Introduction
Renal cell carcinoma (RCC) is the most common
malignant cancer of the kidney in adults, accounting for almost 4%
of all tumors (1). It is estimated
that there are ~30,000 new cases and 12,000 mortalities due to RCC
every year in the USA (2). RCC may be
histologically classified into three subtypes, including clear cell
RCC, papillary RCC and chromophobe RCC. Clear cell RCC is the most
common of these subtypes, accounting for 75 to 80% of RCC cases,
and also is the most aggressive (3–5). The
standard curative treatment for patients with localized RCC is
surgical excision with total nephrectomy. However, in ~30% of cases
the tumors have already metastasized at the time of initial
presentation (6,7). In addition, the benefit of
chemotherapeutic and radiological approaches for RCC is limited,
which results in a poor prognosis and a low five-year survival rate
(8). It is therefore an urgent
requirement to investigate novel molecular biomarkers which may be
used for early detection and targeted therapy, based on the
developing knowledge of the tumorigenesis and progression
mechanisms of RCC.
MicroRNAs (miRNAs/miRs), small non-coding RNA
molecules of ~19–25 nucleotides (nt) in length, exist in a number
of organisms and are cleaved from 70 to 100-nt-long hairpin
pre-miRNA precursors by the enzyme Drosha (9,10). miRNAs
suppress gene expression at the post-transcriptional level through
imperfect complementary sequence pairing to the 3′ untranslated
regions (3′UTRs) of their target genes and inducing mRNA
degradation or translational repression (11,12). It is
estimated that >30% of human genes are regulated by miRNAs, and
that they are involved in a variety of biological functions,
including proliferation, differentiation, cell cycle, apoptosis,
angiogenesis, invasion and metastasis (13,14). An
accumulating number of studies have reported that miRNA expression
levels are dysregulated in a variety of human cancer types
(15–17). miRNAs play key functions in the
initiation and progression of tumors by serving as oncogenes or
tumor suppressors, primarily depending on the roles of their target
genes (18–20). Therefore, miRNAs may be therapeutic
targets for novel treatment strategies against RCC.
In this study, miR-138 expression was observed to be
downregulated in RCC cell lines and tissues, and the expression of
miR-138 was correlated with histological grade, tumor stage and
lymph node metastasis. miR-138 was identified as a tumor suppressor
in RCC, by inhibiting RCC cell proliferation and invasion.
Furthermore, SOX9 was demonstrated to be a direct and functional
target gene of miR-138 in RCC. These findings from the present
study indicate that miR-138 may be a potential therapeutic target
for RCC.
Materials and methods
Tissue samples and cell lines
Sixty-two pairs of RCC tissues and matched normal
adjacent tissues (NATs) were collected from patients with RCC who
had undergone surgery at Shandong Provincial Hospital Affiliated to
Shandong University, China. The matched NATs were kidney tissues
located 2 cm away from the visible tumor lesions. All tissue
samples were frozen in liquid nitrogen immediately after resection
and stored at −80°C until RNA extraction. This study was approved
by the ethics committee of Shandong Provincial Hospital Affiliated
to Shandong University.
The human RCC cell lines 786-O, ACHN, A498, OS-RC-2
and Caki-1 were purchased from the American Type Culture Collection
(ATCC; Manassas, VA, USA), and cultured in Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin (Gibco; Thermo
Fisher Scientific, Inc.) and 100 mg/l streptomycin (Gibco; Thermo
Fisher Scientific, Inc.). The non-malignant SV-40 immortalized
renal cell line HK-2 was obtained from Shanghai Institute of
Biochemistry and Cell Biology (Shanghai, China), and maintained in
keratinocyte serum-free medium (Invitrogen; Thermo Fisher
Scientific, Inc.). All cells were maintained at 37°C in a
humidified incubator (5% CO2).
Cell transfection
The following oligonucleotides were used in the
present study: miR-138 mimics, scrambled control (NC), SOX9 siRNA
(si-SOX9) and control siRNA (si-NC). Cells were transfected with
these oligonucleotides using Lipofectamine® 2000
transfection reagent (Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
Total RNA was extracted from cell lines or tissue
samples using TRIzol (Invitrogen), following to the manufacturer's
protocol. For miR-138 expression, RT-qPCR was performed in a CFX96
Real-Time PCR Detection system (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) using a One Step SYBR® PrimeScript™
miRNA RT-PCR kit (Takara Bio, Inc., Otsu, Japan). For analysis of
SOX9 mRNA, cDNA was synthesized using the Moloney Murine Leukemia
Virus Reverse Transcription system (Promega Corporation, Madison,
WI, USA), and qPCR was performed with a SYBR-Green I mix (Takara
Bio), according to the manufacturer's protocol. The relative
expression of miR-138 and SOX9 mRNA was determined using the
2−∆∆Ct analysis method, where U6 was used as an internal
control for miR-138 and GADPH for SOX9 mRNA.
