Introduction
Pituitary adenomas are generally benign, arise from
adenohypophyseal cells and are one of the most frequent types of
intracranial tumor (1). It is
reported that pituitary gonadotroph adenomas account for 15–22% of
all pituitary adenomas and include the majority of clinically
nonfunctional pituitary adenomas (2,3). Surgery
remains the first line of treatment for the majority of patients,
but it cannot effectively control invasive pituitary adenomas
(4). Therefore, novel and effective
therapeutic approaches are required. Understanding the molecular
mechanisms of pituitary gonadotroph adenoma development may provide
new insights for their treatment.
Heaney et al (5) proposed peroxisome proliferator-activated
receptor-γ ligands as candidates for the management of
nonfunctioning pituitary tumors. Chesnokova et al (6) indicated that Forkhead box gene
transcription factor L2 (FOXL2) activated the clusterin promoter in
gonadotroph pituitary cells. Chesnokova et al (7) reported that clusterin and FOXL2
regulated the growth of pituitary gonadotroph adenoma. Lee et
al (8) suggested that
somatostatin receptor 3 may be a possible target for the treatment
of pituitary gonadotroph adenomas. Furthermore, the downregulation
of miRNA targeting high mobility group AT-hook 1 (HMGA) 1 and 2 and
E2F transcription factor 1 may contribute to pituitary
tumorigenesis (9). The downregulation
of miR-23b and miR-130b expression may contribute to pituitary
tumorigenesis (10). In addition,
targeting phosphoinositide 3-kinase/mechanistic target of rapamycin
signaling may activate antitumor effects against nonfunctioning
pituitary adenomas (11). Despite
results demonstrated in these previous studies, knowledge about the
underlying molecular mechanisms of pituitary gonadotroph adenoma
development may be insufficient and further research is
required.
Lee et al (12), the contributors of the GSE23207
microarray dataset, demonstrated that the multiple endocrine
neoplasia (MENX) rat model could be used as an experimental tool to
study the pathological mechanisms for human pituitary
tumorigenesis. Zhang et al (13) used the GSE23207 dataset to identify
that differentially expressed genes (DEGs) associated with ‘cell
cycle’, ‘cell division’, ‘neuroactive ligand-receptor interaction’,
‘pituitary gland’, ‘adenohypophysis’ and ‘endocrine system’ may
serve important roles in the pathogenesis of pituitary adenomas via
DEG screening, gene ontology (GO) and pathway enrichment analysis,
and protein-protein interaction (PPI) network construction. The
present study used the microarray GSE23207 dataset in addition to
the above techniques; however, miRNA-DEG regulatory network
analysis was also performed. Important genes, miRNAs and pathways
associated with the development of pituitary gonadotroph adenomas
were identified, in order to aid the clarification of the molecular
mechanisms of pituitary adenomas.
Materials and methods
Microarray data
The raw microarray data from GSE23207, as deposited
by Lee et al (12), and
GPL6247, as produced with the Affymetrix Rat Gene 1.0 ST array
[transcript (gene) version; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA] were downloaded from the Gene Expression Omnibus
(GEO; http://www.ncbi.nlm.nih.gov/geo)
database. From these datasets, data from a total of 21 pituitary
samples, including 16 from MENX rats with homozygous mutations
(p27Kip1/Cdknb1) and 5 healthy rat pituitary tissue samples were
used in the present study.
Data preprocessing
The processing of the raw data, including format
conversion, the supplementation of missing values, background
correction and quartile normalization, was performed using the affy
package in R (14).
Screening of DEGs
The DEGs in the pituitary gonadotroph adenoma group
compared with the control group were screened with the limma
package (15). The P-values for DEGs
were calculated by a t-test in the limma package. Then, the
P-values were adjusted to false discovery rate (FDR) values using
the Benjamini-Hochberg procedure (16). |log2 fold change (FC) |≥1 and FDR
<0.05 were set as cut-off criteria for all DEGs. The heatmap for
DEGs was drawn with the pheatmap package in R (17).
Functional enrichment analysis
tools
Gene ontology (GO) annotation (18), including the categories of molecular
function (MF), biological process (BP) and cellular component (CC),
and Kyoto Encyclopedia of Genes and Genomes (KEGG) (19) pathway enrichment analyses were
performed to identify upregulated and downregulated DEGs. The
functional enrichment analysis was performed using the Multifaceted
Analysis Tool for Human Transcriptome (MATHT; www.biocloudservice.com). The Gene Set
Function-Functional Enrichment-mRNA Enrichment module, based on
Fisher's test, was used for the enrichment analysis of gene sets
with a cut off of P<0.05.
PPI network
The Search Tool for the Retrieval of Interacting
Genes database was used to predict and analyze the interactions of
proteins encoded by DEGs (20). PPI
networks were constructed using Cytoscape software (version 3.2.0)
(21). The input gene sets were the
identified DEGs, and the species was set to Rattus
norvegicus. DEGs were constructed into a PPI network with a
weight threshold of ≤0.4 (medium confidence). The top 20 genes were
added one-by-one according to degree value from high to low,
starting with Cdk1, in the cluster analysis until the top 20 genes
clustered correctly. Clustering analysis was performed using
molecular context detection Cytoscape plug-in (MCODE; version
1.4.1; http://baderlab.org/Software/MCODE).
miRNA-target genes regulatory network
analysis
The miRNA that potentially regulated the DEGs were
predicted using WebGestal (22,23), and a
miRNA-target regulatory network was constructed using Cytoscape.
Experimentally verified miRNA-DEG interactions were downloaded from
Mirwalk2 (24).
