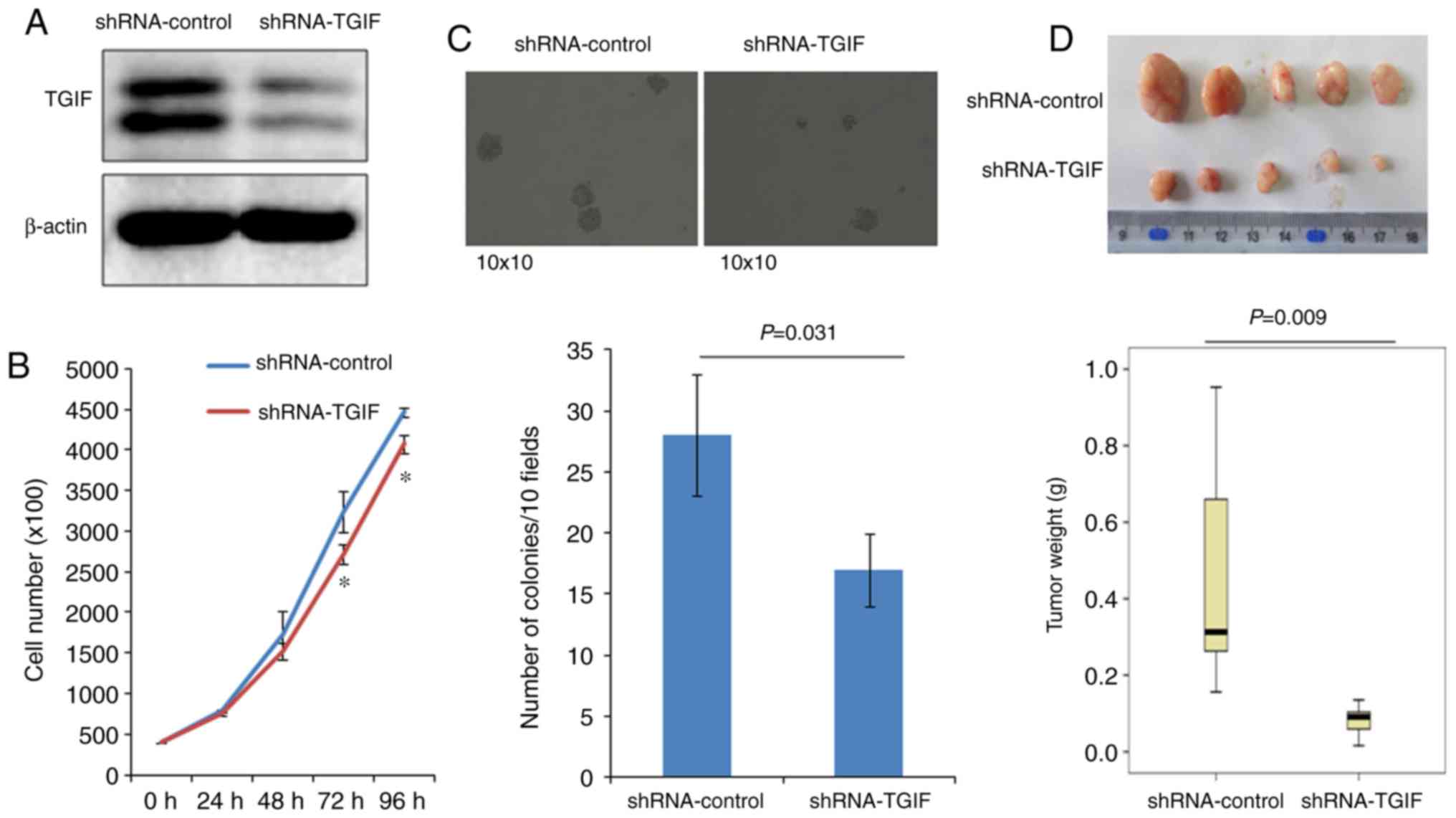
As shown in Fig. 1A,
the expression of TGIF protein was significantly reduced in
EC109-shRNA-TGIF cells, compared with EC109-shRNA-control cells,
which suggests that a stable TGIF-knocked down EC109 cell line was
successfully established.