MTT assay
The capacity for cellular proliferation was
determined with a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Sigma-Aldrich; EMD Millipore, Billerica, MA, USA) assay. Cells were
plated at 3,000 cells per well in 96-well culture plates. Following
transfection for 24, 48, 72 and 96 h, 20 µl MTT solution (5 mg/ml)
was added to the wells and incubated for an additional 4 h. The
supernatant was then removed and 200 µl dimethylsulphoxide was
added to each well to dissolve the precipitate. Finally, absorbance
was measured at a wavelength of 490 nm using a microplate reader
(model 680; Bio-Rad Laboratories, Inc.).
Transwell invasion assay
The capacity for cellular invasion was evaluated
using Transwell chambers (8.0 mm pore size; BD Biosciences, San
Jose, CA, USA). The chambers were coated with Matrigel (BD
Biosciences) and incubated for another 5 h. Transfected cells were
collected 48 h after transfection, and re-suspended in FBS-free
DMEM. A total of 1×104 cells were plated into the upper
chamber, while culture medium containing 20% FBS in the lower
chamber served as a chemoattractant. Following incubation for 48 h,
the cells that did not invade through the pores were carefully
removed with a cotton swab. The invaded cells were fixed with 95%
ethanol, stained with 0.1% crystal violet and washed with
phosphate-buffered saline (Gibco; Thermo Fisher Scientific, Inc.).
The number of invaded cells was counted in five randomly selected
fields using an Eclipse TS 100 microscope (Nikon Corporation,
Tokyo, Japan).
Western blot analysis
At 72 h after transfection, total protein was
extracted using radioimmunoprecipitation assay lysis buffer (50 mM
Tris Cl, pH 7.4, 150 mM NaCl, 5 mM EDTA, 1% Nonidet P-40, 1% sodium
deoxycholate, 0.1% SDS/1% aprotinin, 50 mM NaF and 0.1 mM
Na3VO4) supplemented with a protease
inhibitor cocktail (Promega Corporation). After denaturing at 100°C
for 5 min, equal amounts of proteins were separated using 10%
SDS-PAGE, and then transferred onto polyvinylidene difluoride
membranes (EMD Millipore). Subsequently, the membranes were blocked
with Tris-buffered saline containing 5% skimmed milk, and probed
with primary antibodies, including mouse anti-human monoclonal SOX9
antibody (1:1,000 dilution; sc-166505; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA) and mouse anti-human monoclonal β-actin
antibody (1:1,000 dilution; sc-130301; Santa Cruz Biotechnology,
Inc.), at 4°C for 12 h. Finally, protein signals were detected
using goat anti-mouse horseradish peroxidase-conjugated secondary
antibody (Santa Cruz Biotechnology, Inc.). Bands were visualized
using enhanced chemiluminescence solution (Pierce; Thermo Fisher
Scientific, Inc.). The relative expression of SOX9 was determined
after normalizing to β-actin.
Luciferase reporter assay
For the luciferase reporter assay, PGL3-SOX9-3′UTR
wild-type (Wt) and PGL3-SOX9-3′UTR mutant (Mut) luciferase reporter
vectors were synthesized and purified by GenePharma (Shanghai,
China). In brief, cells were seeded into 24-well plates at a
confluence of 60–70%. Following incubation overnight, cells were
transfected with miR-138 mimics or NC together with luciferase
reporter vectors using Lipofectamine® 2000 transfection
reagent. At 48 h after transfection, luciferase activity was
quantified using the Dual-Luciferase Reporter Assay system (Promega
Corporation). The results were normalized to Renilla
luciferase activity.
Statistical analysis
Data are expressed as the means ± standard deviation
from at least three separate experiments, and compared using SPSS
version 18.0 software (SPSS, Inc., Chicago, IL, USA). Values of
P<0.05 were considered to indicate a statistically significant
difference.