Verification
The GSE26966 dataset, including 10 gonadotroph
tumors and 9 normal human pituitaries from autopsy samples, were
downloaded from the GEO database. Data normalization and DEG
analysis was performed with the affy and limma packages,
respectively. |log FC |>1 and P<0.05 were used as cut-off
criteria. GO-BP and KEGG pathway enrichment analyses were performed
using the Database for Annotation, Visualization and Integrated
Discovery (25). FDR<0.05 was set
as a cut-off value. In addition, the DEGs in GSE26966 were compared
with the DEGs in GSE23207.
Results
DEGs analysis
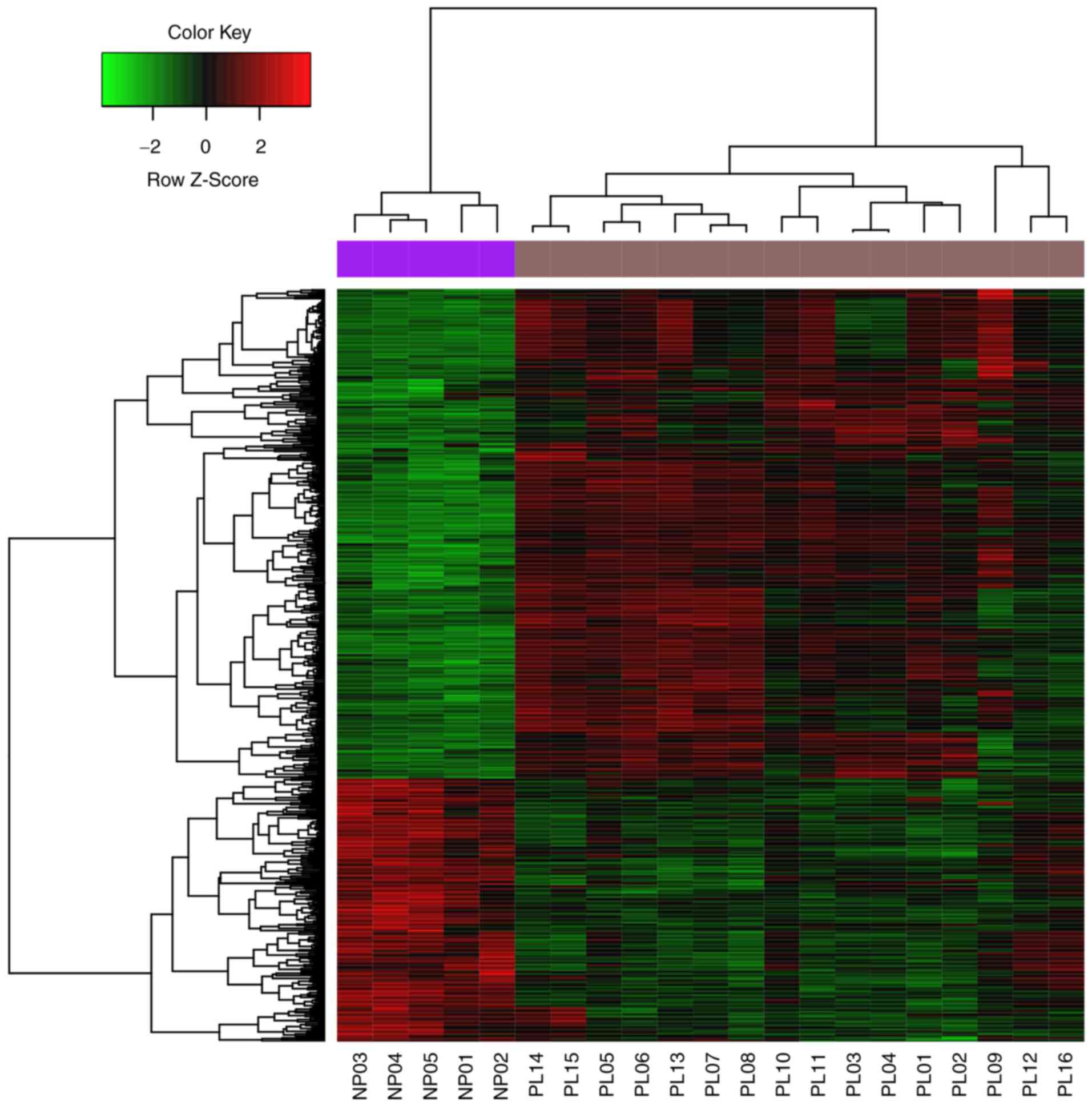
In total, 557 DEGs, including 187 significantly
upregulated and 370 significantly downregulated DEGs, were
identified in the pituitary gonadotroph adenoma group compared with
the control group (Fig. 1).