Effects of TGIF knockdown on EC109
cell proliferation
Fig. 1B indicated that
EC109-shRNA-TGIF cells grew significantly slowly, compared with
EC109-shRNA-control cells from 72 h, which suggests that TGIF
knockdown suppressed EC109 cell proliferation.
Effects of TGIF knockdown on colony
formation
Results of colony formation were presented in
Fig. 1C, which showed that
EC109-shRNA-TGIF cells formed significantly less colonies (17±4/10
fields) than EC109-shRNA-control cells did (28±5/10 fields).
Effects of TGIF knockdown on tumor
xenograft formation in nude mice
Fig. 1D demonstrated
the results of tumor xenograft formation in nude mice. Our data
indicated that the significantly decreased tumor weight was
observed in EC109-shRNA-TGIF cells, compared with
EC109-shRNA-control cells (Fig. 1D),
which suggests that TGIF knockdown significantly suppressed tumor
formation and tumor growth of EC109 cells in vivo.
Effects of TGIF knockdown on cell
cycle distribution
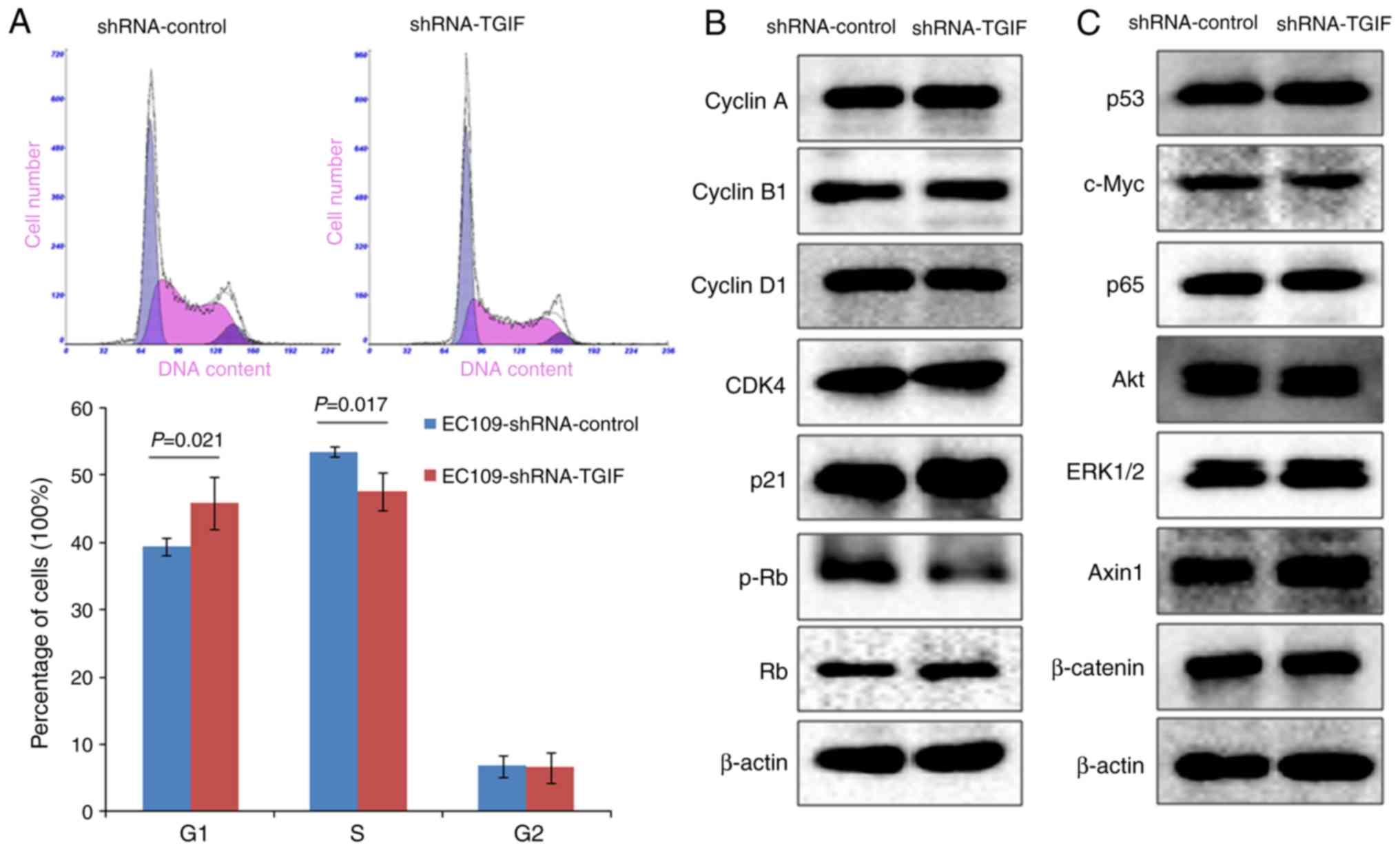
As shown in Fig. 2A,
EC109-shRNA-TGIF cells had the significantly increased percentage
of G1 phase cells (45.8±3.9%) accompanied with the significantly