Results
miR-138 is downregulated in RCC and is
associated with clinicopathological features
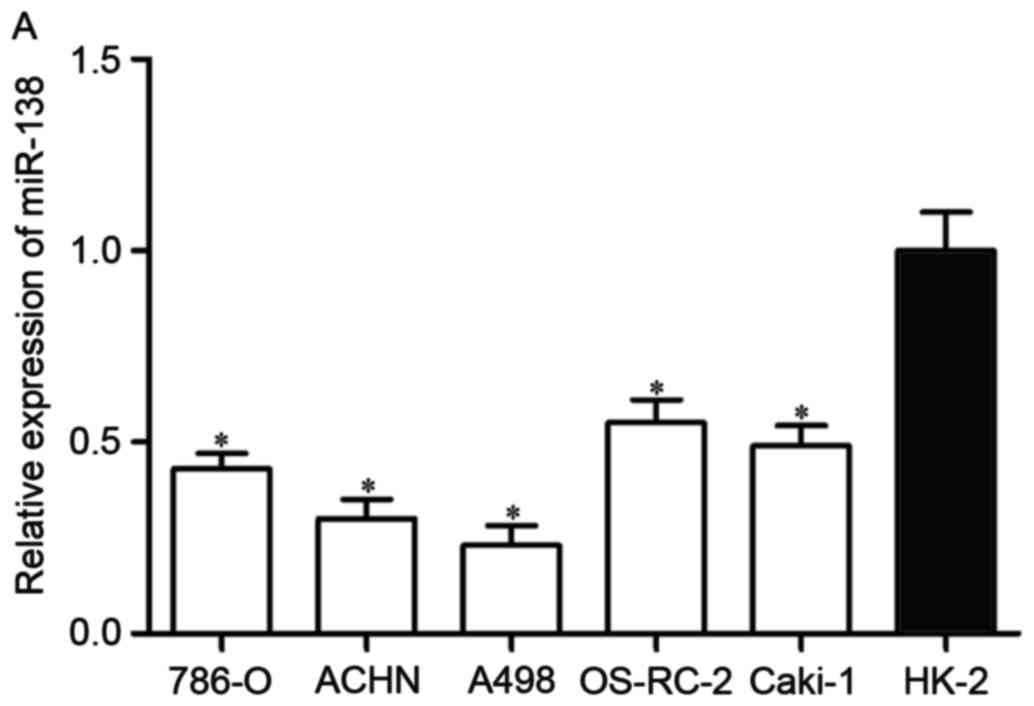
First, miR-138 expression was measured in five RCC
cell lines (786-O, ACHN, A498, OS-RC-2 and Caki-1) and the
non-malignant SV-40 immortalized renal cell line (HK-2). Expression
levels of miR-138 were observed to be significantly lower in RCC
cell lines than in HK-2 (Fig. 1A,
P<0.05). miR-138 expression was also evaluated in RCC tissues
and matched NATs. As shown in Fig.
1B, miR-138 was downregulated in RCC tissues compared with
matched NATs (P<0.05).
Next, the association between miR-138 expression
levels and clinicopathological factors of RCC patients was
assessed. As shown in Table I, low
expression levels of miR-138 were significantly correlated with
histological grade, tumor stage and lymph node metastasis
(P<0.05). However, there were no notable differences between
miR-138 expression and other clinicopathological features in this
study (P>0.05). These findings suggest that miR-138 was
downregulated in RCC, and that low expression levels of miR-138 may
serve a significant role in RCC carcinogenesis and progression.
 | Table I.Correlation between miR-138 expression
and clinicopathological features. |
Table I.
Correlation between miR-138 expression
and clinicopathological features.