Functional enrichment analysis
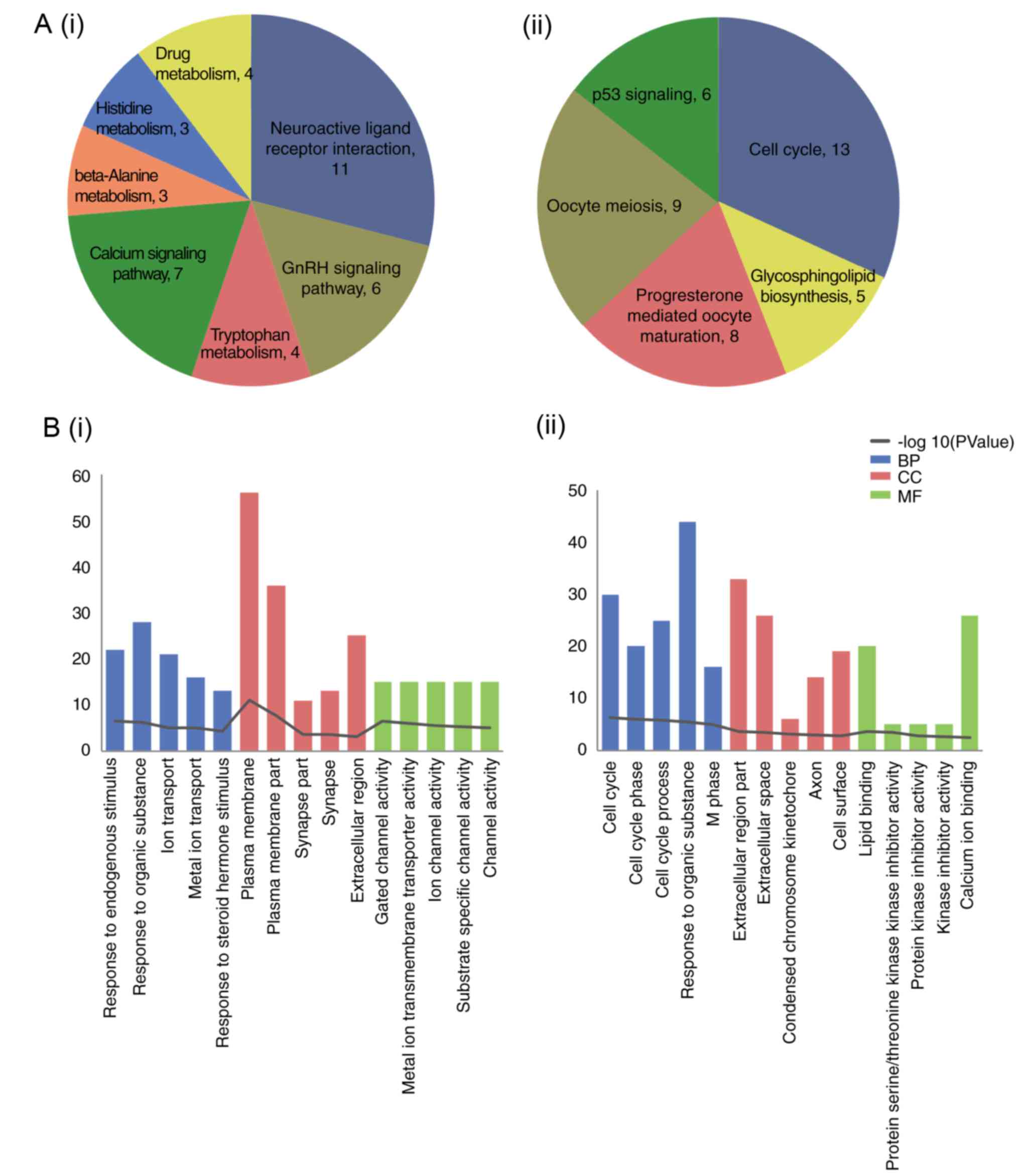
The results of functional enrichment analysis for
the up- and downregulated DEGs are included in Fig. 2. The upregulated DEGs were
predominately enriched in ‘neuroactive ligand-receptor interaction’
and the ‘GnRH signaling pathway’; downregulated DEGs were enriched
in ‘cell cycle’.
PPI network
There were 413 nodes and 1,601 protein pairs in the
PPI network (Fig. 3). The top 20
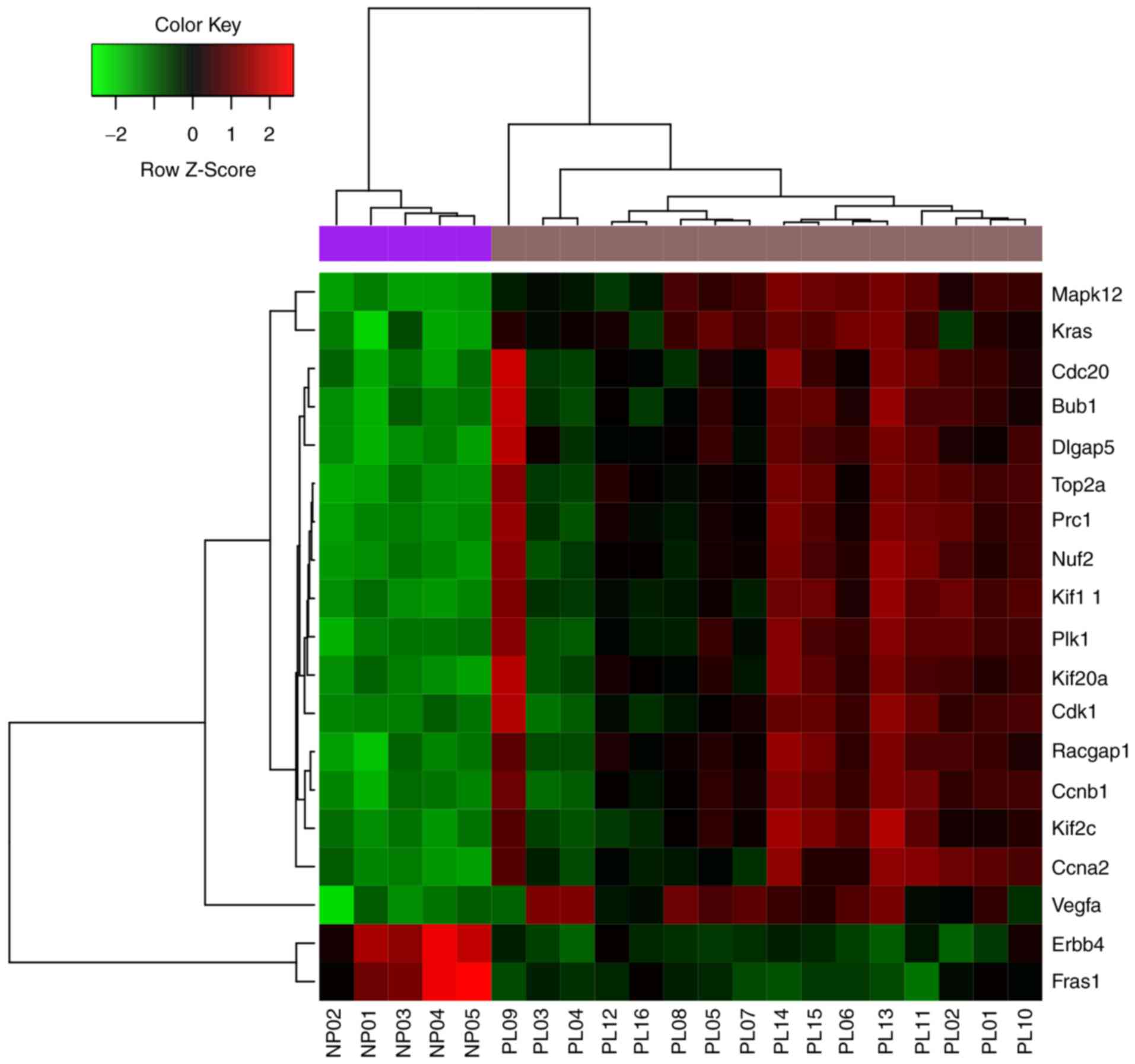
nodes with the highest degrees are included in Table I. The top 5 nodes were Cdk1 (degree,
52), topoisomerase (DNA) II α (Top2α; degree, 51), Ccna2; (degree,
50), Fraser extracellular matrix complex subunit 1 (Fras1; degree,
46) and Ccnb1 (degree, 46). The expression of the top 20 nodes from
the PPI network is illustrated in Fig.