decreased percentage of S phase cells (47.6±2.7%) as compared with
EC109-shRNA-control cells (39.5±1.3 and 53.5±0.7%, respectively).
Our data suggested that TGIF knockdown might induce the inhibition
of EC109 cell growth by arresting the cell cycle in the G1
phase.
TGIF knockdown suppressed the
expression of phospho-Rb
Fig. 2B presented the
alterations of cell cycle-related protein expression, while TGIF
knocking down in EC109 cells. Our findings showed that knockdown of
TGIF suppressed the expression of phospho-Rb protein. There was no
obvious alterations in the expression of the cyclin A, cyclin B1,
cyclin D1, CDK4 and p21 proteins between EC109-shRNA-TGIF cells and
EC109-shRNA-control cells. In addition, we did not observe
significant difference in the expression of p53, c-Myc, p65, Akt,
ERK1/2, Axin1 and β-catenin proteins between EC109-shRNA-TGIF cells
and EC109-shRNA-control cells (Fig.
2C).
Cisplatin suppressed the expression of
TGIF
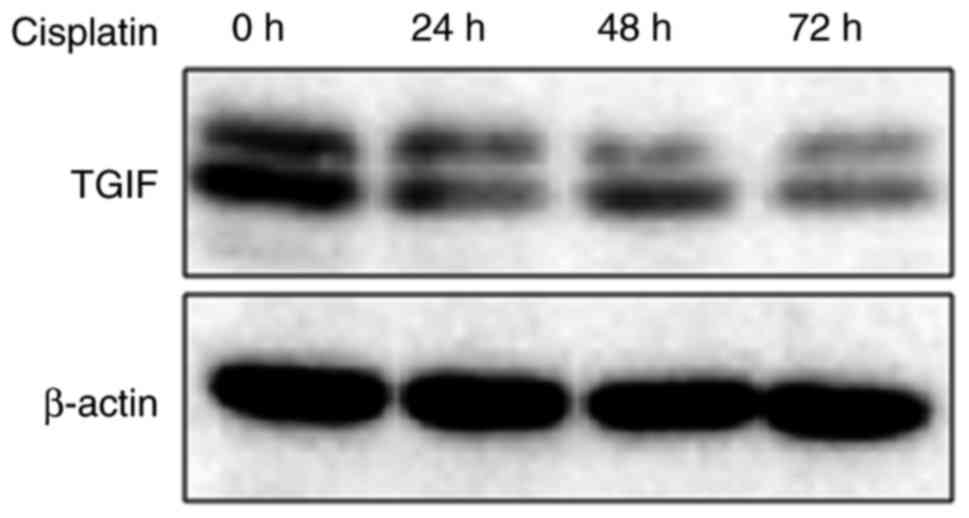
Fig. 3 exhibited the
effects of cisplatin on the expression of TGIF protein in EC109
cells. Our data showed that 12.5 µg/ml of cisplatin could suppress
the expression of TGIF protein from 24 h (Fig. 3).