|
|
| miR-138
expression |
|
|---|
|
|
|
|
|
|---|
| Parameter | Cases | Low | High | P-value |
|---|
| Gender |
|
|
| 0.933 |
| Male | 28 | 17 | 11 |
|
|
Female | 34 | 21 | 13 |
|
| Age |
|
|
| 0.275 |
| <60
years | 36 | 20 | 16 |
|
| ≥60
years | 26 | 18 | 8 |
|
| Tumor size |
|
|
| 0.213 |
| <4
cm | 30 | 16 | 14 |
|
| ≥4
cm | 32 | 22 | 10 |
|
| Histological
grade |
|
|
| 0.001 |
|
I–II | 32 | 13 | 19 |
|
|
III–IV | 30 | 25 | 5 |
|
| Tumor stage |
|
|
| 0.009 |
|
T1-T2 | 26 | 11 | 15 |
|
|
T3-T4 | 36 | 27 | 9 |
|
| Lymph node
metastasis |
|
|
| 0.016 |
|
Negative | 32 | 15 | 17 |
|
|
Positive | 30 | 23 | 7 |
|
miR-138 inhibits proliferation and
invasion of RCC cells
The significantly low expression of miR-138 in RCC
cell lines and tissues prompted the investigation of the
contribution of miR-138 in RCC carcinogenesis and progression. To
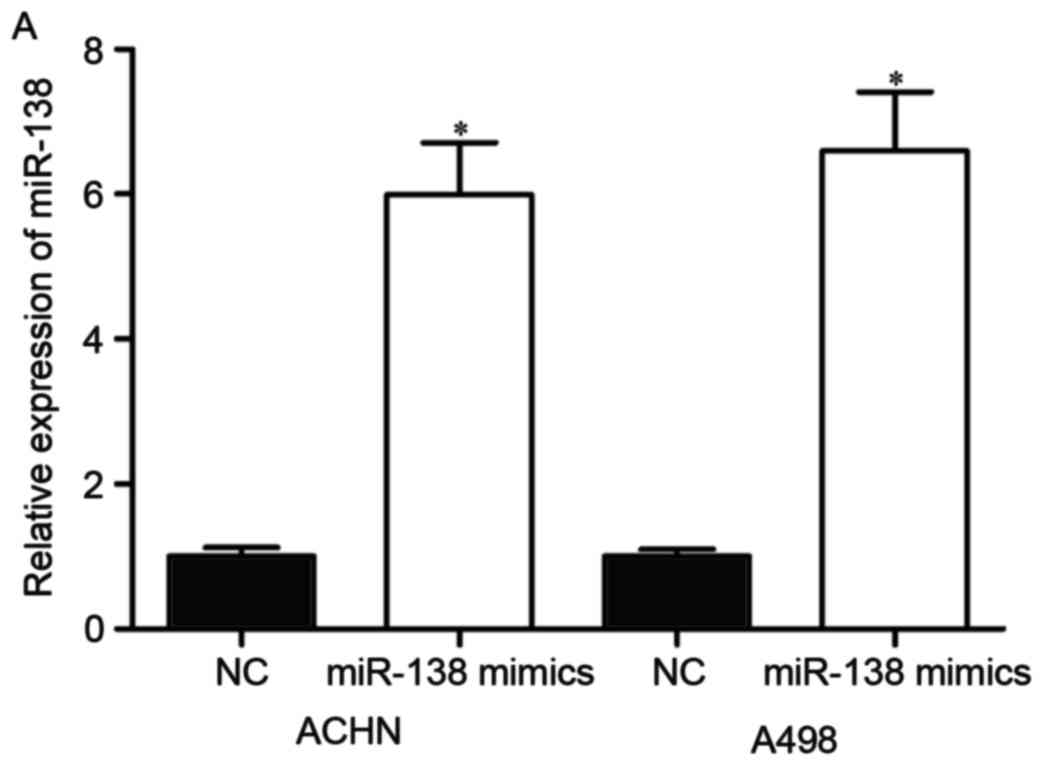
determine this, its expression was increased in RCC cells. miR-138
mimics or NC were transfected into ACHN and A498 cells.
Upregulation of miR-138 in ACHN and A498 cells was confirmed by
RT-qPCR (Fig. 2A, P<0.05).
The effect of miR-138 on RCC cell proliferation was
evaluated by MTT assay. As shown in Fig.
2B, ectopic miR-138 expression significantly inhibited the
proliferation of ACHN and A498 cells compared with NC (Fig. 2B, P<0.05).
Furthermore, the effect of miR-138 on RCC cell
invasion was assessed by Transwell invasion assay. The results
revealed that overexpression of miR-138 decreased RCC cell invasion
ability (Fig. 2C, P<0.05). This
indicated that miR-138 acted as a tumor suppressor in RCC by
inhibiting cell proliferation and invasion.
SOX9 is a target gene of miR-138
To elucidate the underlying molecular mechanisms by
which miR-138 exerts its suppressive functions, the direct target
genes of miR-138 were explored. Using TargetScan (http://www.targetscan.org/vert_60/), SOX9 was
observed to be a potential target of miR-138 based on putative
target sequences at position 1082–1089 of the SOX9 3′UTR (Fig. 3A).