4.
 | Table I.Top 20 nodes with the highest degrees
in the protein-protein interaction network. |
Table I.
Top 20 nodes with the highest degrees
in the protein-protein interaction network.
| Node | Degree |
|---|
| Cdk1 | 52 |
| Top2a | 51 |
| Ccna2 | 50 |
| Fras1 | 46 |
| Ccnb1 | 46 |
| Plk1 | 45 |
| Mapk12 | 44 |
| Cdc20 | 44 |
| Kif11 | 41 |
| Racgap1 | 37 |
| Bub1 | 37 |
| Prc1 | 36 |
| Bub1b | 36 |
| Kras | 35 |
| Kif20a | 35 |
| Kif2c | 34 |
| Vegfa | 34 |
| Dlgap5 | 33 |
| Erbb4 | 32 |
| Nuf2 | 32 |
Network cluster module
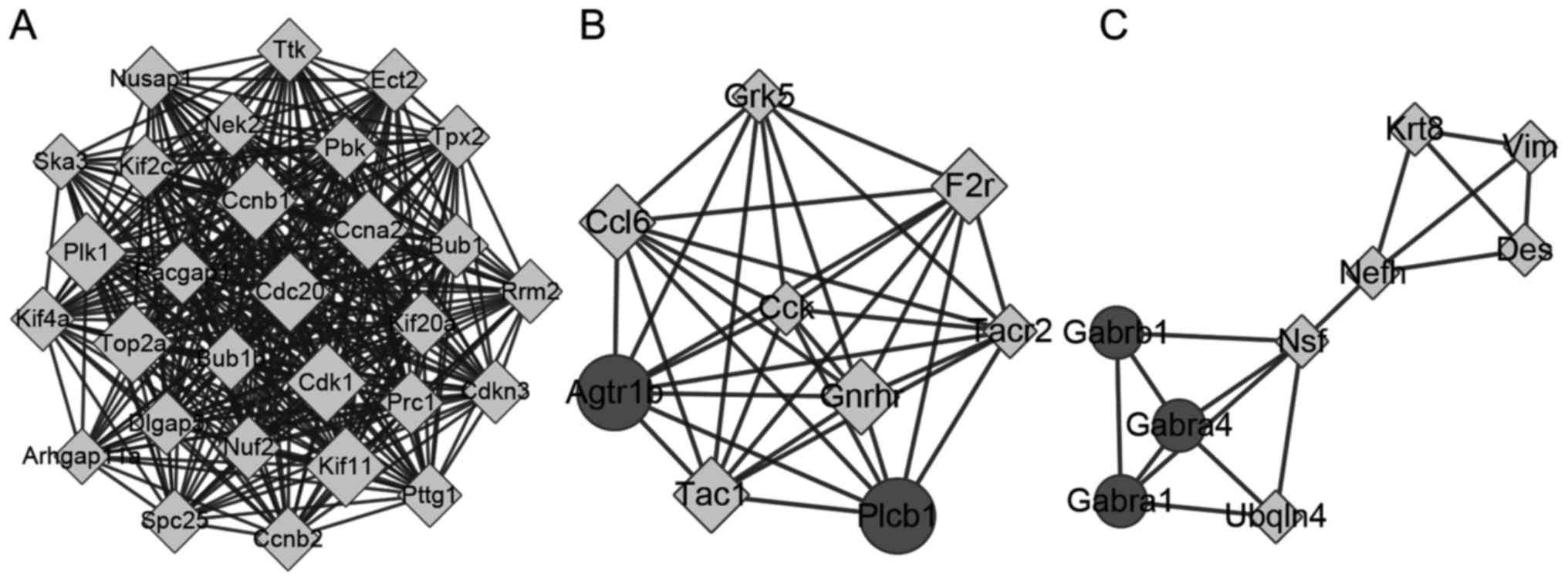
A total of 3 cluster modules were obtained (A-C;
Fig. 5). Module a contained 29 nodes
and 365 edges, module b contained 9 nodes and 35 edges and module c
contained 9 nodes and 16 edges. All genes in module a were
upregulated, and all of the top 20 genes were included in this
module. The KEGG pathways significantly enriched by these modules
are included in Table II. The DEGs
in module a were enriched in the ‘cell cycle’ and ‘oocyte meiosis’
pathways. The DEGs in module b were enriched in the ‘chemokine
signaling pathway’, ‘calcium signaling pathway’, and ‘neuroactive
ligand-receptor interaction’. DEGs in module c were enriched in
‘neuroactive ligand-receptor interaction’.
| Table II.The Kyoto Encyclopedia of Genes and
Genomes pathways significantly enriched in the three identified
modules. |
Table II.