TGIF knockdown promoted
cisplatin-indcued apoptosis
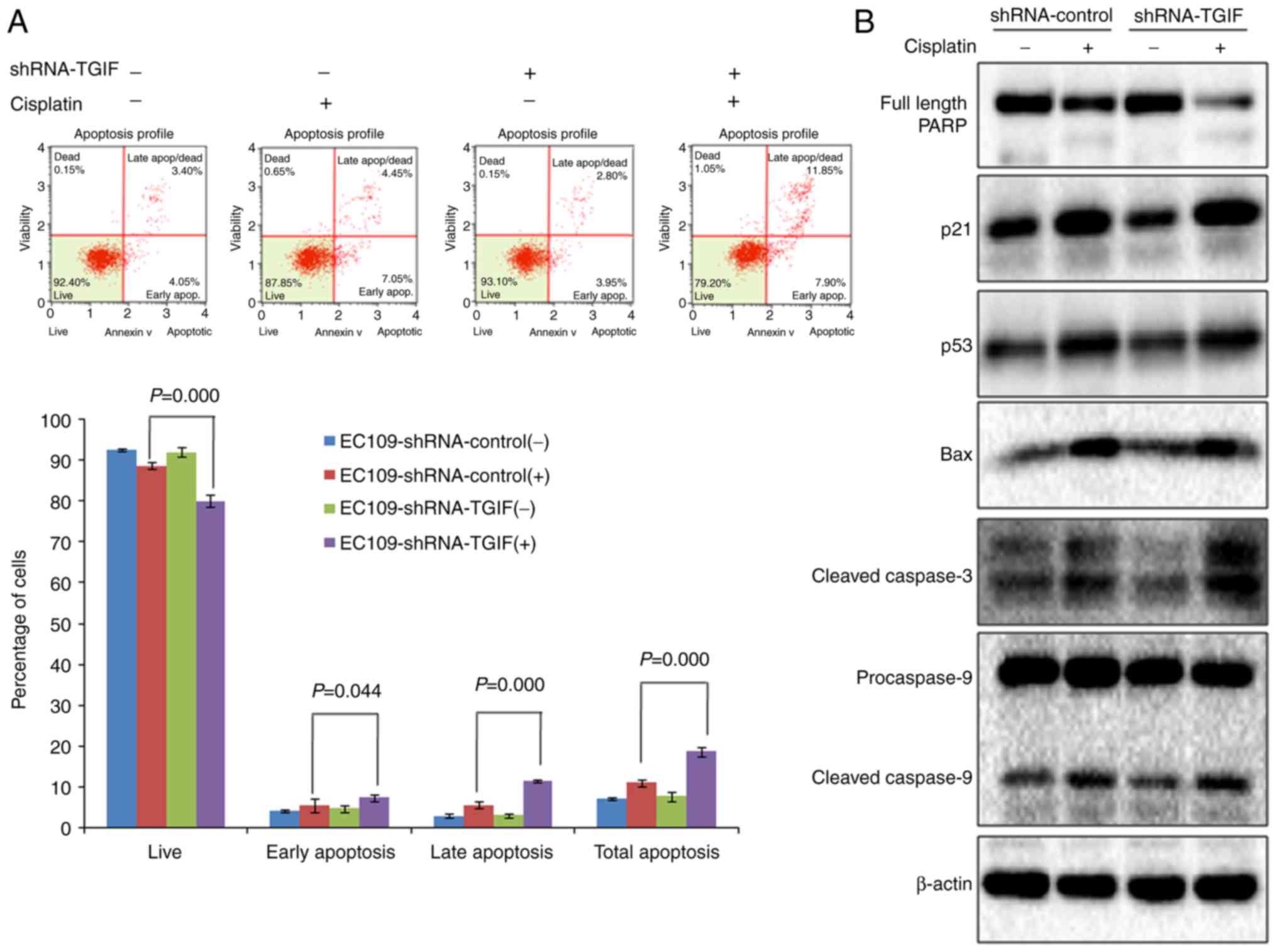
Results from flow cytometry indicated that the
percentage of total apoptosis (early apoptosis and late apoptosis)
was significantly higher in EC109-shRNA-control cells treated with
12.5 µg/ml of cisplatin than that in negative control (Fig. 4A), which suggests that cisplatin could
induce apoptosis in EC109 cells. When TGIF gene was knocked down by
shRNA in EC109 cells, we observed that the percentage of total
apoptosis was significantly higher in EC109-shRNA-TGIF cells
treated with 12.5 µg/ml of cisplatin than that in
EC109-shRNA-control cells treated with 12.5 µg/ml of cisplatin
(Fig. 4A), which suggests that TGIF
knockdown promoted cisplatin-induced apoptosis in EC109 cells.