To investigate whether SOX9 was a direct target of
miR-138, luciferase reporter assay was performed. PGL3-SOX9-3′UTR
Wt or PGL3-SOX9-3′UTR Mut luciferase reporter vectors, together
with miR-138 mimics or NC, were co-transfected into ACHN and A498
cells. The results revealed that luciferase activity was notably
suppressed by miR-138 mimics in PGL3-SOX9-3′UTR Wt (Fig. 3B, P<0.05), whereas the inhibitory
effect of miR-138 mimics was abolished in PGL3-SOX9-3′UTR Mut.
To examine the inhibitory effect of miR-138 on SOX9
expression, RT-qPCR and western blot analysis were performed in
ACHN and A498 cells transfected with miR-138 mimics or NC. The
results revealed that the mRNA (Fig.
3C, P<0.05) and protein (Fig.
3D, P<0.05) levels of SOX9 in ACHN and A498 cells
transfected with miR-138 mimics were significantly decreased
compared with those in the NC groups. Taken together, these results
strongly indicated that SOX9 was a direct target of miR-138 in
RCC.
SOX9 knockdown inhibits RCC cell
proliferation and invasion
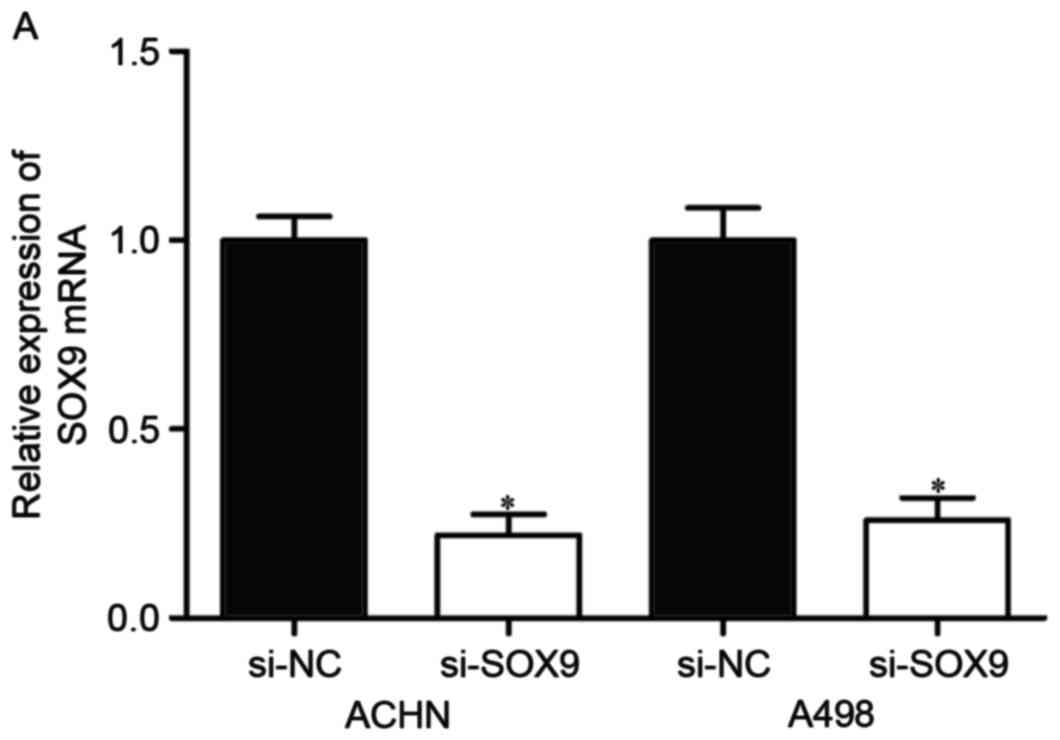
To explore whether SOX9 affected RCC cell
proliferation and invasion, si-SOX9 or si-NC was introduced into
ACHN and A498 cells. Following transfection, the expression of SOX9
mRNA was significantly downregulated in ACHN and A498 cells
transfected with si-SOX9 (Fig. 4A,
P<0.05).
To determine the effect of SOX9 knockdown on RCC
cell proliferation, MTT assay was performed in ACHN and A498 cells
transfected with si-SOX9 or si-NC. As shown in Fig. 4B, cell proliferation was significantly
suppressed by si-SOX9 in ACHN and A498 cells (P<0.05).