The Kyoto Encyclopedia of Genes and
Genomes pathways significantly enriched in the three identified
modules.
| A, enriched terms
associated with module a |
|---|
|
|---|
| Pathway ID | Pathway name | Count | P-value | Genes |
|---|
| rno04110 | Cell cycle | 9 |
4.71×10−13 | CCNB1, CCNB2, PLK1,
BUB1, BUB1B, TTK, CDC20, PTTG1, CCNA2 |
| rno04114 | Oocyte meiosis | 6 |
3.34×10−7 | CCNB1, CCNB2, PLK1,
BUB1, CDC20, PTTG1 |
| rno04914 |
Progesterone-mediated oocyte
maturation | 5 |
6.50×10−6 | CCNB1, CCNB2, PLK1,
BUB1, CCNA2 |
| rno04115 | p53 signaling
pathway | 3 |
4.69×10−3 | CCNB1, CCNB2,
RRM2 |
|
| B, enriched
terms associated with module b |
|
| Pathway
ID | Pathway
name | Count | P-value | Genes |
|
| rno04062 | Chemokine signaling
pathway | 4 |
5.26×10−4 | AGTR1B, GRK5,
PLCB1, CCL6 |
| rno04020 | Calcium signaling
pathway | 4 |
6.32×10−4 | AGTR1B, TACR2,
PLCB1, F2R |
| rno04080 | Neuroactive
ligand-receptor interaction | 4 |
1.83×10−3 | AGTR1B, TACR2,
GNRHR, F2R |
|
| C, enriched
terms associated with module c |
|
| Pathway
ID | Pathway
name | Count | P-value | Genes |
|
| Brno04080 | Neuroactive
ligand-receptor interaction | 3 |
1.23×10−2 | GABRA1, GABRA4,
GABRB1 |
miRNA-target regulatory network
analysis
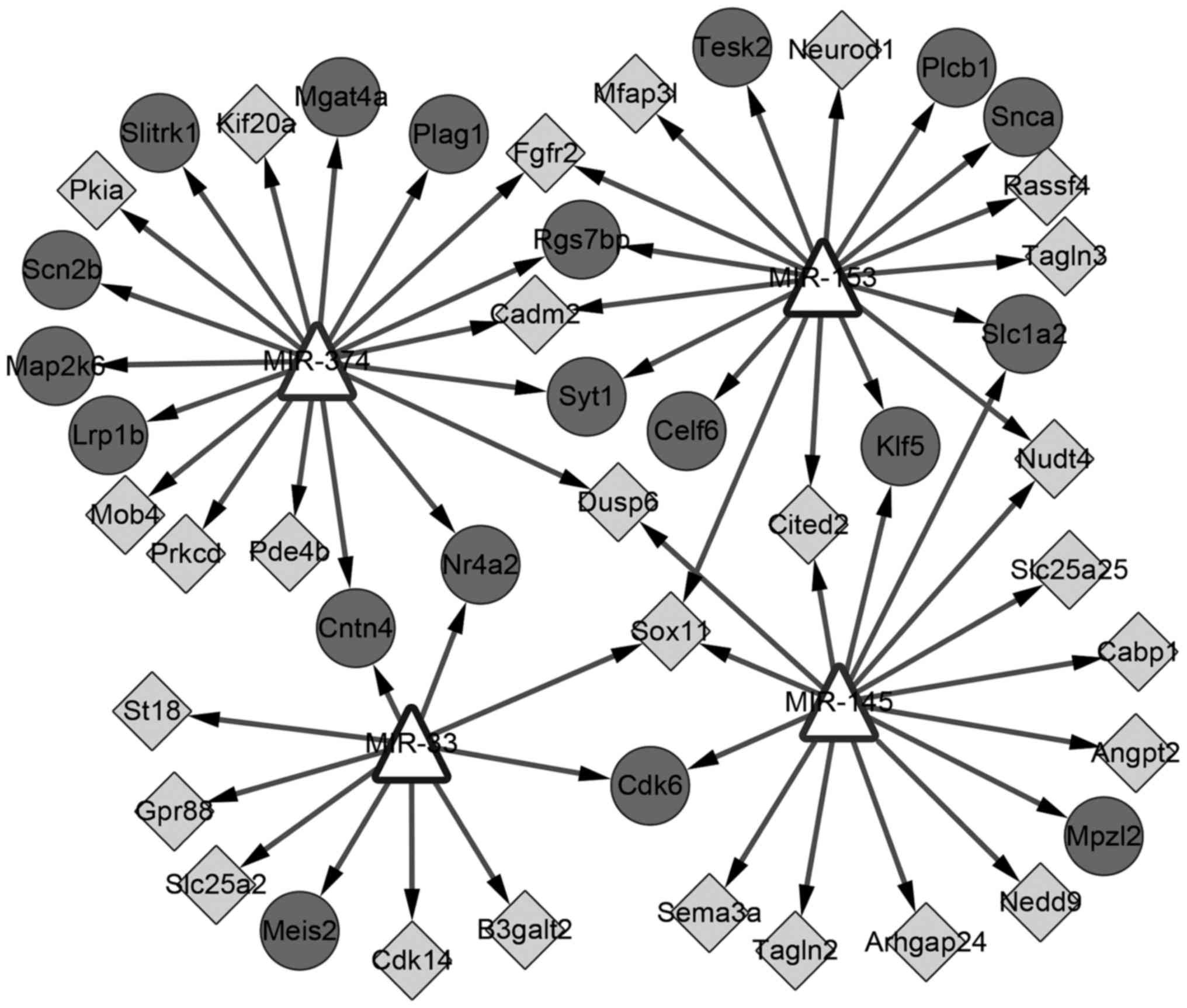
In total, 4 miRNAs were identified (miR-374, −153,
−145 and −33); miR-374 and −145 had been experimentally verified to
be able to regulate DEGs (26,27). A
total of 50 nodes and 60 edges were included in this regulatory
network (Fig. 6). The KEGG pathway
enriched by target genes in this network was ‘GnRH signaling’.