In addition, we observed the significantly decreased
expression of full length PARP in EC109-shRNA-TGIF cells treated
with 12.5 µg/ml of cisplatin as compared with EC109-shRNA-control
cells treated with 12.5 µg/ml of cisplatin (Fig. 4B). We observed the significantly
increased expression of cleaved caspase-3 in EC109-shRNA-TGIF cells
treated with 12.5 µg/ml of cisplatin as compared with
EC109-shRNA-control cells treated with 12.5 µg/ml of cisplatin
(Fig. 4B). Our data suggested that
TGIF knockdown had effects on the expression of apoptosis-related
markers in EC109 cells treated with cisplatin.
Discussion
In our study, the functional role of TGIF in the
proliferation and tumorigenicity of esophageal cancer cell line of
EC109 was investigated. The abilities of EC109 cell growth and
tumor formation in vitro and in vivo were inhibited
when the expression of TGIF was knocked down by shRNA specifically
targeting TGIF, which suggests that TGIF may act as an oncogene in
the development of esophageal cancer. Knockdown of TGIF arrested
the cell cycle of EC109 cells in the G1 phase by downregulating
phospho-Rb. In addition, knockdown of TGIF promoted
cisplatin-induced apoptosis of EC109 cells.
Cell cycle arrest is one of the major causes of
cancer cell proliferation inhibition (23,24).
Dysregualtion of several key factors, including CDK4, cyclin D1,
p21 and phospho-Rb could result in G1 phase arrest (25,26). In
this study, we observed that knockdown of TGIF induced cell cycle
arrest in the G1 phase accompanied with significantly decreased
expression of phospho-Rb protein, while other proteins such as
CDK4, cyclin D1 and p21 did not significantly change. Studies have
shown that activation of cyclin D1-CDK4 complex can phosphorylate
Rb and keep Rb inactivation, thus promote G1/S phase transition
(27,28). Our previous data showed that silencing
of TGIF induced G1 phase cell cycle arrest along with the decreased
expression of phospho-Rb, cyclin D1 and CDK4 in lung cancer cells
(10). Together, the current
observations suggests that knockdown of TGIF led to the decreased
expression of phospho-Rb not through regulating CDK4 and cyclin D1
expression in esophageal cancer cells. Further studies should focus
on the mechanisms linking TGIF and phospho-Rb in esophageal
cancer.
Previous studies have shown that wnt/β-catenin
pathway is involved in the development of esophageal cancer
(29,30) and β-catenin is the key regulator in
the wnt/β-catenin signaling pathway. Deng et al reported
that aberrant expression of β-catenin was identified in 54.3% (114
of 265) of ESCC (31). The level of
β-catenin expression in ESCC was significantly higher than that in
the adjacent non-cancerous tissues (32,33). The
overexpression of β-catenin was aggressively associated with lymph
node metastasis, advanced pathological stage and prognosis of the
patients with ESCC (32). In
addition, Xu and Lu reported that β-catenin was involved in miR-214
inhibiting esophageal cancer cell growth and invasion (33). Jia et al found that RAP1B
activated wnt/β-catenin signaling in ESCC (34). However, in this present study, we
found that knockdown of TGIF had no obvious effects on the
expression of β-catenin and Axin1 proteins in esophageal cancer
cells, which suggests that the wnt/β-catenin signaling pathway
might not be involved in knockdown of TGIF inhibiting the
tumorigenicity of esophageal cancer cells. Previous studies showed
that TGIF could regulate the expression of β-catenin protein in
breast cancer (9) and lung cancer
(10,12). Taken together, the regulation of
β-catenin by TGIF might be dependent on tumor types.