Transwell invasion assay was adopted to evaluate the
effect of SOX9 knockdown on RCC cell invasion. As shown in Fig. 4C, the number of invading cells was
significantly decreased in ACHN and A498 cells transfected with
si-SOX9 compared with the si-NC groups. Collectively, knockdown of
SOX9 notably inhibited RCC cell proliferation and invasion, with a
similar effect to that induced by miR-138, rendering SOX9 a
functional target of miR-138 in RCC.
Discussion
In the present study, it was revealed that miR-138
was significantly downregulated in RCC cell lines and tissue
samples compared with the non-malignant SV-40 immortalized renal
cell line and matched NATs, respectively. Decreased expression
levels of miR-138 were notably correlated with histological grade,
tumor stage and lymph node metastasis in RCC patients. In addition,
enforced miR-138 expression reduced cell proliferation and invasion
of RCC. Next, SOX9 was validated as a direct target gene of miR-138
in RCC. The effects of SOX9 knockdown on RCC cell proliferation and
invasion were similar to those induced by miR-138 overexpression,
rendering SOX9 a functional target of miR-138 in RCC. From these
data, it was concluded that miR-138 may serve as a potential
treatment target to inhibit the growth and metastasis of RCC.
Downregulation of miR-138 has been identified in
several cancer types, including breast cancer (21), bladder cancer (22), larynx carcinoma (23), colorectal cancer (24,25),
non-small cell lung cancer (26,27), oral
squamous cell carcinoma (28),
gallbladder carcinoma (29),
pancreatic cancer (30),
hepatocellular carcinoma (31) and
glioblastoma (32). More
significantly, miR-138 expression levels were observed to be
correlated with clinicopathological features of cancer patients.
For example, in breast cancer, miR-138 expression was correlated
with lymph node metastasis and invasion (21). In non-small cell lung cancer, low
expression levels of miR-138 were significantly associated with
advanced tumor-node-metastasis (TNM) stage and positive lymph node
metastasis. In addition, non-small cell lung cancer patients with
low miR-138 expression levels had a shorter overall survival time
than those with high miR-138 expression levels (27). In hepatocellular carcinoma, miR-138
downregulation was significantly correlated with advanced TNM stage
and the presence of portal vein invasion and lymph node metastasis.
In addition, multivariate survival analysis indicated that low
expression of miR-138 was an independent prognostic factor for
patients with hepatocellular carcinoma (31). In glioblastoma, high expression levels
of miR-138 were associated with a long overall and progression-free
survival time (32). These findings
suggested that miR-138 may represent a promising prognostic
biomarker of human cancer.
Previous studies have reported that miR-138 is
involved in tumorigenesis and progression of several tumor types.
Zhang et al reported that miR-138 overexpression inhibited
breast cancer metastasis and epithelial-mesenchymal transition
through negative regulation of vimentin (21). Sun et al noted that miR-138
targeted zinc finger E-box-binding homeobox 2 to suppress bladder
cancer invasion and metastases (22).
Long et al demonstrated that inhibition of miR-138 in
colorectal cancer cells resulted in a significant reduction of
migration and invasion capacity by directly targeting TWIST2
(25). In non-small cell lung cancer,
miR-138 decreased cell proliferation, migration in vitro and
tumor growth in vivo, and increased cisplatin sensitivity
through targeting multiple genes, including G protein-coupled
receptor kinase interacting ArfGAP 1 (GIT1), semaphorin 4C, cyclin
D3, Glucose regulated protein 124 (GRP124), enhancer of zeste
homolog 2 and pyruvate dehydrogenase kinase 1 (26,33–35). In
oral squamous cell carcinoma, miR-138 inhibited cell proliferation
via blockade of yes-associated protein 1 (28). Ma et al revealed that miR-138
inhibited gallbladder carcinoma cell proliferation by targeting
BCL2-associated athanogene 1 (29).
In pancreatic cancer, miR-138 overexpression reduced pancreatic
cancer cell growth through directly targeting forkhead box C1
(30). In hepatocellular carcinoma,
restoration of miR-138 expression suppressed cell viability and
colony formation, and decreased tumor cell growth in xenograft nude
mice by directly targeting cyclin D3 (36). Thus, re-expression of miR-138 in these
human cancer types may be a new potential therapeutic approach.