Verification
A total of 1,432 DEGs (including 651 upregulated,
781 downregulated) were identified in the pituitary gonadotroph
adenoma group compared with the control group. In total, 13 common
upregulated and 38 common downregulated DEGs were obtained from
comparing the GSE26966 and GSE23207 data. The significantly
enriched GO-BP and KEGG pathways for GSE26966 were also identified;
the top 10 upregulated GO-BP terms were ‘ion transmembrane
transport’, ‘positive regulation of synapse assembly’, ‘adherens
junction organization’, ‘central nervous system development’,
‘membrane depolarization during cardiac muscle cell action
potential’, ‘chloride transmembrane transport’, ‘heterophilic
cell-cell adhesion via plasma membrane cell adhesion molecules’,
‘sodium ion transport’, ‘potassium ion transport’ and ‘negative
regulation of the mitotic cell cycle’.
Discussion
A total of 557 DEGs, including 187 significantly
upregulated and 370 downregulated DEGs, were identified in the
pituitary gonadotroph adenoma group compared with the control
group. Cdk1 (degree, 52) had the highest degree in the PPI network.
Ccna2 (degree, 50) and Ccnb1 (degree, 46) were also enriched in the
‘cell cycle’. Furthermore, the upregulated DEGs were predominately
enriched in ‘neuroactive ligand-receptor interaction’ and
downregulated DEGs were mainly enriched in the ‘cell cycle’. DEGs
in module were also mainly enriched in ‘cell cycle’. DEGs in module
b and c were enriched in ‘neuroactive ligand-receptor interaction’.
Furthermore, miR-374, −153, −145 and −33 were identified as miRNAs
that may have regulated the DEGs.
Cdk1, Ccna2 and Ccnb1 demonstrated the highest
connectivity degrees in the PPI network. A number of previous
studies have suggested that Ccnb1 and Ccna2 are abnormally
expressed in pituitary adenomas (10,28).
Another study suggested that the downregulation of miR-410, which
targets Ccnb1, contributes to pituitary gonadotroph tumor
development (4). The overexpression
of Ccna2 serves a key function in pituitary tumorigenesis (10). p27 belongs to the Cdk1 family, and is
downregulated in invasive and recurrent adenomas as well as
pituitary carcinomas (29). These
studies suggest that Cdk1, Ccna2 and Ccnb1 may have important roles
in the development of pituitary gonadotroph adenomas, which
supports the results of the present study.
Furthermore, in this study, the upregulated DEGs
were predominately enriched in ‘neuroactive ligand-receptor
interaction’ and downregulated DEGs were enriched in ‘cell cycle’.
DEGs in module a were also mainly enriched in ‘cell cycle’. DEGs in
modules b and c were enriched in ‘neuroactive ligand-receptor
interactions’. Previous research has demonstrated cell cycle
dysregulation in pituitary adenomas (30–32).
Quereda et al (31) indicated
that the dysregulation of the cell cycle in pituitary disease had a
significant effect on treatment. Cell cycle dysregulation is the
main mechanism by which HMGA proteins cause the development of
pituitary adenomas (33). The damage
to cell cycle regulation in pituitary adenomas caused by Wee1
downregulation may cause the G2/M checkpoint to be lost, leading to
the accumulation of DNA damage and the induction of tumor
development (34). Thus, the
dysregulation of the cell cycle may be associated with the
progression of pituitary gonadotroph adenomas. In addition, Zhang
et al (13) indicated that
genes associated with neuroactive ligand-receptor interaction,
including the γ-aminobutyric acid type a receptor α1, α4 and β1
subunits, may serve essential roles in the pathogenesis of
pituitary adenomas. Although the role of neuroactive
ligand-receptor interaction in pituitary adenoma development have
not been fully characterized, the present study identified that the
upregulated DEGs were predominately enriched in ‘neuroactive
ligand-receptor interaction’, and that DEGs in module b and c were
particularly enriched in ‘neuroactive ligand-receptor interaction’.
Thus, it may be that a neuroactive ligand-receptor interaction is
critical in the pathogenesis of pituitary gonadotroph adenomas.
Ccna2 and Ccnb1 were also enriched in ‘cell cycle’
in the present study. Therefore, Ccna2 and Ccnb1 may be associated
with the development of pituitary gonadotroph adenomas via an
effect on the cell cycle.
The results also identified that miR-374, −153, −145
and −33 may have been important miRNAs for regulating DEGs. miRNAs
function as antisense regulators of gene expression, and contribute
to the progression and development of cancer (35). One study demonstrated that miR-374 was
upregulated in pituitary gonadotroph adenomas compared with the
normal pituitary by reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) analysis (4). Palumbo et al (36) further confirmed that miR-145 was
downregulated in pituitary tumors compared with normal tissues by
performing RT-qPCR. Sachdeva and Mo (37) suggested that miR-145 could be regarded
as a potential cancer biomarker and a novel target for the therapy
of cancer. Furthermore, miR-374 and −145 have been verified to be
able to regulate DEGs by previous experiments (26,27). Thus,
the present study corroborates previous research by demonstrating
that miR-374 and −145 may contribute to the development of
pituitary gonadotroph adenomas via regulating the expression of
target genes. To the best of our knowledge, no previous
experimental studies concerning the association between miR-153 and
−33 and pituitary adenomas have been performed; therefore, this
topic should be further researched.