In this study, we observed that knockdown of TGIF
suppressed the tumorigenicity of esophageal cancer cell of EC109
and cisplatin could repress the expression of TGIF protein. We
further investigated the potential role of TGIF in
cisplatin-induced apoptosis of EC109 cells. Our data showed that
knockdown of TGIF promoted cisplatin-induced apoptosis of EC109
cells, along with the alterations of apoptosis-related markers,
such as the decreased level of full length PARP protein expression
and the increased level of cleaved caspase-3 protein expression.
Studies suggested the cleavage of caspase-3 was an early event in
apoptosis induced by chemotherapeutic agents (35). Activation of caspase-3 was partially
or totally responsible for proteolytic cleavage of many key
proteins such as PARP (36,37). Liu et al reported that
knockdown of TGIF enhanced arsenic trioxide-induced apoptosis in
HepG2 cells (38). Together, our
findings indicated that TGIF was likely to be a potential
therapeutic target for the treatment of esophageal cancer.
To the best of our knowledge, only one published
study reported the association of TGIF amplifications with
esophageal cancer (22). Although, in
this study, we primarily obtained exciting data on the potential
role of TGIF in the proliferation and tumorigenicity of esophageal
cancer cells, some limitations should be acknowledged. First, our
data are only based on one esophageal cancer cell line, other cell
lines should be applied to verify our findings. Second, the pattern
of TGIF expression in esophageal cancer tissue should be
investigated in future studies. Third, the effects of TGIF
overexpression on the proliferation and tumorigenicity in
esophageal cancer cells should be addressed further. In addition,
transgenic animal model could be applied to assess the functional
role of TGIF in esophageal tumorigenesis.
In conclusion, the present study indicates that
knockdown of TGIF induces growth inhibition of EC109 cells via
arresting cell cycle in the G1 phase by downregulating phospho-Rb.
This study also indicates that TGIF plays an important role in
modulating the tumorigenicity of EC109 cells and cisplatin-induced
apoptosis. Therefore, this study enriches our understanding of the
oncogenesis of esophageal cancer and suggests that TGIF is likely
to be a new therapeutic target for esophageal cancer treatment.
Acknowledgements
This study was supported by a grant from National
Natural Science Foundation of China (no. U1404815). The funder had
no role in study design, data collection and analysis, decision to
publish, or preparation of the manuscript.
References
|
1
|
Xie SH, Wahlin K and Lagergren J: Cause of
death in patients diagnosed with esophageal cancer in Sweden: A
population-based study. Oncotarget. 8:51800–51809. 2017.PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang M, Hao C, Ma Q, Song G, Ma S, Zhao D,
Zhao L, Li X and Wei W: DNA image cytometry test for primary
screening of esophageal cancer: A population-based multi-center
study in high-risk areas in China. Chin J Cancer Res. 28:404–412.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sohda M and Kuwano H: Current status and
future prospects for esophageal cancer treatment. Ann Thorac
Cardiovasc Surg. 23:1–11. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Song G, Liu K, Yang X, Mu B, Yang J, He L,
Hu X, Li Q, Zhao Y, Cai X and Feng G: SATB1 plays an oncogenic role
in esophageal cancer by up-regulation of FN1 and PDGFRB.