miRNAs serve crucial gene regulatory functions by
affecting the expression of multiple genes. To better understand
the roles of miR-138 in RCC carcinogenesis and progression, miR-138
target genes were searched. In the present study, SOX9 was
identified as a novel direct target gene of miR-138. First,
TargetScan analysis revealed that SOX9 was a potential target of
miR-138 based on putative target sequences at position 1082–1089 of
the SOX9 3′UTR. Second, luciferase reporter assay revealed that
luciferase activity was suppressed by miR-138 in PGL3-SOX9-3′UTR
Wt, whereas the inhibitory effect of miR-138 was abolished in
PGL3-SOX9-3′UTR Mut, indicating that SOX9 could be targeted by
miR-138. Third, RT-qPCR and western blot analysis demonstrated an
inhibitory effect of miR-138 on SOX9 expression at the mRNA and
protein levels. Finally, knockdown of SOX9 notably inhibited RCC
cell proliferation and invasion, with an effect similar to that
induced by miR-138, rendering SOX9 a functional target of miR-138
in RCC. These results demonstrated that SOX9 was a direct and
functional target of miR-138 in RCC. miR-138 repressed RCC cell
proliferation and invasion through negative regulation of SOX9
expression.
In conclusion, miR-138 was significantly
downregulated in RCC, and low expression levels of miR-138 were
associated with histological grade, tumor stage and lymph node
metastasis. In addition, miR-138 acted as a tumor suppressor in RCC
by inhibiting RCC cell proliferation and invasion. SOX9 was also
identified as a direct target gene of miR-138. The
miR-138/SOX9-based molecular network may serve a critical role in
RCC carcinogenesis and progression, and serve as a novel
therapeutic target for patients with RCC.
References
|
1
|
Gupta K, Miller JD, Li JZ, Russell MW and
Charbonneau C: Epidemiologic and socioeconomic burden of metastatic
renal cell carcinoma (mRCC): A literature review. Cancer Treat Rev.
34:193–205. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peng J, Mo R, Ma J and Fan J: let-7b and
let-7c are determinants of intrinsic chemoresistance in renal cell
carcinoma. World J Surg Oncol. 13:1752015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nerich V, Hugues M, Paillard MJ, Borowski
L, Nai T, Stein U, Nguyen Tan Hon T, Montcuquet P, Maurina T,
Mouillet G, et al: Clinical impact of targeted therapies in
patients with metastatic clear-cell renal cell carcinoma. Onco
Targets Ther. 7:365–374. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang L, Williamson SR, Wang M, Davidson
DD, Zhang S, Baldridge LA, Du X and Cheng L: Molecular subtyping of
metastatic renal cell carcinoma: Implications for targeted therapy.
Mol Cancer. 13:392014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Youssef YM, White NM, Grigull J, Krizova
A, Samy C, Mejia-Guerrero S, Evans A and Yousef GM: Accurate
molecular classification of kidney cancer subtypes using microRNA
signature. Eur Urol. 59:721–730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Motzer RJ and Russo P: Systemic therapy
for renal cell carcinoma. J Urol. 163:408–417. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kurozumi A, Kato M, Goto Y, Matsushita R,
Nishikawa R, Okato A, Fukumoto I, Ichikawa T and Seki N: Regulation
of the collagen cross-linking enzymes LOXL2 and PLOD2 by
tumor-suppressive microRNA-26a/b in renal cell carcinoma. Int J
Oncol. 48:1837–1846. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Coppin C, Kollmannsberger C, Le L,
Porzsolt F and Wilt TJ: Targeted therapy for advanced renal cell
cancer (RCC): A cochrane systematic review of published randomised
trials. BJU Int. 108:1556–1563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zeng Y: Principles of micro-RNA production
and maturation. Oncogene. 25:6156–6162. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Carrington JC and Ambros V: Role of
microRNAs in plant and animal development. Science. 301:336–338.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Engels BM and Hutvagner G: Principles and
effects of microRNA-mediated post-transcriptional gene regulation.
Oncogene. 25:6163–6169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Giannakakis A, Coukos G, Hatzigeorgiou A,
Sandaltzopoulos R and Zhang L: miRNA genetic alterations in human
cancers. Expert Opin Biol Ther. 7:1375–1386. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goto Y, Kurozumi A, Enokida H, Ichikawa T
and Seki N: Functional significance of aberrantly expressed
microRNAs in prostate cancer. Int J Urol. 22:242–252. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dias F, Teixeira AL, Santos JI, Gomes M,
Nogueira A, Assis J and Medeiros R: Renal cell carcinoma
development and miRNAs: A possible link to the EGFR pathway.
Pharmacogenomics. 14:1793–1803. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Z, Yu X, Shen J and Jiang Y: MicroRNA
dysregulation in uveal melanoma: A new player enters the game.