The upregulated and downregulated DEGs identified
using the GSE26966 human dataset were not significantly enriched in
the ‘neuroactive ligand-receptor interaction’ pathways or ‘cell
cycle’ pathways. However, as the neuroactive ligand-receptor
interaction pathway is a collection of receptors and ligands
associated with intracellular and extracellular signaling pathways
on the plasma membrane (38), a total
of 5 of the top 10 upregulated GO-BP terms (including ‘ion
transmembrane transport’, ‘positive regulation of synapse
assembly’, ‘chloride transmembrane transport’, ‘sodium ion
transport’ and ‘potassium ion transport’) were therefore associated
with the neuroactive ligand-receptor interaction pathway.
The downregulated DEGs were predominately enriched
in the GO-BP terms ‘negative regulation of cell proliferation’,
‘regulation of insulin-like growth factor receptor signaling
pathway’, ‘aging’ and ‘regulation of cell growth’, among which
‘negative regulation of cell proliferation’, ‘aging’ and
‘regulation of cell growth’ were associated with ‘cell cycle’.
Other enriched BP terms identified in this study include ‘cell
differentiation’, ‘positive regulation of osteoblast
differentiation’, ‘fat cell differentiation’, ‘positive regulation
of cell proliferation’ and ‘cell cycle arrest’ were also associated
with ‘cell cycle’. Therefore, the association of neuroactive
ligand-receptor interactions and the cell cycle pathway were
indirectly verified for the human array data of GSE26966,
suggesting that the mechanisms associated with neuroactive
ligand-receptor interactions and the cell cycle pathway may also
serve important roles in the development of human pituitary
gonadotroph adenomas.
Although the array data, GSE23207, which was used to
perform the present analysis, had been analyzed by a previously
published study (12), there were
several differences between the present study and that of Zhang
et al (13). First, as
described in the introduction section, in addition to the screening
of DEGs, PPI network analysis and GO and KEGG pathway enrichment
analysis, miRNA-target gene regulatory network analysis was also
performed. Secondly, verification was achieved using the human
array data from GSE26966. Therefore, the present study provides new
information to advance the understanding of the development of
pituitary gonadotroph adenomas.
In conclusion, Cdk1, Ccna2, Ccnb1, ‘cell cycle’ and
‘neuroactive ligand-receptor interaction’ pathways may serve
important roles in the development of pituitary gonadotroph
adenomas, and Ccna2 and Ccnb1 may be involved in the development of
pituitary gonadotroph adenomas via the cell cycle pathway.
Furthermore, miR-374 and −145 may contribute to the development of
pituitary gonadotroph adenomas via regulating the expression of
target genes. These genes, pathways and miRNAs are potential
targets of pituitary gonadotroph adenoma therapy. However, there
were several limitations in this study, including the relatively
small sample size and the lack of miRNA array data for human
pituitary gonadotroph adenomas for the verification of the up- and
downregulated miRNAs. Further studies concerning the association
between miR-153 and −33, and pituitary gonadotroph
adenomas/pituitary adenomas, are required.
References
|
1
|
Trouillas J, Roy P, Sturm N, Dantony E,
Cortet-Rudelli C, Viennet G, Bonneville JF, Assaker R, Auger C,
Brue T, et al: A new prognostic clinicopathological classification
of pituitary adenomas: A multicentric case-control study of 410
patients with 8 years post-operative follow-up. Acta Neuropatholo.
126:123–135. 2013. View Article : Google Scholar
|
|
2
|
Daly AF, Rixhon M, Adam C, Dempegioti A,
Tichomirowa MA and Beckers A: High prevalence of pituitary
adenomas: A cross-sectional study in the province of liege,
belgium. J Clin Endocrinol Metab. 91:4769–4775. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fernandez A, Karavitaki N and Wass JA:
Prevalence of pituitary adenomas: A community-based,
cross-sectional study in banbury (Oxfordshire, UK). Clin Endocrinol
(Oxf). 72:377–382. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Müssnich P, Raverot G, Jaffrain-Rea ML,
Fraggetta F, Wierinckx A, Trouillas J, Fusco A and D'Angelo D:
Downregulation of miR-410 targeting the cyclin B1 gene plays a role
in pituitary gonadotroph tumors. Cell Cycle. 14:2590–2597. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heaney AP, Fernando M and Melmed S:
PPAR-gamma receptor ligands: Novel therapy for pituitary adenomas.
J Clin Invest. 111:1381–1388. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chesnokova V, Zonis S, Zhou C, Ben-Shlomo
A, Wawrowsky K, Toledano Y, Tong Y, Kovacs K, Scheithauer B and
Melmed S: Lineage-specific restraint of pituitary gonadotroph cell
adenoma growth. PLoS One. 6:e179242011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chesnokova V, Zonis S, Wawrowsky K, Tani
Y, Ben-Shlomo A, Ljubimov V, Mamelak A, Bannykh S and Melmed S:
Clusterin and foxl2 act concordantly to regulate pituitary
gonadotroph adenoma growth. Mol Endocrinol. 26:2092–2103. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee M, Lupp A, Mendoza N, Martin N,
Beschorner R, Honegger J, Schlegel J, Shively T, Pulz E, Schulz S,
et al: SSTR3 is a putative target for the medical treatment of
gonadotroph adenomas of the pituitary. Endocr Relat Cancer.