Oncotarget. 8:17771–17784. 2017.PubMed/NCBI
|
|
6
|
Bertolino E, Reimund B, Wildt-Perinic D
and Clerc RG: A novel homeobox protein which recognizes a TGT core
and functionally interferes with a retinoid-responsive motif. J
Biol Chem. 270:31178–31188. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gripp KW, Wotton D, Edwards MC, Roessler
E, Ades L, Meinecke P, Richieri-Costa A, Zackai EH, Massagué J,
Muenke M and Elledge SJ: Mutations in TGIF cause holoprosencephaly
and link NODAL signalling to human neural axis determination. Nat
Genet. 25:205–208. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wotton D, Lo RS, Lee S and Massagué J: A
Smad transcriptional corepressor. Cell. 97:29–39. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang MZ, Ferrigno O, Wang Z, Ohnishi M,
Prunier C, Levy L, Razzaque M, Horne WC, Romero D, Tzivion G, et
al: TGIF governs a feed-forward network that empowers Wnt signaling
to drive mammary tumorigenesis. Cancer Cell. 27:547–560. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Pan T, Wang H, Li L, Li J, Zhang C
and Yang H: Silencing of TGIF attenuates the tumorigenicity of A549
cells in vitro and in vivo. Tumour Biol. 37:12725–12730. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Wang H, Gao H, Xu B, Zhai W, Li J
and Zhang C: Elevated expression of TGIF is involved in lung
carcinogenesis. Tumour Biol. 36:9223–9231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiang G, Yi Y, Weiwei H and Weiming W:
TGIF1 promoted the growth and migration of cancer cells in nonsmall
cell lung cancer. Tumour Biol. 36:9303–9310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Borlak J, Meier T, Halter R, Spanel R and
Spanel-Borowski K: Epidermal growth factor-induced hepatocellular
carcinoma: Gene expression profiles in precursor lesions, early
stage and solitary tumours. Oncogene. 24:1809–1819. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu ZM, Tseng HY, Tsai HW, Su FC and Huang
HS: Transforming growth factor β-interacting factor-induced
malignant progression of hepatocellular carcinoma cells depends on
superoxide production from Nox4. Free Radic Biol Med. 84:54–64.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang HS, Liu ZM, Chen PC, Tseng HY and
Yeh BW: TG-interacting factor-induced superoxide production from
NADPH oxidase contributes to the migration/invasion of urothelial
carcinoma. Free Radic Biol Med. 53:769–778. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yeh BW, Wu WJ, Li WM, Li CC, Huang CN,
Kang WY, Liu ZM and Huang HS: Overexpression of TG-interacting
factor is associated with worse prognosis in upper urinary tract
urothelial carcinoma. Am J Pathol. 181:1044–1055. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hamid R and Brandt SJ: Transforming
growth-interacting factor (TGIF) regulates proliferation and
differentiation of human myeloid leukemia cells. Mol Oncol.
3:451–463. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hamid R, Patterson J and Brandt SJ:
Genomic structure, alternative splicing and expression of
TG-interacting factor, in human myeloid leukemia blasts and cell
lines. Biochim Biophys Acta. 1779:347–355. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Willer A, Jakobsen JS, Ohlsson E, Rapin N,
Waage J, Billing M, Bullinger L, Karlsson S and Porse BT: TGIF1 is
a negative regulator of MLL-rearranged acute myeloid leukemia.
Leukemia. 29:1018–1031. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Libório TN, Ferreira EN, Xavier Aquino FC,
Carraro DM, Kowalski LP, Soares FA and Nunes FD: TGIF1 splicing
variant 8 is overexpressed in oral squamous cell carcinoma and is
related to pathologic and clinical behavior. Oral Surg Oral Med
Oral Pathol Oral Radiol. 116:614–625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Matizonkas-Antonio LF, Libório TN, Xavier
Aquino FC, Silva-Valenzuela Md, Michaluarte-Júnior P and Nunes FD:
Detection of TGIF1 homeobox gene in oral squamous cell carcinoma
according to histologic grading. Oral Surg Oral Med Oral Pathol
Oral Radiol Endod. 111:218–224. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nakakuki K, Imoto I, Pimkhaokham A, Fukuda
Y, Shimada Y, Imamura M, Amagasa T and Inazawa J: Novel targets for
the 18p11.3 amplification frequently observed in esophageal
squamous cell carcinomas. Carcinogenesis. 23:19–24. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chan AS, Mowla SN, Arora P and Jat PS:
Tumour suppressors and cellular senescence. IUBMB Life. 66:812–822.