Oncotarget. 6:4562–4568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lv C, Bai Z, Liu Z, Luo P and Zhang J:
MicroRNA-495 suppresses human renal cell carcinoma malignancy by
targeting SATB1. Am J Transl Res. 7:1992–1999. 2015.PubMed/NCBI
|
|
21
|
Zhang J, Liu D, Feng Z, Mao J, Zhang C, Lu
Y, Li J, Zhang Q, Li Q and Li L: MicroRNA-138 modulates metastasis
and EMT in breast cancer cells by targeting vimentin. Biomed
Pharmacother. 77:135–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun DK, Wang JM, Zhang P and Wang YQ:
MicroRNA-138 regulates metastatic potential of bladder cancer
through ZEB2. Cell Physiol Biochem. 37:2366–2374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gao S, Wang J, Xie J, Zhang T and Dong P:
Role of miR-138 in the regulation of larynx carcinoma cell
metastases. Tumour Biol. Oct 24–2015.(Epub ahead of print).
|
|
24
|
Cristobal I, Torrejon B, González-Alonso
P, Manso R, Rojo F and García-Foncillas J: Downregulation of
miR-138 as a contributing mechanism to Lcn-2 overexpression in
colorectal cancer with liver metastasis. World J Surg.
40:1021–1022. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Long L, Huang G, Zhu H, Guo Y, Liu Y and
Huo J: Down-regulation of miR-138 promotes colorectal cancer
metastasis via directly targeting TWIST2. J Transl Med. 11:2752013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang H, Zhang H, Zhao M, Lv Z, Zhang X,
Qin X, Wang H, Wang S, Su J, Lv X, et al: MiR-138 inhibits tumor
growth through repression of EZH2 in non-small cell lung cancer.
Cell Physiol Biochem. 31:56–65. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han L, Zhang G, Zhang N, Li H, Liu Y, Fu A
and Zheng Y: Prognostic potential of microRNA-138 and its target
mRNA PDK1 in sera for patients with non-small cell lung cancer. Med
Oncol. 31:1292014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu R, Zeng G, Gao J, Ren Y, Zhang Z, Zhang
Q, Zhao J, Tao H and Li D: miR-138 suppresses the proliferation of
oral squamous cell carcinoma cells by targeting Yes-associated
protein 1. Oncol Rep. 34:2171–2178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ma F, Zhang M, Gong W, Weng M and Quan Z:
MiR-138 suppresses cell proliferation by targeting bag-1 in
gallbladder carcinoma. PLoS One. 10:e01264992015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu C, Wang M, Li Z, Xiao J, Peng F, Guo X,
Deng Y, Jiang J and Sun C: MicroRNA-138-5p regulates pancreatic
cancer cell growth through targeting FOXC1. Cell Oncol (Dordr).
38:173–181. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang B, Li H, Huang L, Luo C and Zhang Y:
Clinical significance of microRNA 138 and cyclin D3 in
hepatocellular carcinoma. J Surg Res. 193:718–723. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qiu S, Huang D, Yin D, Li F, Li X, Kung HF
and Peng Y: Suppression of tumorigenicity by microRNA-138 through
inhibition of EZH2-CDK4/6-pRb-E2F1 signal loop in glioblastoma
multiforme. Biochim Biophys Acta. 1832:1697–1707. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ye XW, Yu H, Jin YK, Jing XT, Xu M, Wan ZF
and Zhang XY: miR-138 inhibits proliferation by targeting
3-phosphoinositide-dependent protein kinase-1 in non-small cell
lung cancer cells. Clin Respir J. 9:27–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han LP, Fu T, Lin Y, Miao JL and Jiang QF:
MicroRNA-138 negatively regulates non-small cell lung cancer cells
through the interaction with cyclin D3. Tumour Biol. 37:291–298.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li J, Wang Q, Wen R, Liang J, Zhong X,
Yang W, Su D and Tang J: miR-138 inhibits cell proliferation and
reverses epithelial-mesenchymal transition in non-small cell lung
cancer cells by targeting GIT1 and SEMA4C. J Cell Mol Med.
19:2793–2805. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang W, Zhao LJ, Tan YX, Ren H and Qi ZT:
MiR-138 induces cell cycle arrest by targeting cyclin D3 in
hepatocellular carcinoma. Carcinogenesis. 33:1113–1120. 2012.
View Article : Google Scholar : PubMed/NCBI
|