22:111–119. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
D'Angelo D, Palmieri D, Mussnich P, Roche
M, Wierinckx A, Raverot G, Fedele M, Croce CM, Trouillas J and
Fusco A: Altered microRNA expression profile in human pituitary GH
adenomas: Down-regulation of miRNA targeting HMGA1, HMGA2, and
E2F1. J Clin Endocrinol Metab. 97:E1128–E1138. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Leone V, Langella C, D'Angelo D, Mussnich
P, Wierinckx A, Terracciano L, Raverot G, Lachuer J, Rotondi S,
Jaffrain-Rea ML, et al: Mir-23b and miR-130b expression is
downregulated in pituitary adenomas. Mol Cell Endocrinol. 390:1–7.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee M, Wiedemann T, Gross C, Leinhäuser I,
Roncaroli F, Braren R and Pellegata NS: Targeting PI3K/mTOR
signaling displays potent antitumor efficacy against nonfunctioning
pituitary adenomas. Clin Cancer Res. 21:3204–3215. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee M, Marinoni I, Irmler M, Psaras T,
Honegger JB, Beschorner R, Anastasov N, Beckers J, Theodoropoulou
M, Roncaroli F and Pellegata NS: Transcriptome analysis of
MENX-associated rat pituitary adenomas identifies novel molecular
mechanisms involved in the pathogenesis of human pituitary
gonadotroph adenomas. Acta Neuropathol. 126:137–150. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang H, Xu C, Sun N, Zhou Y, Yu X, Yan X
and Zhang Q: Gene expression profiling analysis of MENX-associated
rat pituitary adenomas contributes to understand molecular
mechanisms of human pituitary adenomas. Oncol Lett. 11:125–133.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: Affy-analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smyth GK: LIMMA: Linear models for
microarray data. Bioinformatics and Computational Biology Solutions
Using R and Bioconductor. Statistics for Biology and Health.
397–420. 2005.
|
|
16
|
Ferreira JA: The Benjamini-Hochberg method
in the case of discrete test statistics. Int J Biostat. 3:112007.
View Article : Google Scholar
|
|
17
|
Wang L, Cao C, Ma Q, Zeng Q, Wang H, Cheng
Z, Zhu G, Qi J, Ma H, Nian H and Wang Y: RNA-seq analyses of
multiple meristems of soybean: Novel and alternative transcripts,
evolutionary and functional implications. BMC Plant Biol.
14:1692014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Szklarczyk D, Franceschini A, Kuhn M,
Simonovic M, Roth A, Minguez P, Doerks T, Stark M, Muller J, Bork
P, et al: The STRING database in 2011: Functional interaction
networks of proteins, globally integrated and scored. Nucleic Acids
Res. 39(Database Issue): D561–D568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang J, Duncan D, Shi Z and Zhang B:
WEB-based gene set analysis toolkit (WebGestalt): Update 2013.
Nucleic Acids Res. 41(Web Server Issue): W77–W83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang B, Kirov S and Snoddy J: WebGestalt:
An integrated system for exploring gene sets in various biological
contexts. Nucleic Acids Res. 33(Web Server Issue): W741–W748. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dweep H and Gretz N: miRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang DW, Sherman BT, Tan Q, Kir J, Liu D,
Bryant D, Guo Y, Stephens R, Baseler MW, Lane HC and Lempicki RA:
DAVID bioinformatics resources: Expanded annotation database and
novel algorithms to better extract biology from large gene lists.
Nucleic Acids Res. 35(Web Server Issue): W169–W175. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jeyaseelan K, Lim KY and Armugam A:
MicroRNA expression in the blood and brain of rats subjected to
transient focal ischemia by middle cerebral artery occlusion.
Stroke. 39:959–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Turczyńska KM, Hellstrand P, Swärd K and
Albinsson S: Regulation of vascular smooth muscle
mechanotransduction by microRNAs and L-type calcium channels.
Commun Integr Biol. 6:e222782013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Raverot G, Wierinckx A, Dantony E, Auger
C, Chapas G, Villeneuve L, Brue T, Figarella-Branger D, Roy P,
Jouanneau E, et al: Prognostic factors in prolactin pituitary
tumors: clinical, histological, and molecular data from a series of
94 patients with a long postoperative follow-up. J Clin Endocrinol
Metab. 95:1708–1716. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kontogeorgos G: Predictive markers of
pituitary adenoma behavior. Neuroendocrinology. 83:179–188. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bahar A, Bicknell JE, Simpson DJ, Clayton
RN and Farrell WE: Loss of expression of the growth inhibitory gene
GADD45 gamma, in human pituitary adenomas, is associated with CpG
island methylation. Oncogene. 23:936–944. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Quereda V and Malumbres M: Cell cycle
control of pituitary development and disease. J Mol Endocrinol.
42:75–86. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chesnokova V and Melmed S: Pituitary
senescence: The evolving role of Pttg. Mol Cell Endocrinol.
326:55–59. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fedele M, Palmieri D and Fusco A: HMGA2: A
pituitary tumour subtype-specific oncogene? Mol Cell Endocrinol.
326:19–24. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Butz H, Likó I, Czirják S, Igaz P, Khan
MM, Zivkovic V, Bálint K, Korbonits M, Rácz K and Patócs A:
Down-regulation of Wee1 kinase by a specific subset of microRNA in
human sporadic pituitary adenomas. J Clin Endocrinol Metab.
95:E181–E191. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Amaral FC, Torres N, Saggioro F, Neder L,
Machado HR, Silva WA Jr, Moreira AC and Castro M: MicroRNAs
differentially expressed in ACTH-secreting pituitary tumors. J Clin
Endocrinol Metab. 94:320–323. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Palumbo T, Faucz FR, Azevedo M, Xekouki P,
Iliopoulos D and Stratakis CA: Functional screen analysis reveals
miR-26b and miR-128 as central regulators of pituitary
somatomammotrophic tumor growth through activation of the PTEN-AKT
pathway. Oncogene. 32:1651–1659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sachdeva M and Mo YY: miR-145-mediated
suppression of cell growth, invasion and metastasis. Am J Transl
Res. 2:170–180. 2010.PubMed/NCBI
|
|
38
|
Lauss M, Kriegner A, Vierlinger K and
Noehammer C: Characterization of the drugged human genome.
Pharmacogenomics. 8:1063–1073. 2007. View Article : Google Scholar : PubMed/NCBI
|