2012. View
Article : Google Scholar
|
|
24
|
López-Sáez JF, de la Torre C, Pincheira J
and Giménez-Martin G: Cell proliferation and cancer. Histol
Histopathol. 13:1197–1214. 1998.PubMed/NCBI
|
|
25
|
Reed SI, Bailly E, Dulic V, Hengst L,
Resnitzky D and Slingerland J: G1 control in mammalian cells. J
Cell Sci Suppl. 18:69–73. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Reed SI: Control of the G1/S transition.
Cancer Surv. 29:7–23. 1997.PubMed/NCBI
|
|
27
|
Sherr CJ: Mammalian G1 cyclins. Cell.
73:1059–1065. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Weinberg RA: The retinoblastoma protein
and cell cycle control. Cell. 81:323–330. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang M, Linghu E, Zhan Q, He T, Cao B,
Brock MV, Herman JG, Xiang R and Guo M: Methylation of DACT2
accelerates esophageal cancer development by activating Wnt
signaling. Oncotarget. 7:17957–17969. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ge C, Wu S, Wang W, Liu Z, Zhang J, Wang
Z, Li R, Zhang Z, Li Z, Dong S, et al: miR-942 promotes cancer stem
cell-like traits in esophageal squamous cell carcinoma through
activation of Wnt/β-catenin signalling pathway. Oncotarget.
6:10964–10977. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Deng F, Zhou K, Cui W, Liu D and Ma Y:
Clinicopathological significance of wnt/β-catenin signaling pathway
in esophageal squamous cell carcinoma. Int J Clin Exp Pathol.
8:3045–3053. 2015.PubMed/NCBI
|
|
32
|
Lv J, Cao XF, Ji L, Zhu B, Wang DD, Tao L
and Li SQ: Association of β-catenin, Wnt1, Smad4, Hoxa9, and Bmi-1
with the prognosis of esophageal squamous cell carcinoma. Med
Oncol. 29:151–160. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Y and Lu S: Regulation of
β-catenin-mediated esophageal cancer growth and invasion by
miR-214. Am J Transl Res. 7:2316–2325. 2015.PubMed/NCBI
|
|
34
|
Jia Z, Yang Y, Dengyan Z, Chunyang Z,
Donglei L, Kai W and Song Z: RAP1B, a DVL2 binding protein,
activates Wnt/beta-catenin signaling in esophageal squamous cell
carcinoma. Gene. 611:15–20. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Henkels KM and Turchi JJ:
Cisplatin-induced apoptosis proceeds by caspase-3-dependent and
-independent pathways in cisplatin-resistant and -sensitive human
ovarian cancer cell lines. Cancer Res. 59:3077–3083.
1999.PubMed/NCBI
|
|
36
|
Horky M, Wurzer G, Kotala V, Anton M,
Vojtĕsek B, Vácha J and Wesierska-Gadek J: Segregation of nucleolar
components coincides with caspase-3 activation in cisplatin-treated
HeLa cells. J Cell Sci. 114:663–670. 2001.PubMed/NCBI
|
|
37
|
Rheaume E, Cohen LY, Uhlmann F, Lazure C,
Alam A, Hurwitz J, Sekaly RP and Denis F: The large subunit of
replication factor C is a substrate for caspase-3 in vitro and is
cleaved by a caspase-3-like protease during Fas-mediated apoptosis.
EMBO J. 16:6346–6354. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu ZM, Tseng JT, Hong DY and Huang HS:
Suppression of TG-interacting factor sensitizes arsenic
trioxide-induced apoptosis in human hepatocellular carcinoma cells.
Biochem J. 438:349–358. 2011. View Article : Google Scholar : PubMed/NCBI
|